Поликрикос
| Поликрикос | |
|---|---|
| |
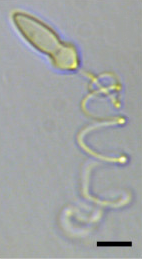
| Светлая микрофотография Polykrikos kofoidii, показывающая выдавленную нематоцисту . Масштабная линейка = 10 мкм. [ 1 ] | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | САР |
| Клэйд : | Альвеолата |
| Тип: | Мизозоа |
| Суперкласс: | Динофлагелляты |
| Сорт: | Динофицеи |
| Заказ: | Гимнодиналес |
| Семья: | Поликриковые |
| Род: | Поликрикос Бючли, 1873 г. |

Британская энциклопедия , 1911 г.

Поликрикос (от греческого «поли» — много и «крикос» — кольцо или круг) — один из родов семейства Polykrikaceae , включающего атекатные псевдоколониеобразующие динофлагелляты. Поликрикосы характеризуются сложной баллистической аппаратурой. [ 2 ] названный комплексом нематоциста-тениоциста, который позволяет видам охотиться на различные организмы. Было обнаружено, что поликрикосы регулируют цветение водорослей, поскольку питаются токсичными динофлагеллятами. [ 3 ] [ 4 ] Однако есть также некоторые данные о том, что Поликрикос токсичен для рыб. [ 5 ]
История знаний
[ редактировать ]Поликрикос был впервые замечен Ульяниным в 1868 году и ошибочно принят за многоклеточную личинку турбеллярий плоских червей. В 1873 году Бутчили повторно исследовал образец и пришел к выводу, что клетка представляла собой необычную инфузорию, а Берг позже, в 1881 году, уточнил родство Polykrikos с динофлагеллятами. [ 6 ]
Морфология
[ редактировать ]Поликрикос — колония зооидов (единиц колониального организма), выполняющих одновременные функции целой клетки.
Все виды Polykrikos имеют: 1) слегка изогнутую продольную борозду, sulcus, идущую к заднему концу организма 2) петлеобразную акробазу, являющуюся передним продолжением борозды 3) поперечную борозду, cingulum, со смещением 4 ) тениоцист-нематоцистных комплексов 5) число ядер в два-четыре раза меньше, чем у зооидов, и 6) способность распадаться на псевдоколонии с меньшим количеством зооидов и только одним ядром. [ 7 ]
Наиболее отличительной чертой этого рода является образование многоядерных псевдоколоний, состоящих из четного числа зооидов. Каждый зооид имеет пару жгутиков (поперечные и продольные жгутики) и имеет собственную поперечную бороздку, цингулум, но продольные борозды зооида, sulci, слиты. [ 8 ] Поперечный жгутик имеет общие для других динофлагеллят латеральные выступы, мастигонемы и исчерченный тяж. [ 9 ] Часто у поликрикосов вдвое меньше ядер, чем у зооидов, и каждая пара зооидов имеет общее ядро. Внутри группы существуют некоторые различия в представлении органелл, но у разных представителей таксона наблюдались трихоцисты, нематоцисты, тениоцисты, мукоцисты и пластиды. [ 8 ]
Цитоплазма Поликрикоса характеризуется многочисленными шероховатыми сетями эндоплазматического ретикулума, комплексами Гольджи и вакуолями. Поликрикосы , как известно, производят выбрасываемые органеллы, экструсомы. Один из них – нематоциста, образующаяся у зооидов. Другой экструсомой, обнаруженной в организме, является палочковидная тениоциста, расположенная дистальнее нематоцисты и ранее ошибочно считавшаяся предшественником нематоцисты. [ 10 ] Вместе эти органеллы образуют комплекс тениоциста-нематоциста, который считается лучшей синапоморфией для клады Поликрикос . [ 8 ] Вакуоли, происходящие из аппарата Гольджи, являются общими для обеих органелл и снабжают каждую молекулами, необходимыми для ее роста, а также участвуют в артикуляции NTC. [ 10 ] Органеллы расположены рядом, но лежат внутри разных мембран и разделены проходом, называемым «желобом». Нематоциста представляет собой более крупную органеллу и лежит позади тениоцисты. Некоторые недавние исследования [ 3 ] показали, что работа двух органелл совмещена: тениоциста прикрепляется к жертве, затем происходит отхождение нематоцисты, приводящее к прокалыванию добычи, и, наконец, извлечение добычи с помощью нити жгута, расположенной на конце нематоцисты вблизи заднего пузырька. Трубочка, встроенная в нематоцисту, разряжается в сторону добычи и, предположительно, используется для ее прокалывания.
Кроме того, Гавелис и др. [ 3 ] глубоко изученная морфология и баллистический механизм NTC, которые, как было показано, фундаментально отличаются от книдарий, продемонстрировали, что нематоцисты развивались независимо в одноклеточных динофлагеллятах. Оболочка извитой канальцевой капсулы, в отличие от книдарий, герметична, что заставляет стилет при выстреле сначала проколоть капсулу изнутри, чтобы освободить нить, и только потом проткнуть добычу. Когда трубочка проходит через сопло, она открывает крышку и после этого разворачивается. Баллистика в нематоцистах книдарий обусловлена синтезом осмотической полигамма-глутаматсинтазы, PgsAA, тогда как у Polykrikos считается, что она возникает из-за индуцированного давления в результате сокращения капсульных волокон в стенке капсулы. [ 3 ] Ядро уникально характеризуется двухслойной фиброзной корой, которая подчеркивает выпяченную ядерную оболочку; Предполагается, что кора обеспечивает прочность и форму ядра, в то время как считается, что выпячивание ядра увеличивает площадь ядерно-цитоплазматического обмена в местах перфорации коры. [ 9 ]
У Polykrikos четко выраженные фиброзные ленты участвуют в ядерно-жгутиковых соединениях, и прикрепление к жгутиковому аппарату может служить для ориентации ядра относительно жгутиков во время процессов движения, митоза и деления клеток. [ 9 ]
Пластиды
[ редактировать ]существуют различия в усвоении питательных веществ Среди видов Polykrikos : некоторые полагаются исключительно на фотосинтез, некоторые являются миксотрофами, а некоторые являются облигатными гетеротрофами, что делает Polykrikos полезной группой для изучения эволюции органелл. Ранневетвящиеся поликрикоиды, Polykrikos geminatum и P. hartmanii , имеют трехмембранные пластиды с тройными стопками тилакоидов, что указывает на вторичные пластиды перидининового типа, характерные для динофлагеллят. Однако миксотрофный P. lebouriae , филогенетически гнездящийся среди гетеротропных поликрикоидов, имеет пластиды, нетипичные для динофлагеллят. Он имеет две мембраны и содержит двойные тилакоиды, которые встречаются у диатомовых водорослей и гаптофитов. Однако анализ молекулярных данных, проведенный Gavelis et al. [ 11 ] продемонстрировали, что P. lebouriae имеют пластиды перидининового типа, которые, скорее всего, были приобретены от предковых поликрикоидов. Транскриптомный анализ продемонстрировал множественные события потери у поликрикоидов, которые можно объяснить с точки зрения энергетики и физиологических ограничений. Постепенное увеличение размера Polykrikos делает эти организмы большими хищниками, но известно, что увеличение размера тела отдельных клеток приводит к усилению самозатенения хлоропластов и уменьшению соотношения площади поверхности к объему, что приводит к снижению эффективности фотосинтеза. Наличие специализированных NTC и большой размер клеток могли спровоцировать множественные потери фотосинтеза. [ 11 ]
Жизненный цикл P. kofoidii
[ редактировать ]Для рода Polykrikos доступны подробные данные о воспроизводстве типового вида (голотипа) P. kofoidii , жизненный цикл которого напоминает общий цикл динофлагеллят, поскольку вегетативные клетки образуют гаметы, которые сливаются с образованием диплоидной (2n) зиготы, которая может инцистировать, но ряд особенностей псевдоколониальный характер придает развитию Поликрика . [ 12 ]
При сытом питании они появлялись в виде 4-зооид-2-ядерных псевдоколоний, а при вегетативном размножении количество зооидов удваивалось с последующим делением ядер, приводящим к стадии 8-зооид-4-ядра с дальнейшим поперечным бинарным делением на два 4-ядра. зооид-2-ядра Поликрикос . Формирование гамет было особенным, поскольку псевдоколония производит 4 гаметы разного размера и морфологии, чем вегетативные клетки. Вегетативная форма удвоила зооиды и впоследствии разделилась на четыре гаметы формы 2-зооид-1-ядро. Две гаметы далее соединились в пары своими вентральными сторонами и слились, образуя плоскозиготу. У P. kofoidii в гаметах наблюдались две копуляционные пальцеобразные структуры, которые предположительно участвуют в контакте и слиянии гамет, но для подтверждения этого необходимы дополнительные данные. Вентрально слитые гаметы потребовали сложной перестройки восьми жгутиков и образования борозд и поясных извилин. 4-зооидная планозигота имела только одно ядро и имела два пути развития в зависимости от наличия пищи. В условиях голодания планозигота распалась на два 2-зооида, у одного из которых отсутствовало ядро, но дальнейшая судьба не рассматривалась. В условиях культивирования большинство организмов подвергалось мейозу и непосредственно вступало в вегетативный цикл. Очень немногие планозиготы прошли стадию покоящейся кисты. Стадия кисты сохранялась в течение 1 мес, что считается относительно коротким периодом по сравнению с другими динофлагеллятами, у которых период облигатного покоя может достигать 6 мес. Продолжительность инцистирования связана с экологическими стратегиями поиска пищи. Короткий период покоя может способствовать быстрому циклическому переходу между стадиями жизненного цикла, что может быть полезно для гетеротрофных видов в случае колебаний доступности пищи. Однако возможность химической передачи сигналов, участвующей в вылуплении кисты, для Поликрикос еще не определен.
Зародыш, единственная зооидная клетка, вышедшая из кисты, имела уникальное развитие, которое никогда не было зарегистрировано ни у одной свободноживущей динофлагелляты. Его морфология явно перешла от 1-зооид-1-ядра, через 2-зооид-1-ядро и 4-зооид-1-ядро к стадии 4-зооид-2-ядра. Эти данные поднимают вопрос, может ли такая схема штриховки отражать филогению псевдоколоний Polykrikos . [ 12 ]
Среда обитания и экология (ниша, пища)
[ редактировать ]Все известные виды Polykrikos встречаются в морской среде. Большинство видов являются планктонными, за исключением Polykrikos lebouriae и Polykrikos herdmanae , обитающих в донной среде обитания. [ 7 ] Существуют также различия в экологии питания, поскольку некоторые виды имеют пластиды и могут использовать фотосинтез для получения питательных веществ, но часто оказываются миксотрофами ( P. barnegatensis, P. lebouriae, P tanit, P. hartmannii ). Известно только один вид исключительно автотрофный ( P. geminatum ), тогда как некоторые лишены пластиды и являются полностью гетеротрофными ( P.grassei, P.herdmanae, P.kofoidii и P.schwartzii ). [ 13 ] [ 8 ]
Polykrikos hartmannii — фототрофная динофлагеллята, обнаруженная в водах Канады, [ 14 ] США, Мексика, Китай, Индия, Япония, Корея. [ 5 ] [ 13 ] P. hartmanii питается видами водорослей путем поглощения после закрепления добычи с помощью комплекса нематоциста-тениоциста (позже названного NTC). [ 13 ] Тан и др. [ 5 ] наблюдал цветение P. hartmannii , которое вызвало 100% смертность молоди овчарки ( Cyprinodon variegates ) в течение 24 часов, что позволяет предположить, что P. hartmannii является ихтиотоксичной и вредной водорослью. P.hartmannii питается образующими цепочки динофлагеллятами Cochlodinium polykrikoides и Gymnodinium catenatum , которые, как известно, также вызывают смертность рыб, поэтому P. hartmannii обладает ферментами, которые детоксифицируют токсины, вырабатываемые этими динофлагеллятами-жертвами. считается, что [ 13 ] Кроме того, сравнение питания трех видов Polykrikos показало, что виды различаются по предпочтениям добычи, а некоторые из них более специализированы, чем другие, так что охота P. hartmanii менее разнообразна (питаются двумя видами добычи), чем P. kofoidii и P. hartmanii. lebouriae , питавшиеся 14 различными видами водорослей. [ 13 ]
Хищничество гетеротрофных поликрикосов стало большой темой интереса, поскольку некоторые организмы поедают динофлагелляты, вызывающие токсичное цветение. сильном хищническом воздействии Polykrikos schwartzii Butschili на токсичную динофлагелляту Alexandrium tamarense Сообщалось о (Lebour) Balech в Аргентине, в то время как Polykrikos kofoidii Chatton контролировал Gymnodium catenatum Graham в прибрежных водах Португалии и Японии. [ 4 ] G. catenatum является одним из видов, вызывающих паралитическое отравление моллюсками (PSP), и встречается в водах Австралии, Японии, Мексики и Испании. [ 15 ] Наблюдения за пищевым поведением позволяют предположить, что P. kofoidii первоначально демонстрировал петлевое плавательное поведение в непосредственной близости от добычи, за которым следовал выброс нематоцисты, втягивание добычи в тело через заднюю борозду и окончательное поглощение добычи. [ 16 ] PSP вызывает социально-экономические и экологические проблемы, поскольку влияет на здоровье как морских млекопитающих, так и человека, а механизм регулирования популяции токсичных микроводорослей P.kofoidii может иметь важное значение для экологического мониторинга и устранения опасности для здоровья.
Практическая значимость
[ редактировать ]Хищничество гетеротрофных динофлагеллят в отношении токсичных микроводорослей является одним из факторов, контролирующих цветение водорослей. Поликрикос , как известно, модулирует популяции динофлагеллят, таких как Alexandrium tamarense . [ 3 ] [ 4 ] и G. catenatum, [ 17 ] которые являются одними из распространенных возбудителей токсичного цветения водорослей. Такие гетеротрофные поликрикоиды могут не только снизить уровень токсичности, вызываемой их добычей в морских пищевых сетях, но и остановить цветение токсичности и могут быть использованы в биоремедиации. [ 17 ] Таким образом, снижение токсичности воды может помочь регулировать баланс морских пищевых сетей и снизить уровень смертности рыб, морских млекопитающих и морских птиц. Дальнейшие исследования молекулярных механизмов детоксикации Поликрикосом могут быть полезны в области биомедицины и мониторинга окружающей среды.
Тем не менее, некоторые Polykrikos представляют риск для здоровья некоторых рыб, в то время как те, которые регулируют цветение, часто становятся жертвами морских беспозвоночных, таких как амфиподы , которые возвращают токсины обратно в пищевую сеть. [ 17 ] Некоторый мониторинг популяции поликрикоидов и исследование динамики токсинов внутри тела травоядных могут обеспечить лучшее понимание планктонных пищевых сетей, оценить степень отравления в экосистемах и предложить потенциальные пути выведения токсинов.
Список видов
[ редактировать ]В настоящее время таксономически приняты следующие 10 видов: [ 18 ]
- Поликрикос барнегатенсис ГВМартин,
- Polycricos geminatus (F.Schütt) DXQiu и Senjie Lin,
- Поликрикос грассей Лекаль,
- Поликрикос Хартманни В.Циммерманн,
- Polykrikos herdmaniae Hoppenrath & Leander,
- Поликрикос кофоидии Чаттон,
- Поликрикос лебуриэ Хердман,
- Поликрикос Шварци Бючли,
- Поликрикос Танит Ренье,
- Polykrikos tentaculatus О.Ветцель
Ссылки
[ редактировать ]- ^ Хоппенрат, Мона; Бачваров, Цветан Р; Хэнди, Сара М; Делвич, Чарльз Ф; Леандер, Брайан С. (2009). «Молекулярная филогения динофлагеллят, несущих оцеллоид (Warnowiaceae), как следует из последовательностей рДНК SSU и LSU» . Эволюционная биология BMC . 9 : 116. Бибкод : 2009BMCEE...9..116H . дои : 10.1186/1471-2148-9-116 . ПМК 2694157 . ПМИД 19467154 .
- ^ Джеймс Горман (31 марта 2017 г.). «Первое четкое представление об одноклеточном гарпунере в действии» . NYTimes.com . Проверено 26 декабря 2017 г.
- ^ Перейти обратно: а б с д и Гавелис, Г.; Уэйкман, К.; Рипкен, К.; Озебек, С.; Гольштейн, Т.; Херранц, М.; Килинг, П.; Леандер, Б. (2017). «Гонка микробных вооружений: баллистические «нематоцисты» в динофлагеллятах представляют собой новую крайность в сложности органелл» . Достижения науки . 3 (3): e1602552. Бибкод : 2017SciA....3E2552G . дои : 10.1126/sciadv.1602552 . ПМЦ 5375639 . ПМИД 28435864 .
- ^ Перейти обратно: а б с Нагай, С.; Мацуяма, Ю.; Такаяма, Х.; Котани, Ю. (2002). «Морфология цист Polykrikos kofoidii и P. schwartzii (Dinophyceae, Polykrikaceae), полученных в культуре». Психология 41 (4): 319. Бибкод : 2002Phyco..41..319N . дои : 10.2216/i0031-8884-41-4-319.1 . S2CID 83728508 .
- ^ Перейти обратно: а б с Тан, ЮЗ; Харк, MJ; Гоблер, CJ (2013). «Морфология, филогения, динамика и ихтиотоксичность изолятов и цветков Pheopolykrikos hartmannii (Dinophyceae) из Нью-Йорка, США». Дж. Фикол . 49 (6): 1084–1094. Бибкод : 2013JPcgy..49.1084T . дои : 10.1111/jpy.12114 . ПМИД 27007629 . S2CID 12140986 .
- ^ Тейлор, FJR (1987). «Биология динофлагеллят». Биологические монографии . 21 .
- ^ Перейти обратно: а б Хоппенрат, М.; Леандер, Б.С. (2007b). «Морфология и филогения псевдоколониальных динофлагеллят Polykrikos lebourae и Polykrikos herdmanae n. sp». Протист . 158 (2): 209–227. дои : 10.1016/j.protis.2006.12.001 . ПМИД 17241813 .
- ^ Перейти обратно: а б с д Хоппенрат, М.; Леандер, Б.С. (2007a). «Эволюция характера поликрикоидных динофлагеллят». Дж. Фикол . 43 (2): 366–377. Бибкод : 2007JPcgy..43..366H . дои : 10.1111/j.1529-8817.2007.00319.x . S2CID 16821791 .
- ^ Перейти обратно: а б с Брэдбери, ПК; Вестфолл, Дж.А.; Таунсенд, Дж. (1983). «Ультраструктура динофлагелляты Polykrikos . Ядро и его связь с жгутиковым аппаратом». Журнал исследований ультраструктуры . 85 (1): 24–32. дои : 10.1016/s0022-5320(83)90113-2 . ПМИД 6686618 .
- ^ Перейти обратно: а б Вестфолл, Дж.А.; Брэдбери, ПК; Таунсенд, Дж. (1983). «Ультраструктура динофлагелляты Поликрикос . Развитие комплекса нематоциста-тениоциста и морфология места экструзии». J Cell Sci . 63 : 245–261. дои : 10.1242/jcs.63.1.245 . ПМИД 6685130 .
- ^ Перейти обратно: а б Гавелис, Г.; Уайт, РА; Саттл, Калифорния; Килинг, П.Дж.; Леандер, Б.С. (2015). «Одноклеточная транскриптомика с использованием сплайсированной лидерной ПЦР: доказательства множественных потерь фотосинтеза у поликрикоидных динофлагеллят» . БМК Геномика . 16 (1): 528. doi : 10.1186/s12864-015-1636-8 . ПМК 4504456 . ПМИД 26183220 .
- ^ Перейти обратно: а б Тильманн, У.; Хоппенрат, М. (2013). «Жизненный цикл псевдоколониальной динофлагелляты Polykrikos kofoidii (Gymnodiniales, Dinoflagellata)». Журнал психологии . 49 (2): 298–317. Бибкод : 2013JPcgy..49..298T . дои : 10.1111/jpy.12037 . ПМИД 27008517 . S2CID 30674349 .
- ^ Перейти обратно: а б с д и Ли, MJ; Чон, HJ; Ли, К.Х.; Джанг, С.Х.; Ким, Дж. Х.; Ким, Кентукки (2015). , несущей комплекс нематоциста-тениоциста «Миксотрофия фототрофной динофлагелляты Polykrikos hartmannii ». Вредные водоросли . 49 : 124–132. дои : 10.1016/j.hal.2015.08.006 .
- ^ Хоппенрат, М.; Юбуки, Н.; Бачваров, С.; Леандер, Б.С. (2010). «Реклассификация Pheopolykrikos hartmannii в Polykrikos (Dinophyceae) частично на основе ультраструктуры сложных экструсом». Евро. Ж. Протистол . 46 (1): 29–37. дои : 10.1016/j.ejop.2009.08.003 . ПМИД 19767184 .
- ^ Мацуяма, Ю; Миямото, М; Котани, Ю. (1999). «Выпасное воздействие гетеротрофной динофлагелляты Polykrikos kofoidii на цветение Gymnodinium catenatum » . Акват Микроб Экол . 17 : 91–98. дои : 10.3354/man017091 .
- ^ Мацуока, К.; Чо, HJ; Джейкобсон, DM (2000). «Наблюдение за пищевым поведением и скоростью роста гетеротрофной динофлагелляты Polykrikos kofoidii (Polykrikaceae. Dinophyceae) 39: 82-86». Психология . 39 : 82–86. дои : 10.2216/i0031-8884-39-1-82.1 . S2CID 86293920 .
- ^ Перейти обратно: а б с Чон, Х.; Парк, К.; Ким, Дж.; Канг, Х.; Ким, К.; Чой, Х.,...; Парк, М. (2003). «Снижение токсичности динофлагелляты Gymnodinium catenatum при питании гетеротрофной динофлагеллятой Polykrikos kofoidii » . Водная микробная экология . 31 : 307–312. дои : 10.3354/ame031307 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гири, доктор медицинских наук «Поликрикос Бучли, 1873» . Основа из водорослей . Проверено 20 февраля 2017 г.