Гомосинаптическая пластичность
Гомосинаптическая пластичность — один из видов синаптической пластичности . [1] Гомосинаптическая пластичность зависит от входного сигнала, то есть изменения в силе синапсов происходят только в постсинаптических мишенях, специально стимулируемых пресинаптической мишенью. [2] Следовательно, распространение сигнала от пресинаптической клетки локализовано.
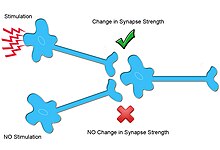
Другой тип синаптической пластичности, гетеросинаптическая пластичность , не является входоспецифичным и по многим механизмам отличается от гомосинаптической пластичности.
Помимо того, что усиление синапса за счет гомосинаптической пластичности зависит от входного сигнала, оно носит ассоциативный характер, поскольку оно зависит от срабатывания пресинаптического и постсинаптического нейрона в близком времени. Эта ассоциативность увеличивает вероятность того, что постсинаптический нейрон также сработает. [3] Предполагается, что эти механизмы лежат в основе обучения и кратковременной памяти . [3]
Обзор
[ редактировать ]Постулат Хебба
[ редактировать ]Дональд Хебб предположил, что усиление синаптических связей происходит из-за скоординированной деятельности между пресинаптическим терминалем и постсинаптическим дендритом. По словам Хебба, эти две клетки укрепляются, потому что их передача сигналов происходит вместе в пространстве и/или времени, что также известно как совпадающая активность. Этот постулат часто резюмируется как « Клетки, которые срабатывают вместе, соединяются вместе ». Это означает, что синапсы, в которых есть нейроны с совпадающим возбуждением, укрепляются, в то время как другие синапсы на этих нейронах остаются неизменными. [3] Постулат Хебба обеспечил концептуальную основу того, как синаптическая пластичность лежит в основе долговременного хранения информации. [1]
Механизмы специфичности входных данных
[ редактировать ]Изменения пластичности часто происходят за счет внедрения или интернализации рецепторов AMPA (AMPAR) в постсинаптическую мембрану синапса, при котором изменяется сила соединения. [1] Что 2+ является одним из сигнальных ионов, который вызывает изменение плотности рецепторов AMPA, вызывая каскад биологических изменений внутри клетки. Чтобы вызвать долговременную потенциацию (LTP), Ca 2+ активирует CAMKII и PKC, вызывая фосфорилирование и вставку AMPAR, в то время как долговременная депрессия (LTD) возникает под действием Ca 2+ активация протеинфосфатаз, которые дефосфорилируют и вызывают интернализацию AMPAR. [1]
Чтобы вызвать специфичные для входа изменения синаптической силы, Ca 2+ сигнал должен быть ограничен конкретными дендритными шипами. Дендритное ограничение Ca 2+ опосредовано несколькими механизмами. Внеклеточный Са 2+ может проникать в позвоночник через NMDA-рецепторы (NMDAR) и потенциалзависимый Ca. 2+ каналы (VGCC). И NMDAR, и VGCC сосредоточены на дендритных шипиках, опосредуя специфичный для шипов Са. 2+ приток. Внутриклеточные запасы Ca 2+ в эндоплазматическом ретикулуме и митохондриях также может способствовать ограничению передачи сигналов в позвоночнике, хотя некоторые исследования не смогли найти доказательств этого. [4] Клиренс Ca 2+ контролируется буферными белками, которые связываются с Ca 2+ и не позволяйте ему просачиваться на другие шипы. Ограниченная диффузия Ca 2+ через шейку дендритного шипа также помогает изолировать его от конкретных дендритов. [4]
Другой механизм долгосрочного потенцирования, специфичного для входных данных, является временным. NMDAR требуют как деполяризации, чтобы снять блокировку магния, так и активации глутамата, чтобы открыть каналы и позволить Ca 2+ приток. Таким образом, LTP локализуется в местах, где каналы NMDA открываются активными синаптическими входами, которые высвобождают глутамат и вызывают деполяризацию постсинаптической клетки, и не влияют на близлежащие неактивные синапсы. [1]
Поддержание долгосрочных изменений
[ редактировать ]Чтобы стабилизировать ДП и продлить ее длительность, в ответ на стимуляцию потенцирующего синапса синтезируются новые белки, поддерживающие это изменение. Возникающая проблема заключается в том, как доставить специфические, вновь синтезированные белки в нужные синапсы, специфичные для ввода, в которых они нужны. Два решения этой проблемы включают синаптическое мечение и локальный синтез белка .
Синаптическая маркировка
[ редактировать ]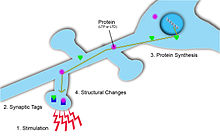
Синаптические метки отмечают, где произошла синаптическая пластичность, и, таким образом, могут предоставить информацию о синаптической силе и потенциале долгосрочных пластических изменений. [5] Метка носит временный характер и включает в себя большое количество белков, активируемых притоком Са. 2+ в постсинаптическую клетку. [5] Кроме того, в зависимости от типа и величины синаптических изменений для мечения используются разные белки. Например, когда пластические изменения приводят к длительной депрессии, кальциневрин применяют . И наоборот, когда пластичность приводит к долговременному потенцированию, CaMKII . используется [5] Чтобы синаптическая пластичность была специфичной для входа, эти синаптические метки необходимы на постсинаптических мишенях, чтобы гарантировать локализацию синаптической потенциации. [5] Эти метки позже инициируют синтез белка, который, в свою очередь, вызовет изменения синаптической пластичности этих активированных нейронов. [1]
Локальный синтез белка
[ редактировать ]Синтез белка на дендритах необходим для гомосинаптической пластичности. Деполяризация и, как следствие, активация рецепторов AMPA и NMDA в постсинаптической клетке вызывает эндоцитоз этих рецепторов. Локальный синтез белка необходим для поддержания количества поверхностных рецепторов в синапсе. Эти новые белки помогают стабилизировать структурные изменения, вызванные гомосинаптической пластичностью. [6] Есть свидетельства наличия в дендритах рибосом, которые могут производить эти белки. Кроме того, есть также свидетельства наличия гранул РНК в дендритах, что указывает на присутствие новообразованных белков. ДП можно индуцировать из дендритов, отделенных от сомы постсинаптического нейрона-мишени. Напротив, LTP может блокироваться в этих дендритах блокаторами синтеза белка, такими как эндомиацин, что дополнительно указывает на место локального синтеза белка. Эти данные показывают, что для стабилизации и поддержания L-LTP необходим локальный синтез белка. [1]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Первс Д., Августин Г.Дж., Фитцпатрик Д., Холл У.К., ЛаМантия А.С., Уайт Л.Е. (2012). Синаптическая пластичность. В неврологии (5-е изд.) (стр. 163–182). Сандерленд, Массачусетс: Sinauer Associates.
- ^ Бирн, Дж. (1997). Синаптическая пластичность. В Neuroscience Online (раздел 1, глава 7).
- ^ Jump up to: а б с Бейли К., Джустетто М., Хуанг Ю., Хокинс Р., Кандел Э. (октябрь 2000 г.). Обзоры: Необходима ли гетеросинаптическая модуляция для стабилизации хеббианской пластичности и памяти? В Macmillan Magazines Ltd (Том 1). Получено с сайта www.nature.com/reviews/neuroscience.
- ^ Jump up to: а б Хигли, М.Дж., Сабатини, Б.Л. (февраль 2012 г.) Передача сигналов кальция в дендритных шипиках. Перспективы Колд-Спринг-Харбор в биологии. Получено с http://cshperspectives.cshlp.org/ . doi:10.1101/cshperspect.a005686.
- ^ Jump up to: а б с д Редондо, Роджер Л. и Ричард Г.М. Моррис. (2011) «Сделаем воспоминания последними: гипотеза синаптической маркировки и захвата». Nature Reviews Neuroscience, 12, 17–30.
- ^ Пфайффер Б.Е., Хубер К.М. (2006). Текущие достижения в области локального синтеза белка и синаптической пластичности. Журнал неврологии, 26(27), 7147-7150.