ПИ-кривая

Кривая PI (или фотосинтез-освещенность ) представляет собой графическое представление эмпирической взаимосвязи между солнечной радиацией и фотосинтезом . Вывод кривой Михаэлиса-Ментен показывает в целом положительную корреляцию между интенсивностью света и скоростью фотосинтеза. Это график зависимости скорости фотосинтеза от интенсивности света (освещенности).
Введение
[ редактировать ]Кривую PI можно применять к наземным и морским реакциям, но чаще всего она используется для объяснения фотосинтетической реакции фитопланктона, обитающего в океане, на изменения интенсивности света. Использование этого инструмента для аппроксимации биологической продуктивности важно, поскольку вклад фитопланктона составляет ~50%. [1] общей глобальной фиксации углерода и являются важными поставщиками морской пищевой сети.
В научном сообществе эту кривую можно назвать PI, PE или кривой светочувствительности. Хотя отдельные исследователи могут иметь свои собственные предпочтения, все они вполне приемлемы для использования в литературе. Независимо от номенклатуры, рассматриваемую скорость фотосинтеза можно описать в терминах количества углерода (C), фиксированного на единицу времени. Поскольку размеры особей различаются, полезно также нормализовать концентрацию углерода по хлорофиллу а (важному фотосинтетическому пигменту) для учета удельной биомассы.
История
[ редактировать ]Еще в 1905 году морские исследователи попытались разработать уравнение, которое можно было бы использовать в качестве стандарта для установления связи между солнечным излучением и фотосинтетической продуктивностью. Несколько групп добились относительного успеха, но в 1976 году сравнительное исследование, проведенное Аланом Джассби и Тревором Платтом, исследователями Бедфордского института океанографии в Дартмуте, Новая Шотландия, пришло к выводу, который укрепил способ построения ПИ-кривой. Оценив восемь наиболее часто используемых уравнений, Яссби и Платт пришли к выводу, что ПИ-кривую лучше всего аппроксимировать гиперболической функцией тангенса, по крайней мере, до тех пор, пока не будет достигнуто фотоингибирование .
Уравнения
[ редактировать ]Есть два простых вывода уравнения, которые обычно используются для построения гиперболической кривой. Первый предполагает, что скорость фотосинтеза увеличивается с увеличением интенсивности света до тех пор, пока не будет достигнута Pmax, и после этого фотосинтез продолжается с максимальной скоростью.
- P = P max [ I ] / ( KI + [ I ])
- P = скорость фотосинтеза при данной интенсивности света
- Обычно обозначается в таких единицах, как (мг C м-3 ч-1) или (мкг C мкг Chl-a-1 ч-1).
- Pmax = максимальная потенциальная скорость фотосинтеза на человека.
- [I] = заданная интенсивность света
- Обычно обозначается в таких единицах, как (мкМоль фотонов м-2 с-1 или (Ватт м-2 ч-1)
- KI = константа полунасыщения; интенсивность света, при которой скорость фотосинтеза составляет ½ Pmax
- Единицы отражают те, которые используются для [I]
- P = скорость фотосинтеза при данной интенсивности света
Как Pmax, так и начальный наклон кривой ΔP/ΔI зависят от вида и зависят от множества факторов, таких как концентрация питательных веществ, температура и физиологические возможности человека. Интенсивность света зависит от широтного положения и подвержена ежедневным и сезонным изменениям, которые также влияют на общую фотосинтетическую способность человека. Эти три параметра предсказуемы и могут использоваться для предопределения общей PI-кривой, которой должна следовать популяция.
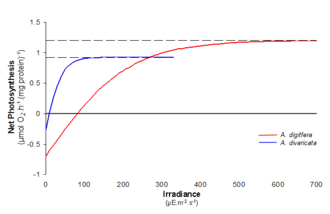
Как видно на графике, два вида могут по-разному реагировать на одни и те же дополнительные изменения интенсивности света. Популяция A (синий цвет) имеет начальную скорость выше, чем популяция B (красный цвет), а также демонстрирует более сильное изменение скорости в сторону увеличения интенсивности света при более низкой освещенности. Следовательно, популяция А будет доминировать в среде с более низкой освещенностью. Хотя популяция B имеет более медленную фотосинтетическую реакцию на увеличение интенсивности света, ее Pmax выше, чем у популяции A. Это позволяет в конечном итоге доминировать в популяции при большей интенсивности света. Существует множество определяющих факторов, влияющих на успех населения; использование PI-кривой для получения прогнозов изменения скорости изменения окружающей среды полезно для мониторинга цветения фитопланктона динамики и стабильности экосистемы.
Второе уравнение объясняет явление фотоингибирования . В верхних нескольких метрах океана фитопланктон может подвергаться воздействию такого уровня радиации, который повреждает хлорофилл — пигмент внутри клетки, что впоследствии снижает скорость фотосинтеза. Кривая реакции изображает фотоингибирование как снижение скорости фотосинтеза при интенсивности света, превышающей ту, которая необходима для достижения Pmax.

Термины, не включенные в приведенное выше уравнение:
- βI = интенсивность света в начале фотоингибирования
- αI = заданная интенсивность света

Примеры
[ редактировать ]Гиперболическая реакция между фотосинтезом и освещенностью, изображаемая ПИ-кривой, важна для оценки динамики популяций фитопланктона, которые влияют на многие аспекты морской среды.
Примечания
[ редактировать ]- ^ Филд и др. 1998 Первичное производство биосферы: интеграция наземных и океанических компонентов, Наука, 10 июля 1998 г.; 281(5374):237-40. doi: 10.1126/science.281.5374.237
Ссылки
[ редактировать ]Эта статья включает список общих ссылок , но в ней отсутствуют достаточные соответствующие встроенные цитаты . ( февраль 2013 г. ) |
- Чалкер Б.Е., Данлэп В.К. и Оливер Дж.К., 1983. Батиметрические адаптации кораллов, образующих рифы, на рифе Дэвис, Большой Барьерный риф, Австралия. II. Кривые светового насыщения для фотосинтеза и дыхания . J Exp Mar Biol Ecol 73:37–87.
- Лалли К.М. и Парсонс Т.Р., 1997. Биологическая океанография: Введение , 2-е изд. Баттерворт-Хайнеманн, Оксфорд.
- Марра Дж., Хайнеманн К. и Ландриау Г. младший, 1985. Наблюденные и предсказанные измерения фотосинтеза в культуре фитопланктона, подвергнутой естественному излучению . Мар Экол. 24:43–50.
- Миллер CB, 2004. Биологическая океанография , Блэквелл.
- Джасби А.Д. и Платт Т., 1976. Математическая формулировка взаимосвязи между фотосинтезом и светом для фитопланктона . Лимнол. Океаногр. 21:540–547.
- Платт Т. и Джасби, А.Д., 1976. Взаимосвязь между фотосинтезом и светом в естественных сообществах прибрежного морского фитопланктона . Дж. Фикол. 12: 421–430.