Дыня (китообразные)

Дыня обнаруженная — это масса жировой ткани, на лбу всех зубатых китов . [1] [2] Он фокусирует и модулирует вокализации животного и действует как звуковая линза. Таким образом, это ключевой орган, участвующий в коммуникации и эхолокации .
Описание
[ редактировать ]Дыня структурно является частью носового аппарата и содержит большую часть массы ткани между дыхалом и кончиком морды. Функция дыни до конца не изучена, но ученые полагают, что это биоакустический компонент, обеспечивающий фокусировку звуков, используемых в эхолокации, и создание сходства между характеристиками ее ткани и окружающей воды, благодаря чему акустическая энергия может вытекать из дыни. голову и в окружающую среду с наименьшими потерями энергии. В прошлом некоторые ученые полагали, что дыня обладает функциями глубокого ныряния и плавучести, но за последние 40 лет эти идеи были отвергнуты, поэтому кетологи больше не считают их обоснованными. [3]
Различный состав дыни создает градиент скорости звука, который направленно преломляет звук. Звуки также отражаются от черепа и воздушных мешков, окружающих дыню. [4]
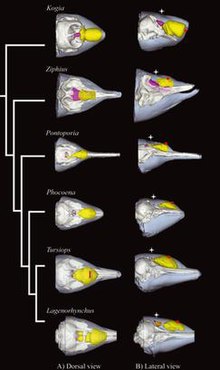
Размер дыни не связан с максимальной глубиной погружения зубатых китов. Особые характеристики дыни, вероятно, больше связаны с филогенией зубатых китов , таксономическими отношениями на протяжении эволюционного времени. У некоторых видов дыни более специализированы, чем у других. самый У кашалота большой нос среди всех животных в мире. Основная часть этого носа состоит из двух крупных жировых структур: спермацета и «мусора». Джанк структурно такой же, как дыня ( гомологичный ей).
Дыня не гомологична спермацетному органу. [1] [5] Исследования экспрессии генов, таких как белок MYH16 , показывают, что он происходит из жевательной мышцы . [6]
Состав
[ редактировать ]Дыня представляет собой смесь триглицеридов и эфиров воска . Точный состав варьируется в зависимости от дыни. Обычно внутренняя сердцевина дыни имеет более высокое содержание воска, чем внешние части, и проводит звук медленнее. Этот градиент преломляет звук и фокусирует его, как линзу.
Липиды дыни не перевариваются животными, так как они метаболически токсичны. У голодающего дельфина крепкая дыня, даже если остальная часть его тела истощена. [1] Липиды в дыне, как правило, имеют более низкую молекулярную массу и более насыщены, чем ворвань .
Дыни Delphinidae ( дельфины) и Physeteroidea (кашалоты) содержат значительное количество воскового эфира, тогда как дыни Phocoenidae (морские свиньи) и Monodontidae (нарвалы и белухи) содержат мало воска или вообще не содержат его. [7] Скорость звука в дыне наименьшая у Delphinidae, Phocoenidae и Monodontidae, промежуточная у Ziphiidae (клюворылы) и наибольшая у Physeteridae и Platanistidae ( речные дельфины Южной Азии ). [8]
Пилотные киты
[ редактировать ]Дыня гринд (Globicephala) представляет собой смесь эфиров воска и триглицеридов. Внутреннее ядро дыни содержит около 33% эфиров воска, а внешний слой — около 5% эфиров воска. Большая часть жиров является насыщенными. [9]
Карликовый кашалот
[ редактировать ]У карликового кашалота ( Kogia brviceps ) дыня состоит из внешнего слоя и внутреннего ядра, которое обычно содержит большую долю эфиров воска, чем внешний слой. [10]

За дыней находится орган в форме рога изобилия, который многие ученые называют «органом спермацета». Этот орган по форме и составу отличается от спермацетового органа кашалота.
| Внешняя дыня | Внутренняя дыня | Спермацетовый орган | ||
|---|---|---|---|---|
| Содержание липидов (вес) | 15–91% | 74–94% | 92–96% | |
| Липид композиция | Эфиры воска | 8–46% | 40–90% | 84–99% |
| Триглицериды | 54–92% | 10–69% | 1–16% | |
| Среднее количество углерода | Эфиры воска | 32–35 | 29–32 | 28–29 |
| Триглицериды | 47–51 | 41–46 | 45 | |
Кашалот
[ редактировать ]Аналогичную структуру у кашалота традиционно называют «мусором», поскольку китобои сочли ее бесполезной для добычи сперматозоида . Он содержит отсеки спермацета, разделенные стенками хряща. [ нужна ссылка ]
Белуха
[ редактировать ]Дыня белухи уникальна еще и тем, что кит может по своему желанию менять форму дыни. [11] Эти изменения формы, вероятно, приводят к изменению размера, формы, направления и частотного состава эхолокационного луча.
Ссылки
[ редактировать ]- ^ Jump up to: а б с Крэнфорд, Тед В.; Амундин, Матс; Норрис, Кеннет С. (июнь 1996 г.). «Функциональная морфология и гомология носового комплекса зубатых китов: значение для генерации звука». Журнал морфологии . 228 (3): 223–285. doi : 10.1002/(SICI)1097-4687(199606)228:3<223::AID-JMOR1>3.0.CO;2-3 . ПМИД 8622183 . S2CID 35653583 .
- ^ Харпер, CJ; Маклеллан, Вашингтон; Роммель, ЮАР; Гей, ДМ; Дилламан, РМ; Пабст, Д.А. (июль 2008 г.). «Морфология дыни и ее сухожильные соединения с лицевыми мышцами афалин (Tursiops truncatus)» (PDF) . Журнал морфологии . 269 (7): 820–839. дои : 10.1002/jmor.10628 . ПМИД 18473369 . S2CID 206090478 .
- ^ Крэнфорд, Тед В. (октябрь 1999 г.). «Нос кашалота: половой отбор в широком масштабе?». Наука о морских млекопитающих . 15 (4): 1133–1157. дои : 10.1111/j.1748-7692.1999.tb00882.x .
- ^ Биология морских млекопитающих: эволюционный подход . стр. 153
- ^ Крэнфорд, Тед В.; Маккенна, Меган Ф.; Сольдевилья, Мелисса С.; Виггинс, Шон М.; Голдбоген, Джереми А.; Шедвик, Роберт Э.; Крысль, Петр; Сен-Леже, Джуди А.; Хильдебранд, Джон А. (апрель 2008 г.). «Анатомическая геометрия передачи и восприятия звука у клюворыла Кювье ( Ziphius cavirostris )» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . 291 (4): 353–378. дои : 10.1002/ar.20652 . ПМИД 18228579 .
- ^ Такеучи, Хаяте; Мацуиси, Такаши Фриц; Хаякава, Такаши (2024). «Эволюция компромисса между акустическими жировыми телами и мышцами черепа у зубатых китов». Джин . 901 : 148167. doi : 10.1016/j.gene.2024.148167 .
- ^ Личфилд, Картер; Акман, Р.Г.; Сипос, Дж. К.; Итон, Калифорния (сентябрь 1971 г.). «Изовалероилтриглицериды из ворвани и дынного масла белухи ( Delphinapterus leucas )». Липиды . 6 (9): 674–681. дои : 10.1007/BF02531529 . ПМИД 5141491 . S2CID 4023319 .
- ^ Личфилд, Картер; Гринберг, Энн Дж.; Колдуэлл, Дэвид К.; Колдуэлл, Мельба К.; Сипос, Дж. К.; Акман, Р.Г. (апрель 1975 г.). «Сравнительный липидный состав слуховых и неакустических жировых тканей дельфинов, морских свиней и зубатых китов». Сравнительная биохимия и физиология. Часть B: Сравнительная биохимия . 50 (4): 591–597. дои : 10.1016/0305-0491(75)90095-4 . ПМИД 1122741 .
- ^ Ведмид, Юрий; Личфилд, Картер; Акман, Р.Г.; Сипос, Дж. К.; Итон, Калифорния; Митчелл, Эд (декабрь 1973 г.). «Неоднородность липидного состава в ткани головной дыни гринды ( Globicephala melaena )». Biochimica et Biophysical Acta (BBA) - Липиды и липидный обмен . 326 (3): 439–447. дои : 10.1016/0005-2760(73)90144-6 . ПМИД 4776442 .
- ^ Jump up to: а б Кароль, Р.; Личфилд, К.; Колдуэлл, Дания; Колдуэлл, MC (1978). «Композиционная топография липидов органов дыни и спермацета у карликового кашалота Kogia brviceps : значение для эхолокации». Морская биология . 47 (2): 115–123. дои : 10.1007/BF00395632 . S2CID 84443478 .
- ^ «Белуха шевелит дыней в аквариуме Ванкувера» . Ютуб . Архивировано из оригинала 21 декабря 2021 года.