Нейронная цепь

Нейронная цепь — это совокупность нейронов , соединенных между собой синапсами для выполнения определенной функции при активации. [ 1 ] Множество нейронных цепей соединяются друг с другом, образуя крупномасштабные мозговые сети . [ 2 ]
Нейронные цепи вдохновили на создание искусственных нейронных сетей , хотя между ними есть существенные различия.
Раннее обучение
[ редактировать ]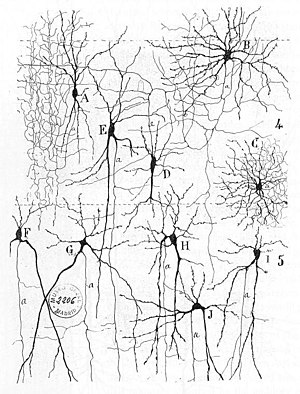
Ранние описания нейронных сетей можно найти в Герберта Спенсера » «Принципах психологии , 3-е издание (1872 г.), Теодора Мейнерта » «Психиатрии (1884 г.), Уильяма Джеймса ( «Принципах психологии» 1890 г.) и Зигмунда Фрейда «Проекте для Научная психология (составлено в 1895 г.). [ 3 ] Первое правило нейронного обучения было описано Хеббом в 1949 году в теории Хебба . Таким образом, хеббианское спаривание пресинаптической и постсинаптической активности может существенно изменить динамические характеристики синаптической связи и, следовательно, либо облегчить, либо ингибировать передачу сигнала . В 1959 году нейробиологи Уоррен Стерджис Маккалок и Уолтер Питтс опубликовали первые работы по обработке нейронных сетей. [ 4 ] Они теоретически показали, что сети искусственных нейронов могут выполнять логические , арифметические и символические функции. упрощенные модели биологических нейронов Были созданы , которые теперь обычно называются перцептронами или искусственными нейронами . Эти простые модели объясняли суммирование нейронов (т.е. потенциалы постсинаптической мембраны суммируются в теле клетки ). Более поздние модели также предусматривали возбуждающую и тормозную синаптическую передачу.
Связи между нейронами
[ редактировать ]
Связи между нейронами в мозге гораздо более сложны, чем связи между искусственными нейронами, используемыми в коннекционистских моделях нейронных вычислений искусственных нейронных сетей . Основным видом связей между нейронами являются синапсы : как химические , так и электрические .
Образование синапсов позволяет нейронам соединяться в миллионы перекрывающихся и взаимосвязанных нейронных цепей. Пресинаптические белки, называемые нейрексинами, играют центральную роль в этом процессе. [ 5 ]
Одним из принципов работы нейронов является нейронное суммирование : потенциалы на постсинаптической мембране суммируются в теле клетки. Если деполяризация нейрона на аксонном холмике превысит пороговое значение, возникнет потенциал действия, который перемещается вниз по аксону к терминальным окончаниям для передачи сигнала другим нейронам. Возбуждающая и тормозная синаптическая передача реализуется преимущественно посредством возбуждающих постсинаптических потенциалов (ВПСП) и тормозных постсинаптических потенциалов (ТПСП).
На электрофизиологическом уровне существуют различные явления, которые изменяют характеристики реакции отдельных синапсов (так называемая синаптическая пластичность ) и отдельных нейронов ( внутренняя пластичность ). Их часто делят на краткосрочную пластичность и долгосрочную пластичность. Долговременная синаптическая пластичность часто считается наиболее вероятным субстратом памяти . Обычно термин « нейропластичность » относится к изменениям в мозге, вызванным деятельностью или опытом.
Связи отображают временные и пространственные характеристики. Временные характеристики относятся к постоянно изменяющейся, зависящей от активности эффективности синаптической передачи, называемой пластичностью, зависящей от времени спайка . В нескольких исследованиях было замечено, что синаптическая эффективность этой передачи может подвергаться кратковременному увеличению (называемому облегчением ) или снижению ( депрессия ) в зависимости от активности пресинаптического нейрона. Индукция долгосрочных изменений синаптической эффективности посредством долгосрочной потенциации (LTP) или депрессии (LTD) сильно зависит от относительного времени возникновения возбуждающего постсинаптического потенциала и постсинаптического потенциала действия. ДП индуцируется серией потенциалов действия, которые вызывают различные биохимические реакции. В конечном итоге реакции вызывают экспрессию новых рецепторов на клеточных мембранах постсинаптических нейронов или повышают эффективность существующих рецепторов за счет фосфорилирования .
Потенциалы обратного распространения действия не могут возникать, поскольку после того, как потенциал действия проходит вниз по заданному сегменту аксона, m-ворота потенциалзависимых натриевых каналов закрываются, таким образом блокируя любое временное открытие h-ворот , вызывающее изменение внутриклеточных ионов натрия ( Na + ) концентрации и предотвращения генерации потенциала действия обратно в тело клетки. Однако в некоторых клетках обратное распространение нейронов действительно происходит посредством дендритного ветвления и может оказывать важное влияние на синаптическую пластичность и вычисления.
Нейрону головного мозга требуется одиночный сигнал нервно-мышечному соединению, чтобы стимулировать сокращение постсинаптической мышечной клетки. Однако в спинном мозге не менее 75 афферентных для возбуждения импульса требуется нейронов. Эта картина еще больше усложняется различиями в постоянной времени между нейронами, поскольку некоторые клетки могут испытывать ВПСП в течение более длительного периода времени, чем другие.
Хотя в синапсах развивающегося мозга особенно часто наблюдается синаптическая депрессия, предполагается, что во взрослом мозге она сменяется облегчением.
Схема
[ редактировать ]
Примером нейронной цепи является трисинаптическая цепь в гиппокампе . Другой — контур Папеса, соединяющий гипоталамус с лимбической долей . имеется несколько нейронных цепей В корково-базальных ганглиях-таламо-кортикальной петле . Эти цепи передают информацию между корой, базальными ганглиями , таламусом и обратно в кору. Самая крупная структура базальных ганглиев — полосатое тело — имеет собственную внутреннюю микросхему. [ 6 ]
Нейронные цепи в спинном мозге , называемые генераторами центральных паттернов, отвечают за управление двигательными инструкциями, участвующими в ритмическом поведении. Ритмическое поведение включает ходьбу, мочеиспускание и эякуляцию . Генераторы центральных паттернов состоят из различных групп спинальных интернейронов . [ 7 ]
Существует четыре основных типа нейронных цепей, которые отвечают за широкий спектр нейронных функций. Этими цепями являются расходящаяся цепь, сходящаяся цепь, реверберационная цепь и параллельная цепь послеразряда. [ 8 ]
В расходящейся цепи один нейрон образует синапсы с несколькими постсинаптическими клетками. Каждый из этих может синапсировать со многими другими, позволяя одному нейрону стимулировать до тысяч клеток. Это иллюстрируется тем, что тысячи мышечных волокон могут быть стимулированы исходным сигналом от одного мотонейрона . [ 8 ]
В сходящейся схеме входные сигналы от многих источников объединяются в один выходной, влияя только на один нейрон или пул нейронов. Примером этого типа схемы является дыхательный центр ствола мозга , который реагирует на ряд входных сигналов из разных источников, выдавая соответствующий паттерн дыхания. [ 8 ]
Реверберирующая схема производит повторяющийся выходной сигнал. В процедуре передачи сигналов от одного нейрона к другому в линейной последовательности один из нейронов может отправить сигнал обратно инициирующему нейрону. Каждый раз, когда срабатывает первый нейрон, другой нейрон, расположенный дальше по последовательности, снова срабатывает, отправляя его обратно к источнику. Это рестимулирует первый нейрон, а также позволяет продолжить путь передачи к его выходу. Результирующий повторяющийся паттерн — это результат, который прекращается только в том случае, если один или несколько синапсов выходят из строя или если тормозящая энергия из другого источника заставляет его остановиться. Этот тип реверберирующей цепи находится в дыхательном центре и посылает сигналы дыхательным мышцам , вызывая вдох. Когда цепь прерывается тормозным сигналом, мышцы расслабляются, вызывая выдох. Этот тип цепи может играть роль в эпилептических припадках . [ 8 ]
В параллельной цепи после разряда нейрон передает сигналы нескольким цепочкам нейронов. Каждая цепочка состоит из разного количества нейронов, но их сигналы сходятся на одном выходном нейроне. Каждый синапс в цепи задерживает сигнал примерно на 0,5 мс, поэтому чем больше синапсов, тем дольше задержка для выходного нейрона. После остановки входа выход будет продолжать работать в течение некоторого времени. Этот тип схемы не имеет петли обратной связи, как реверберационная схема. Продолжение разряда после прекращения стимула называется послеразрядом . Этот тип цепи встречается в рефлекторных дугах некоторых рефлексов . [ 8 ]
Методы исследования
[ редактировать ]различные методы нейровизуализации Для исследования активности нейронных цепей и сетей были разработаны . Использование «сканеров мозга» или функциональной нейровизуализации для исследования структуры или функций мозга является обычным явлением либо просто как способ лучшей оценки черепно-мозговой травмы с помощью изображений с высоким разрешением, либо путем изучения относительных активаций различных областей мозга. Такие технологии могут включать функциональную магнитно-резонансную томографию (фМРТ), позитронно-эмиссионную томографию головного мозга (ПЭТ головного мозга) и компьютерную аксиальную томографию (КТ). Функциональная нейровизуализация использует специальные технологии визуализации мозга для получения изображений мозга, обычно когда человек выполняет определенную задачу, в попытке понять, как активация определенных областей мозга связана с этой задачей. При функциональной нейровизуализации, особенно фМРТ, которая измеряет гемодинамическую активность (с использованием визуализации с ЖИРНЫМ контрастом ПЭТ и электроэнцефалография ), которая тесно связана с нервной активностью, используется (ЭЭГ).
Коннекционистские модели служат испытательной платформой для различных гипотез представления, обработки информации и передачи сигналов. Исследования повреждений в таких моделях, например, в искусственных нейронных сетях , где части узлов намеренно уничтожаются, чтобы увидеть, как работает сеть, также могут дать важную информацию о работе нескольких сборок клеток. Точно так же моделирование дисфункциональных нейротрансмиттеров при неврологических заболеваниях (например, дофамина в базальных ганглиях пациентов с болезнью Паркинсона ) может дать представление о механизмах, лежащих в основе моделей когнитивного дефицита, наблюдаемых в конкретной группе пациентов. Прогнозы этих моделей можно проверить на пациентах или с помощью фармакологических манипуляций, а эти исследования, в свою очередь, можно использовать для информирования моделей, что делает процесс итеративным.
Современный баланс между коннекционистским подходом и одноклеточным подходом в нейробиологии был достигнут в результате длительной дискуссии. В 1972 году Барлоу объявил о революции одиночных нейронов : «наше восприятие обусловлено активностью довольно небольшого числа нейронов, выбранных из очень большой популяции преимущественно молчащих клеток». [ 9 ] Этот подход был стимулирован идеей бабушкиной ячейки , выдвинутой двумя годами ранее. Барлоу сформулировал «пять догм» нейронной доктрины. Недавние исследования « бабушкиной клетки » и феномена разреженного кодирования развивают и модифицируют эти идеи. [ 10 ] В экспериментах с отдельными клетками использовались внутричерепные электроды в медиальной височной доле (гиппокамп и окружающая кора). Современное развитие теории концентрации меры (теоремы стохастического разделения) с применением к искусственным нейронным сетям дает математическое обоснование неожиданной эффективности небольших нейронных ансамблей в многомерном мозге. [ 11 ]
Клиническое значение
[ редактировать ]Иногда нейронные цепи могут становиться патологическими и вызывать проблемы, например, при болезни Паркинсона , когда базальные ганглии . вовлекаются [ 12 ] Проблемы в цепи Папеса также могут привести к ряду нейродегенеративных заболеваний, включая болезнь Паркинсона.
См. также
[ редактировать ]- Обратная связь
- Список регионов человеческого мозга
- Сетевая наука
- Нейронное кодирование
- Нейронная инженерия
- Нейронные колебания
- Сети с импульсной связью
- Системная нейробиология
- Коннектомика
- Нервный тракт
- Нейронный путь
- Нервное сплетение
Ссылки
[ редактировать ]- ^ Первс, Дейл (2011). Нейронаука (5-е изд.). Сандерленд, Массачусетс: Синауэр. п. 507. ИСБН 9780878936953 .
- ^ «Нейронные цепи | Центр передового опыта в области интегративной функции мозга» . Центр передового опыта в области интегративной функции мозга . 13 июня 2016 г. Проверено 4 июня 2018 г.
- ^ Майкл С.К. Томас; Джеймс Л. Макклелланд. «Коннекционистские модели познания» (PDF) . Стэнфордский университет . Архивировано из оригинала (PDF) 6 сентября 2015 г. Проверено 31 августа 2015 г.
- ^ Дж. Я. Леттвин; ХР Матурана; У. С. Маккалок; WH Pitts (1959), «Что глаз лягушки говорит мозгу лягушки», Proc. Инст. Радиоинж. , нет. 47, стр. 1940–1951.
- ^ Зюдхоф, TC (2 ноября 2017 г.). «Синаптические комплексы нейрексина: молекулярный код логики нейронных цепей» . Клетка . 171 (4): 745–769. дои : 10.1016/j.cell.2017.10.024 . ПМЦ 5694349 . ПМИД 29100073 .
- ^ Стокко, Андреа; Лебьер, Кристиан; Андерсон, Джон Р. (2010). «Условная маршрутизация информации в кору: модель роли базальных ганглиев в когнитивной координации» . Психологический обзор . 117 (2): 541–74. дои : 10.1037/a0019077 . ПМК 3064519 . ПМИД 20438237 .
- ^ Гертин, Пенсильвания (2012). «Центральный генератор паттернов локомоции: анатомические, физиологические и патофизиологические соображения» . Границы в неврологии . 3 : 183. doi : 10.3389/fneur.2012.00183 . ПМЦ 3567435 . ПМИД 23403923 .
- ^ Jump up to: а б с д и Саладин, К. (2011). Анатомия человека (3-е изд.). МакГроу-Хилл. п. 364. ИСБН 9780071222075 .
- ^ Барлоу, HB (1 декабря 1972 г.). «Отдельные единицы и ощущение: нейронная доктрина для перцептивной психологии?» . Восприятие . 1 (4): 371–394. дои : 10.1068/p010371 . ПМИД 4377168 . S2CID 17487970 .
- ^ Куиан Кирога, R; Редди, Л; Крейман, Г; Кох, С; Фрид, я (23 июня 2005 г.). «Инвариантное визуальное представление одиночными нейронами человеческого мозга» . Природа . 435 (7045): 1102–1107. Бибкод : 2005Natur.435.1102Q . дои : 10.1038/nature03687 . ПМИД 15973409 .
- ^ Горбань, Александр Н.; Макаров Валерий А.; Тюкин, Иван Юрьевич (июль 2019 г.). «Необоснованная эффективность небольших нейронных ансамблей в многомерном мозге» . Обзоры физики жизни . 29 : 55–88. arXiv : 1809.07656 . Бибкод : 2019PhLRv..29...55G . дои : 10.1016/j.plrev.2018.09.005 . ПМИД 30366739 .
- ^ французский, ИТ; Мутусами, штат Калифорния (2018). «Обзор педункулопонтинного ядра при болезни Паркинсона» . Границы стареющей неврологии . 10:99 . дои : 10.3389/fnagi.2018.00099 . ПМЦ 5933166 . ПМИД 29755338 .
Дальнейшее чтение
[ редактировать ]- Внутренняя пластичность Роберт Х. Кадмор, Нирадж С. Десаи Scholarpedia 3 (2): 1363. doi:10.4249/scholarpedia.1363
Внешние ссылки
[ редактировать ]- Сравнение нейронных сетей мозга и искусственных нейронных сетей
- Конспекты лекций в MIT OpenCourseWare
- Вычисления в мозгу
- Набор инструментов для биологических нейронных сетей. Архивировано 14 июля 2016 г. на Wayback Machine — бесплатный набор инструментов Matlab для моделирования сетей из нескольких различных типов нейронов.
- WormWeb.org: Интерактивная визуализация нейронной сети C. elegans . C. elegans, нематода с 302 нейронами, является единственным организмом, у которого была обнаружена вся нейронная сеть. Используйте этот сайт для просмотра сети и поиска путей между любыми двумя нейронами.
- Введение в нейроны и нейрональные сети , Neuroscience Online (электронный учебник по нейробиологии)
- Сети задержек импульсов (сети волновых помех). Архивировано 30 марта 2016 г. на Wayback Machine.

| Базы данных органов управления : Национальные |
|---|