Полуконсервативная репликация
Полуконсервативная репликация описывает механизм репликации ДНК во всех известных клетках. Репликация ДНК происходит в нескольких точках начала репликации вдоль цепей матрицы ДНК. Поскольку двойная спираль ДНК раскручивается хеликазой , репликация происходит отдельно на каждой цепи матрицы в антипараллельных направлениях. Этот процесс известен как полуконсервативная репликация, поскольку создаются две копии исходной молекулы ДНК, каждая копия сохраняет (реплицирует) информацию из половины исходной молекулы ДНК. [1] [2] Каждая копия содержит одну исходную цепь и одну вновь синтезированную цепь. (Обе копии должны быть идентичными, но это не совсем гарантировано.) Структура ДНК (расшифрованная Джеймсом Д. Уотсоном и Фрэнсисом Криком в 1953 году) предполагает, что каждая нить двойной спирали может служить матрицей для синтеза ДНК. новая прядь. Неизвестно, как вновь синтезированные цепи объединяются с нитями матрицы, образуя две двухспиральные молекулы ДНК. [3] [4]
Открытие [ править ]
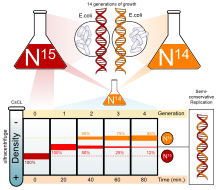
Было проведено множество экспериментов, чтобы определить, как реплицируется ДНК. Полуконсервативная модель была предвосхищена Николаем Кольцовым и позже поддержана экспериментом Мезельсона-Шталя . [4] [5] который подтвердил, что ДНК реплицируется полуконсервативно, проведя эксперимент с использованием двух изотопов : азота-15 ( 15
Н
) и азот-14 ( 14
Н
). Когда 14
Н
был добавлен к тяжелому 15
Н
- 15
Н
ДНК, гибрид 15
Н
- 14
Н
был замечен в первом поколении. После второго поколения гибрид остался, но легкая ДНК( 14
Н
- 14
Н
) тоже было замечено. Это указывало на то, что ДНК реплицируется полуконсервативно. Этот режим репликации ДНК позволял каждой дочерней цепи оставаться связанной со своей матричной цепью. [6]
Природа репликации ДНК
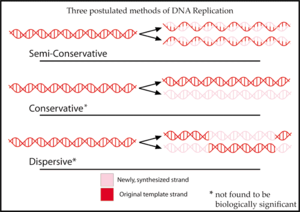
Полуконсервативная репликация получила свое название от того факта, что этот механизм транскрипции был одной из трех первоначально предложенных моделей. [3] [4] для репликации ДНК :
- Полуконсервативная репликация привела бы к образованию двух копий, каждая из которых содержала бы одну исходную цепь ДНК и одну новую цепь. [3] Полуконсервативная репликация полезна для восстановления ДНК. Во время репликации новая цепь ДНК приспосабливается к изменениям, внесенным в цепь матрицы. [7]
- Консервативная репликация оставит две исходные цепи ДНК-матрицы вместе в двойной спирали и создаст копию, состоящую из двух новых цепей, содержащих все новые пары оснований ДНК. [3]
- Дисперсионная репликация приведет к образованию двух копий ДНК, каждая из которых будет содержать отдельные участки ДНК, состоящие либо из обеих исходных цепей, либо из обеих новых цепей. [3] Первоначально считалось, что нити ДНК разрываются на каждой десятой паре оснований, чтобы добавить новую матрицу ДНК. В конце концов, после многих поколений репликации вся новая ДНК составит двойную спираль. [8]
Разделение и рекомбинация двухцепочечной ДНК [ править ]
Для осуществления полуконсервативной репликации двойная спираль ДНК должна быть разделена, чтобы новая цепь матрицы могла быть связана с комплементарными парами оснований. Топоизомераза — это фермент, который способствует расстегиванию и рекомбинации двойной спирали. В частности, топоизомераза предотвращает сверхспираль или слишком тугое закручивание двойной спирали. В этом процессе участвуют три фермента топоизомеразы: топоизомераза типа IA , топоизомераза типа IB и топоизомераза типа II . [9] Топоизомераза типа I раскручивает двухцепочечную ДНК, а топоизомераза типа II разрывает водородные связи, соединяющие комплементарные пары оснований ДНК. [8]
Скорость и точность [ править ]
Скорость полуконсервативной репликации ДНК в живой клетке впервые была измерена как скорость удлинения цепи ДНК фага Т4 в инфицированной фагом E. coli . [10] В период экспоненциального роста ДНК при 37 °С скорость удлинения цепи составляла 749 нуклеотидов в секунду. Частота мутаций на пару оснований за раунд репликации во время синтеза ДНК фага Т4 составляет 2,4 × 10. −8 . [11] Таким образом, полуконсервативная репликация ДНК является одновременно быстрой и точной.
Приложения [ править ]
Полуконсервативная репликация дает ДНК множество преимуществ. Он быстрый, точный и позволяет легко восстанавливать ДНК. Он также отвечает за фенотипическое разнообразие у некоторых видов прокариот. [12] Процесс создания вновь синтезированной цепи из шаблонной цепи позволяет метилировать старую цепь в отдельное время от новой цепи. Это позволяет ферментам репарации корректировать новую цепь и исправлять любые мутации или ошибки. [7]
ДНК может обладать способностью активировать или деактивировать определенные области вновь синтезированной цепи, что позволяет фенотип изменить клетки. Это может быть выгодно для клетки, поскольку ДНК может активировать более благоприятный фенотип, способствующий выживанию. Благодаря естественному отбору более благоприятный фенотип сохранится у всего вида. Это порождает идею наследственности или того, почему одни фенотипы наследуются больше, чем другие. [7]
См. также [ править ]
- Молекулярная структура нуклеиновых кислот: структура нуклеиновой кислоты дезоксирибозы
- репликация ДНК
Ссылки [ править ]
- ^ Экундайо Б., Блейхерт Ф. (сентябрь 2019 г.). «Истоки репликации ДНК» . ПЛОС Генетика . 15 (9): e1008320. дои : 10.1371/journal.pgen.1008320 . ПМЦ 6742236 . ПМИД 31513569 .
- ^ Молитесь, Лесли А. «Полуконсервативная репликация ДНК: Мезельсон и Шталь». Природное образование . 1(1):98.
- ^ Jump up to: Перейти обратно: а б с д и Гриффитс А.Дж., Миллер Дж.Х., Сузуки Д.Т., Левонтин Р.К., Гелбарт В.М. (1999). «Глава 8: Структура и репликация ДНК» . Введение в генетический анализ . Сан-Франциско: WH Freeman. ISBN 978-0-7167-3520-5 .
- ^ Jump up to: Перейти обратно: а б с Мезельсон М., Шталь Ф.В. (июль 1958 г.). «Репликация ДНК в Escherichia Coli» . Труды Национальной академии наук Соединенных Штатов Америки . 44 (7): 671–82. Бибкод : 1958ПНАС...44..671М . дои : 10.1073/pnas.44.7.671 . ПМК 528642 . ПМИД 16590258 .
- ^ Мезельсон М., Шталь Ф.В. (2007). «Демонстрация полуконсервативного режима дупликации ДНК». В Кэрнс Дж., Стент Г.С., Уотсон Дж.Д. (ред.). Фаг и истоки молекулярной биологии . Колд-Спринг-Харбор: Лабораторный пресс Колд-Спринг-Харбор. ISBN 978-0-87969-800-3 .
- ^ ПК Ханавальт (декабрь 2004 г.). «Плотность имеет значение: полуконсервативная репликация ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (52): 17889–94. дои : 10.1073/pnas.0407539101 . ПМК 539797 . ПМИД 15608066 .
- ^ Jump up to: Перейти обратно: а б с Норрис V (июнь 2019 г.). «Способствует ли полуконсервативный характер репликации ДНК согласованному фенотипическому разнообразию?» . Журнал бактериологии . 201 (12). дои : 10.1128/jb.00119-19 . ПМК 6531617 . ПМИД 30936370 .
- ^ Jump up to: Перейти обратно: а б Уотсон Дж.Д., Ганн А., Бейкер Т.А., Левин М., Белл С.П., Лосик Р. (2014). Молекулярная биология гена (Седьмое изд.). Бостон. ISBN 978-0-321-76243-6 . OCLC 824087979 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Браун Т.А. (2002). «Репликация генома» . Геномы (2-е изд.). Вили-Лисс.
- ^ Маккарти Д., Миннер С., Бернштейн Х., Бернштейн С. (октябрь 1976 г.). «Скорость элонгации ДНК и распределение точек роста фага Т4 дикого типа и янтарного мутанта с задержкой ДНК». Журнал молекулярной биологии . 106 (4): 963–81. дои : 10.1016/0022-2836(76)90346-6 . ПМИД 789903 .
- ^ Дрейк Дж.В., Чарльзуорт Б., Чарльзуорт Д., Кроу Дж.Ф. (апрель 1998 г.). «Темпы спонтанных мутаций» . Генетика . 148 (4): 1667–86. дои : 10.1093/генетика/148.4.1667 . ПМК 1460098 . ПМИД 9560386 .
- ^ Маккарти Д., Миннер С., Бернштейн Х., Бернштейн С. (октябрь 1976 г.). «Скорость элонгации ДНК и распределение точек роста фага Т4 дикого типа и янтарного мутанта с задержкой ДНК». Журнал молекулярной биологии . 106 (4): 963–81. дои : 10.1016/0022-2836(76)90346-6 . ПМИД 789903 .