Расширение зародышевой полосы
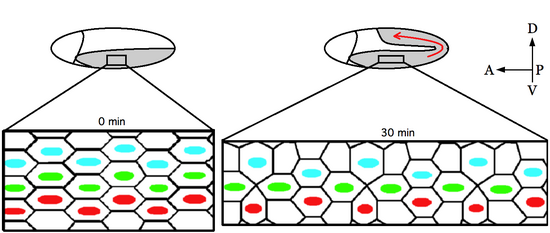
Расширение зародышевого листка — это морфогенный процесс, широко изучаемый в развитии Drosophila melanogaster, при котором зародышевый клубок , развивающийся в сегментированный туловище зародыша, удваивается в длине вдоль передне-задней оси, а затем сужается вдоль дорсально-задней оси. вентральная ось. [ 1 ]
Расширение зародышевого круга начинается вскоре после гаструляции и делится на две фазы. Быстрая фаза, во время которой происходит большая часть растяжения, занимает около 25 минут. Оставшееся расширение продолжается во время медленной фазы и завершается в течение следующих 70 минут. [ 2 ] Во время этого процесса вентральная зародышевая полоска распространяется вокруг заднего конца зародыша, эффективно складываясь на дорсальную сторону яйца. Множественные отдельные клетки, интеркалирующие медиолатерально к передне-задней оси, приводят к глобальному удлинению эмбриона. Кроме того, для полного удлинения частично необходимы изменения формы клеток и ориентированные деления клеток в заднем зародышевом диске. [ 3 ] Однако удлинение оси тела, по-видимому, в первую очередь связано с изменениями в отношениях соседства клеток.
В этой статье описывается удлинение оси у дрозофилы . Однако основы удлинения зародышевых полосок применимы ко многим организмам, включая других беспозвоночных и позвоночных . [ 4 ]
Клеточная основа
[ редактировать ]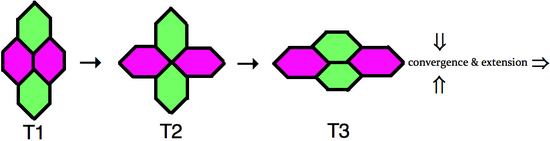
Для того чтобы клетки могли интеркалировать друг с другом, адгезионные соединения , поддерживающие целостность эпителиальной ткани, должны динамически ремоделироваться. Покадровая микроскопия зафиксировала этот процесс обмена соседями клеток, который схематически представлен справа. В конфигурации типа 1 две клетки контактируют друг с другом вдоль передне-задней оси, тогда как две дорсо-вентральные клетки не контактируют напрямую. Затем граница клеток между двумя передне-задними соседями выборочно сужается, что приводит к обязательному промежуточному соединению типа 2, где четыре клетки имеют общую вершину. После разрешения соединения типа 2 образуется новое соединение типа 3, перпендикулярное исходной конфигурации типа 1. В ходе этого процесса две дорсо-вентральные клетки стали соседями. [ 5 ] Когда множественные кластеры клеток интеркалируют в дорсально-вентральной оси посредством обмена соседними узлами, результатом является расширение зародышевого круга в передне-задней оси.
Помимо простого обмена соседями, наблюдались розеточные образования более высокого порядка, в которых пять или более ячеек встречаются в вершине. Многоклеточные розетки формируются и разрешаются направленным образом, что способствует удлинению зародышевых полосок. [ 6 ] Обмен соседями и образование многоклеточных розеток включают ориентированное ремоделирование соединений, что указывает на то, что интеркалирующие клетки внутренне поляризованы в плоскости эпителия.
Молекулярная основа
[ редактировать ]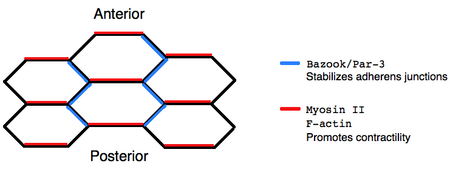
Дорсально-вентральная интеркаляция клеток во время расширения зародышевых полосок в конечном итоге возникает из-за асимметричной локализации белков внутри отдельных клеток. Иммунофлуоресценция показывает, что немышечный миозин II локализуется на передне-задних границах клеток, дестабилизируя слипчивые соединения, тогда как комплекс Базука/Par-3 локализуется на дорсально-вентральных границах, стабилизируя слипчивые соединения. Более того, мутанты базуки дефектны в расширении зародышевых полос, что подтверждает идею о том, что локализация поляризованного белка имеет решающее значение для клеточных перестроек. [ 7 ] Одним из механизмов, с помощью которого миозин II может способствовать ремоделированию поляризованных клеток, является сократительная активность, которая создает ориентирующую напряжение разборку соединений. [ 8 ] Однако точный механизм, при котором асимметрично локализованные белковые комплексы способствуют направленной интеркаляции, остается спорным.
Источник, который устанавливает плоскую полярность во время расширения зародышевых полосок, остается неуловимым. Поляризованная интеркаляция практически не затрагивается у мутантных эмбрионов, у которых отсутствуют типы дорсально-вентральных клеток. Тем не менее, мутации, которые нарушают формирование сегментного паттерна вдоль передне-задней оси, такие как eve и runt , уменьшают интеркаляцию клеток и последующее удлинение зародышевых полосок. [ 1 ] Более того, эктопическая экспрессия eve или runt достаточна для локальной переориентации полярности близлежащих клеток. Эти данные доказывают, что плоская полярность устанавливается межклеточными взаимодействиями, а не дальнодействующим поляризационным сигналом . Таким образом, поляризующая информация может распространяться от одной клетки к другой, следуя за зависимым от Евы сигналом, который еще предстоит идентифицировать. [ 7 ]
Растягивающие силы
[ редактировать ]Исследователи предполагают, что мезодермы инвагинация обеспечивает передне-заднюю тянущую силу, которая способствует расширению зародышевых полосок за счет пассивных изменений формы клеток. Хотя мутанты с передне-задним паттерном не могут полностью удлинить свои зародышевые полосы, во время быстрой фазы длина удлинения нормальна, несмотря на дефекты в интеркаляции поляризованных клеток. Промежуточный анализ показал, что увеличение растяжения формы клеток по передне-задней оси компенсирует аберрантную интеркаляцию клеток, независимо от формирования передне-заднего паттерна. Более того, во время развития дикого типа было высказано предположение, что поведение интеркалярных клеток ослабляет стресс, налагаемый на зародышевый диск, позволяя растянутым клеткам восстанавливаться до изометрической формы. [ 3 ]
Кроме того, имеются данные о том, что механическое напряжение необходимо и достаточно для кортикальной локализации миозина II. Таким образом, миозин II может не только генерировать напряжение, но также может регулироваться силами растяжения, создавая петлю положительной обратной связи , которая позволяет клеткам динамически реагировать на колебания в их механической среде. [ 9 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Ирвин, К.; Вишаус, Э. (1994). «Интеркаляция клеток во время дрозофилы расширения зародышевых полосок и ее регуляция с помощью генов сегментации по парному правилу» . Разработка . 120 (4): 827–841. дои : 10.1242/dev.120.4.827 . ПМИД 7600960 .
- ^ да Силва, С.М.; Винсент, Ж.-П. (2007). «Ориентированное деление клеток в расширяющемся зародыше дрозофилы » . Разработка . 134 (17): 3049–3054. дои : 10.1242/dev.004911 . ПМИД 17652351 .
- ^ Перейти обратно: а б Батлер, LC; Бланшар, Великобритания; Кабла, Эй Джей; Лоуренс, Нью-Джерси; Уэлчман, ДП; Махадеван, Л.; Адамс, Р.Дж.; Сансон, Б. (2009). «Изменения формы клеток указывают на роль внешних сил растяжения в дрозофилы расширении зародышевых групп ». Природная клеточная биология . 11 (7): 859–864. дои : 10.1038/ncb1894 . ПМИД 19503074 .
- ^ Келлер, Р. (2006). «Механизмы элонгации в эмбриогенезе» . Разработка . 133 (12): 2291–2302. дои : 10.1242/dev.02406 . ПМИД 16720874 .
- ^ Берте, К.; Сулак, Л.; Лекуит, Т. (2004). «Ремоделирование миозин-зависимого соединения контролирует интеркаляцию плоских клеток и удлинение оси». Природа . 429 (6992): 667–671. дои : 10.1038/nature02590 . ПМИД 15190355 .
- ^ Бланкеншип, Дж. Т.; Бакович, СТ; Санни, Дж. С.; Вайц, О.; Заллен, Дж. А. (2006). «Образование многоклеточных розеток связывает полярность плоских клеток с морфогенезом тканей» . Развивающая клетка . 11 (4): 459–470. дои : 10.1016/j.devcel.2006.09.007 . ПМИД 17011486 .
- ^ Перейти обратно: а б Заллен, Дж.А.; Вишаус, Э. (2004). «Узорчатая экспрессия генов управляет биполярной планарной полярностью у дрозофилы » . Развивающая клетка . 6 (3): 343–355. дои : 10.1016/s1534-5807(04)00060-7 . ПМИД 15030758 .
- ^ Баум, Б. (2004). «Развитие животных: борьба с толпой» . Современная биология . 14 (17): Р716–Р718. дои : 10.1016/j.cub.2004.08.047 . ПМИД 15341764 .
- ^ Фернандес-Гонсалес Р.; де Матос Симоэс, С.; Репер Ж.-К.; Итон, С.; Заллен, Дж. А. (2009). «Динамика миозина II регулируется напряжением в интеркалирующих клетках» . Развивающая клетка . 17 (5): 736–743. дои : 10.1016/j.devcel.2009.09.003 . ПМЦ 2854079 . ПМИД 19879198 .