Бактериальная конъюгация
Бактериальная конъюгация – это передача генетического материала между бактериальными клетками путем прямого контакта между клетками или посредством мостообразной связи между двумя клетками. [1] Это происходит через пилус . [2] [ нужна полная цитата ] Это парасексуальный способ размножения бактерий.
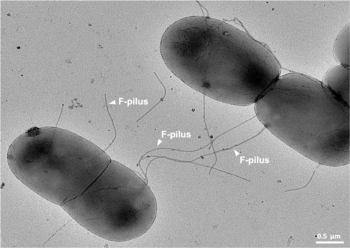
Это механизм горизонтального переноса генов , как и трансформация и трансдукция, хотя эти два других механизма не предполагают межклеточного контакта. [4]
Классическую бактериальную конъюгацию E. coli часто рассматривают как бактериальный эквивалент полового размножения или спаривания, поскольку она включает обмен генетическим материалом. Однако это не половое размножение, поскольку не происходит обмена гамет и даже образования нового организма : вместо этого трансформируется существующий организм. Во время классической E. coli конъюгации донорская клетка обеспечивает конъюгативный или мобилизуемый генетический элемент, который чаще всего представляет собой плазмиду или транспозон . [5] Большинство конъюгативных плазмид имеют системы, гарантирующие, что клетка- реципиент еще не содержит аналогичный элемент.
Передаваемая генетическая информация часто полезна для получателя. Преимущества могут включать устойчивость к антибиотикам , толерантность к ксенобиотикам или возможность использования новых метаболитов . [1] Другие элементы могут быть вредными и могут рассматриваться как бактериальные паразиты .
Конъюгация у Escherichia coli путем спонтанного зигогенеза. [6] и у Mycobacterium smegmatis путем дистрибутивного супружеского переноса. [7] [8] отличаются от более изученной классической конъюгации E. coli тем, что в этих случаях происходит существенное смешение родительских геномов .
История [ править ]
Процесс был открыт Джошуа Ледербергом и Эдвардом Татумом. [9] в 1946 году.
Механизм [ править ]
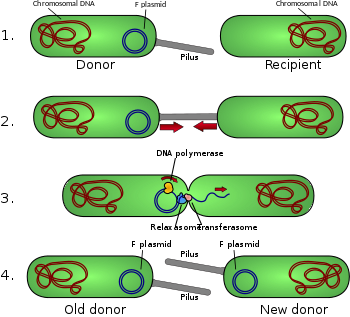
Диаграмма сопряжения
- Донорская клетка производит пилус .
- Pilus прикрепляется к клетке-реципиенту и объединяет две клетки.
- Мобильная плазмида разрезается, и затем одна цепь ДНК переносится в клетку-реципиент.
- Обе клетки синтезируют комплементарную цепь для производства двухцепочечной кольцевой плазмиды, а также воспроизводят пили; обе клетки теперь являются жизнеспособными донорами F-фактора. [1]
F -фактор представляет собой эписому (плазмиду, которая может интегрироваться в бактериальную хромосому путем гомологичной рекомбинации длиной около 100 т.п.н. ) Он несет в себе источник репликации , oriV , и источник передачи, или oriT . [5] В данной бактерии может быть только одна копия F-плазмиды, свободная или интегрированная, а бактерии, обладающие копией, называются F-положительными или F-плюс (обозначаются F + ). Клетки, в которых отсутствуют F-плазмиды, называются F-отрицательными или F-минус (F − ) и как таковые могут функционировать как клетки-реципиенты. [ нужна ссылка ]
Помимо прочей генетической информации, F-плазмида несет tra и trb локус , которые вместе имеют длину около 33 т.п.н. и состоят примерно из 40 генов . Локус tra включает ген пилина и регуляторные гены, которые вместе образуют пили на поверхности клетки. Локус также включает гены белков , которые прикрепляются к поверхности F. − бактерии и инициируют конъюгацию. Хотя ведутся споры о точном механизме конъюгации, похоже, что пили — это структуры, посредством которых происходит обмен ДНК. F-пили чрезвычайно устойчивы к механическим и термохимическим воздействиям, что гарантирует успешную конъюгацию в различных средах. [10] Некоторые белки, кодируемые локусами tra или trb , по-видимому, открывают канал между бактериями, и считается, что фермент traD, расположенный в основании ворсинок, инициирует слияние мембран.
Когда конъюгация инициируется сигналом, релаксаза фермент создает разрыв в одной из цепей конъюгативной плазмиды в oriT . Релаксаза может действовать отдельно или в комплексе из более чем дюжины белков, известных под общим названием релаксосома . В системе F-плазмид фермент релаксаза называется TraI, а релаксосома состоит из TraI, TraY, TraM и интегрированного фактора хозяина IHF. Разрезанная цепь, или Т-цепь , затем разматывается из неразрывной цепи и переносится в клетку-реципиент в направлении от 5'-конца к 3'-концу. Оставшаяся цепь реплицируется либо независимо от конъюгативного действия (вегетативная репликация, начинающаяся с oriV ), либо совместно с конъюгацией (конъюгативная репликация, подобная по катящемуся кругу репликации фага лямбда ). Для конъюгативной репликации может потребоваться второй разрыв, прежде чем произойдет успешный перенос. В недавнем отчете утверждается, что удалось ингибировать конъюгацию с химическими веществами, имитирующими промежуточный этап этого второго события разрыва. [11]
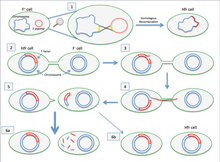
Если перенесенная F-плазмида ранее была интегрирована в геном донора (с образованием штамма Hfr («Высокая частота рекомбинации»)), некоторая часть хромосомной ДНК донора также может быть перенесена вместе с плазмидной ДНК. [4] Количество передаваемой хромосомной ДНК зависит от того, как долго две конъюгирующие бактерии остаются в контакте. У обычных лабораторных штаммов E. coli перенос всей бактериальной хромосомы занимает около 100 минут. Перенесенная ДНК затем может быть интегрирована в геном реципиента посредством гомологичной рекомбинации .
Культура клеток, содержащая в своей популяции клетки с неинтегрированными F-плазмидами, обычно содержит и несколько клеток, случайно интегрировавших свои плазмиды. Именно эти клетки ответственны за низкочастотные переносы хромосомных генов, происходящие в таких культурах. Некоторые штаммы бактерий с интегрированной F-плазмидой можно выделить и вырастить в чистой культуре. их называют Hfr ( высокочастотная очень эффективно переносят хромосомные гены , рекомбинация штаммы Поскольку такие ). Геном E. coli первоначально был картирован в ходе экспериментов по прерывистому спариванию, в которых различные клетки Hfr в процессе конъюгации отделялись от реципиентов менее чем через 100 минут (первоначально с использованием блендера Waring). Затем были исследованы перенесенные гены.
Поскольку интеграция F-плазмиды в хромосому E. coli является редким спонтанным явлением и поскольку многочисленные гены, способствующие переносу ДНК, находятся в геноме плазмиды, а не в бактериальном геноме, утверждалось, что конъюгативный перенос бактериальных генов, как он встречается в системе Hfr E. coli , не является эволюционной адаптацией бактериального хозяина и, вероятно, не является предком эукариотического пола. [13]
Спонтанный зигогенез у E. coli
В дополнение к классической бактериальной конъюгации, описанной выше для E. coli наблюдается форма конъюгации, называемая спонтанным зигогенезом (сокращенно Z-спариванием) , у некоторых штаммов E. coli . [6] При Z-спаривании происходит полное генетическое смешивание, и образуются нестабильные диплоиды , отбрасывающие фенотипически гаплоидные клетки, некоторые из которых демонстрируют родительский фенотип , а некоторые являются настоящими рекомбинантами .
Супружеский перенос микобактерий [ править ]
Конъюгация у Mycobacteria smegmatis , как и конъюгация у E. coli , требует стабильного и продолжительного контакта между донором и штаммом-реципиентом, устойчива к ДНКазе, а перенесенная ДНК включается в хромосому-реципиента путем гомологичной рекомбинации. Однако, в отличие от конъюгации Hfr E. coli , микобактериальная конъюгация основана на хромосоме, а не на плазмиде. [7] [8] Более того, в отличие от конъюгации Hfr E. coli , у M. smegmatis все участки хромосомы переносятся с сопоставимой эффективностью. Длина донорских сегментов широко варьируется, но их средняя длина составляет 44,2 КБ. Поскольку в среднем переносится 13 участков, среднее количество перенесенной ДНК на геном составляет 575 КБ. [8] Этот процесс называется «распределительная супружеская передача». [7] [8] Грей и др. [7] обнаружили существенное смешение родительских геномов в результате конъюгации и сочли это смешение напоминающим то, что наблюдается в мейотических продуктах полового размножения.
перенос ДНК у гипертермофильных Конъюгационно- подобный архей
Гипертермофильные археи кодируют пили, структурно сходные с бактериальными конъюгативными пилями. [14] Однако, в отличие от бактерий, где аппарат конъюгации обычно опосредует перенос мобильных генетических элементов, таких как плазмиды или транспозоны, у гипертермофильных архей используется конъюгативный аппарат, называемый Ced (кренархейная система обмена ДНК). [15] и Тед (система Thermoproteales для обмена ДНК), [14] по-видимому, отвечает за передачу клеточной ДНК между представителями одного и того же вида. Было высказано предположение, что у этих архей полностью приручен механизм конъюгации, способствующий репарации ДНК посредством гомологичной рекомбинации, а не распространения мобильных генетических элементов. [14] Помимо VirB2-подобного конъюгативного пилуса, системы Ced и Ted включают компоненты VirB6-подобной трансмембранной спаривающейся поры и VirB4-подобную АТФазу. [14]
Перенос между королевствами [ править ]

Бактерии, родственные азотфиксирующим ризобиям, представляют собой интересный случай межцарственной конъюгации . [16] Например, плазмида, индуцирующая опухоль (Ti) Agrobacterium , и плазмида, индуцирующая опухоль корня (Ri) A. rhizogenes, содержат гены, которые способны передаваться в растительные клетки. Экспрессия этих генов эффективно превращает растительные клетки в фабрики по производству опина . Опины используются бактериями в качестве источников азота и энергии. Инфицированные клетки образуют коронковую галловую или корневую опухоли . Таким образом, плазмиды Ti и Ri являются эндосимбионтами бактерий, которые, в свою очередь, являются эндосимбионтами (или паразитами) инфицированного растения. [ нужна ссылка ]
Плазмиды Ti и Ri также могут передаваться между бактериями с использованием системы ( оперон tra , или перенос ), которая отличается и независима от системы, используемой для переноса между царствами ( оперон vir , или вирулентность ). Такие передачи создают вирулентные штаммы из ранее авирулентных штаммов. [ нужна ссылка ]
инженерии генной Приложения
Конъюгация — удобный способ переноса генетического материала к различным мишеням. В лабораториях сообщалось об успешных передачах от бактерий к дрожжам. [17] растения, клетки млекопитающих, [18] [19] диатомовые водоросли [20] и изолированные митохондрии млекопитающих . [21] мишени Конъюгация имеет преимущества перед другими формами генетического переноса, включая минимальное нарушение клеточной оболочки и способность переносить относительно большие количества генетического материала (см. выше обсуждение переноса хромосом E. coli ). В растениеводстве Agrobacterium -подобная конъюгация дополняет другие стандартные носители, такие как вирус табачной мозаики (TMV). Хотя ВТМ способен инфицировать многие семейства растений, в первую очередь это травянистые двудольные растения . Agrobacterium -подобная конъюгация также в основном используется для двудольных растений, но реципиенты однодольных растений не являются редкостью. [ нужна ссылка ]
См. также [ править ]
- Половая конъюгация у водорослей и инфузорий
- Трансфекция
- Тройное спаривание
- Зиготическая индукция
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с Холмс, Рэндалл К.; Джоблинг, Майкл Г. (1996). «Генетика». У барона С. (ред.). Генетика: Конъюгация. в: Медицинская микробиология Барона (4-е изд.). Университет Техасского медицинского отделения. ISBN 0-9631172-1-1 . ПМИД 21413277 .
- ^ Доктор Црамарао, магистр наук, доктор философии. (1991). Бакалавр ботаники, том 1 .
- ^ Патковски, Йонаш (21 апреля 2023 г.). «F-pilus, совершенная бактериальная секс-машина» . Сообщество микробиологов Nature Portfolio .
- ^ Jump up to: Перейти обратно: а б с Гриффитс, AJF (1999). Введение в генетический анализ (7-е изд.). Сан-Франциско: WH Freeman. ISBN 978-0-7167-3520-5 . Архивировано из оригинала 08 февраля 2020 г. Проверено 11 августа 2023 г.
- ^ Jump up to: Перейти обратно: а б Райан, Кей Джей; Рэй, CG, ред. (2004). Медицинская микробиология Шерриса (4-е изд.). Нью-Йорк: МакГроу Хилл. стр. 60–64. ISBN 978-0-8385-8529-0 .
- ^ Jump up to: Перейти обратно: а б Грация, Жан-Пьер; Тири, Марк (1 сентября 2003 г.). «Спонтанный зигогенез у Escherichia coli, форма истинной сексуальности у прокариот» . Микробиология . 149 (9): 2571–2584. дои : 10.1099/mic.0.26348-0 . ПМИД 12949181 .
- ^ Jump up to: Перейти обратно: а б с д Грей, Тодд А.; Кривой, Джанет А.; Гарольд, Джессика; Палумбо, Майкл Дж.; Дербишир, Кейт М. (9 июля 2013 г.). «Дистрибутивный супружеский перенос у микобактерий порождает потомство с мейотическиподобным полногеномным мозаицизмом, что позволяет картировать локус идентичности спаривания» . ПЛОС Биология . 11 (7): e1001602. дои : 10.1371/journal.pbio.1001602 . ПМЦ 3706393 . ПМИД 23874149 .
- ^ Jump up to: Перейти обратно: а б с д Дербишир, Кейт М.; Грей, Тодд А. (17 января 2014 г.). «Дистрибутивная супружеская передача: новый взгляд на горизонтальный перенос генов и генетический обмен у микобактерий» . Микробиологический спектр . 2 (1). doi : 10.1128/microbiolspec.MGM2-0022-2013 . ПМЦ 4259119 . ПМИД 25505644 .
- ^ Ледерберг, Джошуа; Татум, Э.Л. (октябрь 1946 г.). «Рекомбинация генов в Escherichia Coli» . Природа . 158 (4016): 558. Бибкод : 1946Natur.158..558L . дои : 10.1038/158558a0 . ПМИД 21001945 . S2CID 1826960 .
- ^ Патковски, Йонаш Б.; Дальберг, Тобиас; Аминь, Химан; Галот, Дхармендер К.; Виджайраджратнам, Сукхитасри; Фогель, Джозеф П.; Фрэнсис, Мэтью С.; Бейкер, Джозеф Л.; Андерссон, Магнус; Кост, Джеймс Р.Д. (5 апреля 2023 г.). «Биомеханическая адаптивность F-pilus ускоряет конъюгативное распространение резистентности к противомикробным препаратам и образование биопленок» . Природные коммуникации . 14 1):1879.doi : ( 10.1038/s41467-023-37600-y . ПМЦ 10076315 . ПМИД 37019921 .
- ^ Лухан, Скотт А.; Гуогас, Лаура М.; Рагонезе, Хизер; Мэтсон, Стивен В.; Рединбо, Мэтью Р. (24 июля 2007 г.). «Нарушение распространения устойчивости к антибиотикам путем ингибирования конъюгативной ДНК-релаксазы» . Труды Национальной академии наук . 104 (30): 12282–12287. Бибкод : 2007PNAS..10412282L . дои : 10.1073/pnas.0702760104 . JSTOR 25436291 . ЧВК 1916486 . ПМИД 17630285 .
- ^ Майер, Джин. «Генетический обмен» . www.microbiologybook.org . Проверено 4 декабря 2017 г.
- ^ Мишод, Ричард Э.; Бернштейн, Харрис; Недельку, Аврора М. (май 2008 г.). «Адаптивное значение пола у микробных патогенов» (PDF) . Инфекция, генетика и эволюция . 8 (3): 267–285. Бибкод : 2008InfGE...8..267M . дои : 10.1016/j.meegid.2008.01.002 . ПМИД 18295550 .
- ^ Jump up to: Перейти обратно: а б с д Бельтран, Летисия К.; Цвиркайте-Крупович, Вирджиния; Миллер, Джессалин; Ван, Фэнбинь; Крейцбергер, Марк А.Б.; Патковски, Йонаш Б.; Коста, Тьяго Р.Д.; Схоутен, Стефан; Левенталь, Илья; Контичелло, Винсент П.; Эгельман, Эдвард Х.; Крупович, Март (07 февраля 2023 г.). «Аппарат для импорта ДНК архей гомологичен бактериальному аппарату конъюгации» . Природные коммуникации . 14 (1): 666. Бибкод : 2023NatCo..14..666B . дои : 10.1038/s41467-023-36349-8 . ISSN 2041-1723 . ПМЦ 9905601 . ПМИД 36750723 .
- ^ ван Вольферен, Марлин; Вагнер, Александр; ван дер Дос, Крис; Альберс, Соня-Верена (01 марта 2016 г.). «Архейная система Ced импортирует ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (9): 2496–2501. Бибкод : 2016PNAS..113.2496V . дои : 10.1073/pnas.1513740113 . ISSN 1091-6490 . ПМЦ 4780597 . ПМИД 26884154 .
- ^ Пан, Шен Ц.; Цзинь, Шоугуан; Бултон, Маргарет И.; Хоуз, Марта; Гордон, Милтон П.; Нестер, Юджин В. (июль 1995 г.). «Для переноса Т-ДНК в растения необходим фактор вирулентности Agrobacterium, кодируемый геном плазмиды Ti или хромосомным геном». Молекулярная микробиология . 17 (2): 259–269. doi : 10.1111/j.1365-2958.1995.mmi_17020259.x . ПМИД 7494475 . S2CID 38483513 .
- ^ Хайнеманн, Джек А.; Спрэг, Джордж Ф. (июль 1989 г.). «Бактериальные конъюгативные плазмиды мобилизуют перенос ДНК между бактериями и дрожжами». Природа . 340 (6230): 205–209. Бибкод : 1989Natur.340..205H . дои : 10.1038/340205a0 . ПМИД 2666856 . S2CID 4351266 .
- ^ Куник, Тайла; Цфира, Цви; Капульник, Йорам; Гафни, Едидия; Дингуолл, Колин; Цитовский, Виталий (13 февраля 2001 г.). «Генетическая трансформация клеток HeLa агробактериями» . Труды Национальной академии наук . 98 (4): 1871–1876. Бибкод : 2001PNAS...98.1871K . дои : 10.1073/pnas.041327598 . ЧВК 29349 . ПМИД 11172043 .
- ^ Уотерс, Вирджиния Л. (декабрь 2001 г.). «Конъюгация между клетками бактерий и млекопитающих». Природная генетика . 29 (4): 375–6. дои : 10.1038/ng779 . ПМИД 11726922 . S2CID 27160 .
- ^ Карас, Богумил Ж.; Дайнер, Рэйчел Э.; Лефевр, Стефан К.; Маккуэйд, Джефф; Филлипс, Алекс PR; Ноддингс, Чари М.; Брансон, Джон К.; Валас, Рубен Э.; Диринк, Томас Дж. (21 апреля 2015 г.). «Дизайнерские эписомы диатомей, доставленные путем бактериальной конъюгации» . Природные коммуникации . 6 : 6925. Бибкод : 2015NatCo...6.6925K . дои : 10.1038/ncomms7925 . ISSN 2041-1723 . ПМЦ 4411287 . ПМИД 25897682 .
- ^ Юн, Молодой Геол; Кооб, Майкл Д. (2005). «Трансформация изолированных митохондрий млекопитающих путем бактериальной конъюгации» . Исследования нуклеиновых кислот . 33 (16): е139. дои : 10.1093/нар/гни140 . ПМК 1201378 . ПМИД 16157861 .
Внешние ссылки [ править ]
- Бактериальная конъюгация (Flash-анимация)