Мшанки
| Мшанки Временной диапазон: Спорные кембрийские записи ( Pywackia , Protomelission [3] ) | |
|---|---|
 | |
| «Мшанки» из Эрнста Геккеля книги «Художественные формы природы» , 1904 год. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Клэйд : | Лофофората |
| Тип: | Мшанки Эренберг , 1831 г. [4] |
| Классы | |
| Синонимы [5] | |
Ectoprocta (Nitsche, 1869) (ранее подтип Bryozoa) | |
Мшанки (также известные как Polyzoa , Ectoprocta или обычно как моховые животные ) [6] — тип простых водных беспозвоночных животных, почти все из которых живут сидячими колониями . Обычно около длину 0,5 миллиметра ( 1/64 фильтрационного Имея дюйма), они имеют особую структуру питания, называемую лофофором , «короной» из щупалец, используемой для питания . Большинство морских мшанок обитают в тропических водах, но некоторые встречаются в океанических желобах и полярных водах. Мшанки подразделяются на морские мшанки (Stenolaemata), пресноводные мшанки (Phylactolaemata) и преимущественно морские мшанки (Gymnolaemata), некоторые представители которых предпочитают солоноватые воды . 5869 современных видов. Известно [7] Первоначально вся кроновая группа Bryozoa была колониальной, но в результате адаптации к мезопсаммальному (интерстициальному пространству в морском песке) образу жизни или к глубоководным местообитаниям с тех пор развились вторично одиночные формы. Одиночные виды описаны в четырех родах ; Aethozooides , Aethozoon , Franzenella и Monobryozoon ). Последний имеет статоцистоподобный орган с предполагаемой выделительной функцией. [8] [9]
Термины Polyzoa и Bryozoa были введены в 1830 и 1831 годах соответственно. [10] [11] Вскоре после того, как ей было присвоено название, была обнаружена еще одна группа животных, механизм фильтрации которых выглядел схожим, поэтому ее включали в состав мшанок до 1869 года, когда было отмечено, что эти две группы сильно различаются внутри. Новой группе было присвоено название « Entoprocta », а первоначальные мшанки назывались «Ectoprocta». Разногласия по поводу терминологии сохранялись и в 20 веке, но сейчас общепринятым термином является «Мшанка». [12] [13]
Колонии принимают самые разные формы, включая веера, кусты и листы. Животные-одиночки, называемые зооидами , живут по всей колонии и не являются полностью независимыми. Эти люди могут выполнять уникальные и разнообразные функции. Во всех колониях есть «аутозооиды», которые отвечают за питание, выделение и доставку питательных веществ в колонию по различным каналам. В некоторых классах есть специализированные зооиды, такие как инкубаторы для оплодотворенных яиц, колониальные защитные структуры и корневые структуры прикрепления. Cheilostomata — самый разнообразный отряд мшанок, возможно, потому, что его представители обладают самым широким спектром специализированных зооидов. Они имеют минерализованный экзоскелет и образуют однослойные листы, которые покрывают поверхность, а некоторые колонии могут ползти очень медленно, используя колючие защитные зооиды в качестве ног.
Каждый зооид состоит из «цистиды», которая обеспечивает стенку тела и образует экзоскелет , и « полипида », удерживающего органы. У зооидов нет специальных органов выделения, и полипиды автозооидов сбрасываются, когда они перегружены отходами; обычно в стенке тела затем вырастает замещающий полипид. Их кишечник имеет U-образную форму: рот находится внутри венца щупалец, а анус — снаружи. Зооиды всех пресноводных видов одновременно являются гермафродитами . Хотя представители многих морских видов функционируют сначала как самцы, а затем как самки, их колонии всегда содержат комбинацию зооидов, находящихся на мужской и женской стадиях. Все виды выделяют сперму в воду. Некоторые также выпускают яйцеклетки в воду, в то время как другие захватывают сперму своими щупальцами для внутреннего оплодотворения яйцеклеток. У некоторых видов личинки имеют крупные желтки , идут на питание и быстро оседают на поверхности. Другие производят личинок, у которых мало желтка, но они плавают и питаются в течение нескольких дней, прежде чем оседать. После расселения все личинки подвергаются радикальному метаморфоз , разрушающий и перестраивающий почти все внутренние ткани. Пресноводные виды также производят статобласты , которые бездействуют до тех пор, пока условия не станут благоприятными, что позволяет линии колонии выжить, даже если суровые условия убивают материнскую колонию.
Хищниками морских мшанок являются морские слизни (голожаберные), рыбы, морские ежи , пикногониды , ракообразные , клещи и морские звезды . На пресноводных мшанок охотятся улитки, насекомые и рыбы. В Таиланде многие популяции одного пресноводного вида были уничтожены интродуцированным видом улиток. [14] Membranipora membranacea , быстрорастущая инвазивная мшанка у северо-восточного и северо-западного побережья США, сократила леса водорослей , что это повлияло на местные популяции рыб и беспозвоночных. настолько [15] Мшанки распространили болезни на рыбные фермы и рыбаков. Химические вещества, извлеченные из морских видов мшанок, исследовались для лечения рака и болезни Альцгеймера , но результаты исследований не обнадежили. [16]
Минерализованные скелеты мшанок впервые появляются в породах раннего ордовика . [1] что делает его последним крупным типом, появившимся в летописи окаменелостей. Это заставило исследователей подозревать, что мшанки возникли раньше, но изначально были неминерализованными и, возможно, значительно отличались от окаменелых и современных форм. В 2021 году некоторые исследования показали, что Protomelission , род, известный с кембрийского периода , может быть примером ранней мшанки, [17] но более поздние исследования показали, что этот таксон может вместо этого представлять собой дазикладную водоросль. [3] Ранние окаменелости в основном имеют прямостоячие формы, но постепенно доминируют корковые формы. Неизвестно, является ли тип монофилетическим . Эволюционные отношения мшанок с другими типами также неясны, отчасти потому, что на взгляд ученых на генеалогическое древо животных в основном влияют более известные типы. Как морфологический , так и молекулярно-филогенический анализ расходятся во мнениях относительно взаимоотношений мшанок с энтопроктами, относительно того, следует ли объединять мшанки с брахиоподами и форонидами в Lophophorata и следует ли считать мшанок протостомами или вторичноротыми .
Описание
[ редактировать ]Отличительные особенности
[ редактировать ]Мшанки, форониды и брахиоподы извлекают пищу из воды с помощью лофофора — «короны» из полых щупалец. Мшанки образуют колонии, состоящие из клонов , называемых зооидами, обычно размером около 0,5 мм ( 1 ⁄ 64 дюйма) в длину. [18] Форониды напоминают зооидов мшанок, но имеют длину от 2 до 20 см (от 1 до 8 дюймов) и, хотя часто растут сгустками, не образуют колоний, состоящих из клонов. [19] Брахиоподы, которых обычно считают близкими родственниками мшанок и форонид, отличаются наличием раковин, напоминающих раковины двустворчатых моллюсков . [20] Все три из этих типов имеют целом , внутреннюю полость, выстланную мезотелием . [18] [19] [20] Некоторые инкрустирующие колонии мшанок с минерализованным экзоскелетом очень похожи на маленькие кораллы. Однако колонии мшанок основаны предком, который имеет круглую, а не форму, как у нормального зооида этого вида. С другой стороны, полип-основатель коралла имеет форму, подобную форме дочерних полипов, а зооиды кораллов не имеют целома или лофофора . [21]
Энтопрокты , другой тип фильтраторов, похожи на мшанок, но их лофофороподобная структура питания имеет твердые щупальца, их анус расположен внутри, а не снаружи основания «короны», и у них нет целома . [22]
| Мшанки [18] (Эктопрокта) | Другие лофофораты [23] | Другие лофотрохозоа | Похожий тип | |||
|---|---|---|---|---|---|---|
| Форониды [19] | Брахиоподы [20] | Аннелида , Моллюска | Энтопрокта [22] | Кораллы (класс типа Cnidaria ) [21] | ||
| целом | Трехчастный, если включена полость эпистома | Трехчастный | По одному на сегмент в базовой форме; слились в некоторые таксоны | никто | ||
| Формирование целома | Неопределенно, поскольку метаморфоз личинок во взрослых особей делает невозможным проследить. | Энтероцелия | шизоцелия | непригодный | ||
| Лофофор | С полыми щупальцами | никто | Похожая структура питания, но с твердыми щупальцами. | никто | ||
| Питающий ток | От кончиков до оснований щупалец | непригодный | От оснований до кончиков щупалец | непригодный | ||
| Мультиреснитчатые клетки эпителия | Да [24] | нет [24] | Да [24] | непригодный | ||
| Положение ануса | Внешнее основание лофофора | Варьируется, отсутствует у некоторых видов | Задняя часть, но нет у Siboglinidae. | Внутреннее основание лофофороподобного органа | никто | |
| Колониальный | В большинстве своем колонии клонов; один единственный род | Сидящие виды часто образуют скопления, но без активного сотрудничества. | Колонии клонов у некоторых видов; некоторые одиночные виды | Колонии клонов | ||
| Форма зооида-основателя | Круглый, в отличие от обычных зооидов. [21] | непригодный | То же, что и другие зооиды | |||
| Минерализованные экзоскелеты | Некоторые таксоны | нет | Двустворчатые раковины | Некоторые сидячие кольчатые черви строят минерализованные трубки; [25] у большинства моллюсков есть раковины, но у большинства современных головоногих моллюсков есть внутренние раковины или они отсутствуют. [26] | нет | Некоторые таксоны |
Виды зооидов
[ редактировать ]Все мшанки колониальные, за исключением одного рода — Monobryozoon . [27] [28] Отдельные члены колонии мшанок составляют около 0,5 мм ( 1/64 ) в дюйма длину и известны как зооиды . [18] поскольку они не являются полностью независимыми животными. [29] Все колонии содержат питающихся зооидов, известных как автозооиды. Представители некоторых групп также содержат непитающиеся гетерозооиды, также известные как полиморфные зооиды, которые выполняют множество функций, помимо питания; [28] Члены колонии генетически идентичны и взаимодействуют друг с другом, как органы более крупных животных. [18] Какой тип зооида растет и где в колонии определяется химическими сигналами, исходящими от колонии в целом, а иногда и в ответ на запах хищников или конкурирующих колоний. [28]
Тела всех типов имеют две основные части. Цистида экзоскелета состоит из стенки тела и , секретируемого эпидермисом любого типа . Экзоскелет может быть органическим ( хитин , полисахарид или белок ) или состоять из минерала карбоната кальция . Последний всегда отсутствует у пресноводных видов. [30] Стенка тела состоит из эпидермиса, базальной пластинки (мата из неклеточного материала), соединительной ткани , мышц и мезотелия , выстилающего целом (основную полость тела). [18] – за исключением того, что у одного класса мезотелий расщепляется на два отдельных слоя: внутренний, образующий перепончатый мешок, свободно плавающий и содержащий целом, и наружный, прикрепленный к стенке тела и заключающий перепончатый мешок в псевдоцелом . [31] Другая основная часть тела мшанки, известная как полипид и почти полностью расположенная внутри цистиды, содержит нервную систему, пищеварительную систему, некоторые специализированные мышцы и питательный аппарат или другие специализированные органы, заменяющие питательный аппарат. [18]
Кормление зооидов
[ редактировать ]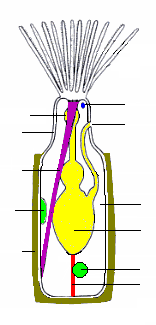
Наиболее распространенным типом зооидов является питающийся автозооид, у которого полипид несет «корону» из полых щупалец, называемую лофофором , которая захватывает частицы пищи из воды. [28] Во всех колониях большой процент зооидов являются автозооидами, а некоторые полностью состоят из автозооидов, часть из которых также занимается размножением. [32]
Основная форма «короны» — полный круг. У пресноводных мшанок ( Pylactolaemata ) крона кажется U-образной, но это впечатление создается глубокой вмятиной по краю коронки, не имеющей просвета в бахроме щупалец. [18] По бокам щупалец покрыты тонкими волосками, называемыми ресничками , движение которых направляет поток воды от кончиков щупалец к их основаниям, откуда он выходит. Частицы пищи, сталкивающиеся с щупальцами, улавливаются слизью , а дальнейшие реснички на внутренних поверхностях щупалец перемещают частицы ко рту в центре. [23] Метод, используемый эктопроктами, называется «восходящим сбором», поскольку частицы пищи захватываются до того, как они пройдут через поле ресничек, создающее питающий ток. Этот метод используют также форониды , брахиоподы и крыложаберные . [33]
Лофофор и рот закреплены на гибкой трубке, называемой «инверт», которую можно вывернуть наизнанку и втянуть в полипид. [18] скорее, как палец резиновой перчатки; в этом положении лофофор лежит внутри инверта и сложен, как спицы зонтика. Инверт вытягивается, иногда в течение 60 миллисекунд , с помощью пары мышц-ретракторов, которые закреплены на дальнем конце цистиды. Датчики на кончиках щупалец могут обнаружить признаки опасности до того, как инверт и лофофор полностью выдвинутся. Разгибание происходит за счет увеличения внутреннего давления жидкости, которое виды с гибким экзоскелетом производят за счет сокращения круговых мышц, лежащих внутри стенки тела. [18] в то время как виды с перепончатым мешком используют круговые мышцы, чтобы сжать его. [31] Некоторые виды с жестким экзоскелетом имеют гибкую мембрану, которая заменяет часть экзоскелета, а поперечные мышцы, закрепленные на дальней стороне экзоскелета, увеличивают давление жидкости, втягивая мембрану внутрь. [18] У других брешь в защитном скелете отсутствует, и поперечные мышцы натягивают гибкий мешок, который снаружи соединен с водой небольшой порой; расширение мешка увеличивает давление внутри тела и выталкивает инверт и лофофор наружу. [18] У некоторых видов втянутая инверсия и лофофор защищены крышечкой («крышкой»), которая закрывается мышцами и открывается под давлением жидкости. У одного класса полая доля, называемая «эпистом», нависает над ртом. [18]
Кишка имеет U-образную форму, идет ото рта, в центре лофофора, вниз внутрь животного, а затем обратно к анальному отверстию , которое расположено на обратной стороне, снаружи и обычно под лофофором. [18] Сеть нитей мезотелия , называемая «фуникулы» («веревочки»). [34] соединяет мезотелий, покрывающий кишечник, с мезотелием, выстилающим стенку тела. Стенка каждой нити состоит из мезотелия и окружает пространство, заполненное жидкостью, предположительно кровью. [18] Зооиды колонии связаны между собой, что позволяет автозооидам делиться пищей друг с другом и с любыми не питающимися гетерозооидами. [18] Способ связи у разных классов мшанок различен: от довольно крупных промежутков в стенках тела до мелких пор, через которые по канатикам проходят питательные вещества. [18] [31]
Вокруг глотки (горла) находится нервное кольцо, а по одну сторону от него расположен ганглий , служащий мозгом. Нервы идут от кольца и узла к щупальцам и остальной части тела. [18] У мшанок нет специализированных органов чувств, но роль сенсоров выполняют реснички на щупальцах. Представители рода Bugula растут по направлению к солнцу и поэтому должны уметь обнаруживать свет. [18] В колониях некоторых видов сигналы передаются между зооидами через нервы, которые проходят через поры в стенках тела и координируют такие действия, как питание и втягивание лофофоров. [18]
Одиночные особи Monobryozoon представляют собой автозооиды с телом грушевидной формы. На более широких концах имеется до 15 коротких мускулистых выступов, с помощью которых животные прикрепляются к песку или гравию. [35] и вытащить себя через отложения. [36]
Авикулярия и вибракула
[ редактировать ]Некоторые авторитеты используют термин авикулярия (множественное число от авикулярий ) для обозначения любого типа зооидов, у которых лофофор заменен расширением, выполняющим некоторую защитную функцию. [32] в то время как другие ограничивают этот термин теми, кто защищает колонию, нападая на захватчиков и мелких хищников, убивая одних и кусая придатки других. [18] У некоторых видов щелкающие зооиды прикреплены к стеблю (стебельку), их птичий внешний вид и послужил причиной возникновения термина - Чарльз Дарвин описал их как «голову и клюв стервятника в миниатюре, сидящих на шее и способных двигаться». ". [18] [32] Черешковые авикулярии располагаются на стеблях в перевернутом виде. [28] «Нижние челюсти» представляют собой модифицированные версии жаберных крышек, которые защищают втянутые лофофоры у автозооидов некоторых видов, и защелкиваются «как мышеловка» аналогичными мышцами. [18] а верхняя челюсть в форме клюва представляет собой перевернутую стенку тела. [28] У других видов авикулярии представляют собой неподвижные коробчатые зооиды, уложенные обычным образом вверх, так что измененная крышка прижимается к стенке тела. [28] У обоих типов видоизмененная жаберная крышка открывается другими прикрепляющимися к ней мышцами. [32] или внутренними мышцами, которые повышают давление жидкости, натягивая гибкую мембрану. [18] Действия этих щелкающих зооидов контролируются небольшими сильно модифицированными полипидами, расположенными внутри «рта» и несущими пучки коротких сенсорных ресничек . [18] [28] Эти зооиды появляются в различных положениях: некоторые заменяют автозооиды, некоторые вписываются в небольшие промежутки между автозооидами, а на поверхности других зооидов могут встречаться небольшие авикулярии. [32]
У вибракулы, которую некоторые считают разновидностью авикулярии, жаберная крышка модифицирована и образует длинную щетину, имеющую широкий диапазон движений. Они могут действовать как защита от хищников и захватчиков или как очистители. У некоторых видов, образующих подвижные колонии, вибракулы по краям используются как ноги для рытья нор и ходьбы. [18] [32]
Структурные полиморфы
[ редактировать ]Кенозооиды (от греческого kenós «пустой») [37] состоят только из стенки тела и тяжей фуникулёра, пересекающих внутреннюю часть, [18] и нет полипида. [28] В функции этих зооидов входит формирование стеблей ветвящихся структур, выступающих в роли прокладок, позволяющих колониям быстро расти в новом направлении; [28] [32] укрепление ветвей колонии и немного приподнятие колонии над субстратом для получения конкурентных преимуществ перед другими организмами. Предполагается, что некоторые кенозооиды способны хранить питательные вещества для колонии. [38] Поскольку функция кенозооидов в целом является структурной, их называют «структурными полиморфами».
Считается, что некоторые гетерозооиды, обнаруженные у вымерших трепостомных мшанок, называемые мезозооидами, функционировали для того, чтобы располагать питающиеся автозооиды на соответствующем расстоянии друг от друга. В тонких срезах окаменелостей трепостомов можно увидеть мезозооиды между трубками, в которых находились автозооиды; это трубки меньшего размера, разделенные по длине диафрагмами, что делает их похожими на ряды коробчатых камер, зажатых между аутозооидальными трубками. [39]
Репродуктивные полиморфы
[ редактировать ]Гонозуиды действуют как выводковые камеры для оплодотворенных яиц. [28] Они есть почти у всех современных круглоротых мшанок, но их бывает трудно обнаружить в колонии, поскольку в одной колонии очень мало гонозооидов. Отверстие гонозооидов, называемое оэциопорой, служит точкой выхода личинок. Некоторые гонозуиды имеют очень сложную форму с автозооидными трубками, проходящими через камеры внутри них. Все личинки, выпущенные из гонозооида, представляют собой клоны, созданные путем деления одного яйца; это называется монозиготной полиэмбрионией , и это репродуктивная стратегия, также используемая броненосцами . [40]
Хейлостомные мшанки также высиживают свои эмбрионы; один из распространенных методов - через яйцеклетки, капсулы, прикрепленные к аутозооидам. Однако автозооиды, обладающие яйцеклетками, обычно все еще способны питаться, поэтому они не считаются гетерозооидами. [41]
«Женские» полиморфы встречаются чаще, чем «мужские», но известны и специализированные зооиды, вырабатывающие сперму. Их называют андрозооидами, и некоторые из них встречаются в колониях Odontoporella Bishopi , вида, который симбиотичен с раками-отшельниками и живет на их панцирях. Эти зооиды меньше остальных и имеют четыре коротких и четыре длинных щупальца, в отличие от автозооидов, у которых 15–16 щупалец. Андрозооиды также встречаются у видов с подвижными колониями, которые могут ползать. Возможно, что андрозооиды используются для обмена спермой между колониями, когда две подвижные колонии или раки-отшельники, покрытые мшанками, встречаются друг с другом. [42]
Другие полиморфы
[ редактировать ]Спинозооиды представляют собой полые подвижные шипы, похожие на очень тонкие маленькие трубочки, присутствующие на поверхности колоний и, вероятно, предназначенные для защиты. [43] У некоторых видов есть миниатюрные нанозооиды с небольшими полипидами с одним щупальцем, и они могут расти на других зооидах или внутри стенок тела деградировавших автозооидов. [32]
Формы и состав колоний
[ редактировать ]

Хотя зооиды микроскопичны, размеры колоний варьируются от От 1 см ( 1 ⁄ 2 дюйма) до более 1 м (3 футов 3 дюйма). [18] Однако большинство из них имеют диаметр менее 10 см (4 дюйма). [21] Формы колоний широко варьируются и зависят от характера почкования, по которому они растут, разнообразия присутствующих зооидов, а также типа и количества выделяемого ими скелетного материала . [18]
Некоторые морские виды имеют кустообразную или веерообразную форму, поддерживаемую «стволами» и «ветвями», образованными кенозооидами, из которых растут питающиеся автозооиды. Колонии этих типов обычно не минерализованы , но могут иметь экзоскелет из хитина . [18] Другие выглядят как маленькие кораллы , образующие тяжелые известковые скелеты. [44] Многие виды образуют колонии, состоящие из листов автозооидов. Эти листы могут образовывать листья, пучки или, у рода Thalamoporella , структуры, напоминающие раскрытый кочан салата. [18]
Однако наиболее распространенной морской формой является корковая, при которой однослойный лист зооидов распространяется по твердой поверхности или водорослям. Некоторые покрытые коркой колонии могут вырастать до более 50 см (1 фут 8 дюймов) и содержать около 2 000 000 зооидов. [18] Эти виды обычно имеют экзоскелеты, усиленные карбонатом кальция , а отверстия, через которые выступают лофофоры, находятся на верхней или внешней поверхности. [18] Внешний вид покрытых коркой колоний, напоминающий мох, и послужил причиной названия типа ( древнегреческие слова βρύον brúon , означающие «мох», и ζῷον zôion, означающие «животное»). [45] Большие колонии инкрустирующих видов часто имеют « дымоходы » — щели в пологе лофофоров, через которые они быстро выбрасывают просеянную воду и таким образом избегают повторной фильтрации уже истощенной воды. [46] Они образованы участками непитающихся гетерозооидов. [47] Новые дымоходы появляются вблизи краев расширяющихся колоний, в местах, где скорость истечения уже высока, и не меняют положения при изменении расхода воды. [48]
Некоторые пресноводные виды выделяют массу студенистого материала диаметром до 1 м (3 фута 3 дюйма), к которому прилипают зооиды. Другие пресноводные виды имеют растительную форму со «стволами» и «ветвями», которые могут стоять прямо или стелиться по поверхности. Некоторые виды могут ползти примерно на 2 см ( дюйма ) 3/4 в день. [18]
Каждая колония растет путем бесполого почкования от одного зооида, известного как предок. [18] который имеет круглую форму, а не форму обычного зооида. [21] Это происходит на кончиках «стволов» или «ветвей» у форм, имеющих такое строение. По их краям растут корковые колонии. У видов с известковым экзоскелетом они не минерализуются до тех пор, пока зооиды не вырастут полностью. Продолжительность жизни колонии колеблется от одного до примерно 12 лет, а недолговечные виды проходят через несколько поколений за один сезон. [18]
Виды, производящие защитные зооиды, делают это только тогда, когда угроза уже появилась, и могут сделать это в течение 48 часов. [28] Теория «индуцированной защиты» предполагает, что создание защиты обходится дорого и что колонии, которые защищаются слишком рано или слишком сильно, будут иметь снижение темпов роста и продолжительности жизни. Такой подход к защите «в последнюю минуту» возможен, поскольку потери зооидов в результате одной атаки вряд ли будут значительными. [28] Колонии некоторых покрывающих корку видов также производят специальные гетерозооиды, чтобы ограничить распространение других покрывающих корку организмов, особенно других мшанок. В некоторых случаях эта реакция становится более агрессивной, если противник меньше, что позволяет предположить, что зооиды на краю колонии каким-то образом могут чувствовать размер противника. Некоторые виды постоянно побеждают некоторые другие, но большинство войн за сферы влияния являются нерешительными, и участники боевых действий вскоре переходят к выращиванию на неоспоримых территориях. [28] Мшанки, конкурирующие за территорию, не используют сложные методы, используемые губками или кораллами , возможно, потому, что короткая продолжительность жизни мшанок делает крупные инвестиции в войны за сферы влияния нерентабельными. [28]
Мшанки внесли свой вклад в осаждение карбонатов в морской жизни начиная с ордовикского периода. Мшанки берут на себя ответственность за многие формы колоний, которые развились в разные таксономические группы и различаются по способности образовывать отложения. Девять основных форм колоний мшанок включают: корковую, куполообразную, пальчатую, листоватую, окончатую, сильное ветвление, тонкое ветвление, сочлененную и свободноживущую. Большая часть этих отложений происходит из двух отдельных групп колоний: купольных, с тонким ветвлением, сильно ветвящихся и пальчатых; и фенестрировать. Колонии фенестратов образуют грубые частицы как в виде осадка, так и в виде компонентов строматопороидных коралловых рифов. Однако хрупкие колонии образуют как грубые осадки, так и ядра глубоководных субфотических биогенных холмиков. Почти все постмшашковые отложения представлены ростовыми формами с добавлением свободноживущих колоний, включающих значительное количество различных колоний. «В отличие от палеозоя, постпалеозойские мшанки образовывали осадки, более широко варьирующиеся в зависимости от размера их зерен; они растут по мере перемещения из ила в песок и гравий». [49]
Таксономия
[ редактировать ]

Первоначально этот тип назывался «Polyzoa», но в конечном итоге это название было заменено термином Эренберга «Bryozoa». [13] [50] [51] Название «Bryozoa» первоначально применялось только к животным, также известным как Ectoprocta ( букв. « вне ануса » ). [52] при котором анус лежит вне «макушки» щупалец. После открытия Entoprocta ( букв . « внутри ануса » ) [53] у которых анус находится внутри «короны» из щупалец, название «Bryozoa» было повышено до уровня типа , включив в него два класса Ectoprocta и Entoprocta. [54] Однако в 1869 году Хинрих Ницше по ряду причин считал эти две группы совершенно разными и придумал название «Ectoprocta» для «Bryozoa» Эренберга. [5] [55] Несмотря на кажущуюся схожесть методов питания, они заметно различались анатомически; Помимо различного положения ануса, эктопрокты имеют полые щупальца и целом , а у энтопроктов твердые щупальца и нет целома. Следовательно, эти две группы теперь широко рассматриваются как отдельные типы, а название «Bryozoa» теперь является синонимом «Ectoprocta». [54] С тех пор это мнение большинства, хотя большинство публикаций предпочитают название «Bryozoa», а не «Ectoprocta». [51] Тем не менее, некоторые известные ученые продолжают считать «Ectoprocta» и Entoprocta близкими родственниками и группировать их под «Bryozoa». [55]
Неясность в отношении объема названия «Bryozoa» привела к предложениям в 1960-х и 1970-х годах избегать его и использовать однозначный термин «Ectoprocta». [56] Однако это изменение затруднило бы поиск более старых работ, в которых этот тип назывался «Мшанки», а желание избежать двусмысленности, если оно последовательно применялось ко всем классификациям, потребовало бы переименования нескольких других типов и многих более низких уровней. группы. [50] На практике зоологическое наименование разделенных или объединенных групп животных является сложным и не вполне последовательным. [57] В работах, начиная с 2000 года, чтобы устранить двусмысленность, использовались различные названия, в том числе: «Мшанка», [18] [21] «Эктопрокта», [24] [28] «Мшанка (Эктопрокта)», [31] и «Эктопрокта (Мшанка)». [58] Некоторые использовали более одного подхода в одной и той же работе. [59]
Общее название «моховые животные» является буквальным значением слова «Bryozoa» от греческого βρυόν («мох») и ζῷα («животные»), основанного на мшистом внешнем виде покрытых коркой видов. [60]
До 2008 года существовали «недостаточно известные и неправильно понятые типовые виды, принадлежащие к семейству круглоротых мшанок Oncousoeciidae». [61] Современные исследования и эксперименты были проведены с использованием сканирующей электронной микроскопии в низком вакууме материала без покрытия для критического изучения и, возможно, пересмотра таксономии трех родов, принадлежащих к этому семейству, включая Oncousoecia , Microeciella и Eurystrotos . Этот метод позволяет получить данные, которые было бы трудно распознать с помощью оптического микроскопа. Действительным типовым видом Oncousoecia оказался Oncousoecia lobulata . Эта интерпретация стабилизирует Oncousoecia , устанавливая типовой вид, который соответствует общему использованию этого рода. В настоящее время считается, что его товарищ по Oncousoeciid Eurystrotos не является конспецифичным O. lobulata , как предполагалось ранее, но демонстрирует достаточное сходство, чтобы его можно было считать младшим синонимом Oncousoecia . Microeciella suborbcularus также недавно была отделена от O. lobulata и O. dilatans с использованием этого современного метода сканирования в низком вакууме, с которым в прошлом ее ошибочно ассоциировали. Недавно был открыт новый род под названием Junerossia в семействе Stomachetosellidae, а также 10 относительно новых видов мшанок, таких как Alderina flaventa , Corbulella extenuata , Puellina septemcryptica , Junerossia copiosa , Calyptotheca kapaaensis , Bryopesanser serratus , Cribellopora Souleorum , Metacleidochasma verrucosa , Dysporella compta и Favo . сипора адунка. [62]
Классификация и разнообразие
[ редактировать ]Количество официально описанных видов колеблется от 4000 до 4500. [63] У Gymnolaemata и особенно Cheilostomata наибольшее количество видов, возможно, из-за широкого спектра специализированных зооидов. [28] Согласно системе классификации Линнея , которая до сих пор используется как удобный способ обозначения групп организмов, [64] живые представители типа Bryozoa делятся на: [18] [28]
| Сорт | Филактолематы | Стенолемата | Гимнолематы | |
|---|---|---|---|---|
| Заказ | Плюмателлида [65] | Циклостоматида | Ктеностоматида | Хейлостоматы |
| Окружающая среда | Пресноводный | Морской | В основном морской | |
| Губовидный эпистом, нависающий над ртом | Да | никто | ||
| Формы колоний | Желатиновые массы или трубчатые ветвящиеся структуры [66] | Прямостоячий или инкрустирующий [67] | Прямостоячие, инкрустированные или свободноживущие | |
| Материал экзоскелета | Студенистый или пленчатый; неминерализованный | минерализованный | Хитин студенистый или пленчатый; неминерализованный | минерализованный |
| Оперкулум («крышка») | никто | никто [67] (кроме семейства Eleidae ) [68] | Нет у большинства видов | Да (кроме рода Bugula ) |
| Форма лофофора | U-образный внешний вид (кроме рода Fredericella , у которого лофофор круглый) | Круговой | ||
| Как лофофор распространялся | Сжатие всей стенки тела | Сдавление перепончатого мешка (отдельный внутренний слой эпителия , выстилающий целом) | Сжатие всей стенки тела | Втягивание внутрь гибкого участка стенки тела или расширение внутреннего мешка. |
| Виды зооидов | Только автозооиды | Ограниченные гетерозооиды, преимущественно гонозооиды. [69] | Столоны и шипы, а также автозооиды [69] | Полный спектр типов |
Ископаемая запись
[ редактировать ]
| Стереоизображение | |||
|---|---|---|---|
| |||
| |||
| |||
| |||
| Ископаемый скелет Архимеда мшанки | |||

Были найдены окаменелости около 15 000 видов мшанок. Мшанки входят в число трех доминирующих групп палеозойских окаменелостей. [70] Мшанки с кальцитовым скелетом были основным источником карбонатных минералов, составляющих известняки, а их окаменелости невероятно распространены в морских отложениях по всему миру, начиная с ордовика. Однако, в отличие от кораллов и других колониальных животных, обнаруженных в летописи окаменелостей, колонии мшанок не достигали больших размеров. [71] Ископаемые колонии мшанок обычно встречаются сильно фрагментированными и разбросанными; Сохранение полной зоарии редко встречается в летописи окаменелостей, и относительно мало исследований было посвящено повторной сборке фрагментированных зоарий. [72] Самые крупные известные ископаемые колонии — это ветвящиеся трепостомные мшанки из ордовикских пород в США, достигающие 66 сантиметров в высоту. [71]
Древнейший вид с минерализованным скелетом встречается в нижнем ордовике . [1] Вполне вероятно, что первые мшанки появились гораздо раньше и были полностью мягкотелыми, а ордовикские окаменелости фиксируют появление минерализованных скелетов в этом типе. [5] К аренигскому этапу раннего ордовика , [21] [73] около 480 миллионов лет назад современные отряды стенолемат , присутствовали все [74] а гребневиков отряд гимнолематов появился к среднему ордовику, около 465 миллионов лет назад . Окаменелости раннего ордовика могут также представлять собой формы, которые уже значительно отличались от первоначальных членов филума. [74] С девона известны гребневики с фосфатизированной мягкой тканью. [75] Примерно в то же время появились и другие виды фильтраторов , что говорит о том, что некоторые изменения сделали окружающую среду более благоприятной для такого образа жизни. [21] Окаменелости хейлостомат , отряда голосематов с минерализованным скелетом, впервые появляются в средней юре , около 172 миллионов лет назад , и это были самые многочисленные и разнообразные мшанки от мела до настоящего времени. [21] Данные, собранные за последние 100 миллионов лет, показывают, что хейлостоматиды постоянно вырастали над циклостоматидами в территориальной борьбе, что может помочь объяснить, как хейлостоматиды заменили циклостоматидами в качестве доминирующих морских мшанок. [76] Морские окаменелости палеозойской эры , завершившейся 251 миллион лет назад , представлены преимущественно прямостоячими формами, мезозойские примерно поровну разделены на прямостоячие и корковые формы, а более поздние — преимущественно корковые. [77] Окаменелости мягких пресноводных филактолематов очень редки. [21] появляются в поздней перми и после нее (начавшейся около 260 миллионов лет назад ) и полностью состоят из прочных статобластов. [66] Окаменелости пресноводных представителей других классов неизвестны. [66]
Эволюционное генеалогическое древо
[ редактировать ]
Ученые расходятся во мнениях относительно того, являются ли мшанки (Ectoprocta) монофилетической группой (включают ли они все и только один вид-предок и все его потомки), каковы ближайшие родственники этого типа в генеалогическом древе животных и даже относительно того, являются ли они следует рассматривать как представителей протостомов или вторичноротых , двух основных групп, которые составляют всех умеренно сложных животных.
Молекулярная филогения, которая пытается разработать эволюционное генеалогическое древо организмов путем сравнения их биохимии и особенно их генов , многое сделала для выяснения отношений между более известными типами беспозвоночных . [54] Однако нехватка генетических данных о «второстепенных типах», таких как мшанки и энтопрокты, оставила неясными их отношения с другими группами. [55]
Традиционный вид
[ редактировать ]Традиционное мнение состоит в том, что Bryozoa представляют собой монофилетическую группу, в которой класс Phylactolaemata наиболее тесно связан с Stenolaemata и Ctenostomatida , классами , которые появляются раньше всего в летописи окаменелостей. [78] Однако в 2005 году исследование молекулярной филогении , посвященное филактолематам, пришло к выводу, что они более тесно связаны с типом Phoronida и особенно с единственным колониальным видом форонид, чем с другими классами эктопроктов. Это означает, что Entoprocta не являются монофилетическими, поскольку Phoronida представляют собой подгруппу эктопроктов, но стандартное определение Entoprocta исключает Phoronida. [78]

В 2009 году другое исследование молекулярной филогении с использованием комбинации генов митохондрий и клеточного ядра пришло к выводу, что мшанки являются монофилетическим типом, другими словами, включают всех потомков общего предка, который сам является мшанками. Анализ также пришел к выводу, что классы Phylactolaemata, Stenolaemata и Gymnolaemata также являются монофилетическими, но не смог определить, являются ли Stenolaemata более близкими родственниками Phylactolaemata или Gymnolaemata . Gymnolaemata традиционно делятся на Ctenostomatida с мягким телом и минерализованные Cheilostomata, но анализ 2009 года показал, что более вероятно, что ни один из этих отрядов не является монофилетическим и что минерализованные скелеты , вероятно, развивались более одного раза в пределах ранних Gymnolaemata. [5]
Отношения мшанок с другими типами неопределенны и противоречивы. Традиционная филогения, основанная на анатомии и развитии взрослых форм из эмбрионов , не привела к устойчивому консенсусу относительно положения эктопроктов. [24] Попытки реконструировать генеалогическое древо животных в значительной степени игнорировали эктопроктов и другие «второстепенные типы», которые мало изучены научными исследованиями, поскольку они, как правило, крошечные, имеют относительно простое строение тела и мало влияют на экономику человека – несмотря на то, что «Младшие типы» включают большую часть разнообразия в истории эволюции животных. [80]
По мнению Рут Дьюэл, Джудит Уинстон и Фрэнка МакКинни, «наша стандартная интерпретация морфологии и эмбриологии мшанок представляет собой конструкцию, возникшую в результате более чем 100-летних попыток синтезировать единую структуру для всех беспозвоночных», и мало учитывает некоторые специфические особенности. Особенности эктопроктов. [74]


У эктопроктов все внутренние органы личинки разрушаются во время метаморфоза во взрослую форму, и органы взрослой особи строятся из эпидермиса и мезодермы личинки , тогда как у других билатерий некоторые органы, включая кишечник, построены из энтодермы . У большинства двусторонних эмбрионов бластопор, вмятина на внешней стенке, углубляется и становится кишкой личинки, но у эктопроктов бластопор исчезает, и новая вмятина становится точкой, из которой вырастает кишечник. Целом эктопрокта не образуется ни за счет процессов, используемых другими билатериями, ни энтероцелии , при которой мешочки, образующиеся на стенке кишки, становятся отдельными полостями, ни шизоцелии , при которой ткань между кишкой и стенкой тела расщепляется, образуя парные полости. [74]
Энтопрокты
[ редактировать ]Когда в 19 веке были открыты энтопрокты, они и мшанки (эктопрокты) рассматривались как классы внутри типа Bryozoa, поскольку обе группы были сидячими животными, которые осуществляли фильтрацию с помощью короны щупалец с ресничками .
Начиная с 1869 года растущее осознание различий, включая положение ануса энтопрокта внутри структуры питания и разницу в раннем характере деления клеток эмбрионов , заставило ученых рассматривать эти две группы как отдельные типы. [55] а «Мшанки» стали всего лишь альтернативным названием эктопроктов, у которых анус находится вне органа питания. [54] Серия исследований молекулярной филогении с 1996 по 2006 год также пришла к выводу, что мшанки (эктопрокты) и энтопрокты не являются сестринскими группами. [55]
Однако два известных зоолога, Клаус Нильсен и Томас Кавальер-Смит , на основании анатомии и развития утверждают, что мшанки и энтопрокты являются членами одного и того же типа, Bryozoa. Исследование молекулярной филогении, проведенное в 2007 году, также подтвердило эту старую идею, а его выводы о других типах согласовались с выводами нескольких других анализов. [55]
Группировка в Lophophorata
[ редактировать ]К 1891 году мшанки (эктопрокты) были сгруппированы с форонидами в супертип под названием «Tentaculata». В 1970-х годах сравнение личинок форонид и личинок cyphonautes некоторых мшанок Gymnolaete привело к предположению, что мшанки, большинство из которых являются колониальными, произошли от полуколониальных видов форонид. [81] Брахиоподы также были отнесены к «Tentaculata», которые были переименованы в Lophophorata , поскольку все они используют лофофор для фильтрационного питания. [54]
С этим согласны большинство учёных. [54] но Клаус Нильсен считает, что эти сходства поверхностны. [24] Lophophorata обычно определяют как животных с лофофором, трехчастным целомом и U-образной кишкой. [81] По мнению Нильсена, лофофоры форонид и брахиопод больше похожи на лофофоры крыложаберных . [24] которые относятся к типу Hemichordata . [82] Щупальца мшанок несут клетки с множественными ресничками , тогда как соответствующие клетки лофофоров форонид, брахиопод и крыложаберных имеют по одной ресничке на клетку; а щупальца мшанок не имеют гемального канала («кровеносного сосуда»), который есть у трех других типов. [24]
Если группировка мшанок с форонидами и брахиоподами в Lophophorata верна, следующий вопрос заключается в том, являются ли Lophophorata протостомами , наряду с большинством типов беспозвоночных, или вторичноротыми , наряду с хордовыми , полухордовыми и иглокожими .
Традиционное мнение заключалось в том, что лофофораты представляют собой смесь особенностей протостома и вторичноротого. Исследования, начиная с 1970-х годов, показали, что это вторичноротые из-за некоторых особенностей, которые считались характерными для вторичноротых: целом, состоящий из трех частей; радиальное, а не спиральное дробление в развитии зародыша; [54] и образование целома путем энтероцелии . [24] Однако целом личинок эктопрокта не имеет признаков разделения на три отдела. [81] а у взрослых эктопроктов отличается от такового у других типов целоматов , поскольку он строится заново из эпидермиса и мезодермы после того, как метаморфоз разрушил личиночный целом. [74]
Молекулярная филогенетика лофофората
[ редактировать ]Анализ молекулярной филогении, начиная с 1995 года, с использованием различных биохимических данных и аналитических методов, поместил лофофораты как протостомов и тесно связанных с кольчатыми червями и моллюсками в супертипе под названием Lophotrochozoa . [54] [83] Анализ «полных доказательств», в котором использовались как морфологические особенности, так и относительно небольшой набор генов, пришел к различным выводам, в основном в пользу тесной связи между лофофоратами и Lophotrochozoa. [83] Исследование 2008 года с использованием большего набора генов пришло к выводу, что лофофораты были ближе к Lophotrochozoa, чем к вторичноротым, но также и что лофофораты не были монофилетическими. Вместо этого он пришел к выводу, что брахиоподы и форониды образуют монофилетическую группу, но мшанки (эктопрокты) были ближе всего к энтопроктам, что подтверждает первоначальное определение «мшанки». [83]
Это единственный крупный тип исключительно клональных животных, состоящий из модульных единиц, известных как зооиды. Поскольку они процветают в колониях, колониальный рост позволяет им развивать неограниченные вариации формы. Несмотря на это, было обнаружено лишь небольшое количество основных форм роста, которые обычно появлялись вновь на протяжении всей истории мшанок. [70]


Молекулярная филогенетика эктопрокта
[ редактировать ]Филогенетическое положение эктопроктовых мшанок остается неопределенным, но остается достоверным, что они принадлежат к Protostomia, а точнее к Lophotrochozoa. Это означает, что личинка эктопрокта представляет собой трохофор, коронка которого является гомологом прототроха; Об этом свидетельствует сходство коронатных личинок с личинками перикалиммы 1-го типа некоторых моллюсков и сипункуланов, у которых зона прототроха расширена и охватывает гипосферу. [84]
Исследование последовательности митохондриальной ДНК предполагает, что мшанки могут быть связаны с Chaetognatha . [85]
Физиология
[ редактировать ]Питание и выделение
[ редактировать ]Большинство видов являются фильтраторами мелкие частицы, в основном фитопланктон (микроскопические плавающие растения). , которые фильтруют из воды [18] Пресноводный вид Plumatella emarginata питается диатомовыми водорослями , зелеными водорослями , цианобактериями , нефотосинтезирующими бактериями , динофлагеллятами , коловратками , простейшими , мелкими нематодами и микроскопическими ракообразными . [86] Хотя токи, которые генерируют мшанки, чтобы доставить пищу ко рту, хорошо изучены, точный метод захвата все еще обсуждается. Все виды также подбрасывают более крупные частицы ко рту с помощью щупалец, а некоторые захватывают зоопланктон (планктонных животных), используя свои щупальца в качестве клеток. Кроме того, щупальца, площадь поверхности которых увеличена за счет микроворсинок (мелких волосков и складок), поглощают органические соединения . растворенные в воде [18] Нежелательные частицы можно отмахнуть щупальцами или закрыть рот, закрыв рот. [18] Исследование 2008 года показало, что как покрытые коркой, так и стоячие колонии питаются быстрее и растут быстрее при слабом течении, чем при сильном. [87]
У некоторых видов первая часть желудка образует мускулистый желудок, выстланный хитиновыми зубами, которые раздавливают бронированную добычу, например, диатомовые водоросли . Волнообразные перистальтические сокращения продвигают пищу через желудок для переваривания. Последний отдел желудка покрыт ресничками (мельчайшими волосками), которые сжимают непереваренные твердые вещества, которые затем проходят через кишечник и выходят через задний проход . [18]
отсутствуют . Нефридии («почки») и другие органы выделения у мшанок [28] Считается, что аммиак диффундирует через стенки тела и лофофор. [18] Более сложные продукты жизнедеятельности не выводятся, а накапливаются в полипиде , который дегенерирует через несколько недель. Часть старых полипидов перерабатывается, но большая их часть остается в виде большой массы умирающих клеток, содержащих накопленные отходы, и она сжимается в «коричневое тело». Когда дегенерация завершена, цистида (внешняя часть животного) производит новый полипид, а коричневое тело остается в целоме или желудке нового полипида и выбрасывается при следующей дефекации животного . [18]
Дыхание и кровообращение
[ редактировать ]нет Органов дыхания, сердца и кровеносных сосудов . Вместо этого зооиды поглощают кислород и удаляют углекислый газ путем диффузии. Мшанки осуществляют диффузию либо с помощью тонкой мембраны (у анасканов и некоторых многоклеточных), либо через псевдопоры, расположенные на внешней дерме зооида. [88] Различные группы мшанок используют различные методы для распределения питательных веществ и кислорода между зооидами: у некоторых есть довольно большие промежутки в стенках тела, позволяющие целомической жидкости свободно циркулировать; у других - канатики (внутренние «веревочки»). [34] соседних зооидов соединяются через небольшие поры в стенке тела. [18] [31]
Размножение и жизненные циклы
[ редактировать ]
Зооиды всех видов филактолематов одновременно являются гермафродитами . Хотя представители многих морских видов являются протандрическими, то есть функционируют сначала как самцы, а затем как самки, их колонии содержат комбинацию зооидов, находящихся на мужской и женской стадиях. У всех видов яичники развиваются на внутренней стороне стенки тела, а семенники - на канатике, соединяющем желудок со стенкой тела. [28] Яйца и сперматозоиды выбрасываются в целом, а сперматозоиды выходят в воду через поры на кончиках некоторых щупалец, а затем захватываются питающими токами зооидов, производящих яйца. [18] Яйца некоторых видов оплодотворяются снаружи после выхода через пору между двумя щупальцами, которая в некоторых случаях находится на кончике небольшого выступа, называемого «межщупальцевым органом», в основании пары щупалец. У других оплодотворение происходит внутри, в межщупальцевом органе или целоме. [18]
Все филактолематы и стенолематы, а также большинство гимнолематов имеют плацентацию и, следовательно, имеют лецитотрофные (не питающиеся) личинки. За исключением круглоротых и небольшого голосематного семейства Epistomiidae, которые являются живородящими, все они являются выводками. Phylactolaemata высиживает свои эмбрионы во внутреннем выводковом мешке, но у Gymnolaemata существуют как внешние мембранные мешки, скелетные камеры (яйцеклетки), так и внутренние выводковые мешки. [89] [90] Развивающийся эмбрион зависит от яичного желтка, внеэмбрионального питания (матротрофии) или того и другого. [91]
У гребневиков мать обеспечивает выводковую камеру для оплодотворенных яиц, а ее полипид распадается, обеспечивая питание зародыша . Stenolaemate производят специализированные зооиды, которые служат выводковыми камерами, и их яйца делятся внутри них, образуя до 100 идентичных эмбрионов. [28]
Планктотрофные (питающиеся) личинки встречаются только в классе Gymnolaemata: в подотряде хейлостоматов Malacostegina они встречаются в двух семействах Membraniporidae и Electridae, а также в трех семействах гребневиков Alcyonidiidae, Farrellidae и Hislopiidae. Кроме того, есть несколько неподтвержденных записей, таких как одиночная форма Aethozoid, личинки которой никогда не наблюдались, но предположительно имеют планктотрофные личинки. [92] [93] [94]
Дробление . яиц мшанок двулучевое, то есть на ранних стадиях двусторонне-симметричное Как формируется целом, неизвестно, поскольку при во взрослую метаморфозе личинки особь разрушаются все внутренние ткани личинки. У многих животных бластопор — отверстие на поверхности раннего эмбриона — туннелирует, образуя кишечник. Однако у мшанок бластопор закрывается, и образуется новое отверстие, образующее рот. [18]
Личинки мшанок различаются по форме, но все они имеют полоску ресничек вокруг тела, которая позволяет им плавать, пучок ресничек наверху и клейкий мешок, который выворачивает и фиксирует их, когда они оседают на поверхность. [18] Некоторые виды Gymnolaemate производят личинок cyphonautes, у которых мало желтка, но хорошо развиты рот и кишечник, и они живут в виде планктона в течение значительного времени, прежде чем оседать. Эти личинки имеют треугольные хитиновые раковины с одним углом вверху и открытым основанием, образующим капюшон вокруг обращенного вниз рта. [28] В 2006 году сообщалось, что реснички личинок cyphonautes используют тот же набор методов, что и взрослые особи, для захвата пищи. [95] Виды, высиживающие зародыши, образуют личинок, питающихся крупными желтками , не имеющих кишечника и не питающихся, такие личинки быстро оседают на поверхности. [18] У всех морских видов личинки образуют коконы, в которых после расселения они полностью метаморфизуются личинки : эпидермис становится выстилкой целома , а внутренние ткани превращаются в запас пищи, питающий развивающегося зооида до тех пор, пока он не будет готов к питанию. [18] Личинки филактолематов образуют множественные полипиды, так что каждая новая колония начинается с нескольких зооидов. [18] У всех видов зооиды-основатели затем выращивают новые колонии, отпочковываясь от своих клонов. У филактолематов зооиды умирают после создания нескольких клонов , поэтому живые зооиды обнаруживаются только по краям колонии. [18]
Филактолематы также могут размножаться бесполым путем, используя метод, который позволяет линии колонии выживать в изменчивых и неопределенных условиях пресноводной среды. [28] В течение лета и осени они производят дискообразные статобласты, массы клеток, которые функционируют как «стручки выживания», подобно геммулам губок . [18] Статобласты образуются на канатике, соединенном с кишечником родителя, который их питает. [28] По мере роста статобласты образуют защитные двустворчатые оболочки из хитина . По мере созревания часть статобластов прикрепляется к родительской колонии, часть падает на дно («сессобласты»), часть содержит воздушные пространства, позволяющие им плавать («флоатобласты»). [18] а некоторые остаются в цистиде родителя, чтобы восстановить колонию в случае ее гибели. [28] Статобласты могут оставаться в состоянии покоя в течение значительных периодов времени и, находясь в состоянии покоя, могут выдерживать суровые условия, такие как замерзание и высыхание . Их можно переносить на большие расстояния с помощью животных, плавучей растительности, течений. [18] и ветры, [28] и даже в кишках более крупных животных. [96] Когда условия улучшаются, створки панциря разделяются, и клетки внутри развиваются в зооида, который пытается сформировать новую колонию. Plumatella emarginata производит как «сессобласты», которые позволяют линии контролировать хорошую территорию, даже если тяжелые времена уничтожают родительские колонии, так и «флоатобласты», которые распространяются на новые участки. Новые колонии Plumatella repens производят в основном «сессобласты», тогда как зрелые переходят на «флоатобласты». [86] По оценкам исследования, одна группа колоний на участке площадью 1 квадратный метр (11 квадратных футов) произвела 800 000 статобластов. [18]
Cupuladriid Bryozoa способны как к половому, так и к бесполому размножению. Колонии, размножающиеся половым путем (аклональные), являются результатом превращения личинки купуладриид во взрослую стадию, тогда как бесполые колонии (клональные) являются результатом превращения фрагмента колонии купуладриид в собственную колонию. Различные формы размножения у купуладриид достигаются различными способами в зависимости от морфологии и классификации зооида. [97]
Экология
[ редактировать ]Места обитания и распространение
[ редактировать ]Большинство морских видов обитают в тропических водах на глубине менее 100 метров (330 футов; 55 саженей). Однако некоторые из них были найдены в глубоководных траншеях . [98] особенно вокруг холодных просачиваний и других вблизи полюсов . [99] [100]
Подавляющее большинство мшанок сидячие . Обычно сидячие мшанки живут на твердых субстратах, включая камни, песок или ракушки. [101] Покрытые коркой формы наиболее распространены в мелководных морях, но по мере увеличения глубины встречаются прямостоячие формы. [99] Пример инкрустации гальки и булыжника можно найти у разнообразных мшанок плейстоцена, обнаруженных в северной Японии, где были обнаружены окаменелости отдельных камней, покрытых более чем 20 видами мшанок. [102] Отложения с более мелкими частицами, такими как песок или ил, обычно не являются подходящей средой обитания для мшанок, но были обнаружены крошечные колонии, инкрустирующие зерна крупного песка. [103] Некоторые виды мшанок специализируются на колонизации морских водорослей, морских трав и даже корней мангровых деревьев; Род Amphibiobeania живет на листьях мангровых деревьев и называется «земноводным», поскольку может выдерживать регулярное воздействие воздуха во время отлива. [104]
Существует множество «свободноживущих» мшанок, которые живут не прикрепленными к субстрату. Некоторые формы, такие как Кристелла, могут двигаться. Лунулитиформные хейлостомы представляют собой группу свободноживущих мшанок с подвижными колониями. Они образуют небольшие круглые колонии, не прикрепленные к субстрату; Было замечено, что колонии рода Selenaria «гуляют» с помощью щетинок. [105] Другое семейство хейлостом, Cupuladriidae, конвергентно развило колонии аналогичной формы, способные передвигаться. При наблюдении в аквариуме было зарегистрировано, что колонии Selenaria maculata ползают со скоростью один метр в час, перелезают друг через друга, движутся к свету и выпрямляются, когда их переворачивают. [106] Более позднее изучение этого рода показало, что нейроэлектрическая активность в колониях увеличивается пропорционально перемещению к источникам света. Предполагается, что способность к передвижению возникла как побочный эффект, когда у колоний появились более длинные щетинки для раскапывания себя из осадка. [106]

Другие свободноживущие мшанки свободно перемещаются под воздействием волн, течений или других явлений. Антарктический состоит из вид Alcyonidium pelagosphaera плавающих колоний. Пелагический вид имеет диаметр от 5,0 до 23,0 мм (от 0,20 до 0,91 дюйма), имеет форму полой сферы и состоит из одного слоя автозооидов. До сих пор неизвестно, являются ли эти колонии пелагическими всю свою жизнь или представляют собой лишь временную и ранее неописанную ювенильную стадию. [99] [107] Колонии вида Alcyonidium Disciforme , имеющего форму диска и столь же свободноживущего, населяют илистое морское дно в Арктике и могут улавливать поглощённые ими песчинки, потенциально используя песок в качестве балласта, чтобы перевернуться на правую сторону после того, как они был отменен. Некоторые виды мшанок могут образовывать бриолиты, свободноживущие колонии сферической формы, которые растут во всех направлениях, катаясь по морскому дну. [108]
В 2014 году сообщалось, что мшанка Fenestrulina Ruula стала доминирующим видом в некоторых частях Антарктиды. Глобальное потепление привело к увеличению скорости размыва айсбергами , и этот вид особенно хорош в повторном заселении очищенных территорий. [109]
Филактолематы обитают во всех типах пресноводной среды – озерах и прудах, реках и ручьях, эстуариях. [66] – и являются одними из самых многочисленных сидячих пресноводных животных. [78] Некоторые гребневики живут исключительно в пресной воде, другие предпочитают солоноватую воду, но могут выжить и в пресной воде. [66] Знания ученых о популяциях пресноводных мшанок во многих частях мира являются неполными, даже в некоторых частях Европы. Долгое время считалось, что некоторые пресноводные виды встречаются по всему миру, но с 2002 года все они были разделены на более локализованные виды. [66]
Мшанки растут клональными колониями. Личинка мшанки поселяется на твердом веществе и образует колонию бесполым путем путем почкования. Эти колонии могут вырастить тысячи отдельных зооидов за относительно короткий период времени. Хотя колонии зооидов растут за счет бесполого размножения, мшанки являются гермафродитами, и новые колонии могут образовываться путем полового размножения и образования свободно плавающих личинок. Однако, когда колонии становятся слишком большими, они могут разделиться на две части. Это единственный случай, когда бесполое размножение приводит к образованию новой колонии, отдельной от ее предшественницы. Большинство колоний стационарны. Действительно, эти колонии имеют тенденцию селиться на неподвижных веществах, таких как осадки и грубые вещества. Есть некоторые колонии пресноводных видов, таких как Cristatella mucedo , которые способны медленно передвигаться на ползучих ногах. [110]
Взаимодействие с нечеловеческими организмами
[ редактировать ]
Морские виды распространены на коралловых рифах , но редко составляют значительную долю общей биомассы . В умеренных водах скелеты погибших колоний составляют значительную часть ракушечника, а живые в этих районах многочисленны. [111] Морская кружевообразная мшанка Membranipora membranacea производит шипы в ответ на нападение нескольких видов морских слизней (голожаберных). [112] Другие хищники морских мшанок включают рыбу, морских ежей , пикногонид , ракообразных , клещей . [113] и морская звезда . [114] В целом морские иглокожие и моллюски поедают массы зооидов, выгрызая кусочки колоний, разрушая их минерализованные «дома», в то время как большинство членистоногих хищников мшанок поедают отдельных зооидов. [115]
В пресной воде мшанки являются одними из наиболее важных фильтраторов , наряду с губками и мидиями . [116] Пресноводные мшанки подвергаются нападению многих хищников, в том числе улиток, насекомых и рыб. [86]
В Таиланде Pomacea интродуцированный вид canaliculata (золотая яблочная улитка), который обычно является разрушительным травоядным животным , уничтожил популяции филактолемата везде, где он появлялся. P. canaliculata также охотится на обычного пресноводного гимнолемата, но с менее разрушительным эффектом. Местные улитки не питаются мшанками. [14]
Несколько видов гидроидных семейства Zancleidae имеют симбиотические отношения с мшанками, некоторые из которых полезны для гидроидов, а другие являются паразитическими . В форме некоторых гидроидов появляются модификации, например, щупальца меньшего размера или корка корней мшанок. [117] Мшанка Alcyonidium nodosum защищает трубача Burnupena papyracea от хищничества мощного и прожорливого омара Jasus lalandii . Хотя панцири трубачей, покрытые коркой мшанок, прочнее, чем те, у которых нет такого усиления, химическая защита, создаваемая мшанками, вероятно, является более важным сдерживающим фактором. [118]

В Банк д'Арген на шельфе Мавритании вид Acanthodesia commensale , который обычно растет прикрепленным к гравию и твердому грунту, сформировал факультативные симбиотические отношения с раками-отшельниками вида Pseudopagurus cf. granulimanus, что приводит к образованию структур размером с яйцо, известных как бриолиты. [119] Зарождаясь на пустой раковине брюхоногих моллюсков, колонии мшанок образуют многоламеллярные скелетные корки, которые образуют сферические корки и расширяют жилую камеру рака-отшельника за счет спирального трубчатого роста.
Некоторые виды филактолематов являются промежуточными хозяевами для группы миксозоа , которые, как также обнаружено, вызывают пролиферативное заболевание почек , которое часто приводит к летальному исходу у лососевых рыб. [120] и привело к значительному сокращению популяций диких рыб в Европе и Северной Америке. [66]
Membranipora membranacea , чьи колонии питаются и растут исключительно быстро в широком диапазоне скоростей течения, впервые была замечена в заливе Мэн в 1987 году и быстро стала самым многочисленным организмом, живущим на водорослях . [87] Это вторжение сократило популяцию водорослей, сломав их листья. [18] так что ее место в качестве доминирующей «растительности» на некоторых участках занял другой вселенец — крупная водоросль Codium хрупкая tomentosoides . [87] Эти изменения сократили площадь среды обитания местных рыб и беспозвоночных. M. membranacea также проникла на северо-западное побережье США. [18] Несколько пресноводных видов также были обнаружены за тысячи километров от их естественного ареала. Некоторые из них могли быть перенесены естественным путем в виде статобластов. Другие, скорее всего, были распространены людьми, например, с привезенными водными растениями или безбилетными пассажирами на кораблях. [96]
Взаимодействие с людьми
[ редактировать ]Рыбные фермы и инкубаторы потеряли поголовье из-за пролиферативного заболевания почек , вызванного одним или несколькими миксозойными организмами , которые используют мшанок в качестве альтернативных хозяев. [120]
Некоторым рыбакам в Северном море пришлось искать другую работу из-за формы экземы (кожного заболевания), известной как « зуд Доггер-Бэнка ». [99] вызвано контактом с мшанками, прилипшими к сетям и ловушкам для омаров. [121]
Морские мшанки часто являются причиной биообрастания корпусов кораблей, доков и пристаней, а также морских сооружений. Они являются одними из первых колонизаторов новых или недавно очищенных построек. [111] Пресноводные виды время от времени попадают в водопроводные трубы, оборудование для очистки питьевой воды, очистные сооружения и охлаждающие трубы электростанций. [66] [122]
Группа химических веществ, называемых бриостатинами, может быть извлечена из морской мшанки Bugula neritina . В 2001 году фармацевтическая компания GPC Biotech лицензировала бриостатин 1 из Университета штата Аризона для коммерческой разработки в качестве средства лечения рака. GPC Biotech прекратила разработку в 2003 году, заявив, что бриостатин 1 показал низкую эффективность и некоторые токсичные побочные эффекты. [123] В январе 2008 года клиническое исследование США было отправлено в Национальные институты здравоохранения для оценки безопасности и эффективности Бриостатина 1 при лечении болезни Альцгеймера . Однако к концу декабря 2008 года, когда исследование планировалось завершить, ни один из участников не был набран. [124] Более поздние исследования показывают, что он оказывает положительное влияние на когнитивные функции у пациентов с болезнью Альцгеймера с небольшим количеством побочных эффектов. [125] Для извлечения необходимо переработать около 1000 кг (2200 фунтов) мшанок. 1 грамм ( 1/32 . эффективны унции) бриостатина. В результате были разработаны синтетические эквиваленты, которые проще производить и, очевидно, не менее [126]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Тейлор, PD; Бернинг, Б.; Уилсон, Массачусетс (ноябрь 2013 г.). «Переосмысление кембрийской мшанки Пивакии как восьмикоралла» . Журнал палеонтологии . 87 (6): 984–990. Бибкод : 2013JPal...87..984T . дои : 10.1666/13-029 . S2CID 129113026 . Архивировано из оригинала 7 июня 2019 года . Проверено 20 апреля 2018 г.
- ^ Ма, Джунье; Тейлор, Пол Д.; Ся, Фэншэн; Чжан, Ренбин (сентябрь 2015 г.). «Самая старая известная мшанка: Prophyllodictya (Cryptostomata) из нижнего тремадока (нижнего ордовика) Люцзячана, юго-запад Хубэй, центральный Китай» . Палеонтология . 58 (5): 925–934. Бибкод : 2015Palgy..58..925M . дои : 10.1111/пала.12189 . S2CID 130040324 .
- ^ Перейти обратно: а б Ян, Цзе; Лан, Тиан; Чжан, Си-гуан; Смит, Мартин Р. (8 марта 2023 г.). «Протомелиссия — это ранняя дазикладная водоросль, а не кембрийская мшанка» . Природа . 615 (7952): 468–471. Бибкод : 2023Natur.615..468Y . дои : 10.1038/s41586-023-05775-5 . ПМИД 36890226 . S2CID 257425218 .
- ^ Эрнст, А. (2007). «Цистопоратный вид мшанок из Цехштейна (поздняя пермь)». Палеонтологический журнал . 81 (2): 113–117. Бибкод : 2007PalZ...81..113E . дои : 10.1007/BF02988385 . S2CID 129637643 .
- ^ Перейти обратно: а б с д Фукс, Дж.; Обст, М.; Сундберг, П. (июль 2009 г.). «Первая комплексная молекулярная филогения мшанок (Ectoprocta) на основе комбинированного анализа ядерных и митохондриальных генов». Молекулярная филогенетика и эволюция . 52 (1): 225–233. Бибкод : 2009МОЛПЭ..52..225Ф . дои : 10.1016/j.ympev.2009.01.021 . ПМИД 19475710 .
- ^ Бруска, Ричард К. (1980) [1973]. «21: Лофофоратный тип» . Обыкновенные приливные беспозвоночные Калифорнийского залива (переработанное и дополненное, 2-е изд.). Тусон, Аризона: Издательство Университета Аризоны. ISBN 9780816506828 . OCLC 1029265317 .
- ^ Бок, П.; Гордон, ДП (август 2013 г.). «Филюм мшанки Эренберга, 1831 » . Зоотакса . 3703 (1): 67–74. дои : 10.11646/zootaxa.3703.1.14 .
- ^ Шваха, Томас Ф.; Островский, Андрей Н.; Ваннингер, Андреас (июнь 2020 г.). «Ключевые новинки в эволюции водного колониального типа Bryozoa: данные морфологии мягких тел» . Биологические обзоры Кембриджского философского общества . 95 (3): 696–729. дои : 10.1111/brv.12583 . ПМЦ 7317743 . ПМИД 32032476 .
- ^ Шваха, Томас; Бернхард, Джоан М.; Эджкомб, Вирджиния П.; Тодаро, М. Антонио (1 августа 2019 г.). «Aethozooides uraniae, новый глубоководный род и вид одиночных мшанок из Средиземного моря, с пересмотренной версией Aethozoidae» . Морское биоразнообразие . 49 (4): 1843–1856. Бибкод : 2019MarBd..49.1843S . дои : 10.1007/s12526-019-00948-w . hdl : 11380/1174838 .
- ^ Томпсон, Джон В. (1830). «Мемуары V: На Полизоа обнаружено новое животное как обитатель некоторых зоофитов» . Зоологические исследования и иллюстрации; или Естественная история неописуемых или недостаточно известных животных . Корк, IE: Король и Райдингс. стр. 89–102.
- ^ Эренберг, Кристиан Г. (1831). «Зоологическая часть. Беспозвоночные животные, кроме насекомых [Invertebrata, кроме Insecta]» . Физические символы или обозначения и описания новых или менее известных природных тел (на латыни). Из академической мастерской в Берлине. стр. 1–126.
- ^ Стеббинг, TRR (1911). «Термины Polyzoa и Bryozoa» . Труды Лондонского Линнеевского общества . Сессия 123 (Симпозиум): 61–72.
- ^ Перейти обратно: а б Мьюир-Вуд, HM (1955). История классификации типа Brachiopoda . Лондон: Британский музей (естественная история). п. 13.
- ^ Перейти обратно: а б Вуд, Т.С. (май 2006 г.). «Сильное хищничество пресноводных мшанок золотой яблочной улитки Pomacea canaliculata » (PDF) . Журнал естествознания Университета Чулалонгкорн . 6 (1): 31–36. Архивировано из оригинала (PDF) 6 октября 2011 года . Проверено 18 августа 2009 г.
- ^ Лауэр, Антье (2016). «Watersipora subtorquata и возможная роль связанных с ней микробов: попытка объяснить исключительный успех инвазии этого морского вида мшанок». Механистические преимущества микробных симбионтов . Достижения экологической микробиологии. Том. 2. Чам: Международное издательство Springer. стр. 239–268. дои : 10.1007/978-3-319-28068-4_9 . ISBN 978-3-319-28066-0 .
Иногда воздействие беспозвоночных захватчиков может быть существенным; такое было зарегистрировано для Membranipora membranacea, быстрорастущего инвазивного вида мшанок, который сократил леса водорослей у северо-восточного и северо-западного побережья США до такой степени, что это повлияло на местные популяции рыб и беспозвоночных.
- ^ «Знакомство с мшанками» . Беркли, Калифорния: Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 8 декабря 2019 года . Проверено 8 декабря 2019 г.
- ^ Чжан, Чжилян; Чжан, Чжифэй; Ма, Дж.; Тейлор, PD; Строц, LC; Жаке, СМ; Форест-Плейс, CB; Чен, Ф.; Хан, Дж.; Брок, Джорджия (2021). «Ископаемые свидетельства раскрывают раннекембрийское происхождение мшанок» . Природа 599 (7884): 251–255. Бибкод : 2021Nature.599..251Z . дои : 10.1038/s41586-021-04033-w . ПМЦ 8580826 . ПМИД 34707285 . S2CID 240073948 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Лофопората» . Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 829–845 . ISBN 978-0-03-025982-1 .
- ^ Перейти обратно: а б с Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Лофопората» . Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 817–821 . ISBN 978-0-03-025982-1 .
- ^ Перейти обратно: а б с Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Лофопората» . Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 821–829 . ISBN 978-0-03-025982-1 .
- ^ Перейти обратно: а б с д и ж г час я дж Рич, TH; Фентон, Массачусетс; Фентон, CL (1997). « Моховые животные, или мшанки» . Книга окаменелостей . Дуврские публикации. стр. 142–152 . ISBN 978-0-486-29371-4 . Проверено 7 августа 2009 г.
- ^ Перейти обратно: а б Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Камптозоа и циклиофора» . Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 808–812 . ISBN 978-0-03-025982-1 .
- ^ Перейти обратно: а б Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Лофопората» . Зоология беспозвоночных (7-е изд.). Брукс / Коул. п. 817 . ISBN 978-0-03-025982-1 .
- ^ Перейти обратно: а б с д и ж г час я Нильсен, К. (июль 2002 г.). «Филогенетическое положение Entoprocta, Ectoprocta, Phoronida и Brachiopoda» . Интегративная и сравнительная биология . 42 (3): 685–691. дои : 10.1093/icb/42.3.685 . ПМИД 21708765 .
- ^ Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Аннелида» . Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 414–420 . ISBN 978-0-03-025982-1 .
- ^ Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 284–291 . ISBN 978-0-03-025982-1 .
- ^ Жир, О. (2009). «Тентакулата» . Мейобентология (2-е изд.). Издательство Спрингер. п. 227. ИСБН 978-3-540-68657-6 . Архивировано из оригинала 8 марта 2023 года . Проверено 7 июля 2009 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Доэрти, Пи Джей (2001). «Лофофораты». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных (2-е изд.). Издательство Оксфордского университета. стр. 363–373. ISBN 978-0-19-551368-4 .
- ^ Литтл, В.; Фаулер, Х.В.; Коулсон, Дж. и Лук, Коннектикут (1964). «Зооид». Краткий Оксфордский словарь английского языка . Издательство Оксфордского университета. ISBN 978-0-19-860613-0 .
- ^ Мшанки и палеоэкологическая интерпретация
- ^ Перейти обратно: а б с д и Нильсен, К. (2001). «Мшанки (Ectoprocta: моховые животные)». Энциклопедия наук о жизни . John Wiley & Sons, Ltd. doi : 10.1038/npg.els.0001613 . ISBN 978-0470016176 .
- ^ Перейти обратно: а б с д и ж г час МакКинни, ФК; Джексон, JBC (1991). «Мшанки как модульные машины» . Эволюция мшанок . Издательство Чикагского университета. стр. 1–13. ISBN 978-0-226-56047-2 . Архивировано из оригинала 8 марта 2023 года . Проверено 29 июля 2009 г.
- ^ Риисгорд, Хьюстон; Нильсен, К; Ларсен, PS (2000). «Последующий сбор в реснитчатых суспензионных питателях: принцип догона» (PDF) . Серия «Прогресс в области морской экологии» . 207 : 33–51. Бибкод : 2000MEPS..207...33R . дои : 10.3354/meps207033 . Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 12 сентября 2009 г.
- ^ Перейти обратно: а б «фуникулер» . Случайный словарь дома . Случайный дом. Архивировано из оригинала 13 мая 2010 года . Проверено 2 августа 2009 г.
- ^ Хейворд, ПиДжей (1985). «Систематическая часть» . Ктеностомные мшанки . Конспекты британской фауны. Линнеевское общество Лондона. стр. 106–107. ISBN 978-90-04-07583-2 . Архивировано из оригинала 8 марта 2023 года . Проверено 2 августа 2009 г.
- ^ Жир, О. (2009). «Тентакулата» . Мейобентология (2-е изд.). Спрингер-Верлаг. п. 227. ИСБН 978-3-540-68657-6 . Архивировано из оригинала 8 марта 2023 года . Проверено 2 августа 2009 г.
- ^ Лидделл, Х.Г.; Скотт Р. (1940). "кенос " Греко-английский лексикон . Кларендон Пресс. ISBN 978-0-19-864226-8 . Архивировано из оригинала 8 марта 2023 года . Проверено 1 августа 2009 г.
- ^ Тейлор 2020 , стр. 72–73.
- ^ Тейлор 2020 , с. 74.
- ^ Тейлор 2020 , с. 59.
- ^ Тейлор 2020 , с. 60.
- ^ Тейлор 2020 , с. 65.
- ^ Тейлор 2020 , с. 75.
- ^ Филиал, МЛ; Гриффитс, CI; Бекли, Л.Е. (2007). «Мшанки: моховые или кружевные животные» . Два океана – Путеводитель по морской жизни Южной Африки . Струйк. стр. 104–110. ISBN 978-1-77007-633-4 . Проверено 2 августа 2009 г.
- ^ Литтл, В.; Фаулер, Х.В.; Коулсон, Дж. и Лук, Коннектикут (1959). «Мшанка». Краткий Оксфордский словарь английского языка . Оксфордский университет. ISBN 978-0-19-860613-0 .
- ^ Экман, Дж. Э.; Окамура, Б. (декабрь 1998 г.). «Модель захвата частиц мшанками в турбулентном потоке: значение формы колонии». Американский натуралист . 152 (6): 861–880. дои : 10.1086/286214 . ПМИД 18811433 . S2CID 5535013 .
- ^ Фогель, С. (1996). «Жизнь в градиентах скорости» . Жизнь в движущихся жидкостях (2-е изд.). Издательство Принстонского университета. п. 191. ИСБН 978-0-691-02616-9 . Архивировано из оригинала 8 марта 2023 года . Проверено 5 августа 2009 г.
- ^ фон Дассов, М. (1 августа 2006 г.). «Функционально-зависимое развитие колониального животного» . Биологический бюллетень . 211 (1): 76–82. дои : 10.2307/4134580 . ISSN 0006-3185 . JSTOR 4134580 . ПМИД 16946244 . Архивировано из оригинала 6 июля 2009 года . Проверено 5 августа 2009 г.
- ^ Тейлор, Пол Д.; Джеймс, Ноэль П. (август 2013 г.). «Вековые изменения колониальных форм и мшанок карбонатных отложений на протяжении геологической истории» . Седиментология . 60 (5): 1184–1212. дои : 10.1111/сед.12032 . S2CID 128939236 .
- ^ Перейти обратно: а б Битти, Дж.А.; Блэквелдер (1974). «Названия типов беспозвоночных». Систематическая зоология . 23 (4): 545–547. дои : 10.2307/2412472 . JSTOR 2412472 .
- ^ Перейти обратно: а б Майр, Э. (июнь 1968 г.). «Мшанка против Эктопрокты». Систематическая зоология . 17 (2): 213–216. дои : 10.2307/2412368 . JSTOR 2412368 .
- ^ Литтл, В.; Фаулер, Х.В.; Коулсон, Дж. и Лук, Коннектикут (1964). «Экто-». Краткий Оксфордский словарь английского языка . Издательство Оксфордского университета. ISBN 978-0-19-860613-0 .
- ^ Литтл, В.; Фаулер, Х.В.; Коулсон, Дж. и Лук, Коннектикут (1964). «Энто-». Краткий Оксфордский словарь английского языка . Издательство Оксфордского университета. ISBN 978-0-19-860613-0 .
- ^ Перейти обратно: а б с д и ж г час Галаныч, К.М. (2004). «Новый взгляд на филогению животных» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 35 : 229–256. doi : 10.1146/annurev.ecolsys.35.112202.130124 . Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 26 августа 2016 г.
- ^ Перейти обратно: а б с д и ж Хаусдорф, Б.; Хельмкампф, М.; Мейер, А.; Витек, А.; Херлин, Х.; Бруххаус, И.; Ханкельн, Т.; Страк, TH; Либ, Б. (2007). «Спиральная филогеномика поддерживает возрождение мшанок, состоящих из Ectoprocta и Entoprocta» . Молекулярная биология и эволюция . 24 (12): 2723–2729. дои : 10.1093/molbev/msm214 . ПМИД 17921486 .
- ^ Каффи, Р.Дж. (1969). «Мшанка против эктопрокты – необходимость точности». Систематическая зоология . 18 (2): 250–251. дои : 10.2307/2412617 . JSTOR 2412617 .
- ^ Гизелин, МТ (1977). «Об изменении названий высших таксонов». Систематическая зоология . 26 (3): 346–349. дои : 10.2307/2412681 . JSTOR 2412681 .
- ^ Ёкобори, С.; Исето, Т; Асакава, С; Сасаки, Т; Симидзу, Н; Ямагиши, А; Осима, Т; Хиросе, Э. (май 2008 г.). «Полные нуклеотидные последовательности митохондриальных геномов двух одиночных энтопроктов, Loxocorone allax и Loxosomella aloxiata : значение для филогении лофотрохозой». Молекулярная филогенетика и эволюция . 47 (2): 612–628. Бибкод : 2008МОЛПЭ..47..612Г . дои : 10.1016/j.ympev.2008.02.013 . ПМИД 18374604 .
- ^ Рейнольдс, КТ (2000). «Таксономически важные особенности поверхности флоатобластов Plumatella (Bryozoa)». Микроскопия и микроанализ . 6 (3): 202–210. Бибкод : 2000MiMic...6..202R . дои : 10.1017/S1431927600000349 . ПМИД 10790488 . Текст начинается со слов «Тип Ectoprocta (Bryozoa)…».
- ^ Трамбл, Вт; Браун, Л. (2002). «Мшанка» . Краткий Оксфордский словарь английского языка . Издательство Оксфордского университета. ISBN 978-0-19-860457-0 .
- ^ Тейлор, Затон, 2008 г.
- ^ Тейлор, Пол Д. (октябрь 2008 г.). «Таксономия родов мшанок Oncousoecia, Microeciella и Eurystrotos». Журнал естественной истории . 42 (39–40): 2557–2574. дои : 10.1080/00222930802277640 . S2CID 84315311 .
- ^ Чепмен, AD (2006). Численность живых видов в Австралии и мире (PDF) . Департамент окружающей среды и наследия правительства Австралии. п. 34. ISBN 978-0-642-56849-6 . Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 7 августа 2009 г.
- ^ Руперт, Э.Э.; Фокс, Р.С. и Барнс, Р.Д. (2004). «Знакомство с беспозвоночными» . Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 2–9 . ISBN 978-0-03-025982-1 .
- ^ «Страница стандартного отчета ITIS: Филактолематы» . Интегрированная система таксономической информации. Архивировано из оригинала 18 января 2018 года . Проверено 12 августа 2009 г.
- ^ Перейти обратно: а б с д и ж г час Массард, Дж.А.; Геймер, Габи (2008). «Глобальное разнообразие мшанок (Bryozoa или Ectoprocta) в пресной воде». Гидробиология . 595 : 93–99. дои : 10.1007/s10750-007-9007-3 . S2CID 13057599 .
- ^ Перейти обратно: а б Фиш, Джей Ди; Фиш, С. (1996). «Мшанка» . Путеводитель по морю для школьников (2-е изд.). Кембридж: Издательство Кембриджского университета. стр. 418–419 . ISBN 978-0-521-46819-0 .
- ^ Яблонски, Д.; Лидгард, С.; Тейлор, П.Д. (1997). «Сравнительная экология излучений мшанок: происхождение новинок у круглоротых и хейлостом». ПАЛЕОС . 12 (6): 505–523. Бибкод : 1997Палай..12..505J . дои : 10.2307/3515408 . JSTOR 3515408 .
- ^ Перейти обратно: а б Хейворд, ПиДжей; Райланд, Дж. С. (1985). «Определитель высших таксонов морских мшанок» . Круглоротые мшанки . Линнеевское общество Лондона. п. 7. ISBN 978-90-04-07697-6 . Архивировано из оригинала 8 марта 2023 года . Проверено 9 августа 2009 г.
- ^ Перейти обратно: а б МакКинни; Фрэнк К; Джереми. «Эволюция мшанок». Бостон: Анвин и Хайман, 1989.
- ^ Перейти обратно: а б Эрнст, Эндрю (2020). «2- Летопись окаменелостей и эволюция мшанок». Тип мшанки . стр. 11–56.
- ^ Джексон, Патрик Н. Вайз; Ки, Маркус М. Ки младший (2019). «Распределение эпизоя и эндоскелетоза в вновь собранных разветвленных стенолаематных мшанках зоариях из верхнего ордовика (катиан) региона арки Цинциннати, США». Австралазийские палеонтологические мемуары . 52 : 169–178.
- ^ Торсвик, TH; Райан, Пол Д.; Тренч, Аллан; Харпер, Дэвид А.Т. (январь 1991 г.). «Кембрийско-ордовикская палеогеография Балтики». Геология . 19 (1): 7–10. Бибкод : 1991Geo....19....7T . doi : 10.1130/0091-7613(1991)019<0007:COPOB>2.3.CO;2 .
- ^ Перейти обратно: а б с д и Дьюэл, РА; Уинстон, Дж. Э.; МакКинни, Ф.Дж. (2002). «Деконструкция бёзоанов: происхождение и последствия уникального строения тела» . В Уайз-Джексдоне, штат Пенсильвания; Баттлер, CE; Спенсер Джонс, Мэн (ред.). Исследования мшанок 2001: материалы Двенадцатой Международной конференции по мшанке . М. Е. Лиссе: Светс и Цайтлингер. стр. 93–96. ISBN 978-90-5809-388-2 . Архивировано из оригинала 8 марта 2023 года . Проверено 13 августа 2009 г.
- ^ Олемпска, Э. (2012). «Исключительная сохранность мягких тканей у сверлящих гребневиков мшанок и связанных с ними «грибных» расточек из раннего девона Подолья, Украина» . Acta Palaeontologica Polonica . 57 (4): 925–940. дои : 10.4202/app.2011.0200 .
- ^ МакКинни, ФК (1994). «Сто миллионов лет конкурентных взаимодействий между кладами мшанок: асимметричные, но не обостряющиеся». Биологический журнал Линнеевского общества . 56 (3): 465–481. дои : 10.1111/j.1095-8312.1995.tb01105.x .
- ^ Вуд, Р. (1999). Эволюция рифов . Издательство Оксфордского университета. стр. 235–237. ISBN 978-0-19-857784-3 . Архивировано из оригинала 8 марта 2023 года . Проверено 11 августа 2009 г.
- ^ Перейти обратно: а б с Вуд, Т.С.; Лор М. (2005). «Высшая филогения филактолематных мшанок, выведенная на основе последовательностей рибосомальной ДНК 18S» (PDF) . В Мояно, Гавайи; Кансино, Дж. М.; Вайз-Джексон, ПН (ред.). Исследования мшанок 2004: Труды 13-й Международной ассоциации мшанок . Лондон: Группа Тейлора и Фрэнсиса. стр. 361–367. Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 24 августа 2009 г.
- ^ Поховский, Р.А. (1978). «Сверлые гребневики мшанки: таксономия и палеобиология на основе полостей в известковых субстратах». Бюллетени американской палеонтологии . 73 : 192с.
- ^ Гэри-младший; Шмидт-Раэса, Андреас (1998). «Основная роль «второстепенных» типов в молекулярных исследованиях эволюции животных» . Американский зоолог . 38 (6): 907–917. дои : 10.1093/icb/38.6.907 .
- ^ Перейти обратно: а б с Нильсен, К. (2001). «Тип Эктопрокта» . Эволюция животных: взаимоотношения живых типов (2-е изд.). Издательство Оксфордского университета. стр. 244–264. ISBN 978-0-19-850681-2 . Архивировано из оригинала 8 марта 2023 года . Проверено 14 августа 2009 г.
- ^ «Знакомство с полухордовыми» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 1 февраля 2019 года . Проверено 22 сентября 2008 г.
- ^ Перейти обратно: а б с Хельмкампф, М.; Бруххаус, Ирис; Хаусдорф, Бернхард (2008). «Филогеномный анализ лофофоратов (брахиопод, форонид и мшанок) подтверждает концепцию Lophotrochozoa» . Труды Королевского общества B: Биологические науки . 275 (1645): 1927–1933. дои : 10.1098/rspb.2008.0372 . ПМК 2593926 . ПМИД 18495619 .
- ^ Нильсен, К.; Ворсаа, К. (сентябрь 2010 г.). «Строение и распространение личинок cyphonautes (Bryozoa, Ectoprocta)». Журнал морфологии . 271 (9): 1094–1109. дои : 10.1002/jmor.10856 . ПМИД 20730922 . S2CID 11453241 .
- ^ Шен, X.; Тиан, М.; Мэн, X.; Лю, Х.; Ченг, Х.; Чжу, К.; Чжао, Ф. (сентябрь 2012 г.). «Полный митохондриальный геном Membranipora grandicella (Bryozoa: Cheilostomatida), определенный с помощью секвенирования следующего поколения: первый представитель подотряда Malacostegina». Сравнительная биохимия и физиология. Часть D: Геномика и протеомика . 7 (3): 248–253. дои : 10.1016/j.cbd.2012.03.003 . ПМИД 22503287 .
- ^ Перейти обратно: а б с Каллаган, ТП; Р., Карлсон (июнь 2002 г.). «Летний покой как убежище от смертности пресноводных мшанок Plumatella emarginata ». Экология . 132 (1): 51–59. Бибкод : 2002Oecol.132...51C . дои : 10.1007/s00442-002-0946-0 . ПМИД 28547286 . S2CID 19925846 .
- ^ Перейти обратно: а б с Пратт, MC (2008). «Жизнь там, где есть правильный поток: как поток влияет на питание мшанок» (PDF) . Интегративная и сравнительная биология . 48 (6): 808–822. дои : 10.1093/icb/icn052 . ПМИД 21669834 .
- ^ Райланд, Дж. С. (1967). «Дыхание у многозоа (эктопрокта)». Природа . 216 (5119): 1040–1041. Бибкод : 1967Natur.216.1040R . дои : 10.1038/2161040b0 . S2CID 4207120 .
- ^ Неклюдова, ЮА; Шваха, ТФ; Котенко, О.Н.; Грубер, Д.; Сайран, Н.; Островский, АН (2021). «Три в одном: Эволюция живорождения, ценоцитарной плаценты и полиэмбрионии у круглоротых мшанок» . BMC Экология и эволюция . 21 (1): 54. дои : 10.1186/s12862-021-01775-z . ПМК 8042935 . ПМИД 33845757 .
- ^ Моосбругер, М.; Шваха, Т.; Уолцл, МГ; Обст, М.; Островский, АН (2012). «Плацентарный аналог и закономерности полового размножения у хейлостомных мшанок Bicellariella ciliata (Gymnolaemata)» . Границы в зоологии . 9 (1): 29. дои : 10.1186/1742-9994-9-29 . ПМЦ 3489856 . ПМИД 23098166 .
- ^ Котенко, О.Н.; Островский, А.Н. (2024). «Разгадка эволюции личинок мшанок» . Палеонтологический журнал . 57 (11): 1306. Бибкод : 2024PalJ...57.1306K . дои : 10.1134/S0031030123110072 .
- ^ Декер, Себастьян; Ваннингер, Андреас; Шваха, Томас (2020). «Морфология и жизненный цикл эпифитной мшанки ферузеллид гребневиков из Средиземного моря» . Разнообразие и эволюция организмов . 20 (3): 417–437. дои : 10.1007/s13127-020-00443-2 .
- ^ Шваха, Томас (23 ноября 2020 г.). Тип мшанки . Вальтер де Грюйтер ГмбХ & Ко КГ. ISBN 978-3-11-058631-2 .
- ^ Шваха, Томас; Зеппилли, Даниэла; Гонсалес-Касаррубиос, Альберт; Сепеда, Диего (2024). «Первая глубоководная мшанка гребневиков из Индийского океана: Aethozoon flavum sp. nov» . Морское биоразнообразие . 54 (2): 19. Бибкод : 2024Март..54... 19С. дои : 10.1007/ s12526-024-01409-9
- ^ Стратманн, Р.Р. (март 2006 г.). «Универсальное поведение ресничек при захвате частиц личинкой мшанок cyphonautes». Акта Зоология . 87 (1): 83–89. дои : 10.1111/j.1463-6395.2006.00224.x .
- ^ Перейти обратно: а б Вуд, Т.С.; Окамура, Бет (декабрь 1998 г.). « Asajirella gelatinosa в Панаме: расширение ареала мшанок в Западном полушарии». Гидробиология . 390 (1–3): 19–23. дои : 10.1023/A:1003502814572 . S2CID 1525771 .
- ^ О'Ди, Аарон; Джексон, Джереми, Британская Колумбия; Тейлор, Пол Д.; Родригес, Феликс (2008). «Способы размножения современных и ископаемых мшанок купуладриид» . Палеонтология . 51 (4): 847–864. Бибкод : 2008Palgy..51..847O . дои : 10.1111/j.1475-4983.2008.00790.x . S2CID 41016220 .
- ^ Эмилиани, К. (1992). «Палеозой» . Планета Земля: космология, геология, эволюция жизни и окружающей среды . Издательство Кембриджского университета. стр. 488–490 . ISBN 978-0-19-503652-7 . Проверено 11 августа 2009 г.
- ^ Перейти обратно: а б с д Джонс, RW (2006). «Основные ископаемые группы» . Прикладная палеонтология . Издательство Кембриджского университета. п. 116. ИСБН 978-0-521-84199-3 . Архивировано из оригинала 8 марта 2023 года . Проверено 11 августа 2009 г.
- ^ Куклинский, П.; Бадер, Беате (2007). «Сравнение комплексов мшанок из двух контрастирующих регионов арктического шельфа». Устьевые, прибрежные и шельфовые науки . 73 (3–4): 835–843. Бибкод : 2007ECSS...73..835K . дои : 10.1016/j.ecss.2007.03.024 .
- ^ Бруска, Р; Бруска, Г. «Беспозвоночные (2-е издание)». Сандерленд, Массачусетс: Sinauer Associates.
- ^ Тейлор 2020 , с. 159.
- ^ Тейлор 2020 , с. 164.
- ^ Тейлор 2020 , с. 162–163.
- ^ Тейлор 2020 , с. 112–113.
- ^ Перейти обратно: а б Тейлор 2020 , с. 79.
- ^ Пек, Л.С.; Хейворд, ПиДжей; Спенсер-Джонс, Мэн (октябрь 1995 г.). «Пелагическая мшанка из Антарктиды» . Морская биология . 123 (4): 757–762. Бибкод : 1995MarBi.123..757P . дои : 10.1007/BF00349118 . S2CID 83529565 . Архивировано из оригинала 3 августа 2017 года . Проверено 28 августа 2017 г.
- ^ Тейлор 2020 , с. 114.
- ^ Мэтт МакГрат (16 июня 2014 г.). « По мере того, как нагреваются берега Антарктики, «сорняки» процветают» . Новости Би-би-си . Архивировано из оригинала 17 июня 2014 года . Проверено 16 июня 2014 г.
- ^ Рамель, Гордон (5 марта 2020 г.). «Мшанки: очаровательные колонии типа Ectoprocta» . Земная жизнь . Архивировано из оригинала 27 марта 2022 года . Проверено 19 мая 2022 г.
- ^ Перейти обратно: а б Маргулис, Л .; Шварц К.В. (1998). «Мшанка» . Пять королевств: иллюстрированный путеводитель по типам жизни на Земле . Эльзевир. п. 335 . ISBN 978-0-7167-3027-9 . Проверено 20 августа 2009 г.
- ^ Айенгар, Э.В.; Харвелл, компакт-диск (2002). «Специфика сигналов, вызывающих защитные шипы у мшанок Membranipora membranacea» . Серия «Прогресс в области морской экологии» . 225 : 205–218. Бибкод : 2002MEPS..225..205I . дои : 10.3354/meps225205 . Архивировано из оригинала 26 января 2012 года . Проверено 18 августа 2009 г.
- ^ Хейворд, ПиДжей; Райланд, Дж. С. (1985). «Хищники» . Круглоротые мшанки: определители и примечания для определения вида . Архив Брилла. п. 27. ISBN 978-90-04-07697-6 . Архивировано из оригинала 8 марта 2023 года . Проверено 18 августа 2009 г.
- ^ Дэй, RW; Осман, RW (январь 1981 г.). «Хищничество Patiria miniata (Asteroidea) на мшанках». Экология . 51 (3): 300–309. Бибкод : 1981Oecol..51..300D . дои : 10.1007/BF00540898 . ПМИД 28310012 . S2CID 19976956 .
- ^ МакКинни, ФК; Тейлор, PD; Лидгард, С. (2003). «Хищничество мшанок и его отражение в летописи окаменелостей» . В Келли, штат Пенсильвания; Ковалевски, М.; Хансен, Т.А. (ред.). Взаимодействие хищник-жертва в летописи окаменелостей . Спрингер. стр. 239–246. ISBN 978-0-306-47489-7 . Архивировано из оригинала 8 марта 2023 года . Проверено 18 августа 2009 г.
- ^ Вуд, Т.С. (октябрь 2006 г.). «Пресноводные мшанки Таиланда (Ectoprocta и Entoprocta)» (PDF) . Журнал естественной истории Университета Чулалонгкорн . 6 (2): 83–119. Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 24 августа 2009 г.
- ^ Пьюс, С. (2007). «Симбиотические отношения гидроидов и мшанок» . Конгресс Международного Общества Симбиоза № 5 . 44 (1–3): 137–143. Архивировано из оригинала 26 января 2012 года . Проверено 18 августа 2009 г.
- ^ Грей, Калифорния; Маккуэйд, компакт-диск; Дэвис-Коулман, штат Монтана (декабрь 2005 г.). «Симбиотическая мшанка, покрытая коркой панциря, обеспечивает сублиторальных трубачей химической защитой от каменных омаров». Африканский журнал морской науки . 27 (3): 549–556. Бибкод : 2005AfJMS..27..549G . дои : 10.2989/18142320509504115 . S2CID 84531235 .
- ^ Кличпера, Андре; Тейлор, Пол Д.; Вестфаль, Хильдегард (30 июля 2013 г.). «Бриолиты, построенные мшанками в симбиотических ассоциациях с крабами-отшельниками в тропической гетерозойной карбонатной системе, Гольф-д'Арген, Мавритания» . Морское биоразнообразие . 43 (4): 429–444. Бибкод : 2013МарБд..43..429К . дои : 10.1007/s12526-013-0173-4 . S2CID 15841444 . Архивировано из оригинала 11 января 2020 года . Проверено 19 августа 2019 г.
- ^ Перейти обратно: а б Андерсон, К.; Каннинг, ЕС; Окамура, Б. (1999). «Молекулярные данные свидетельствуют о том, что мшанки являются хозяевами PKX (тип Myxozoa) и идентифицируют кладу паразитов мшанок внутри Myxozoa» . Паразитология . 119 (6): 555–561. дои : 10.1017/S003118209900520X . ПМИД 10633916 . S2CID 2851575 . Архивировано из оригинала 26 января 2012 года . Проверено 18 августа 2009 г.
- ^ Клин, Б. (2008). «Профессиональная светочувствительная экзема рыбаков при контакте с мшанками: инвалидизирующий профессиональный дерматоз» (PDF) . Международное морское здравоохранение . 59 (1–4): 1–4. ПМИД 19227737 . Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 18 августа 2009 г.
- ^ Вуд, Т.С.; Марш, Терренс Дж. (февраль 1999 г.). «Биообрастание очистных сооружений пресноводной мшанкой Plumatella vaihiriae ». Исследования воды . 33 (3): 609–614. дои : 10.1016/S0043-1354(98)00274-7 .
- ^ «Бриостатин 1» . 19 июня 2006 года. Архивировано из оригинала 9 мая 2007 года . Проверено 20 августа 2009 г.
- ^ «Исследование безопасности, эффективности, фармакокинетики и фармакодинамики бриостатина 1 у пациентов с болезнью Альцгеймера» . Национальные институты здравоохранения. 19 августа 2009 года. Архивировано из оригинала 13 июня 2011 года . Проверено 20 августа 2009 г.
- ^ Нельсон, Ти Джей; Вс, МК; Лим, К.; Сен, А.; Хан, Т.; Кирила, Ф.В.; Алкон, Д.Л. (2017). «Влияние бриостатина на когнитивные функции и PKCɛ в фазе IIA болезни Альцгеймера и исследованиях расширенного доступа» . Журнал болезни Альцгеймера . 58 (2): 521–535. дои : 10.3233/JAD-170161 . ПМЦ 5438479 . ПМИД 28482641 .
- ^ Вендер, Пенсильвания; Барыза, Дж.Л.; Беннетт, CE; Би, ФК; Бреннер, SE; Кларк, Миссури; Хоран, JC; Кан, К; и др. (20 ноября 2002 г.). «Практический синтез нового и сильнодействующего аналога бриостатина». Журнал Американского химического общества . 124 (46): 13648–13649. дои : 10.1021/ja027509+ . ПМИД 12431074 .
Библиография
[ редактировать ]- Тейлор, Пол Д. (2020). Палеобиология мшанок . Лондон: Музей естественной истории. ISBN 9781118455005 .
Дальнейшее чтение
[ редактировать ]- Холл, СР; Тейлор, PD; Дэвис, ЮАР; Манн, С. (2002). «Электронографические исследования известковых скелетов мшанок». Журнал неорганической биохимии . 88 (3–4): 410–419. дои : 10.1016/S0162-0134(01)00359-2 . ПМИД 11897358 .
- Хейворд, П.Г., Дж.С. Райланд и П.Д. Тейлор (ред.), 1992. Биология и палеобиология мшанок , Олсен и Олсен, Фреденсборг, Дания. [ ISBN отсутствует ]
- Уинстон, Дж. Э. (2010). «Жизнь в колониях: изучение инопланетного образа жизни колониальных организмов» . Интегративная и сравнительная биология . 50 (6): 919–933. дои : 10.1093/icb/icq146 . ПМИД 21714171 .
- Робисон, Р.А. (редактор), 1983. Трактат по палеонтологии беспозвоночных , Часть G, Мшанки (пересмотренный). Геологическое общество Америки и Канзасский университет Press. [ ISBN отсутствует ]
- Шарп, Дж. Х.; Уинсон, МК; Портер, Дж. С. (2007). «Метаболиты мшанок: экологическая перспектива» (PDF) . Отчеты о натуральных продуктах . 24 (4): 659–673. дои : 10.1039/b617546e . HDL : 2160/3792 . ПМИД 17653353 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- Тейлор, П; Уилсон, Массачусетс (2003). «Палеоэкология и эволюция сообществ морского твердого субстрата» (PDF) . Обзоры наук о Земле . 62 (1): 1–103. Бибкод : 2003ESRv...62....1T . дои : 10.1016/S0012-8252(02)00131-9 . Архивировано из оригинала (PDF) 25 марта 2009 года.
- Винн, Олев; Уилсон, Марк А.; Мытус, Мари-Анн; Тоом, Урсула (2014). «Самый ранний мшанок-паразит: средний ордовик (дарривилий) острова Осмуссаар, Эстония» . Палеогеография, Палеоклиматология, Палеоэкология . 414 : 129–132. Бибкод : 2014PPP...414..129В . дои : 10.1016/j.palaeo.2014.08.021 .
- Вуллакотт, Р.М. и Р.Л. Циммер (редакторы), 1977. Биология мшанок , Academic Press, Нью-Йорк. [ ISBN отсутствует ]
Внешние ссылки
[ редактировать ]- Указатель домашней страницы Bryozoa Bryozoa находился в RMIT; сейчас bryozoa.net
- Другие ресурсы WWW по мшанкам
- Международной ассоциации бриозоологии Официальный сайт
- Неогеновые мшанки Британии
- Мшанки Введение
- Тип Ectoprocta (Bryozoa)
- Тип мшанки в Wikispecies
- Мшанки в реке Коннектикут
- Информационный бюллетень о мшанках