Перестановка дерева
Перестановки деревьев — это детерминированные алгоритмы, предназначенные для поиска оптимальной структуры филогенетического дерева . Они могут быть применены к любому набору данных, которые естественным образом организованы в виде дерева, но находят наибольшее применение в вычислительной филогенетике , особенно при с максимальной экономией и максимальным правдоподобием поиске филогенетических деревьев , которые стремятся идентифицировать одно среди многих возможных деревьев, которое лучше всего объясняет эволюционная история конкретного гена или вида .
Основные перестановки деревьев
[ редактировать ]-
 Ближайшая соседская развязка (NNI)
Ближайшая соседская развязка (NNI) -
 Обрезка и пересадка поддеревьев (SPR)
Обрезка и пересадка поддеревьев (SPR) -
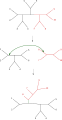 Разделение дерева пополам и пересоединение (TBR)
Разделение дерева пополам и пересоединение (TBR)



Простейшая перестановка дерева, известная как обмен ближайшими соседями , меняет связность четырех поддеревьев внутри основного дерева. Поскольку существует три возможных способа соединения четырех поддеревьев, [ 1 ] и один из них — исходная связность, каждый обмен создает два новых дерева. Полный поиск возможных ближайших соседей для каждого возможного набора поддеревьев — самый медленный, но наиболее оптимизирующий способ выполнения этого поиска. Альтернативный, более широкий поиск, обрезка и пересадка поддерева (SPR), выбирает и удаляет поддерево из основного дерева и повторно вставляет его в другое место основного дерева для создания нового узла. Наконец, разделение дерева пополам и повторное соединение (TBR) отделяет поддерево от основного дерева во внутреннем узле, а затем пытается использовать все возможные соединения между ребрами двух созданных таким образом деревьев. Возрастающая сложность метода перестановки дерева коррелирует с увеличением вычислительного времени, необходимого для поиска, хотя и не обязательно с их производительностью. [ 2 ]
SPR можно разделить на uSPR: некорневой SPR, rSPR: корневой SPR. uSPR применяется к деревьям без корней и действует следующим образом: сломайте любой край. Присоедините один конец ребра (выбранного произвольно) к любому другому ребру в дереве. rSPR применяется к корневым деревьям* и заключается в следующем: сломать любое ребро, кроме ребра, ведущего к корневому узлу. Соедините один конец ребра (точнее: конец ребра, находящийся САМЫМ ДАЛЕЕ от корня) и прикрепите его к любому другому краю дерева. [ 3 ]
* В этом примере корень дерева отмечен узлом первой степени, что означает, что все узлы в дереве имеют степень 1 или степень 3. Альтернативный подход, используемый в Бордевиче и Семпле, состоит в том, чтобы рассматривать корневой узел как иметь степень 2 и иметь специальное правило для rSPR.
Количество СПР [ 4 ] или ТБР [ 5 ] Ходы, необходимые для перехода от одного дерева к другому, можно рассчитать, создав лес максимального согласия, включающий (соответственно) укорененные или неукорененные деревья. Эта проблема NP сложна, но решаема с фиксированным параметром.
Слияние деревьев
[ редактировать ]Самый простой тип слияния деревьев начинается с двух деревьев, которые уже определены как почти оптимальные; таким образом, они, скорее всего, имеют правильное большинство своих узлов, но могут не суметь правильно разрешить отдельные «листья» дерева; например, разделение ((A,B),(C,D)) на кончике ветки по сравнению с ((A,C),(B,D)) может быть неразрешенным. [ 1 ] Слияние деревьев заменяет эти два решения двумя в остальном почти оптимальными деревьями. Варианты метода используют стандартные генетические алгоритмы с определенной целевой функцией для замены поддеревьев с высокими показателями на основные деревья, которые в целом имеют высокие показатели. [ 6 ]
Секторальный поиск
[ редактировать ]Альтернативная стратегия — отделить часть дерева (которую можно выбрать случайным образом или использовать более стратегический подход) и выполнить TBR/SPR/NNI на этом поддереве. Это оптимизированное поддерево затем можно заменить в главном дереве, что, возможно, улучшит p-показатель. [ 7 ]
Дерево дрейфует
[ редактировать ]Чтобы избежать попадания в локальные оптимумы, можно использовать подход «имитированного отжига», при котором алгоритму иногда разрешается использовать неоптимальные деревья-кандидаты с вероятностью, связанной с тем, насколько они далеки от оптимума. [ 7 ]
Слияние деревьев
[ редактировать ]После того, как собран ряд одинаково оптимальных деревьев, часто можно найти лучшее дерево, объединив «хорошие части» отдельных деревьев. Подгруппы с одинаковым составом, но разной топологией можно переключать и оценивать полученные деревья. [ 7 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Фельзенштейн, Джозеф (2004). Выводы о филогениях . Sinauer Associates: Сандерленд, Массачусетс. ISBN 9780878931774 .
- ^ Такахаши, Кей; Ней, Масатоши (август 2000 г.). «Эффективность быстрых алгоритмов филогенетического вывода по критериям максимальной экономии, минимальной эволюции и максимального правдоподобия при использовании большого количества последовательностей» . Молекулярная биология и эволюция . 17 (8): 1251–1258. doi : 10.1093/oxfordjournals.molbev.a026408 . ПМИД 10908645 .
- ^ Бордевич, Магнус; Семпл, Чарльз (2005). «О вычислительной сложности корневого поддерева при обрезке и перепрививке расстояния» . Анналы комбинаторики . 8 (4): 409–423. дои : 10.1007/s00026-004-0229-z . S2CID 13002129 .
- ^ Уидден, Крис; Бейко, Роберт Г.; Зех, Норберт (2016). «Алгоритмы с фиксированными параметрами и аппроксимации для лесов максимального согласия разветвленных деревьев». Алгоритмика . 74 (3): 1019–1054. arXiv : 1305.0512 . дои : 10.1007/s00453-015-9983-z . S2CID 14297537 .
- ^ Чен, Цзянер; Фань, Цзя-Хао; Сзе, Синг-Хой (2015). «Алгоритмы параметризации и аппроксимации леса максимального согласия в разветвленных деревьях» . Теоретическая информатика . 562 : 496–512. дои : 10.1016/j.tcs.2014.10.031 .
- ^ Мацуда, Х. (1996). «Филогенетический вывод белка с использованием максимального правдоподобия с помощью генетического алгоритма» (PDF) . Тихоокеанский симпозиум по биокомпьютингу, 1996 г. стр. 512–523.
- ^ Перейти обратно: а б с Голобов, Пабло А. (1999). «Анализ больших наборов данных в разумные сроки: решения для составной оптимумы» . Кладистика . 15 (4): 415–428. дои : 10.1006/клад.1999.0122 . ПМИД 34902941 .