Hfr-ячейка
Эта статья нуждается в дополнительных цитатах для проверки . ( декабрь 2009 г. ) |
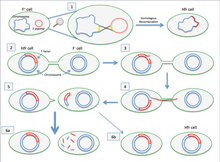
2. Клетка Hfr образует половые пили и прикрепляется к F-клетке-реципиенту.
3. Создается разрыв в одной цепи хромосомы клетки Hfr.
4.ДНК начинает переноситься из клетки Hfr в клетку-реципиент, пока реплицируется вторая цепь ее хромосомы.
5. Пилюс отделяется от клетки-реципиента и втягивается. В идеале клетка Hfr хочет передать весь свой геном клетке-реципиенту. Однако из-за своего большого размера и неспособности поддерживать контакт с клеткой-реципиентом он не способен этого сделать.
6. F-клетка остается F-, поскольку вся последовательность F-фактора не была получена. Поскольку гомологичной рекомбинации не произошло, перенесенная ДНК разрушается ферментами. В очень редких случаях фактор F будет полностью перенесен и F-клетка станет клеткой Hfr. [1]
Клетка высокочастотной рекомбинации (клетка Hfr) (также называемая штаммом Hfr ) представляет собой бактерию с конъюгативной плазмидой (например, F-фактором ), интегрированной в ее хромосомную ДНК. Интеграция плазмиды в хромосому клетки происходит посредством гомологичной рекомбинации . Конъюгативная плазмида, способная к интеграции хромосом, также называется эписомой (участок ДНК, который может существовать как плазмида или интегрироваться в хромосому). Когда происходит конъюгация, клетки Hfr очень эффективно доставляют хромосомные гены клетки реципиенту F. − клетки, у которых отсутствует эписома.
История
[ редактировать ]Штамм Hfr был впервые охарактеризован Лукой Кавалли-Сфорца . Уильям Хейс также независимо выделил еще один штамм Hfr. [2]
Перенос бактериальной хромосомы клетками Hfr
[ редактировать ]Клетка Hfr может переносить часть бактериального генома. Несмотря на то, что фактор F клеток Hfr интегрирован в хромосомную ДНК бактерий, он все же может инициировать конъюгативный перенос без предварительного вырезания из бактериальной хромосомы. Из-за присущей фактору F тенденции к переносу во время конъюгации остальная часть бактериального генома увлекается за ним. Поэтому, в отличие от обычного F + В клетке штаммы Hfr попытаются перенести всю свою ДНК через мост спаривания способом, аналогичным обычной конъюгации. При типичной конъюгации клетка-реципиент также становится F. + после конъюгации он получает полную копию плазмиды фактора F; но это не относится к конъюгации, опосредованной клетками Hfr. Из-за большого размера бактериальной хромосомы очень редко вся хромосома переносится в F. − клетки, поскольку требуемое время просто слишком велико для того, чтобы клетки могли поддерживать физический контакт. Следовательно, поскольку конъюгативный перенос не является полным (круговая природа плазмиды и бактериальной хромосомы требует полного переноса для переноса фактора F, поскольку он может быть разрезан посередине), реципиент F − клетки не получают полную последовательность F-фактора и не становятся F + из-за его неспособности образовывать половой пилус . [3]
Прерванная техника спаривания
[ редактировать ]При конъюгации, опосредованной клетками Hfr, перенос ДНК начинается в начале переноса ( oriT ), расположенном внутри F-фактора, а затем продолжается по или против часовой стрелки в зависимости от ориентации F-фактора в хромосоме. Следовательно, длина хромосомной ДНК, перенесенная в F − ячейки пропорциональна времени, в течение которого может произойти конъюгация. Это приводит к последовательному переносу генов на бактериальную хромосому. Бактериальные генетики используют этот принцип для картирования генов бактериальной хромосомы. Этот метод называется прерывистым спариванием, поскольку генетики позволяют конъюгации происходить в течение разных периодов времени, прежде чем остановить конъюгацию с помощью высокоскоростного блендера. Используя Hfr и F − штаммы, в которых один штамм несет мутации в нескольких генах, каждый из которых влияет на метаболическую функцию или вызывает устойчивость к антибиотикам , и исследуя фенотип клеток-реципиентов на чашках с селективным агаром, можно сделать вывод, какие гены передаются в клетки-реципиенты первыми и, следовательно, ближе к последовательность oriT на хромосоме.
F-первичная ячейка
[ редактировать ]F-прайм-клетка содержит F-плазмиду, которая интегрируется с хромосомной ДНК и несет с собой часть хромосомной ДНК при вырезании из хромосомы. Таким образом, F-прайм-плазмида представляет собой плазмиду, содержащую часть хромосомной ДНК, которая может быть перенесена в клетку-реципиент вместе с плазмидой во время конъюгации. [4]
Ссылки
[ редактировать ]- ^ «Генетический обмен» . www.microbiologybook.org . Проверено 4 декабря 2017 г.
- ^ Брукер, Роберт Дж. (2012). Генетика: анализ и принципы (4-е изд.). Нью-Йорк: МакГроу-Хилл. п. 186. ИСБН 9780073525280 .
- ^ Брукер, Роберт Дж. (2012). Генетика: анализ и принципы (4-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 186–187. ISBN 9780073525280 .
- ^ "микробиология-эволюционирующая-наука.pdf" . Drive.google.com . Проверено 01 июня 2020 г.