Гипотеза адаптера

Гипотеза адаптера — это теоретическая схема в молекулярной биологии, объясняющая, как информация, закодированная в последовательностях нуклеиновых кислот информационной РНК (мРНК), используется для определения аминокислот, входящих в состав белков, в процессе трансляции . Она была сформулирована Фрэнсисом Криком в 1955 году в неофициальной публикации Клуба связей РНК , а позже разработана в 1957 году вместе с центральной догмой молекулярной биологии и гипотезой последовательности . Официально она была опубликована как статья «О синтезе белка» в 1958 году. Название «гипотеза адаптера» было дано Сиднеем Бреннером .
Крик предположил, что должна существовать небольшая молекула, которая точно распознает и связывает последовательности мРНК во время синтеза аминокислот. Позже было установлено, что гипотетическая молекула-адаптер представляет собой до сих пор неизвестную нуклеиновую кислоту, транспортную РНК (тРНК).
Разработка
[ редактировать ]В 1953 году английский биофизик Фрэнсис Крик и американский биолог Джеймс Уотсон , работая вместе в Кавендишской лаборатории Кембриджского университета, разработали правильное описание структуры ДНК — одного из основных генетических материалов. [1] В своей последующей статье в том же году они представили концепцию генетической информации наряду с представлением о том, что ДНК и белковое облако связаны между собой. [2] К 1954 году стало понятно, что ДНК , РНК (в то время понималась только информационная РНК, но только как неопределенная нуклеиновая кислота, идентифицированная как таковая только в 1960 году). [3] ) и белки были связаны как компоненты одного и того же пути генетической информации. Однако структура РНК и детали того, как эти биологические молекулы связаны и взаимодействуют друг с другом, все еще оставались загадкой, особенно то, как белки могут быть синтезированы из нуклеиновых кислот. [4] В своем письме Крику Уотсон назвал эту проблему «тайнами жизни». [3] Уотсон и Александр Рич обсуждали это в PNAS , заявив: «Мы не сможем проверить структурную взаимосвязь между РНК и синтезом белка или между РНК и ДНК, пока не узнаем структуру РНК». [5]
С 1940-х годов накапливались данные о том, что синтез белка происходит одновременно с повышением уровня РНК в цитоплазме. [6] [7] [8] Взаимосвязь между ДНК и РНК для синтеза белка была впервые выдвинута гипотезой французскими биологами Андре Буавеном и Роджером Вендрели в 1947 году. Они написали в журнале Experientia (позже переименованном в Cellular and Molecular Life Sciences ): «Благодаря каталитическому действию макромолекулярные дезоксирибонуклеиновые кислоты управляют построением макромолекулярных молекул. рибонуклеиновые кислоты, а они, в свою очередь, контролируют выработку цитоплазматических ферментов. Действительно, ферментативный аппарат возникает одновременно от действия рибонуклеиновых кислот (каталитическое действие) и от действия субстратов (массовое действие). [9]
Гипотезы о шаблоне нуклеиновой кислоты
[ редактировать ]Александр Даунс из Медицинского центра Рочестерского университета в Нью-Йорке подошел близко к правильной интерпретации синтеза белка из РНК. [10] В 1952 году он предложил первую концепцию, назвав ее «гипотезой матрицы нуклеиновой кислоты». [11] что белки состоят из РНК, а не из ДНК. Однако, поскольку точная химическая структура ДНК или РНК в то время не была известна, он ошибочно предположил, что белки и РНК имеют комплементарные последовательности, как он описал в «Энзимологии» : «специфическое расположение аминокислотных остатков в данной пептидной цепи обусловлено от специфического расположения нуклеотидных остатков в соответствующей специфической молекуле нуклеиновой кислоты». [12] это было встречено критикой в журнале Nature . В следующем году [13] на что Даунс защищал свою гипотезу:
В предложенной мной гипотезе в качестве матрицы сознательно была выбрана рибонуклеиновая кислота, поскольку из ряда соображений и экспериментальных исследований представляется вероятным, что синтез белка может происходить как в цитоплазме, так и в ядрах. В настоящее время не может быть возражений против предположения, что гены являются матрицами, но нет необходимости предполагать, что гены действуют непосредственно как матрицы для синтеза белка. [11]
Даунс также был близок к правильному и фундаментальному объяснению взаимосвязи генов с белками, поскольку он сказал: «Взаимосвязь между генами плазмы и ядерными генами [постулированная Солом Шпигельманом и Мартином Дэвидом Кэменом в 1946 году ] [14] ] может быть объяснено матричной последовательностью дезоксирибонуклеиновая кислота — рибонуклеиновая кислота — белок с дополнительным постулатом о сосуществовании независимого синтеза рибонуклеиновой кислоты, при этом рибонуклеиновая кислота действует как собственная матрица». [11] Правильная интерпретация передачи генетической информации (ДНК → РНК → белок), разработанная Криком в 1957 году, стала центральной догмой молекулярной биологии . [15] [16]
Гипотеза прямой матрицы ДНК
[ редактировать ]Советско-американский физик Джордж Гамов первым предложил последовательную схему синтеза белка из ДНК. [17] Основываясь на модели Уотсона-Крика , он предположил, что сама ДНК является непосредственной матрицей для синтеза белка. [18] Предполагая, что четыре основания ДНК могут образовывать 20 различных комбинаций в виде триплетов, он предположил, что разные аминокислоты должны соответствовать двадцатибуквенному алфавиту нуклеотидной последовательности. [19] В такой конфигурации ДНК напрямую производит белки из свободных молекул аминокислот. [20] В выпуске журнала Nature от 13 февраля 1954 года он объяснил:
Мне кажется, что такую процедуру трансляции можно легко установить, рассмотрев отношения « ключ-замок » между различными аминокислотами и ромбовидные «дырки», образованные различными нуклеотидами в цепи дезоксирибонуклеиновой кислоты... Один Можно предположить, что свободные аминокислоты из окружающей среды захватываются «дырками» молекул дезоксирибонуклеиновой кислоты и таким образом объединяются в соответствующие пептидные цепи. [21]
Уотсон, работавший тогда в Калифорнийском технологическом институте в Массачусетсе, тоже придумал эту проблему. [22] В отличие от Гамова, он осознавал возможную важность РНК как промежуточной стадии синтеза белка. [3] Первоначально он предполагал, что ДНК сначала превращается в РНК в результате химической реакции, но затем изменил свое мнение о ДНК как о матрице для синтеза РНК. [22] Эта последняя модель, которую он назвал «не безобразной», начинала становиться более убедительной. В письме Крику от 11 декабря 1954 года, обсуждая, как РНК может быть получена из ДНК, поскольку они содержат принципиально схожий нуклеотидный состав, он написал: «Я подозреваю, что ответ смотрит нам в лицо». [22]
Чтобы разгадать загадку синтеза белка, Гамов (как Синтезатор) создал в 1954 году неформальную группу ученых, которую он назвал Клубом приливов РНК. Согласно его предсказанию, в клубе могло быть только 20 членов с назначенными офицеры: Крик (как Пессимист), Рич (Лорд-хранитель печати) и Ватсон (Оптимист) среди них. Отчасти это было задумано как юмористическое. [23]
Гипотеза Крика
[ редактировать ]Наткнувшись на статью Гамова, Крик сразу заметил некоторые проблемы в схеме. Во-первых, он и Ватсон никогда раньше не считали 20 аминокислот или возможные 20 триплетов нуклеотидов. [19] (хотя впоследствии была доказана правильность Гамова, и его модель стала первым предсказанием генетического кода [17] [23] ). Другая проблема заключалась в том, что синтез РНК и белка, по-видимому, происходил исключительно в цитоплазме, а не в ядре, что указывает на то, что путь ДНК к белку требует не просто прямого химического связывания, но должен быть задействован какой-то медиатор. [10]
Крик начал думать о решении этой проблемы в духе идеи Уотсона о том, что роль РНК упускается из виду. Он отверг предположение Гамова о том, что такие молекулы, как аминокислоты, которые химически слишком отличаются от нуклеиновой кислоты, структурно невозможно связать с ДНК. Вместо этого он визуализировал определенную, но неизвестную молекулу, которая связывает нуклеиновые кислоты и переносит аминокислоты в рибосомы для образования пептидных цепей. [3] Когда он обсуждал со своим коллегой Сидни Бреннером особенности такой молекулы, Бреннер был впечатлен этой идеей и назвал эту концепцию «гипотезой адаптера». [24] Адаптером тогда была гипотетическая молекула, которая, по-видимому, способствует непрямому взаимодействию между ДНК, РНК и аминокислотами во время синтеза белка. [3] Как объяснил Крик:
Я не думаю, что кто-либо, глядя на ДНК или РНК, мог бы думать о них как о шаблонах для аминокислот, если бы не другие, косвенные доказательства... [В одной возможной схеме] каждая аминокислота соединялась бы химически, с помощью специального фермента, с небольшой молекулой, которая, имея специфическую поверхность с водородными связями, будет специфически соединяться с матрицей нуклеиновой кислоты. Эта комбинация также обеспечит энергию, необходимую для полимеризации. В своей простейшей форме существовало бы 20 различных типов молекул-адаптеров, по одному на каждую аминокислоту, и 20 различных ферментов, соединяющих аминокислоты с их адаптерами. Сидни Бреннер, с которым я обсуждал эту идею, называет это «гипотезой адаптера». «Поскольку каждая аминокислота снабжена адаптером для перехода к матрице... Гипотеза адаптера предполагает, что фактический набор из двадцати аминокислот, обнаруженных в белках, возник либо в результате исторической случайности, либо в результате биологического отбора на чрезвычайно примитивном уровне. этап." [24]
Крик признал, что понятия не имеет, какое химическое вещество является адаптером, но затем благоразумно предположил, что оно состоит из нуклеотидов, своего рода РНК, имеющих общие черты как ДНК, так и РНК. [3] Он даже предположил, что «например, инсулин, вероятно, представляет собой белки, созданные РНК. Возможно, существует особый класс белков, созданных ДНК, почти всегда в небольших количествах (и поэтому обычно игнорируемых). [24]
Матрица может состоять, возможно, из одной цепи РНК… Каждая молекула-адаптер, содержащая, скажем, ди- или тринуклеотид, будет присоединена к своей аминокислоте с помощью специального фермента. Эти молекулы затем диффундируют к микросомальным частицам и прикрепляются к нужному месту на основе РНК путем спаривания оснований.
Статья, разосланная членам Клуба связей РНК в январе 1955 года под названием «О вырожденных шаблонах и гипотезе адаптера: заметка для клуба связей РНК», описывается как «одна из самых важных неопубликованных статей в истории науки». [25] [26] и «самая известная неопубликованная статья в анналах молекулярной биологии». [27]
Гипотеза адаптера была полностью формализована в 1957 году. Крик представил подробное объяснение в лекции под названием «Синтез белка» 19 сентября 1957 года на симпозиуме Общества экспериментальной биологии по биологической репликации макромолекул, проходившем в Университетском колледже Лондона .
В дальнейшем лекция была расширена и опубликована под названием «О синтезе белка» в 1958 году, о чем историк науки Хорас Фриланд Джадсон заметил: «Эта статья навсегда изменила логику биологии». [28] Это потому, что это было первое всестороннее понимание генетической информации (позже названной центральной догмой молекулярной биологии), синтеза белка (известного как гипотеза последовательности ), роли РНК (гипотезы адаптера), а также существования генетического кода. . [3]
Объяснение
[ редактировать ]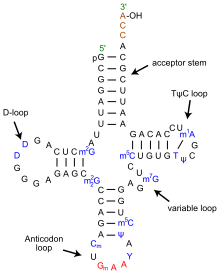
Гипотеза адаптера была сформулирована, чтобы объяснить, как информация может быть извлечена из нуклеиновой кислоты и использована для объединения цепочки аминокислот в определенной последовательности, причем эта последовательность определяется нуклеотидной последовательностью матрицы нуклеиновой кислоты (ДНК или РНК ). Крик предположил, что каждая аминокислота сначала присоединяется к своему специфическому «адаптерному» участку нуклеиновой кислоты (в результате реакции , катализируемой ферментами ). Порядок сборки аминокислот затем определяется специфическим узнаванием между адаптером и нуклеиновой кислотой, которая служит информационной матрицей. Таким образом, аминокислоты могут быть выстроены по матрице в определенном порядке. Соединение между соседними аминокислотами затем приведет к синтезу полипептида, последовательность которого определяется матричной нуклеиновой кислотой.
Основа
[ редактировать ]Мысли Крика, лежащие в основе этого предложения, были основаны на общем рассмотрении химических свойств двух классов молекул — нуклеиновых кислот и белков. Аминокислоты характеризуются наличием множества боковых цепей, которые варьируются от гидрофильных до гидрофобных : их индивидуальные особенности заключаются в совершенно разных свойствах, которыми обладают эти боковые цепи. Напротив, нуклеиновая кислота состоит из цепочки нуклеотидов, последовательность которых представляет собой геометрически определенную поверхность для образования водородных связей . Это позволяет нуклеиновым кислотам хорошо распознавать друг друга, но плохо различать различные боковые цепи аминокислот. Именно это очевидное отсутствие какой-либо возможности специфического распознавания боковых цепей аминокислот с помощью нуклеотидной последовательности привело Крика к выводу, что аминокислоты сначала присоединяются к небольшой нуклеиновой кислоте — адаптеру — и что это происходит путем спаривания оснований с матрица (предположительно, возникающая между нитями ДНК в двойной спирали) будет нести аминокислоты, которые должны быть выстроены в линию на матрице.
Доказательство
[ редактировать ]Гамов не был полностью не прав. Хотя его концепция о прямом синтезе аминокислот из двойных цепей ДНК оказалась ложной, его предсказания о природе аминокислот и о том, как они кодируются последовательностями ДНК (триплетными нуклеотидами), подтвердились — ключевые компоненты синтеза белка. [17] Это широкое понятие теперь известно как генетический код. Именно Крик вместе с Дж. С. Гриффитом и Лесли Оргелом (членом Клуба связей РНК), описывая предсказание из 20 аминокислот как «магическое число», представили подходящую концепцию в 1957 году:
[Порядок] аминокислот определяется порядком нуклеотидов нуклеиновой кислоты. В белках обычно встречается около двадцати встречающихся в природе аминокислот, но (обычно) только четыре различных нуклеотида. Проблема того, как последовательность из четырех вещей (нуклеотидов) может определять последовательность двадцати вещей (аминокислот), известна как проблема «кодирования». [29]
Когда Крик читал лекцию по своей гипотезе, такие адаптеры действительно существуют в природе, это уже было обнаружено командой Махлона Хогланда и Пола Замечника , чья статья была опубликована в следующем году, в марте 1958 года. [30] Эти «растворимые РНК» теперь называются транспортными РНК и опосредуют трансляцию информационных РНК на рибосомах в соответствии с правилами, содержащимися в генетическом коде . Крик предполагал, что его адаптеры будут небольшими, длиной примерно 5–10 нуклеотидов. На самом деле они намного крупнее, играют более сложную роль в синтезе белка и имеют длину около 100 нуклеотидов.
Ссылки
[ редактировать ]- ^ Уотсон Дж.Д., Крик Ф.Х. (1953). «Молекулярная структура нуклеиновых кислот: структура нуклеиновой кислоты дезоксирибозы» . Природа . 171 (4356): 737–8. Бибкод : 1953Natur.171..737W . дои : 10.1038/171737a0 . ПМИД 13054692 . S2CID 4253007 .
- ^ Уотсон, доктор медицинских наук; Крик, FH (30 мая 1953 г.). «Генетическое значение структуры дезоксирибонуклеиновой кислоты» . Природа . 171 (4361): 964–967. Бибкод : 1953Natur.171..964W . дои : 10.1038/171964b0 . ISSN 0028-0836 . ПМИД 13063483 . S2CID 4256010 .
- ^ Перейти обратно: а б с д и ж г Кобб, Мэтью (2017). «60 лет назад Фрэнсис Крик изменил логику биологии» . ПЛОС Биология . 15 (9): e2003243. дои : 10.1371/journal.pbio.2003243 . ПМК 5602739 . ПМИД 28922352 .
- ^ Парди, AB (1954). «Предшественники нуклеиновых кислот и синтез белка» . Труды Национальной академии наук Соединенных Штатов Америки . 40 (5): 263–270. Бибкод : 1954PNAS...40..263P . дои : 10.1073/pnas.40.5.263 . ПМК 534118 . ПМИД 16589470 .
- ^ Рич, А.; Уотсон, доктор медицинских наук (1954). «Некоторые отношения между ДНК и РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 40 (8): 759–764. Бибкод : 1954ПНАС...40..759Р . дои : 10.1073/pnas.40.8.759 . ПМК 534159 . ПМИД 16589555 .
- ^ Дж, Браше (1942). «Локализация пентонуклеиновых кислот в тканях животных и развивающихся яйцах амфибий» . Архивы биологии . 53 : 207–257.
- ^ Касперссон, Т. (1947). «Связь между нуклеиновой кислотой и синтезом белка» . Симпозиумы Общества экспериментальной биологии (1): 127–151. ISSN 0081-1386 . ПМИД 20257022 .
- ^ Туласне, Р.; Вендрели, Р. (1947). «Демонстрация бактериальных ядер с помощью рибонуклеазы» . Природа . 160 (4059): 225–226. Бибкод : 1947Natur.160..225T . дои : 10.1038/160225a0 . ISSN 0028-0836 . ПМИД 20256204 . S2CID 4105312 .
- ^ Бойвен, Андре; Вендрели, Роджер (1947). «О возможной роли двух нуклеиновых кислот в живой клетке» . Experientia (на французском языке). 3 (1): 32–34. дои : 10.1007/BF02155119 . ISSN 0014-4754 . ПМИД 20239698 . S2CID 46528362 .
- ^ Перейти обратно: а б Кобб, Мэтью (29 июня 2015 г.). «Кто открыл информационную РНК?» . Современная биология . 25 (13): 526–532 рэндов. Бибкод : 2015CBio...25.R526C . дои : 10.1016/j.cub.2015.05.032 . ISSN 1879-0445 . ПМИД 26126273 . S2CID 16443054 .
- ^ Перейти обратно: а б с Даунс, Алабама (19 сентября 1953 г.). «Матричная гипотеза нуклеиновой кислоты» . Природа . 172 (4377): 541. Бибкод : 1953Natur.172..541D . дои : 10.1038/172541a0 . ISSN 0028-0836 . ПМИД 13099258 . S2CID 4191207 .
- ^ Даунс, Алабама (1952). «Механизм дублирования пептидной цепи и синтеза нуклеиновых кислот» . Энзимология . 15 (5): 251–258. ISSN 0013-9424 . ПМИД 13033864 .
- ^ Кэмпбелл, Пенсильвания; Работа, ТС (06.06.1953). «Биосинтез белков» . Природа . 171 (4362): 997–1001. Бибкод : 1953Natur.171..997C . дои : 10.1038/171997a0 . ISSN 0028-0836 . ПМИД 13063503 . S2CID 4146549 .
- ^ Шпигельман, С.; Камен, доктор медицины (1946). «Гены и нуклеопротеины в синтезе ферментов» . Наука . 104 (2712): 581–584. Бибкод : 1946Sci...104..581S . дои : 10.1126/science.104.2712.581 . ISSN 0036-8075 . ПМИД 17749260 .
- ^ Крик, Ф. (8 августа 1970 г.). «Центральная догма молекулярной биологии» . Природа . 227 (5258): 561–563. Бибкод : 1970Natur.227..561C . дои : 10.1038/227561a0 . ISSN 0028-0836 . ПМИД 4913914 . S2CID 4164029 .
- ^ Иль, Александр М.; Ламонт, Ханна; Мэтьюз, Майкл Б. (23 февраля 2022 г.). «Возвращение к центральной догме: идеи синтеза белка, CRISPR и не только» . Междисциплинарные обзоры Wiley. РНК . 13 (5): e1718. дои : 10.1002/wrna.1718 . ISSN 1757-7012 . ПМИД 35199457 . S2CID 247082499 .
- ^ Перейти обратно: а б с Стегманн, Ульрих Э. (01 сентября 2016 г.). « Переосмысление« генетического кодирования »: анализ фактического использования» . Британский журнал философии науки . 67 (3): 707–730. дои : 10.1093/bjps/axv007 . ISSN 0007-0882 . ПМЦ 4990703 . ПМИД 27924115 .
- ^ Хейс, Брайан (1998). «Информатика: изобретение генетического кода» . Американский учёный . 86 (1): 8–14. дои : 10.1511/1998.17.3338 . ISSN 0003-0996 . JSTOR 27856930 . S2CID 121907709 .
- ^ Перейти обратно: а б Сегре, Джино (2000). «Большой взрыв и генетический код» . Природа . 404 (6777): 437. дои : 10.1038/35006517 . ПМИД 10761891 . S2CID 205005362 .
- ^ Рич, Александр (2009). «Эра пробуждения РНК: Структурная биология РНК в первые годы» . Ежеквартальные обзоры биофизики . 42 (2): 117–137. дои : 10.1017/S0033583509004776 . ISSN 0033-5835 . ПМИД 19638248 . S2CID 2285884 .
- ^ Гамов, Г. (1954). «Возможная связь между дезоксирибонуклеиновой кислотой и белковыми структурами» . Природа . 173 (4398): 318. Бибкод : 1954Natur.173..318G . дои : 10.1038/173318a0 . S2CID 4279494 .
- ^ Перейти обратно: а б с «Письмо Джеймса Д. Уотсона Фрэнсису Крику» . Фрэнсис Крик - Профили в науке . Проверено 22 июля 2022 г.
- ^ Перейти обратно: а б Штраус, Бернард С. (01 марта 2019 г.). «Мартинас Йчас: «Архивариус» Галстук-клуба РНК» . Генетика . 211 (3): 789–795. дои : 10.1534/genetics.118.301754 . ISSN 1943-2631 . ПМК 6404253 . ПМИД 30846543 .
- ^ Перейти обратно: а б с Крик, Фрэнсис (1955). «О вырожденных шаблонах и гипотезе адаптера: заметка для Клуба связей РНК» . Национальная медицинская библиотека . Проверено 21 июля 2022 г.
- ^ «Фрэнсис Крик — Профили в результатах научного поиска» . Profiles.nlm.nih.gov . Проверено 21 июля 2022 г.
- ^ «Фрэнсис Крик - Профили в науке: определение проблемы генетического кодирования, 1954–1957» . Национальная медицинская библиотека . 12 марта 2019 г. Проверено 21 июля 2022 г.
- ^ Фрай, Майкл (2022). «Гипотеза адаптера Крика и открытие транспортной РНК: эксперимент, превосходящий теоретические предсказания» . Философия, теория и практика биологии . 14 . дои : 10.3998/ptpbio.2628 . ISSN 2475-3025 . S2CID 249112573 .
- ^ Джадсон, Гораций Фриланд (1996). Восьмой день творения: творцы революции в биологии . Лабораторный пресс Колд-Спринг-Харбор. п. 330. ИСБН 978-0-87969-478-4 .
- ^ Крик, FH; Гриффит, Дж. С.; Оргель, Л.Е. (15 мая 1957 г.). «Коды без запятых» . Труды Национальной академии наук Соединенных Штатов Америки . 43 (5): 416–421. Бибкод : 1957PNAS...43..416C . дои : 10.1073/pnas.43.5.416 . ISSN 0027-8424 . ПМК 528468 . ПМИД 16590032 .
- ^ Хоугланд, МБ; Стивенсон, ML; Скотт, Дж. Ф.; Хехт, Л.И.; Замечник, ПК (1958). «Растворимая рибонуклеиновая кислота, промежуточное соединение в синтезе белка» . Журнал биологической химии . 231 (1): 241–257. дои : 10.1016/S0021-9258(19)77302-5 . ПМИД 13538965 .