Глицеральдегид-3-фосфатдегидрогеназа
| ГАФДГ | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | GAPDH , GAPD, G3PD, HEL-S-162eP, глицеральдегид-3-фосфатдегидрогеназа | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 138400 ; МГИ : 5434255 ; Гомологен : 107053 ; GeneCards : GAPDH ; ОМА : GAPDH – ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||


| Глицеральдегид-3-фосфатдегидрогеназа, НАД-связывающий домен | |||
|---|---|---|---|
 детерминанты термостабильности фермента, наблюдаемые в молекулярной структуре thermus aquaticus d-глицеральдегид-3-фосфатдегидрогеназы при разрешении 2,5 ангстрем | |||
| Идентификаторы | |||
| Символ | Gp_dh_N | ||
| Пфам | PF00044 | ||
| Пфам Клан | CL0063 | ||
| ИнтерПро | ИПР020828 | ||
| PROSITE | PDOC00069 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1гд1 / СКОПе / СУПФАМ | ||
| |||
| Глицеральдегид-3-фосфатдегидрогеназа, С-концевой домен | |||
|---|---|---|---|
 кристаллическая структура глицеральдегид-3-фосфатдегидрогеназы пирококка хорикоши ot3 | |||
| Идентификаторы | |||
| Символ | Gp_dh_C | ||
| Пфам | PF02800 | ||
| Пфам Клан | CL0139 | ||
| ИнтерПро | ИПР020829 | ||
| PROSITE | PDOC00069 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1гд1 / СКОПе / СУПФАМ | ||
| |||
Глицеральдегид-3-фосфатдегидрогеназа (сокращенно ГАФД ) ( EC 1.2.1.12 ) представляет собой фермент массой около 37 кДа, который катализирует шестую стадию гликолиза и, таким образом, служит для расщепления глюкозы для получения энергии и молекул углерода. В дополнение к этой давно установленной метаболической функции, GAPDH недавно была вовлечена в несколько неметаболических процессов, включая активацию транскрипции , инициацию апоптоза и т.д. [ 4 ] Перемещение пузырьков ЭР к Гольджи и быстрый аксональный или аксоплазматический транспорт . [ 5 ] специфичный для семенников изофермент GAPDHS В сперме экспрессируется .
Структура
[ редактировать ]В нормальных клеточных условиях цитоплазматическая ГАФД существует преимущественно в виде тетрамера . Эта форма состоит из четырех идентичных субъединиц массой 37 кДа, каждая из которых содержит одну каталитическую тиоловую группу и имеет решающее значение для каталитической функции фермента. [ 6 ] [ 7 ] Ядерная ГАФД имеет повышенную изоэлектрическую точку (pI) при pH 8,3–8,7. [ 7 ] Следует отметить, что цистеина остаток фермента C152 в активном центре необходим для индукции апоптоза при окислительном стрессе . [ 7 ] Примечательно, что посттрансляционные модификации цитоплазматической ГАФД способствуют ее функциям вне гликолиза. [ 6 ]
GAPDH кодируется одним геном, который продуцирует один транскрипт мРНК с 8 вариантами сплайсинга, хотя изоформа существует в виде отдельного гена, который экспрессируется только в сперматозоидах . [ 7 ]
Реакция
[ редактировать ]| глицеральдегид-3-фосфат | глицеральдегидфосфатдегидрогеназа | D - глицерат 1,3-бисфосфат | |

|
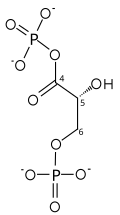
| ||
| ОНИ + +П я | НАДН + Н + | ||
| ОНИ + +П я | НАДН + Н + | ||
Соединение C00118 в базе данных KEGG Pathway. Фермент 1.2.1.12 в базе данных KEGG Pathway. Реакция R01063 в базе данных KEGG Pathway. Соединение C00236 в базе данных KEGG Pathway.
Двухэтапное преобразование G3P
[ редактировать ]Первая реакция — окисление глицеральдегид-3-фосфата (G3P) по положению-1 (на схеме он показан как 4-й углерод гликолиза), при котором альдегид превращается в карбоновую кислоту (ΔG°'=- 50 кДж/моль (-12 ккал/моль)) и НАД+ одновременно эндергонически восстанавливается до НАДН.
Энергия, выделяемая в результате этой высокоэкзергонической реакции окисления, запускает вторую эндергоническую реакцию (ΔG°'=+50 кДж/моль (+12 ккал/моль)), в которой молекула неорганического фосфата переносится на промежуточный продукт GAP с образованием продукта с высокий потенциал переноса фосфорила: 1,3-бисфосфоглицерат (1,3-БФГ).
Это пример фосфорилирования в сочетании с окислением, и общая реакция является несколько эндергонической (ΔG°'=+6,3 кДж/моль (+1,5)). Энергетическая связь здесь стала возможной благодаря GAPDH.
Механизм
[ редактировать ]GAPDH использует ковалентный катализ и общий основной катализ для уменьшения очень большой энергии активации второй стадии (фосфорилирования) этой реакции.
1: Окисление
[ редактировать ]Во-первых, остаток цистеина в активном центре GAPDH атакует карбонильную группу G3P, создавая полутиоацеталь- промежуточный продукт (ковалентный катализ).
Гемитиоацеталь депротонируется остатком гистидина в активном центре фермента (общий основной катализ). Депротонирование способствует реформированию карбонильной группы в последующем промежуточном тиоэфире и выбросу гидрид-иона .
Далее соседняя, прочно связанная молекула НАД. + принимает ион гидрида , образуя НАДН , в то время как гемитиоацеталь окисляется до тиоэфира .
Эта разновидность тиоэфира имеет гораздо более высокую энергию (менее стабильна), чем разновидность карбоновой кислоты , которая могла бы возникнуть, если бы G3P окислялся в отсутствие GAPDH (виды карбоновой кислоты настолько низки по энергии, что энергетический барьер для второй стадии реакции (фосфорилирование) было бы слишком сильным, а реакция, следовательно, слишком медленной и неблагоприятной для живого организма).
2: Фосфорилирование
[ редактировать ]НАДН покидает активный центр и заменяется другой молекулой НАД. + , положительный заряд которого стабилизирует отрицательно заряженный карбонильный кислород в переходном состоянии следующей и последней стадии. Наконец, молекула неорганического фосфата атакует тиоэфир и образует тетраэдрический промежуточный продукт, который затем разрушается с высвобождением 1,3-бисфосфоглицерата и тиоловой группы цистеинового остатка фермента.
Регулирование
[ редактировать ]Этот белок может использовать морфеиновую модель аллостерической регуляции . [ 8 ]
Функция
[ редактировать ]Метаболический
[ редактировать ]Как следует из названия, глицеральдегид-3-фосфатдегидрогеназа (ГАФД) катализирует превращение глицеральдегид-3-фосфата в D - глицерат-1,3-бисфосфат . Это шестой этап гликолитического распада глюкозы, важного пути доставки энергии и молекул углерода, который происходит в цитозоле эукариотических клеток. Преобразование происходит в два связанных шага. Первое благоприятно и позволяет произойти второму неблагоприятному шагу.
Адгезия
[ редактировать ]GAPDH Одной из подрабатывающих функций является ее роль в адгезии и связывании с другими партнерами. Известно, что бактериальная GAPDH из Mycoplasma и Streptococcus и грибковая GAPDH из Paracoccidioides brasiliensis связываются с компонентом внеклеточного матрикса человека и действуют посредством адгезии. [ 9 ] [ 10 ] [ 11 ] Обнаружено, что GAPDH связан с поверхностью, способствуя адгезии, а также конкурентному исключению вредных патогенов. [ 12 ] GAPDH Candida albicans связан с клеточной стенкой и связывается с фибронектином и ламинином . [ 13 ] Известно, что GAPDH из видов пробиотиков связывает муцин толстой кишки человека и ЕСМ, что приводит к усилению колонизации пробиотиков в кишечнике человека. [ 14 ] [ 15 ] [ 16 ] Patel D. и др. показали, что Lactobacillus acidophilus GAPDH связывается с муцином, действуя в качестве адгезии. [ 17 ]
Транскрипция и апоптоз
[ редактировать ]GAPDH сама может активировать транскрипцию . Транскрипционный коактиваторный комплекс OCA -S содержит GAPDH и лактатдегидрогеназу , два белка, которые, как считалось ранее, участвуют только в метаболизме . GAPDH перемещается между цитозолем и ядром и, таким образом, может связывать метаболическое состояние с транскрипцией гена. [ 18 ]
В 2005 году Хара и др. показали, что GAPDH инициирует апоптоз . Это не третья функция, но ее можно рассматривать как активность, опосредованную связыванием GAPDH с ДНК , как при активации транскрипции, обсуждавшейся выше. Исследование показало, что GAPDH S-нитрозилируется NO в ответ на клеточный стресс, что заставляет его связываться с белком SIAH1 , убиквитинлигазой . Комплекс перемещается в ядро, где Siah1 нацелен на деградацию ядерных белков , тем самым инициируя контролируемое отключение клеток. [ 19 ] В последующем исследовании группа продемонстрировала, что депренил , который использовался клинически для лечения болезни Паркинсона , сильно снижает апоптотическое действие ГАФД, предотвращая ее S-нитрозилирование, и, таким образом, может использоваться в качестве лекарственного средства. [ 20 ]
Метаболический переключатель
[ редактировать ]GAPDH действует как обратимый метаболический переключатель при окислительном стрессе. [ 21 ] Когда клетки подвергаются воздействию окислителей , им необходимо избыточное количество антиоксидантного кофактора НАДФН . В цитозоле НАДФН восстанавливается из НАДФ+ с помощью нескольких ферментов, три из которых катализируют первые этапы пентозофосфатного пути . Окислительная обработка вызывает инактивацию GAPDH. Эта инактивация временно перенаправляет метаболический поток с гликолиза на пентозофосфатный путь, позволяя клетке генерировать больше НАДФН. [ 22 ] В условиях стресса НАДФН необходим некоторым антиоксидантным системам, включая глутаредоксин и тиоредоксин , а также необходим для переработки глутатиона .
Транспорт от скорой помощи к Гольджи
[ редактировать ]GAPDH также, по-видимому, участвует в транспорте везикул из эндоплазматического ретикулума (ЭР) в аппарат Гольджи , который является частью пути доставки секретируемых белков. Было обнаружено, что GAPDH рекрутируется с помощью rab2 в везикулярно-трубчатые кластеры ЭР, где он помогает формировать пузырьки COP 1 . GAPDH активируется посредством тирозина фосфорилирования с помощью Src . [ 23 ]
Дополнительные функции
[ редактировать ]GAPDH, как и многие другие ферменты, выполняет множество функций. Помимо катализа 6-й стадии гликолиза , недавние данные указывают на участие GAPDH в других клеточных процессах. Было описано, что GAPDH демонстрирует мультифункциональность более высокого порядка в контексте поддержания клеточного гомеостаза железа. [ 24 ] в частности, в качестве белка-шаперона для лабильного гема внутри клеток. [ 25 ] Это стало неожиданностью для исследователей, но с эволюционной точки зрения имеет смысл повторно использовать и адаптировать существующие белки вместо того, чтобы создавать новый белок с нуля.
Использование в качестве контроля загрузки
[ редактировать ]Поскольку ген GAPDH часто стабильно и конститутивно экспрессируется на высоких уровнях в большинстве тканей и клеток, его считают геном домашнего хозяйства . По этой причине GAPDH обычно используется исследователями-биологами в качестве контроля нагрузки для вестерн-блоттинга и в качестве контроля для qPCR . Однако исследователи сообщили о различной регуляции GAPDH в определенных условиях. [ 26 ] Например, транскрипционный фактор MZF-1 регулирует ген GAPDH. было показано, что [ 27 ] Гипоксия также сильно активирует GAPDH. [ 28 ] Следовательно, использование GAPDH в качестве контроля нагрузки должно быть тщательно продумано.
Сотовое распространение
[ редактировать ]Все стадии гликолиза происходят в цитозоле , как и реакция, катализируемая ГАФД. В эритроцитах ГАФД и некоторые другие гликолитические ферменты собираются в комплексы на внутренней стороне клеточной мембраны . Этот процесс, по-видимому, регулируется фосфорилированием и оксигенацией. [ 29 ] Ожидается, что сближение нескольких гликолитических ферментов значительно увеличит общую скорость распада глюкозы. Недавние исследования также показали, что GAPDH экспрессируется железозависимым образом на внешней стороне клеточной мембраны, где она играет роль в поддержании клеточного гомеостаза железа. [ 30 ] [ 31 ]
Клиническое значение
[ редактировать ]Рак
[ редактировать ]GAPDH сверхэкспрессируется при многих видах рака человека, таких как меланома кожи , и ее экспрессия положительно коррелирует с прогрессированием опухоли. [ 32 ] [ 33 ] Его гликолитические и антиапоптотические функции способствуют пролиферации и защите опухолевых клеток, способствуя онкогенезу . Примечательно, что GAPDH защищает от укорочения теломер, вызванного химиотерапевтическими препаратами, которые стимулируют сфинголипид- церамид . Между тем, такие состояния, как окислительный стресс , нарушают функцию GAPDH, что приводит к клеточному старению и смерти. [ 7 ] Более того, истощение GAPDH привело к старению опухолевых клеток, что представляет собой новую терапевтическую стратегию контроля роста опухоли. [ 34 ]
нейродегенерация
[ редактировать ]GAPDH участвует в ряде нейродегенеративных заболеваний и расстройств, в основном за счет взаимодействия с другими белками, специфичными для этого заболевания или расстройства. Эти взаимодействия могут влиять не только на энергетический обмен, но и на другие функции GAPDH. [ 6 ] Например, взаимодействия GAPDH с белком-предшественником бета-амилоида (betaAPP) могут мешать его функции в отношении цитоскелета или мембранного транспорта, тогда как взаимодействия с хантингтином могут мешать его функции в отношении апоптоза, ядерной тРНК транспорта , репликации ДНК и репарации ДНК . (БП) сообщалось о ядерной транслокации ГАФД Кроме того, при болезни Паркинсона , а некоторые антиапоптотические препараты, такие как разагилин , действуют, предотвращая ядерную транслокацию ГАФД. Предполагается, что гипометаболизм может быть одним из факторов, способствующих БП, но точные механизмы, лежащие в основе участия GAPDH в нейродегенеративных заболеваниях, еще предстоит выяснить. [ 35 ] SNP rs3741916 в 5'- UTR гена GAPDH может быть связан с поздним началом болезни Альцгеймера . [ 36 ]
Взаимодействия
[ редактировать ]Партнеры по связыванию белков
[ редактировать ]GAPDH участвует в ряде биологических функций посредством межбелковых взаимодействий с:
- тубулин для облегчения связывания микротрубочек; [ 6 ]
- актин для облегчения полимеризации актина; [ 6 ]
- VDAC1 индуцирует проницаемость митохондриальных мембран (MMP) и апоптоз; [ 6 ]
- Инозитол-1,4,5-трифосфатный рецептор для регуляции внутриклеточной Ca2+ передачи сигналов ; [ 6 ]
- Oct-1 для образования коактиваторного комплекса OCA-S, который необходим для гистона H2B синтеза во время S-фазы клеточного цикла ; [ 7 ]
- p22 для помощи в организации микротрубочек ; [ 7 ]
- Rab2 для облегчения эндоплазматической сети (ER) – Гольджи ; транспорта [ 7 ]
- Трансферрин на поверхности разнообразных клеток и во внеклеточной жидкости; [ 7 ] [ 31 ] [ 37 ]
- лактатдегидрогеназа ; [ 7 ]
- Лактоферрин; [ 38 ]
- Апуриновая/апиримидиновая эндонуклеаза ( APE1 ), преобразующая таким образом окисленный APE1 в восстановленную форму, чтобы возобновить его эндонуклеазную активность; [ 7 ]
- Белок промиелоцитарного лейкоза (PML) РНК -зависимым образом; [ 7 ]
- Rheb для секвестрации ГТФазы в условиях низкого уровня глюкозы; [ 7 ]
- Siah1 образует комплекс, который перемещается в ядро, где убиквитинирует и разрушает ядерные белки в условиях нитрозативного стресса; [ 7 ]
- Конкурент GAPDH белка Siah улучшает жизнь (GOSPEL), чтобы блокировать взаимодействие GAPDH с Siah1 и, таким образом, гибель клеток в ответ на окислительный стресс; [ 7 ]
- p300/ CREB-связывающий белок (CBP), который ацетилирует GAPDH и, в свою очередь, усиливает ацетилирование дополнительных апоптотических мишеней; [ 7 ]
- специфичная для скелетных мышц Са2+/кальмодулинзависимая протеинкиназа; [ 7 ]
- Акт ; [ 7 ]
- Белок -предшественник бета-амилоида (бетаАРР); [ 35 ]
- Хантингтин . [ 35 ]
- ГАФД может самоассоциироваться в гомотипические олигомеры/агрегаты.
Партнеры по связыванию нуклеиновых кислот
[ редактировать ]GAPDH связывается с одноцепочечной РНК [ 39 ] ДНК и ряд партнеров по связыванию нуклеиновых кислот были идентифицированы: [ 7 ]
- тРНК ,
- гепатита А , РНК вируса
- гепатита В , РНК вируса
- гепатита С , РНК вируса
- ВПЧ3,
- лимфокина , мРНК
- ИФН-γ , мРНК
- мРНК JEV и
- теломерная ДНК.
Ингибиторы
[ редактировать ]Интерактивная карта маршрутов
[ редактировать ]Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [ § 1 ]

- ^ Интерактивную карту маршрутов можно редактировать на WikiPathways: «Гликолиз-Глюконеогенез_WP534» .
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000111640 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Тарз А., Денио А., Ле Брас М., Майе Э., Молле Д., Ларошетт Н., Замзами Н., Ян Г., Кремер Г., Бреннер С. (апрель 2007 г.). «GAPDH, новый регулятор проапоптотической проницаемости митохондриальных мембран». Онкоген . 26 (18): 2606–2620. дои : 10.1038/sj.onc.1210074 . ПМИД 17072346 . S2CID 20291542 .
- ^ Зала Д., Хинкельман М.В., Ю Х., Лира да Кунья М.М., Лиот Г., Кордельер Ф.П., Марко С., Сауду Ф. (январь 2013 г.). «Везикулярный гликолиз обеспечивает запас энергии для быстрого аксонального транспорта» . Клетка . 152 (3): 479–491. дои : 10.1016/j.cell.2012.12.029 . ПМИД 23374344 .
- ^ Jump up to: а б с д и ж г Тристан С., Шахани Н., Седлак Т.В., Сава А. (февраль 2011 г.). «Разнообразные функции GAPDH: взгляды из разных субклеточных компартментов» . Сотовая сигнализация . 23 (2): 317–323. doi : 10.1016/j.cellsig.2010.08.003 . ПМЦ 3084531 . ПМИД 20727968 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с Николлс С., Ли Х., Лю Дж. П. (август 2012 г.). «ГАФД: общий фермент с необычными функциями». Клиническая и экспериментальная фармакология и физиология . 39 (8): 674–679. дои : 10.1111/j.1440-1681.2011.05599.x . ПМИД 21895736 . S2CID 23499684 .
- ^ Селвуд Т., Яффе Е.К. (март 2012 г.). «Динамическая диссоциация гомоолигомеров и контроль функции белка» . Архив биохимии и биофизики . 519 (2): 131–143. дои : 10.1016/j.abb.2011.11.020 . ПМЦ 3298769 . ПМИД 22182754 .
- ^ Думке Р., Хауснер М., Джейкобс Э. (август 2011 г.). «Роль глицеральдегид-3-фосфатдегидрогеназы Mycoplasma pneumoniae (GAPDH) в опосредовании взаимодействий с внеклеточным матриксом человека» . Микробиология . 157 (Часть 8): 2328–2338. дои : 10.1099/mic.0.048298-0 . ПМИД 21546586 .
- ^ Брассар Дж., Готшалк М., Кесси С. (август 2004 г.). «Клонирование и очистка глицеральдегид-3-фосфатдегидрогеназы Streptococcus suis серотипа 2 и ее участие в качестве адгезина». Ветеринарная микробиология . 102 (1–2): 87–94. дои : 10.1016/j.vetmic.2004.05.008 . ПМИД 15288930 .
- ^ Барбоза М.С., Бао С.Н., Андреотти П.Ф., де Фариа Ф.П., Фелипе М.С., душ Сантос Фейтоса Л., Мендес-Джаннини М.Дж., Соареш К.М. (январь 2006 г.). «Глицеральдегид-3-фосфатдегидрогеназа Paracoccidioides brasiliensis представляет собой белок клеточной поверхности, участвующий в адгезии грибов к белкам внеклеточного матрикса и взаимодействии с клетками» . Инфекция и иммунитет . 74 (1): 382–389. дои : 10.1128/IAI.74.1.382-389.2006 . ПМЦ 1346668 . ПМИД 16368993 .
- ^ Рамиа К., ван Ринен, Калифорния, Дикс Л.М. (1 июля 2008 г.). «Поверхностно-связанные белки Lactobacillus plantarum 423, которые способствуют адгезии клеток Caco-2, и их роль в конкурентном исключении и вытеснении Clostridium sporogenes и Enterococcus faecalis» . Исследования в области микробиологии . 159 (6): 470–475. дои : 10.1016/j.resmic.2008.06.002 . ПМИД 18619532 .
- ^ Госальбо Д., Хиль-Наварро I, Асорин I, Ренау-Пикерас Х., Мартинес Х.П., Хиль М.Л. (май 1998 г.). «Связанная с клеточной стенкой глицеральдегид-3-фосфатдегидрогеназа Candida albicans также является белком, связывающим фибронектин и ламинин» . Инфекция и иммунитет . 66 (5): 2052–2059. дои : 10.1128/IAI.66.5.2052-2059.1998 . ПМЦ 108162 . ПМИД 9573088 .
- ^ Дэн З, Дай Т, Чжан В, Чжу Дж, Луо XM, Фу Д, Лю Дж, Ван Х (декабрь 2020 г.). «Глицеральдегид-3-фосфатдегидрогеназа увеличивает адгезию Lactobacillus reuteri к муцину хозяина, усиливая пробиотические эффекты» . Международный журнал молекулярных наук . 21 (24): 9756. doi : 10.3390/ijms21249756 . ПМЦ 7766874 . ПМИД 33371288 .
- ^ Киносита Х, Учида Х, Каваи Ю, Кавасаки Т, Вакахара Н, Мацуо Х, Ватанабе М, Китазава Х, Онума С, Миура К, Хории А, Сайто Т (июнь 2008 г.). «Клеточная поверхность Lactobacillus plantarum LA 318 глицеральдегид-3-фосфатдегидрогеназа (GAPDH) прикрепляется к муцину толстой кишки человека». Журнал прикладной микробиологии . 104 (6): 1667–1674. дои : 10.1111/j.1365-2672.2007.03679.x . ПМИД 18194256 . S2CID 22346488 .
- ^ Киносита Х, Вакахара Н, Ватанабэ М, Кавасаки Т, Мацуо Х, Каваи Ю, Китазава Х, Онума С, Миура К, Хории А, Сайто Т (1 ноября 2008 г.). «Глицеральдегид-3-фосфатдегидрогеназа клеточной поверхности (ГАФДГ) Lactobacillus plantarum LA 318 распознает антигены групп крови человека A и B» . Исследования в области микробиологии . 159 (9–10): 685–691. дои : 10.1016/j.resmic.2008.07.005 . ПМИД 18790050 .
- ^ Патель Д.К., Шах К.Р., Паппачан А., Гупта С., Сингх Д.Д. (октябрь 2016 г.). «Клонирование, экспрессия и характеристика муцин-связывающей GAPDH из Lactobacillus acidophilus». Международный журнал биологических макромолекул . 91 : 338–346. doi : 10.1016/j.ijbiomac.2016.04.041 . ПМИД 27180300 .
- ^ Чжэн Л., Рёдер Р.Г., Луо Ю (июль 2003 г.). «Активация S-фазы промотора гистона H2B с помощью OCA-S, коактиваторного комплекса, который содержит GAPDH в качестве ключевого компонента» . Клетка . 114 (2): 255–266. дои : 10.1016/S0092-8674(03)00552-X . ПМИД 12887926 . S2CID 5543647 .
- ^ Хара М.Р., Агравал Н., Ким С.Ф., Касио М.Б., Фуджимуро М., Озэки Ю., Такахаси М., Чеа Дж.Х., Танкоу С.К., Хестер Л.Д., Феррис К.Д., Хейворд С.Д., Снайдер Ш., Сава А. (июль 2005 г.). «S-нитрозилированная GAPDH инициирует апоптотическую гибель клеток путем ядерной транслокации после связывания Siah1». Природная клеточная биология . 7 (7): 665–674. дои : 10.1038/ncb1268 . ПМИД 15951807 . S2CID 1922911 .
- ^ Хара М.Р., Томас Б., Касио М.Б., Бэ Б.И., Хестер Л.Д., Доусон В.Л., Доусон Т.М., Сава А., Снайдер Ш. (март 2006 г.). «Нейропротекция путем фармакологической блокады каскада смерти GAPDH» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (10): 3887–3889. Бибкод : 2006PNAS..103.3887H . дои : 10.1073/pnas.0511321103 . ПМК 1450161 . ПМИД 16505364 .
- ^ Агарвал А.Р., Чжао Л., Санчети Х., Сундар И.К., Рахман И., Каденас Э. (ноябрь 2012 г.). «Кратковременное воздействие сигаретного дыма вызывает обратимые изменения в энергетическом обмене и окислительно-восстановительном статусе клеток независимо от воспалительных реакций в легких мышей». Американский журнал физиологии. Клеточная и молекулярная физиология легких . 303 (10): Л889–Л898. дои : 10.1152/ajplung.00219.2012 . ПМИД 23064950 .
- ^ Ральсер М., Вамелинк М.М., Ковальд А., Гериш Б., Херен Г., Стрейс Е.А., Клипп Е., Якобс С., Брайтенбах М., Лерах Х., Кробич С. (декабрь 2007 г.). «Динамическое изменение потока углеводов является ключом к противодействию окислительному стрессу» . Журнал биологии . 6 (4): 10. дои : 10.1186/jbiol61 . ПМК 2373902 . ПМИД 18154684 .
- ^ Тисдейл Э.Дж., Арталехо Ч.Р. (июнь 2007 г.). «Мутант GAPDH, дефектный в Src-зависимом фосфорилировании тирозина, препятствует событиям, опосредованным Rab2» . Трафик . 8 (6): 733–741. дои : 10.1111/j.1600-0854.2007.00569.x . ПМЦ 3775588 . ПМИД 17488287 .
- ^ Борадиа В.М., Радже М., Радже С.И. (декабрь 2014 г.). «Подработка белка в метаболизме железа: глицеральдегид-3-фосфатдегидрогеназа (ГАФДГ)». Труды Биохимического общества . 42 (6): 1796–1801. дои : 10.1042/BST20140220 . ПМИД 25399609 .
- ^ Суини Э.А., Сингх А.Б., Чакраварти Р., Мартинес-Гузман О, Сайни А., Хак М.М., Гари Дж., Дэнс П.Д., Ганнибал Л., Редди А.Р., Стюр DJ (сентябрь 2018 г.). «Глицеральдегид-3-фосфатдегидрогеназа является шапероном, выделяющим в клетках лабильный гем» . Журнал биологической химии . 293 (37): 14557–14568. дои : 10.1074/jbc.RA118.004169 . ПМК 6139559 . ПМИД 30012884 .
- ^ Барбер Р.Д., Хармер Д.В., Коулман Р.А., Кларк Б.Дж. (май 2005 г.). «GAPDH как ген домашнего хозяйства: анализ экспрессии мРНК GAPDH на панели из 72 тканей человека». Физиологическая геномика . 21 (3): 389–395. CiteSeerX 10.1.1.459.7039 . doi : 10.1152/физиологгеномика.00025.2005 . ПМИД 15769908 .
- ^ Пищатовский Р.Т., Рафферти Б.Дж., Розадо А., Тобак С., Ленц Н.Х. (август 2014 г.). «Ген глицеральдегид-3-фосфатдегидрогеназы (GAPDH) регулируется миелоидным цинковым пальцем 1 (MZF-1) и индуцируется кальцитриолом». Связь с биохимическими и биофизическими исследованиями . 451 (1): 137–141. дои : 10.1016/j.bbrc.2014.07.082 . ПМИД 25065746 .
- ^ Сайгин Д., Табиб Т., Биттар Х.Е., Валензи Э., Сембрат Дж., Чан С.Ю., Рохас М., Лафиатис Р. (2008). «Транскрипционное профилирование популяций клеток легких при идиопатической легочной артериальной гипертензии» . Легочное кровообращение . 10 (1): 239–243. дои : 10.1007/s11684-008-0045-7 . ПМК 7052475 . ПМИД 32166015 . S2CID 85327763 .
- ^ Кампанелла М.Э., Чу Х., Лоу PS (февраль 2005 г.). «Сборка и регуляция гликолитического ферментного комплекса на мембране эритроцитов человека» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (7): 2402–2407. Бибкод : 2005PNAS..102.2402C . дои : 10.1073/pnas.0409741102 . ПМК 549020 . ПМИД 15701694 .
- ^ Сировер М.А. (декабрь 2014 г.). «Структурный анализ функционального разнообразия глицеральдегид-3-фосфатдегидрогеназы» . Международный журнал биохимии и клеточной биологии . 57 : 20–26. doi : 10.1016/j.biocel.2014.09.026 . ПМЦ 4268148 . ПМИД 25286305 .
- ^ Jump up to: а б Кумар С., Шеоканд Н., Мхадешвар М.А., Радже С.И., Радже М. (январь 2012 г.). «Характеристика глицеральдегид-3-фосфатдегидрогеназы как нового рецептора трансферрина». Международный журнал биохимии и клеточной биологии . 44 (1): 189–199. дои : 10.1016/j.biocel.2011.10.016 . ПМИД 22062951 .
- ^ Рамос Д., Пеллин-Карселен А., Агусти Х., Мурги А., Хорда Е., Пеллин А., Монтеагудо К. (январь 2015 г.). «Дерегуляция экспрессии глицеральдегид-3-фосфатдегидрогеназы во время прогрессирования опухоли меланомы кожи человека». Противораковые исследования . 35 (1): 439–444. ПМИД 25550585 .
- ^ Ван Д., Мутхарт Д.Р., Лоуи Д.Р., Цянь Икс (2013). «Экспрессия генов, связанных с клеточным циклом глицеральдегид-3-фосфатдегидрогеназы (GACC), коррелирует со стадией рака и плохой выживаемостью у пациентов с солидными опухолями» . ПЛОС ОДИН . 8 (4): e61262. Бибкод : 2013PLoSO...861262W . дои : 10.1371/journal.pone.0061262 . ПМЦ 3631177 . ПМИД 23620736 .
- ^ Фадке М., Кринецкая Н., Мишра А., Кринецкий Е. (июль 2011 г.). «Фенотип ускоренного клеточного старения клеток карциномы легких человека, обедненных GAPDH» . Связь с биохимическими и биофизическими исследованиями . 411 (2): 409–415. дои : 10.1016/j.bbrc.2011.06.165 . ПМК 3154080 . ПМИД 21749859 .
- ^ Jump up to: а б с Маццола Дж.Л., Сировер М.А. (октябрь 2002 г.). «Изменение внутриклеточной структуры и функции глицеральдегид-3-фосфатдегидрогеназы: общий фенотип нейродегенеративных нарушений?». Нейротоксикология . 23 (4–5): 603–609. дои : 10.1016/s0161-813x(02)00062-1 . ПМИД 12428732 .
- ^ Аллен М., Кокс С., Белбин О., Ма Л., Бишельо Г.Д., Уилкокс С.Л., Хауэлл К.С., Хантер Т.А., Калли О., Уокер Л.П., Карраскильо М.М., Диксон Д.В., Петерсен Р.К., Графф-Рэдфорд Н.Р., Юнкин С.Г., Эртекин-Танер Н (январь 2012 г.). «Ассоциация и гетерогенность локуса GAPDH при болезни Альцгеймера» . Нейробиология старения . 33 (1): 203.e25–203.e33. doi : 10.1016/j.neurobiolaging.2010.08.002 . ПМК 3017231 . ПМИД 20864222 .
- ^ Радже С.И., Кумар С., Харл А., Нанда Дж.С., Радже М. (февраль 2007 г.). «Глицеральдегид-3-фосфатдегидрогеназа на поверхности клеток макрофагов представляет собой новый рецептор трансферрина» . Журнал биологической химии . 282 (5): 3252–3261. дои : 10.1074/jbc.M608328200 . ПМИД 17121833 .
- ^ Чаухан А.С., Рават П., Малхотра Х., Шеоканд Н., Кумар М., Патидар А., Чаудхари С., Джахар П., Радже С.И., Радже М. (декабрь 2015 г.). «Секретируемая многофункциональная глицеральдегид-3-фосфатдегидрогеназа связывает лактоферрин и железо в клетки неканоническим путем» . Научные отчеты . 5 : 18465. Бибкод : 2015NatSR...518465C . дои : 10.1038/srep18465 . ПМК 4682080 . ПМИД 26672975 .
- ^ Уайт М.Р., Хан М.М., Деридж Д., Росс Ч.Р., Квинтин Р., Зуккони Б.Е., Высоцкий В.Х., Уинтрод П.Л., Уилсон Г.М., Гарсин Э.Д. (январь 2015 г.). «Мутация димерного интерфейса глицеральдегид-3-фосфатдегидрогеназы регулирует ее связывание с РНК, богатой AU» . Журнал биологической химии . 290 (3): 1770–1785. дои : 10.1074/jbc.M114.618165 . ПМК 4340419 . ПМИД 25451934 .
- ^ Jump up to: а б с Герлах М., Райхманн Х., Ридерер П. (2012). «Критический обзор доказательств доклинических различий между разагилином и селегилином». Базальные ганглии . 2 (4): С9–С15. дои : 10.1016/j.baga.2012.04.032 .
Дальнейшее чтение
[ редактировать ]- Воет Д., Воет Дж.Г. (2010). Биохимия . Нью-Йорк: Уайли. ISBN 978-0-470-57095-1 .
- Страйер Л., Берг Дж.М., Тимочко Дж.Л. (2002). Биохимия, пятое издание и тетрадь лекций . Сан-Франциско: WH Freeman. ISBN 978-0-7167-9804-0 .
- схема механизма реакции GAPDH из Lodish MCB на книжной полке NCBI
- аналогичная диаграмма из книги Alberts The Cell на книжной полке NCBI
Внешние ссылки
[ редактировать ]- PDBe-KB предоставляет обзор всей информации о структуре, доступной в PDB для глицеральдегид-3-фосфатдегидрогеназы человека.
галерея PDB |
|---|












