Белок

Белки — это крупные биомолекулы и макромолекулы , которые содержат одну или несколько длинных цепочек аминокислотных остатков . Белки выполняют широкий спектр функций внутри организмов, включая катализацию метаболических реакций , репликацию ДНК , реакцию на стимулы , обеспечение структуры клеток и организмов и транспортировку молекул из одного места в другое. Белки отличаются друг от друга прежде всего последовательностью аминокислот, которая определяется нуклеотидной последовательностью их генов и обычно приводит к сворачиванию белка в специфическую трехмерную структуру , определяющую его активность.
Линейная цепь аминокислотных остатков называется полипептидом . Белок содержит по крайней мере один длинный полипептид. Короткие полипептиды, содержащие менее 20–30 остатков, редко считаются белками и обычно называются пептидами . Отдельные аминокислотные остатки связаны между собой пептидными связями и соседними аминокислотными остатками. Последовательность генетическом аминокислотных остатков в белке определяется последовательностью гена , который закодирован в коде . В общем, генетический код определяет 20 стандартных аминокислот; но у некоторых организмов генетический код может включать селеноцистеин и — у некоторых архей — пирролизин . Вскоре после или даже во время синтеза остатки белка часто химически модифицируются путем посттрансляционной модификации , которая изменяет физические и химические свойства, укладку, стабильность, активность и, в конечном итоге, функцию белков. К некоторым белкам присоединены непептидные группы, которые можно назвать простетическими группами или кофакторами . Белки также могут работать вместе для достижения определенной функции, и они часто объединяются, образуя стабильные белковые комплексы .
После образования белки существуют только в течение определенного периода, а затем разлагаются и перерабатываются клеточными механизмами в процессе белкового обмена . Продолжительность жизни белка измеряется периодом его полураспада и охватывает широкий диапазон. Они могут существовать в течение минут или лет со средней продолжительностью жизни 1–2 дня в клетках млекопитающих. Аномальные или неправильно свернутые белки разлагаются быстрее либо из-за того, что они предназначены для разрушения, либо из-за своей нестабильности.
Подобно другим биологическим макромолекулам, таким как полисахариды и нуклеиновые кислоты , белки являются важными частями организмов и участвуют практически во всех процессах внутри клеток . Многие белки представляют собой ферменты , которые катализируют биохимические реакции и имеют жизненно важное значение для обмена веществ . Белки также выполняют структурные или механические функции, такие как актин и миозин в мышцах и белки цитоскелета , которые образуют систему каркаса , поддерживающую форму клеток. Другие белки важны для передачи сигналов в клетках, иммунных реакций , клеточной адгезии и клеточного цикла . В рационе животных белки необходимы для обеспечения незаменимых аминокислот , которые не могут быть синтезированы . Пищеварение расщепляет белки для метаболического использования.
История и этимология
и другие признали белки как отдельный класс биологических молекул В восемнадцатом веке Антуан Фуркрой , отличающихся способностью молекул коагулировать или флокулировать при обработке теплом или кислотой. [1] Известные примеры в то время включали альбумин из яичных белков , альбумин сыворотки крови , фибрин и пшеничный глютен .
Белки были впервые описаны голландским химиком Герардусом Йоханнесом Мюлдером и названы шведским химиком Йонсом Якобом Берцелиусом в 1838 году. [2] [3] Малдер провел элементный анализ обычных белков и обнаружил, что почти все белки имеют одну и ту же эмпирическую формулу : C 400 H 620 N 100 O 120 P 1 S 1 . [4] Он пришел к ошибочному выводу, что они могут состоять из молекул одного типа (очень больших). Термин «белок» для описания этих молекул был предложен соратником Малдера Берцелиусом; Белок происходит от греческого слова πρώτειος ( протеиос ), что означает «первичный», [5] «впереди» или «стоять впереди», [6] + -в . Далее Малдер идентифицировал продукты деградации белков, такие как аминокислота лейцин, для которой он обнаружил (почти правильную) молекулярную массу 131 Да . [4] До слова «белок» использовались другие названия, например, «альбумины» или «белковые материалы» ( Eiweisskörper , на немецком языке). [6]
Ранние ученые-диетологи, такие как немец Карл фон Войт, считали, что белок является наиболее важным питательным веществом для поддержания структуры тела, поскольку обычно считалось, что «плоть делает плоть». [7] Карл Генрих Риттхаузен расширил известные формы белка, идентифицировав глутаминовую кислоту . На сельскохозяйственной экспериментальной станции Коннектикута подробный обзор растительных белков составил Томас Берр Осборн . Работая с Лафайетом Менделем и применяя закон минимума Либиха при кормлении лабораторных крыс , незаменимые в питании аминокислоты были установлены . Работу продолжил и сообщил Уильям Камминг Роуз . Понимание белков как полипептидов пришло благодаря работе Франца Хофмайстера и Германа Эмиля Фишера в 1902 году. [8] [9] Центральная роль белков как ферментов в живых организмах не была полностью оценена до 1926 года, когда Джеймс Б. Самнер показал, что фермент уреаза на самом деле является белком. [10]
Трудность очистки белков в больших количествах делала их изучение очень трудным для первых биохимиков белков. Поэтому ранние исследования были сосредоточены на белках, которые можно было очистить в больших количествах, например, белках крови, яичном белке, различных токсинах и пищеварительных/метаболических ферментах, полученных на скотобойнях. В 1950-х годах компания Armor Hot Dog Co. очистила 1 кг чистой бычьей панкреатической рибонуклеазы А и предоставила ее ученым бесплатно; этот жест помог рибонуклеазе А стать основной мишенью биохимических исследований на следующие десятилетия. [4]
Лайнусу Полингу приписывают успешное предсказание регулярных вторичных структур белков на основе водородных связей — идея, впервые выдвинутая Уильямом Эстбери в 1933 году. [11] Более поздняя работа Вальтера Каузмана по денатурации . [12] [13] частично основано на предыдущих исследованиях Кая Линдерстрем-Ланга , [14] способствовал пониманию сворачивания и структуры белков , опосредованных гидрофобными взаимодействиями .
Первым белком, который секвенировали, был инсулин , предложенный Фредериком Сэнгером в 1949 году. Сэнгер правильно определил аминокислотную последовательность инсулина, тем самым убедительно продемонстрировав, что белки состоят из линейных полимеров аминокислот, а не из разветвленных цепей, коллоидов или циклолов . [15] За это достижение он получил Нобелевскую премию в 1958 году. [16]

С развитием рентгеновской кристаллографии стало возможным секвенировать белковые структуры. [17] Первыми белковыми структурами, которые нужно было решить, были гемоглобин Макса Перуца и миоглобин Джона Кендрю в 1958 году. [18] [19] Использование компьютеров и увеличение вычислительной мощности также способствовали секвенированию сложных белков. В 1999 году Роджеру Корнбергу удалось секвенировать весьма сложную структуру РНК-полимеразы с помощью рентгеновских лучей высокой интенсивности от синхротронов . [17]
С тех пор криоэлектронная микроскопия (крио-ЭМ) крупных макромолекулярных ансамблей [20] был разработан. В крио-ЭМ используются замороженные образцы белков, а не кристаллы, и пучки электронов, а не рентгеновские лучи. Это наносит меньше вреда образцу, позволяя ученым получить больше информации и проанализировать более крупные структуры. [17] Вычислительное предсказание структуры белка малых белковых доменов [21] также помогло исследователям приблизиться к разрешению белковых структур на атомном уровне.По состоянию на апрель 2024 г. [update]Банк данных белков содержит 181 018 рентгеновских, 19 809 ЭМ и 12 697 ЯМР белковых структур. [22]
Количество белков, закодированных в геномах
Число белков, кодируемых в геноме, примерно соответствует числу генов (хотя может существовать значительное количество генов, кодирующих РНК белка, например рибосомальные РНК ). Вирусы обычно кодируют от нескольких до нескольких сотен белков, археи и бактерии — от нескольких сотен до нескольких тысяч, тогда как эукариоты обычно кодируют от нескольких тысяч до десятков тысяч белков ( Размер генома список примеров см. в разделе « »).
Классификация
Белки в первую очередь классифицируются по последовательности и структуре, хотя обычно используются и другие классификации. Специально для ферментов система нумерации ЕС обеспечивает схему функциональной классификации. Точно так же онтология генов классифицирует как гены, так и белки по их биологическим и биохимическим функциям, а также по их внутриклеточному расположению.
Сходство последовательностей используется для классификации белков как с точки зрения эволюционного, так и функционального сходства. При этом могут использоваться либо целые белки, либо белковые домены , особенно в многодоменных белках . Белковые домены позволяют классифицировать белки по сочетанию последовательности, структуры и функции, и их можно комбинировать разными способами. В раннем исследовании 170 000 белков примерно двум третям был присвоен по крайней мере один домен, при этом более крупные белки содержали больше доменов (например, белки, содержащие более 600 аминокислот , имели в среднем более 5 доменов). [23]
Биохимия

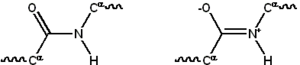
Большинство белков состоят из линейных полимеров, построенных из серий до 20 различных L -α- аминокислот. Все протеиногенные аминокислоты обладают общими структурными особенностями, включая α-углерод, которым аминогруппа , карбоксильная группа и вариабельная боковая цепь связана с . Только пролин отличается от этой базовой структуры, поскольку он содержит необычное кольцо у N-концевой аминогруппы, которое заставляет амидный фрагмент CO–NH принять фиксированную конформацию. [24] Боковые цепи стандартных аминокислот, подробно описанные в списке стандартных аминокислот , имеют большое разнообразие химических структур и свойств; именно совокупное воздействие всех боковых цепей аминокислот в белке в конечном итоге определяет его трехмерную структуру и химическую активность. [25] Аминокислоты в полипептидной цепи связаны пептидными связями . После соединения в белковой цепи отдельная аминокислота называется остатком , а связанный ряд атомов углерода, азота и кислорода известен как основная цепь или основная цепь белка. [26] : 19
Пептидная связь имеет две резонансные формы, которые придают некоторый характер двойной связи и препятствуют вращению вокруг своей оси, так что альфа-углероды примерно копланарны . Два других двугранных угла в пептидной связи определяют локальную форму, принимаемую основной цепью белка. [26] : 31 Конец со свободной аминогруппой известен как N-конец или аминоконец, тогда как конец белка со свободной карбоксильной группой известен как С-конец или карбокси-конец (последовательность белка записывается с N-конца). от конца к С-концу, слева направо).
Слова «белок» , «полипептид» и «пептид» немного двусмысленны и могут перекрываться по смыслу. Белок обычно используется для обозначения полной биологической молекулы в стабильной конформации , тогда как пептид обычно используется для обозначения коротких олигомеров аминокислот, часто не имеющих стабильной трехмерной структуры. Но граница между ними нечетко определена и обычно лежит в районе 20–30 остатков. [27] Полипептид может относиться к любой отдельной линейной цепи аминокислот, обычно независимо от ее длины, но часто подразумевает отсутствие определенной конформации .
Взаимодействия
Белки могут взаимодействовать со многими типами молекул, в том числе с другими белками , с липидами , углеводами и ДНК . [28] [29] [26] [30]
Обилие в клетках
Было подсчитано, что бактерии среднего размера содержат около 2 миллионов белков на клетку (например, E. coli и Staphylococcus aureus ). Меньшие бактерии, такие как микоплазма или спирохеты , содержат меньше молекул — примерно от 50 000 до 1 миллиона. Напротив, эукариотические клетки крупнее и, следовательно, содержат гораздо больше белка. Например, по оценкам, дрожжевые клетки содержат около 50 миллионов белков, а человеческие клетки — от 1 до 3 миллиардов. [31] Концентрация отдельных копий белка колеблется от нескольких молекул на клетку до 20 миллионов. [32] Не все гены, кодирующие белки, экспрессируются в большинстве клеток, и их количество зависит, например, от типа клеток и внешних раздражителей. Например, из примерно 20 000 белков, кодируемых геномом человека, только 6 000 обнаруживаются в лимфобластоидных клетках. [33]
Синтез
Биосинтез

Белки собираются из аминокислот с использованием информации, закодированной в генах. Каждый белок имеет свою уникальную аминокислотную последовательность, которая определяется нуклеотидной последовательностью гена, кодирующего этот белок. Генетический код представляет собой набор наборов из трех нуклеотидов, называемых кодонами , и каждая комбинация из трех нуклеотидов обозначает аминокислоту, например AUG ( аденин - урацил - гуанин ) является кодом метионина . Поскольку ДНК содержит четыре нуклеотида, общее число возможных кодонов равно 64; следовательно, в генетическом коде существует некоторая избыточность: некоторые аминокислоты определяются более чем одним кодоном. [30] : 1002–42 Гены, закодированные в ДНК, сначала транскрибируются в пре- мессенджерную РНК (мРНК) с помощью таких белков, как РНК-полимераза . Большинство организмов затем обрабатывают пре-мРНК (также известную как первичный транскрипт ), используя различные формы посттранскрипционной модификации в качестве матрицы для синтеза белка , чтобы сформировать зрелую мРНК, которая затем используется рибосомой . У прокариот мРНК может либо использоваться сразу после образования, либо связываться с рибосомой после удаления от нуклеоида . Напротив, эукариоты производят мРНК в ядре клетки , а затем перемещают ее через ядерную мембрану в цитоплазму , где синтез белка затем происходит . Скорость синтеза белка у прокариот выше, чем у эукариот, и может достигать до 20 аминокислот в секунду. [34]
Процесс синтеза белка из матрицы мРНК известен как трансляция . мРНК загружается на рибосому и считывается по три нуклеотида за раз путем сопоставления каждого кодона с спаривания оснований антикодоном , расположенным на молекуле транспортной РНК , которая несет аминокислоту, соответствующую распознаваемому ею кодону. Фермент аминоацил-тРНК-синтетаза «заряжает» молекулы тРНК правильными аминокислотами. Растущий полипептид часто называют зарождающейся цепью . Белки всегда биосинтезируются от N-конца к С-концу . [30] : 1002–42
Размер синтезированного белка можно измерить по количеству содержащихся в нем аминокислот и по его общей молекулярной массе , которая обычно выражается в дальтонах ( синоним атомных единиц массы ) или производной единице килодальтон (кДа). Средний размер белка увеличивается от архей к бактериям и эукариотам (283, 311, 438 остатков и 31, 34, 49 кДа соответственно) за счет большего числа белковых доменов, составляющих белки у высших организмов. [35] Например, дрожжевые белки имеют длину в среднем 466 аминокислот и массу 53 кДа. [27] Самыми крупными известными белками являются тайтины , компонент мышечного саркомера , с молекулярной массой почти 3000 кДа и общей длиной почти 27 000 аминокислот. [36]
Химический синтез
Короткие белки также можно синтезировать химическим путем с помощью семейства методов, известных как пептидный синтез , которые основаны на методах органического синтеза, таких как химическое лигирование, для получения пептидов с высоким выходом. [37] Химический синтез позволяет вводить неприродные аминокислоты в полипептидные цепи, например прикреплять флуоресцентные зонды к боковым цепям аминокислот. [38] Эти методы полезны в лабораторной биохимии и клеточной биологии , но, как правило, не для коммерческого применения. Химический синтез неэффективен для полипептидов длиной более 300 аминокислот, и синтезированные белки могут с трудом принять свою нативную третичную структуру . Большинство методов химического синтеза действуют от С-конца к N-концу, в противоположность биологической реакции. [39]
Структура

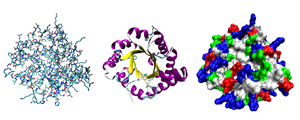
Большинство белков складываются в уникальные трехмерные структуры. Форма, в которую естественным образом сворачивается белок, известна как его нативная конформация . [26] : 36 Хотя многие белки могут сворачиваться без посторонней помощи, просто благодаря химическим свойствам их аминокислот, другим требуется помощь молекулярных шаперонов . для сворачивания в нативное состояние [26] : 37 Биохимики часто называют четыре различных аспекта структуры белка: [26] : 30–34
- Первичная структура : аминокислотная последовательность . Белок – это полиамид .
- Вторичная структура : регулярно повторяющиеся локальные структуры, стабилизированные водородными связями . Наиболее распространенными примерами являются α-спираль , β-лист и витки . Поскольку вторичные структуры локальны, в одной и той же белковой молекуле может присутствовать множество областей различной вторичной структуры.
- Третичная структура : общая форма отдельной белковой молекулы; пространственное соотношение вторичных структур друг к другу. Третичная структура обычно стабилизируется за счет нелокальных взаимодействий, чаще всего за счет образования гидрофобного ядра , а также за счет солевых мостиков , водородных связей, дисульфидных связей и даже посттрансляционных модификаций . Термин «третичная структура» часто используется как синоним термина « складка» . Третичная структура – это то, что контролирует основную функцию белка.
- Четвертичная структура : структура, образованная несколькими белковыми молекулами (полипептидными цепями), обычно называемыми в этом контексте белковыми субъединицами , которые функционируют как единый белковый комплекс .
- Пятинарная структура : признаки поверхности белка, которые организуют переполненную клеточную внутреннюю часть. Пятинарная структура зависит от временных, но существенных макромолекулярных взаимодействий, которые происходят внутри живых клеток.
Белки не являются полностью жесткими молекулами. В дополнение к этим уровням структуры белки могут переключаться между несколькими родственными структурами во время выполнения своих функций. В контексте этих функциональных перестроек эти третичные или четвертичные структуры обычно называют « конформациями », а переходы между ними называют конформационными изменениями. Такие изменения часто вызываются связыванием молекулы субстрата фермента с активным центром или физической областью белка, которая участвует в химическом катализе. В растворе структура белков также меняется из-за тепловой вибрации и столкновений с другими молекулами. [30] : 368–75

Белки можно неофициально разделить на три основных класса, которые коррелируют с типичными третичными структурами: глобулярные белки , волокнистые белки и мембранные белки . Почти все глобулярные белки растворимы , многие из них являются ферментами. Фиброзные белки часто являются структурными, например, коллаген , основной компонент соединительной ткани, или кератин , белковый компонент волос и ногтей. Мембранные белки часто служат рецепторами или обеспечивают каналы для прохождения полярных или заряженных молекул через клеточную мембрану . [30] : 165–85
Особый случай внутримолекулярных водородных связей внутри белков, плохо защищенных от воздействия воды и, следовательно, способствующих их собственной дегидратации , называется дегидронами . [40]
Белковые домены
Многие белки состоят из нескольких белковых доменов , то есть сегментов белка, которые складываются в отдельные структурные единицы. Домены обычно также имеют специфические функции, такие как ферментативная активность (например, киназа ), или они служат связывающими модулями (например, домен SH3 связывается с богатыми пролином последовательностями в других белках).
Мотив последовательности
Короткие аминокислотные последовательности внутри белков часто служат сайтами узнавания для других белков. [41] Например, домены SH3 обычно связываются с короткими мотивами PxxP (т.е. 2 пролинами [P], разделенными двумя неуказанными аминокислотами [x], хотя окружающие аминокислоты могут определять точную специфичность связывания). Многие такие мотивы собраны в базе данных Eukaryotic Linear Motif (ELM).
Топология белка
Топология белка описывает перепутывание основной цепи и расположение контактов внутри свернутой цепи. [42] две теоретические основы: теория узлов и топология цепей Для характеристики топологии белка были применены . Возможность описать топологию белка открывает новые пути для белковой инженерии и фармацевтических разработок, а также расширяет наше понимание заболеваний, связанных с неправильным сворачиванием белков, таких как нервно-мышечные расстройства и рак.
Клеточные функции
Белки являются главными действующими лицами в клетке и, как говорят, выполняют обязанности, определенные информацией, закодированной в генах. [27] За исключением некоторых типов РНК , большинство других биологических молекул являются относительно инертными элементами, на которые действуют белки. Белки составляют половину сухой массы клетки Escherichia coli , тогда как другие макромолекулы, такие как ДНК и РНК, составляют только 3% и 20% соответственно. [43] Набор белков, экспрессируемых в определенной клетке или типе клеток, известен как ее протеом .

Главной характеристикой белков, которая также обеспечивает их разнообразный набор функций, является их способность специфически и прочно связывать другие молекулы. Область белка, ответственная за связывание другой молекулы, известна как место связывания и часто представляет собой углубление или «карман» на поверхности молекулы. Эта связывающая способность опосредована третичной структурой белка, которая определяет карман сайта связывания, а также химическими свойствами боковых цепей окружающих аминокислот. Связывание с белками может быть чрезвычайно прочным и специфичным; например, белок -ингибитор рибонуклеазы связывается с ангиогенином человека с субфемтомолярной константой диссоциации (<10 −15 М), но вообще не связывается со своим гомологом онконазой амфибий (> 1 М). Чрезвычайно незначительные химические изменения, такие как добавление одной метильной группы к партнеру по связыванию, иногда могут быть достаточными для почти полного устранения связывания; например, аминоацил-тРНК-синтетаза, специфичная для аминокислоты валин, дискриминирует очень похожую боковую цепь аминокислоты изолейцин . [44]
Белки могут связываться с другими белками, а также с низкомолекулярными субстратами. Когда белки специфически связываются с другими копиями той же молекулы, они могут олигомеризоваться с образованием фибрилл; этот процесс часто происходит в структурных белках, которые состоят из глобулярных мономеров, которые самоассоциируются с образованием жестких волокон. Белково-белковые взаимодействия также регулируют ферментативную активность, контролируют ход клеточного цикла и позволяют собирать большие белковые комплексы , которые выполняют множество тесно связанных реакций с общей биологической функцией. Белки также могут связываться с клеточными мембранами или даже интегрироваться в них. Способность партнеров по связыванию вызывать конформационные изменения в белках позволяет строить чрезвычайно сложные сигнальные сети. [30] : 830–49 Поскольку взаимодействия между белками обратимы и во многом зависят от доступности различных групп белков-партнеров для формирования агрегатов, способных выполнять дискретные наборы функций, изучение взаимодействий между конкретными белками является ключом к пониманию важных аспектов клеточных функций. и, в конечном счете, свойства, которые отличают определенные типы клеток. [45] [46]
Ферменты
Самая известная роль белков в клетке — это ферменты , катализирующие химические реакции. Ферменты обычно высокоспецифичны и ускоряют только одну или несколько химических реакций. Ферменты осуществляют большинство реакций, участвующих в метаболизме , а также манипулируют ДНК в таких процессах, как репликация ДНК , репарация ДНК и транскрипция . Некоторые ферменты действуют на другие белки, добавляя или удаляя химические группы в процессе, известном как посттрансляционная модификация. Известно около 4000 реакций, катализируемых ферментами. [47] Ускорение скорости, обеспечиваемое ферментативным катализом, часто бывает огромным — до 10 17 -кратное увеличение скорости по сравнению с некатализируемой реакцией в случае оротатдекарбоксилазы (78 миллионов лет без фермента, 18 миллисекунд с ферментом). [48]
Молекулы, связанные и подвергающиеся воздействию ферментов, называются субстратами . Хотя ферменты могут состоять из сотен аминокислот, обычно лишь небольшая часть остатков вступает в контакт с субстратом, а еще меньшая часть — в среднем три-четыре остатка — принимает непосредственное участие в катализе. [49] Область фермента, которая связывает субстрат и содержит каталитические остатки, известна как активный центр .
Белки-диригенты относятся к классу белков, определяющих стереохимию соединений, синтезируемых другими ферментами. [50]
Передача сигналов в клетках и связывание лигандов
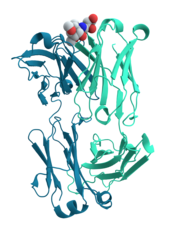
Многие белки участвуют в процессе клеточной передачи сигналов и передачи сигналов . Некоторые белки, такие как инсулин , являются внеклеточными белками, которые передают сигнал от клетки, в которой они были синтезированы, к другим клеткам в отдаленных тканях . Другие представляют собой мембранные белки , действующие как рецепторы , основная функция которых заключается в связывании сигнальной молекулы и индукции биохимического ответа в клетке. Многие рецепторы имеют сайт связывания, расположенный на поверхности клетки, и эффекторный домен внутри клетки, который может обладать ферментативной активностью или может претерпевать конформационные изменения, обнаруживаемые другими белками внутри клетки. [29] : 251–81
Антитела — это белковые компоненты адаптивной иммунной системы , основная функция которых — связывать антигены или чужеродные вещества в организме и нацеливать их на разрушение. Антитела могут секретироваться во внеклеточную среду или закрепляться в мембранах специализированных В-клеток, известных как плазматические клетки . В то время как ферменты ограничены в своей аффинности связывания со своими субстратами из-за необходимости проведения реакции, антитела таких ограничений не имеют. Аффинность связывания антитела с его мишенью чрезвычайно высока. [30] : 275–50
Многие белки-переносчики лигандов связывают отдельные небольшие биомолекулы и транспортируют их в другие места тела многоклеточного организма. Эти белки должны иметь высокую аффинность связывания, когда их лиганд присутствует в высоких концентрациях, но также должны высвобождать лиганд, когда он присутствует в низких концентрациях в тканях-мишенях. Каноническим примером лиганд-связывающего белка является гемоглобин , который переносит кислород из легких в другие органы и ткани у всех позвоночных и имеет близких гомологов во всех биологических царствах . [30] : 222–29 Лектины представляют собой сахарсвязывающие белки , которые очень специфичны в отношении своих сахарных фрагментов. Лектины обычно играют роль в явлениях биологического распознавания , затрагивающих клетки и белки. [51] Рецепторы и гормоны представляют собой высокоспецифичные связывающие белки.
Трансмембранные белки также могут служить белками-переносчиками лигандов, которые изменяют проницаемость клеточной мембраны для небольших молекул и ионов. Сама по себе мембрана имеет гидрофобное ядро, через которое полярные или заряженные молекулы не могут диффундировать . Мембранные белки содержат внутренние каналы, которые позволяют таким молекулам входить в клетку и выходить из нее. Многие белки ионных каналов специализируются на выборе только определенного иона; например, калиевые и натриевые каналы часто различают только один из двух ионов. [29] : 232–34
Структурные белки
Структурные белки придают жесткость и жесткость биологическим компонентам, которые в противном случае были бы жидкими. Большинство структурных белков являются волокнистыми белками ; например, коллаген и эластин являются важнейшими компонентами соединительной ткани , такой как хрящ , а кератин содержится в твердых или нитевидных структурах, таких как волосы , ногти , перья , копыта и панцири некоторых животных . [30] : 178–81 Некоторые глобулярные белки также могут выполнять структурные функции, например, актин и тубулин являются глобулярными и растворимы в виде мономеров, но полимеризуются с образованием длинных жестких волокон, составляющих цитоскелет , что позволяет клетке сохранять свою форму и размер.
Другими белками, выполняющими структурные функции, являются моторные белки, такие как миозин , кинезин и динеин , которые способны генерировать механические силы. Эти белки имеют решающее значение для клеточной подвижности одноклеточных организмов и спермы многих многоклеточных организмов, которые размножаются половым путем . Они также генерируют силы, возникающие при сокращении мышц. [30] : 258–64, 272 и играют важную роль во внутриклеточном транспорте.
Эволюция белка
Ключевой вопрос молекулярной биологии заключается в том, как развиваются белки, т.е. как мутации (или, скорее, изменения в последовательности аминокислот ) могут привести к появлению новых структур и функций? Большинство аминокислот в белке можно изменить без нарушения активности или функции, как это видно из многочисленных гомологичных белков разных видов (собранных в специализированных базах данных для семейств белков , например PFAM ). [52] Чтобы предотвратить драматические последствия мутаций, ген можно дублировать, прежде чем он сможет свободно мутировать. Однако это также может привести к полной потере функции генов и, следовательно, псевдогенов . [53] Чаще всего изменения отдельных аминокислот имеют ограниченные последствия, хотя некоторые из них могут существенно изменить функцию белков, особенно ферментов . Например, многие ферменты могут изменить свою субстратную специфичность в результате одной или нескольких мутаций. [54] Изменениям в субстратной специфичности способствует неразборчивость субстратов , т.е. способность многих ферментов связывать и обрабатывать несколько субстратов . Когда происходят мутации, специфичность фермента может увеличиваться (или уменьшаться) и, следовательно, его ферментативная активность. [54] Таким образом, бактерии (или другие организмы) могут адаптироваться к различным источникам пищи, включая такие неестественные субстраты, как пластик. [55]
Методы исследования
Методы, обычно используемые для изучения структуры и функции белка, включают иммуногистохимию , сайт-направленный мутагенез , рентгеновскую кристаллографию , ядерный магнитный резонанс и масс-спектрометрию .
Активность и структуру белков можно исследовать in vitro , in vivo и in silico . Исследования очищенных белков in vitro в контролируемых средах полезны для изучения того, как белок выполняет свою функцию: например, исследования кинетики ферментов изучают химический механизм каталитической активности фермента и его относительное сродство к различным возможным молекулам субстрата. Напротив, эксперименты in vivo могут предоставить информацию о физиологической роли белка в клетке или даже в целом организме . В исследованиях in silico для изучения белков используются вычислительные методы.
Очистка белка
Белки могут быть очищены от других клеточных компонентов с использованием различных методов, таких как ультрацентрифугирование , осаждение , электрофорез и хроматография ; появление генной инженерии сделало возможным ряд методов облегчения очистки.
Для проведения анализа in vitro белок необходимо очистить от других клеточных компонентов. Этот процесс обычно начинается с лизиса клеток , при котором клеточная мембрана разрушается и ее внутреннее содержимое высвобождается в раствор, известный как сырой лизат . Полученную смесь можно очистить с помощью ультрацентрифугирования , при котором различные клеточные компоненты фракционируются на фракции, содержащие растворимые белки; мембранные липиды и белки; клеточные органеллы и нуклеиновые кислоты . Осаждение методом, известным как высаливание, позволяет сконцентрировать белки из этого лизата. Затем используются различные типы хроматографии для выделения интересующего белка или белков на основе таких свойств, как молекулярная масса, суммарный заряд и аффинность связывания. [26] : 21–24 Уровень очистки можно контролировать с помощью различных типов гель-электрофореза желаемого белка молекулярная масса и изоэлектрическая точка , если известны , с помощью спектроскопии, если белок имеет различимые спектроскопические характеристики, или с помощью ферментативного анализа, если белок обладает ферментативной активностью. Кроме того, белки можно изолировать в зависимости от их заряда с помощью электрофокусирования . [56]
Для натуральных белков может потребоваться ряд стадий очистки, чтобы получить белок, достаточно чистый для лабораторного применения. Чтобы упростить этот процесс, часто используется генная инженерия , чтобы придать белкам химические свойства, которые облегчают их очистку, не влияя на их структуру или активность. Здесь «метка», состоящая из определенной аминокислотной последовательности, часто серии остатков гистидина (« His-метка »), прикреплена к одному концу белка. В результате, когда лизат пропускают через хроматографическую колонку, содержащую никель , остатки гистидина связывают никель и прикрепляются к колонке, в то время как непомеченные компоненты лизата проходят беспрепятственно. Был разработан ряд различных меток, которые помогают исследователям очищать определенные белки из сложных смесей. [57]
Клеточная локализация
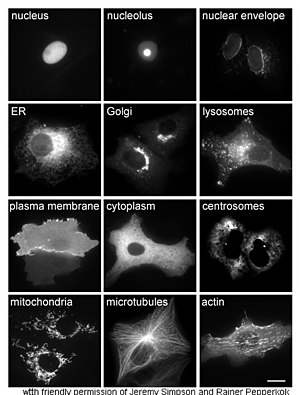
Изучение белков in vivo часто связано с синтезом и локализацией белка внутри клетки. Хотя многие внутриклеточные белки синтезируются в цитоплазме , а мембраносвязанные или секретируемые белки — в эндоплазматическом ретикулуме , особенности того, как белки направляются на определенные органеллы или клеточные структуры, часто неясны. Полезный метод оценки клеточной локализации использует генную инженерию для экспрессии в клетке слитого белка или химеры, состоящей из интересующего природного белка, связанного с « репортером », таким как зеленый флуоресцентный белок (GFP). [58] Положение слитого белка внутри клетки можно затем четко и эффективно визуализировать с помощью микроскопии . [59] как показано на рисунке напротив.
Другие методы выяснения клеточного расположения белков требуют использования известных компартментарных маркеров для таких областей, как ЭР, Гольджи, лизосомы или вакуоли, митохондрии, хлоропласты, плазматическая мембрана и т. д. С использованием флуоресцентно меченных версий этих маркеров или антител к известным маркерам становится гораздо проще определить локализацию интересующего белка. Например, непрямая иммунофлуоресценция позволит колокализировать флуоресценцию и продемонстрировать ее местоположение. Флуоресцентные красители используются для маркировки клеточных компартментов с той же целью. [60]
Существуют и другие возможности. Например, в иммуногистохимии обычно используются антитела к одному или нескольким интересующим белкам, которые конъюгированы с ферментами, дающими либо люминесцентные, либо хромогенные сигналы, которые можно сравнивать между образцами, что позволяет получить информацию о локализации. Другим применимым методом является кофракционирование в градиентах сахарозы (или другого материала) с использованием изопикнического центрифугирования . [61] Хотя этот метод не доказывает колокализацию компартмента известной плотности и интересующего белка, он увеличивает вероятность и более поддается крупномасштабным исследованиям.
Наконец, золотым стандартом клеточной локализации является иммуноэлектронная микроскопия . В этом методе наряду с классическими методами электронной микроскопии также используются антитела к интересующему белку. Образец готовят для обычного электронного микроскопического исследования, а затем обрабатывают антителом к интересующему белку, которое конъюгировано с чрезвычайно электроплотным материалом, обычно с золотом. Это позволяет локализовать как ультраструктурные детали, так и интересующий белок. [62]
С помощью другого приложения генной инженерии, известного как сайт-направленный мутагенез , исследователи могут изменить последовательность белка и, следовательно, его структуру, клеточную локализацию и восприимчивость к регуляции. Этот метод позволяет даже включать в белки неприродные аминокислоты с использованием модифицированных тРНК. [63] и может позволить рационально создавать новые белки с новыми свойствами. [64]
Протеомика
Полный набор белков, присутствующих одновременно в клетке или типе клеток, известен как ее протеом , и изучение таких крупномасштабных наборов данных определяет область протеомики , названную по аналогии со смежной областью геномики . Ключевые экспериментальные методы в протеомике включают 2D-электрофорез , [65] который позволяет разделять многие белки, масс-спектрометрию , [66] который позволяет быстро и высокопроизводительно идентифицировать белки и секвенировать пептиды (чаще всего после расщепления в геле ), белковые микрочипы , которые позволяют определять относительные уровни различных белков, присутствующих в клетке, и двухгибридный скрининг , который позволяет систематически исследовать белок-белковые взаимодействия . [67] Полный набор биологически возможных таких взаимодействий известен как интерактом . [68] Систематическая попытка определить структуру белков, представляющих все возможные складки, известна как структурная геномика . [69]
Определение структуры
Открытие третичной структуры белка или четвертичной структуры его комплексов может дать важные сведения о том, как белок выполняет свою функцию и как на него можно повлиять, например, при разработке лекарств . Поскольку белки слишком малы, чтобы их можно было увидеть под световым микроскопом , для определения их структуры приходится использовать другие методы. Общие экспериментальные методы включают рентгеновскую кристаллографию и ЯМР-спектроскопию , оба из которых могут предоставить структурную информацию с атомным разрешением. Однако эксперименты ЯМР могут предоставить информацию, на основе которой можно оценить подмножество расстояний между парами атомов, а окончательные возможные конформации белка определяются путем решения проблемы геометрии расстояний . Интерферометрия двойной поляризации — это количественный аналитический метод измерения общей конформации белка и конформационных изменений, вызванных взаимодействиями или другими стимулами. Круговой дихроизм — еще один лабораторный метод определения внутреннего β-листного/α-спирального состава белков. Криоэлектронная микроскопия используется для получения структурной информации с более низким разрешением об очень больших белковых комплексах, включая собранные вирусы ; [29] : 340–41 вариант, известный как электронная кристаллография, также может в некоторых случаях давать информацию с высоким разрешением, особенно для двумерных кристаллов мембранных белков. [70] Решенные структуры обычно хранятся в Банке данных белков (PDB), свободно доступном ресурсе, из которого можно получить структурные данные о тысячах белков в форме декартовых координат для каждого атома белка. [71]
Известно гораздо больше последовательностей генов, чем белковых структур. Кроме того, набор решенных структур смещен в сторону белков, которые можно легко подвергнуть условиям, необходимым для рентгеновской кристаллографии , одного из основных методов определения структуры. В частности, глобулярные белки сравнительно легко кристаллизовать при подготовке к рентгеновской кристаллографии. Мембранные белки и крупные белковые комплексы, напротив, трудно кристаллизовать, и они недостаточно представлены в PDB. [72] Инициативы в области структурной геномики попытались исправить эти недостатки путем систематического решения репрезентативных структур основных классов складок. Методы прогнозирования структуры белка пытаются предоставить средства создания вероятной структуры белков, структура которых не была определена экспериментально. [73]
Прогнозирование структуры

В дополнение к области структурной геномики, прогнозирование структуры белков разрабатывает эффективные математические модели белков для вычислительного предсказания молекулярных образований в теории вместо обнаружения структур с помощью лабораторных наблюдений. [74] Самый успешный тип предсказания структуры, известный как моделирование гомологии , основан на существовании «шаблонной» структуры с последовательностью, сходной с моделируемым белком; Цель структурной геномики - обеспечить достаточное представительство в решенных структурах для моделирования большинства из тех, которые остались. [75] Хотя создание точных моделей остается проблемой, когда доступны только отдаленно родственные матричные структуры, было высказано предположение, что выравнивание последовательностей является узким местом в этом процессе, поскольку можно создать довольно точные модели, если известно «идеальное» выравнивание последовательностей. [76] Многие методы предсказания структуры послужили основой для новой области белковой инженерии , в которой уже созданы новые белковые складки. [77] Кроме того, белки (у эукариот ~33%) содержат большие неструктурированные, но биологически функциональные сегменты и могут быть классифицированы как внутренне неупорядоченные белки . [78] Таким образом, прогнозирование и анализ белковых нарушений является важной частью характеристики структуры белка. [79]
Биоинформатика
Для анализа структуры, функций и эволюции белков был разработан широкий спектр вычислительных методов. Разработка таких инструментов была обусловлена большим количеством геномных и протеомных данных, доступных для различных организмов, включая геном человека . Экспериментально изучить все белки просто невозможно, поэтому лишь некоторые из них подвергаются лабораторным экспериментам, а для экстраполяции на аналогичные белки используются вычислительные инструменты. Такие гомологичные белки можно эффективно идентифицировать в отдаленно родственных организмах путем выравнивания последовательностей . Геном и последовательности генов можно искать с помощью различных инструментов по определенным свойствам. Инструменты профилирования последовательностей могут находить рестрикционных ферментов сайты , открытые рамки считывания в нуклеотидных последовательностях и прогнозировать вторичные структуры . филогенетические деревья можно построить и разработать эволюционные гипотезы С помощью специального программного обеспечения, такого как ClustalW, относительно происхождения современных организмов и генов, которые они экспрессируют. Область биоинформатика теперь незаменима для анализа генов и белков.
Моделирование динамических процессов in silico
Более сложная вычислительная проблема — предсказание межмолекулярных взаимодействий, например, при стыковке молекул . [80] сворачивание белков , белок-белковое взаимодействие и химическая реактивность. Математические модели для моделирования этих динамических процессов включают молекулярную механику , в частности, молекулярную динамику . В связи с этим при моделировании in silico было обнаружено сворачивание небольших α-спиральных белковых доменов, таких как головной убор злодеев , [81] белок ВИЧ вспомогательный [82] и гибридные методы, сочетающие стандартную молекулярную динамику с квантово-механической математикой, исследовали электронные состояния родопсинов . [83]
Помимо классической молекулярной динамики, методы квантовой динамики позволяют моделировать белки в атомистических деталях с точным описанием квантово-механических эффектов. Примеры включают многоуровневый, многоконфигурационный, зависящий от времени метод Хартри (MCTDH) и подход иерархических уравнений движения (HEOM), которые применялись для выращивания криптохромов. [84] и светособирающие комплексы бактерий, [85] соответственно. Как квантовое, так и классическое механическое моделирование систем биологического масштаба чрезвычайно требовательны к вычислительным ресурсам, поэтому инициативы по распределенным вычислениям (например, Folding@home) проект [86] ) облегчают молекулярное моделирование , используя достижения в области параллельной обработки графических процессоров и Монте-Карло методов .
Химический анализ
Общий азот органического вещества в основном формируется за счет аминогрупп белков. Общий азот по Кьельдалю ( TKN ) — это показатель азота, широко используемый при анализе (сточных) вод, почвы, пищевых продуктов, кормов и органических веществ в целом. Как следует из названия, метод Кьельдаля применяется . Существуют более чувствительные методы. [87] [88]
Питание
Большинство микроорганизмов и растений могут биосинтезировать все 20 стандартных аминокислот , в то время как животные (включая человека) должны получать часть аминокислот из рациона . [43] Аминокислоты, которые организм не может синтезировать самостоятельно, называются незаменимыми аминокислотами . У животных отсутствуют ключевые ферменты, синтезирующие определенные аминокислоты, такие как аспартокиназа , которая катализирует первый этап синтеза лизина , метионина и треонина из аспартата . Если аминокислоты присутствуют в окружающей среде, микроорганизмы могут сохранять энергию, поглощая аминокислоты из окружающей среды и подавляя свои пути биосинтеза.
У животных аминокислоты получаются при употреблении в пищу продуктов, содержащих белок. Поглощенные белки затем расщепляются на аминокислоты посредством пищеварения , которое обычно включает денатурацию белка под воздействием кислоты и гидролиз ферментами, называемыми протеазами . Некоторые поступившие в организм аминокислоты используются для биосинтеза белка, тогда как другие преобразуются в глюкозу посредством глюконеогенеза или поступают в цикл лимонной кислоты . Такое использование белка в качестве топлива особенно важно в условиях голодания , поскольку оно позволяет использовать собственные белки организма для поддержания жизни, особенно те, которые содержатся в мышцах . [89]
У животных, таких как собаки и кошки, белок поддерживает здоровье и качество кожи, способствуя росту и кератинизации волосяных фолликулов и тем самым снижая вероятность возникновения проблем с кожей, вызывающих неприятный запах. [90] Белки низкого качества также влияют на здоровье желудочно-кишечного тракта, увеличивая вероятность возникновения метеоризма и неприятных запахов у собак, поскольку, когда белки достигают толстой кишки в непереваренном состоянии, они ферментируются с образованием сероводорода, индола и скатола. [91] Собаки и кошки переваривают животные белки лучше, чем растительные, но плохо перевариваются продукты некачественного животного происхождения, в том числе кожа, перья и соединительная ткань. [91]
Механические свойства
Механические свойства белков весьма разнообразны и часто играют центральную роль в их биологических функциях, как в случае с такими белками, как кератин и коллаген . [92] Например, способность мышечной ткани постоянно расширяться и сокращаться напрямую связана с эластичными свойствами лежащего в ее основе белка. [93] [94] Помимо волокнистых белков, конформационная динамика ферментов [95] и структура биологических мембран , среди других биологических функций, определяется механическими свойствами белков. За пределами биологического контекста уникальные механические свойства многих белков, а также их относительная устойчивость по сравнению с синтетическими полимерами сделали их желанными объектами для разработки материалов следующего поколения. [96] [97]
Модуль Юнга
Юнга Модуль E рассчитывается как осевое напряжение σ в зависимости от результирующей деформации ε. Это мера относительной жесткости материала. В контексте белков эта жесткость часто напрямую коррелирует с биологической функцией. Например, коллаген , содержащийся в соединительной ткани , костях и хрящах , и кератин , содержащийся в ногтях, когтях и волосах, обладают жесткостью, которая на несколько порядков выше, чем у эластина . [98] который, среди прочего, придает эластичность таким структурам, как кровеносные сосуды , легочная ткань и ткань мочевого пузыря . [99] [100] По сравнению с этим глобулярные белки , такие как бычий сывороточный альбумин , которые относительно свободно плавают в цитозоле и часто функционируют как ферменты (и, таким образом, претерпевают частые конформационные изменения), имеют сравнительно гораздо более низкие модули Юнга. [101] [102]
Модуль Юнга отдельного белка можно найти с помощью молекулярно-динамического моделирования. Используя либо атомистические силовые поля, такие как CHARMM или GROMOS , либо крупнозернистые силовые поля, такие как Мартини, [103] отдельная молекула белка может быть растянута одноосной силой, в то время как полученное растяжение регистрируется для расчета деформации. [104] [105] Экспериментально такие методы, как атомно-силовая микроскопия . для получения аналогичных данных можно использовать [106]
На макроскопическом уровне модуль Юнга сшитых белковых сетей можно получить с помощью более традиционных механических испытаний . Экспериментально наблюдаемые значения для нескольких белков можно увидеть ниже.
| Белок | Класс белка | Модуль Юнга |
|---|---|---|
| Кератин (сшитый) | Фиброзный | 1,5-10 ГПа [107] |
| Эластин (сшитый) | Фиброзный | 1 МПа [98] |
| Фибрин (сшитый) | Фиброзный | 1-10 МПа [98] |
| Коллаген (сшитый) | Фиброзный | 5-7,5 ГПа [98] [108] |
| Резилин (сшитый) | Фиброзный | 1-2 МПа [98] |
| Бычий сывороточный альбумин (сшитый) | Шаровидный | 2,5-15 КПа [101] |
| Белки внешней мембраны β-цилиндра | Мембрана | 20-45 ГПа [109] |
Вязкость
Помимо функции ферментов внутри клетки, глобулярные белки часто действуют как ключевые транспортные молекулы. Например, сывороточные альбумины , ключевой компонент крови , необходимы для транспортировки множества небольших молекул по всему организму. [110] Из-за этого поведение этих белков в растворе, зависящее от концентрации, напрямую связано с функцией системы кровообращения . Одним из способов количественной оценки этого поведения является вязкость раствора.
Вязкость η обычно является мерой сопротивления жидкости деформации. Его можно рассчитать как соотношение между приложенным напряжением и скоростью изменения результирующей деформации сдвига, то есть скорости деформации. Вязкость сложных жидких смесей, таких как кровь, часто сильно зависит от температуры и концентрации растворенного вещества. [111] Для сывороточного альбумина, особенно бычьего сывороточного альбумина следующую зависимость между вязкостью, температурой и концентрацией . , можно использовать [112]
![{\displaystyle \eta =\exp \left[{\frac {c}{\alpha -\beta \ c}}\left(-B+DT+{\frac {\Delta E}{RT}}\right)\ верно]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/c875efcb377556dd7908fc43705d64ab6b749489)
Где c — концентрация, T — температура, R — газовая постоянная , а α, β, B , D и ΔE — константы свойств, зависящие от материала. Это уравнение имеет форму уравнения Аррениуса , определяющего вязкость экспоненциальной зависимостью от температуры и концентрации.
См. также
Ссылки
- ^ Осборн туберкулез (1909). «История» . Растительные белки . стр. 1–6.
- ^ Малдер Г.Дж. (1838). «О составе некоторых животных веществ» . Бюллетень физических и естественных наук в Нидерландах : 104.
- ^ Хартли Х. (август 1951 г.). «Происхождение слова «белок» » . Природа . 168 (4267): 244. Бибкод : 1951Natur.168..244H . дои : 10.1038/168244a0 . ПМИД 14875059 . S2CID 4271525 .
- ^ Jump up to: Перейти обратно: а б с Перретт Д. (август 2007 г.). «От белка к истокам клинической протеомики». Протеомика. Клинические применения . 1 (8): 720–738. дои : 10.1002/prca.200700525 . ПМИД 21136729 . S2CID 32843102 .
- ^ «Белок (сущ.)». Оксфордский словарь английского языка . Июль 2023 г. doi : 10.1093/OED/5657543824 .
- ^ Jump up to: Перейти обратно: а б Рейнольдс Дж. А., Танфорд С. (2003). Роботы природы: история белков (Оксфордские книги в мягкой обложке) . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета. п. 15. ISBN 978-0-19-860694-9 .
- ^ Бишофф Т.Л., Войт С. (1860). Законы питания травоядных, установленные новыми исследованиями (на немецком языке). Лейпциг, Гейдельберг.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ «Гофмейстер, Франц» . энциклопедия.com. Архивировано из оригинала 5 апреля 2017 года . Проверено 4 апреля 2017 г.
- ^ Кошланд Д.Э., Хауровиц Ф. «Белок. Раздел: Классификация белков» . britannica.com. Архивировано из оригинала 4 апреля 2017 года . Проверено 4 апреля 2017 г.
- ^ Самнер Дж. Б. (август 1926 г.). «Выделение и кристаллизация фермента уреазы» . Журнал биологической химии . 69 (2): 435–441. дои : 10.1016/S0021-9258(18)84560-4 .
- ^ Полинг Л., Кори Р.Б. (май 1951 г.). «Координаты атомов и структурные факторы двух спиральных конфигураций полипептидных цепей» . Труды Национальной академии наук Соединенных Штатов Америки . 37 (5): 235–240. Бибкод : 1951ПНАС...37..235П . дои : 10.1073/pnas.37.5.235 . ПМЦ 1063348 . ПМИД 14834145 .
- ^ Каузманн В. (май 1956 г.). «Структурные факторы денатурации белка». Журнал клеточной физиологии. Добавка . 47 (Приложение 1): 113–131. дои : 10.1002/jcp.1030470410 . ПМИД 13332017 .
- ^ Каузманн В. (1959). «Некоторые факторы интерпретации денатурации белка». Достижения в области химии белков. Том 14 . Том. 14. стр. 1–63. дои : 10.1016/S0065-3233(08)60608-7 . ISBN 978-0-12-034214-3 . ПМИД 14404936 .
- ^ Кальман С.М., Линдерстрем-Ланг К., Оттесен М., Ричардс Ф.М. (февраль 1955 г.). «Деградация рибонуклеазы субтилизином». Биохимика и биофизика Acta . 16 (2): 297–299. дои : 10.1016/0006-3002(55)90224-9 . ПМИД 14363272 .
- ^ Сэнгер Ф (1949). «Концевые пептиды инсулина» . Биохимический журнал . 45 (5): 563–574. дои : 10.1042/bj0450563 . ПМК 1275055 . ПМИД 15396627 .
- ^ Сэнгер Ф. (1958), Нобелевская лекция: Химия инсулина (PDF) , Nobelprize.org, заархивировано (PDF) из оригинала 19 марта 2013 г. , получено 9 февраля 2016 г.
- ^ Jump up to: Перейти обратно: а б с Стоддарт С. (1 марта 2022 г.). «Структурная биология: как белки оказались крупным планом» . Знающий журнал . doi : 10.1146/knowable-022822-1 .
- ^ Мюрхед Х., Перуц М.Ф. (август 1963 г.). «. Структура гемоглобина. Трехмерный Фурье-синтез восстановленного гемоглобина человека с разрешением 5,5 Å». Природа . 199 (4894): 633–638. Бибкод : 1963Natur.199..633M . дои : 10.1038/199633a0 . ПМИД 14074546 . S2CID 4257461 .
- ^ Кендрю Дж.К., Бодо Дж., Динцис Х.М., Пэрриш Р.Г., Вайкофф Х., Филлипс, округ Колумбия (март 1958 г.). «Трехмерная модель молекулы миоглобина, полученная методом рентгеноструктурного анализа». Природа . 181 (4610): 662–666. Бибкод : 1958Natur.181..662K . дои : 10.1038/181662a0 . ПМИД 13517261 . S2CID 4162786 .
- ^ Чжоу Чж. (апрель 2008 г.). «На пути к структурному определению атомного разрешения с помощью одночастичной криоэлектронной микроскопии» . Современное мнение в области структурной биологии . 18 (2): 218–228. дои : 10.1016/j.sbi.2008.03.004 . ПМК 2714865 . ПМИД 18403197 .
- ^ Кескин О., Тункбаг Н., Гурсой А. (апрель 2008 г.). «Характеристика и прогнозирование белковых интерфейсов для определения сетей белок-белкового взаимодействия». Современная фармацевтическая биотехнология . 9 (2): 67–76. дои : 10.2174/138920108783955191 . hdl : 11511/32640 . ПМИД 18393863 .
- ^ «Сводная статистика» . РЦСБ ПДБ . Проверено 20 апреля 2024 г.
- ^ Экман Д., Бьёрклунд А.К., Фрей-Скотт Дж., Элофссон А. (апрель 2005 г.). «Мультидоменные белки в трех царствах жизни: сиротские домены и другие неназначенные области». Журнал молекулярной биологии . 348 (1): 231–243. дои : 10.1016/j.jmb.2005.02.007 . ПМИД 15808866 .
- ^ Нельсон Д.Л., Кокс М.М. (2005). Принципы биохимии Ленингера (4-е изд.). Нью-Йорк, Нью-Йорк: WH Freeman and Company.
- ^ Гаттеридж А., Торнтон Дж. М. (ноябрь 2005 г.). «Понимание каталитического инструментария природы». Тенденции биохимических наук . 30 (11): 622–629. дои : 10.1016/j.tibs.2005.09.006 . ПМИД 16214343 .
- ^ Jump up to: Перейти обратно: а б с д и ж г Мюррей РФ, Харпер Х.В., Граннер Д.К., Мэйес П.А., Родвелл В.В. (2006). Иллюстрированная биохимия Харпера . Нью-Йорк: Lange Medical Books/McGraw-Hill. ISBN 978-0-07-146197-9 .
- ^ Jump up to: Перейти обратно: а б с Лодиш Х., Берк А., Мацудайра П., Кайзер К.А., Кригер М., Скотт М.П. и др. (2004). Молекулярно-клеточная биология (5-е изд.). Нью-Йорк, Нью-Йорк: WH Freeman and Company.
- ^ Ардеяни М.С., Пауэрс И.Т., Келли Дж.В. (август 2017 г.). «Использование кооперативно свернутых пептидов для измерения энергии взаимодействия и конформационных склонностей» . Отчеты о химических исследованиях . 50 (8): 1875–1882. doi : 10.1021/acs.accounts.7b00195 . ПМЦ 5584629 . ПМИД 28723063 .
- ^ Jump up to: Перейти обратно: а б с д Брэнден С., Туз Дж. (1999). Введение в структуру белка . Нью-Йорк: Паб Garland. ISBN 978-0-8153-2305-1 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Ван Холде К.Е., Мэтьюз К.К. (1996). Биохимия . Менло-Парк, Калифорния: Паб Benjamin/Cummings. компании, Inc. ISBN 978-0-8053-3931-4 .
- ^ Майло Р. (декабрь 2013 г.). «Каково общее количество белковых молекул в объеме клетки? Призыв переосмыслить некоторые опубликованные значения» . Биоэссе . 35 (12): 1050–1055. doi : 10.1002/bies.201300066 . ПМК 3910158 . ПМИД 24114984 .
- ^ Бек М., Шмидт А., Мальмстрем Дж., Клаассен М., Ори А., Шимборска А. и др. (ноябрь 2011 г.). «Количественный протеом линии клеток человека» . Молекулярная системная биология . 7 : 549. дои : 10.1038/msb.2011.82 . ПМК 3261713 . ПМИД 22068332 .
- ^ Ву Л., Кандилль С.И., Чой Ю., Се Д., Цзян Л., Ли-Пук-Тан Дж. и др. (июль 2013 г.). «Изменение и генетический контроль содержания белка у человека» . Природа . 499 (7456): 79–82. Бибкод : 2013Natur.499...79W . дои : 10.1038/nature12223 . ПМЦ 3789121 . ПМИД 23676674 .
- ^ Добсон CM (2000). «Природа и значение сворачивания белка». В боли Р.Х. (ред.). Механизмы сворачивания белка . Оксфорд, Оксфордшир: Издательство Оксфордского университета. стр. 1–28. ISBN 978-0-19-963789-8 .
- ^ Козловский Л.П. (январь 2017 г.). «Протеом-pI: база данных изоэлектрических точек протеома» . Исследования нуклеиновых кислот . 45 (Д1): Д1112–Д1116. дои : 10.1093/nar/gkw978 . ПМК 5210655 . ПМИД 27789699 .
- ^ Фултон А.Б., Айзекс В.Б. (апрель 1991 г.). «Титин, огромный эластичный саркомерный белок, который, вероятно, играет роль в морфогенезе». Биоэссе . 13 (4): 157–161. дои : 10.1002/bies.950130403 . ПМИД 1859393 . S2CID 20237314 .
- ^ Брукдорфер Т., Мардер О., Альберисио Ф. (февраль 2004 г.). «От производства пептидов в миллиграммах для исследований до многотонных количеств лекарств будущего». Современная фармацевтическая биотехнология . 5 (1): 29–43. дои : 10.2174/1389201043489620 . ПМИД 14965208 .
- ^ Шварцер Д., Коул П.А. (декабрь 2005 г.). «Полусинтез белка и выраженное лигирование белка: в погоне за хвостом белка». Современное мнение в области химической биологии . 9 (6): 561–569. дои : 10.1016/j.cbpa.2005.09.018 . ПМИД 16226484 .
- ^ Кент С.Б. (февраль 2009 г.). «Тотальный химический синтез белков». Обзоры химического общества . 38 (2): 338–351. дои : 10.1039/b700141j . ПМИД 19169452 . S2CID 5432012 .
- ^ Фернандес А., Скотт Р. (сентябрь 2003 г.). «Дегидрон: структурно закодированный сигнал взаимодействия белков» . Биофизический журнал . 85 (3): 1914–1928. Бибкод : 2003BpJ....85.1914F . дои : 10.1016/S0006-3495(03)74619-0 . ПМЦ 1303363 . ПМИД 12944304 .
- ^ Дэйви Н.Э., Ван Рой К., Уэзеритт Р.Дж., Тоедт Г., Уяр Б., Альтенберг Б. и др. (январь 2012 г.). «Признаки коротких линейных мотивов». Молекулярные биосистемы . 8 (1): 268–281. дои : 10.1039/c1mb05231d . ПМИД 21909575 .
- ^ Скальвини Б., Шейххассани В., Вудард Дж., Аупич Дж., Дам Р.Т., Джерала Р. и др. (июль 2020 г.). «Топология свернутых молекулярных цепей: от одиночных биомолекул до искусственного оригами». Тенденции в химии . 2 (7): 609–622. doi : 10.1016/j.trechm.2020.04.009 . hdl : 1887/3245505 . S2CID 218957613 .
- ^ Jump up to: Перейти обратно: а б Фут D, Фут JG. (2004). Биохимия Том 1 3-е изд. Wiley: Хобокен, Нью-Джерси.
- ^ Шанкаранарайанан Р., Морас Д. (2001). «Правильность перевода генетического кода» . Акта Биохимика Полоника . 48 (2): 323–335. дои : 10.18388/abp.2001_3918 . ПМИД 11732604 .
- ^ Копленд Дж.А., Шеффилд-Мур М., Колджич-Зиванович Н., Джентри С., Лампроу Г., Цорцату-Статопулу Ф. и др. (июнь 2009 г.). «Рецепторы половых стероидов при дифференцировке скелета и эпителиальной неоплазии: возможно ли тканеспецифическое вмешательство?». Биоэссе . 31 (6): 629–641. doi : 10.1002/bies.200800138 . ПМИД 19382224 . S2CID 205469320 .
- ^ Самарин С., Нусрат А. (январь 2009 г.). «Регуляция эпителиального апикального соединительного комплекса ГТФазами семейства Rho» . Границы бионауки . 14 (3): 1129–1142. дои : 10.2741/3298 . ПМИД 19273120 .
- ^ Байрох А. (январь 2000 г.). «База данных ENZYME в 2000 году» . Исследования нуклеиновых кислот . 28 (1): 304–305. дои : 10.1093/нар/28.1.304 . ПМК 102465 . ПМИД 10592255 .
- ^ Радзичка А., Вулфенден Р. (январь 1995 г.). «Опытный фермент». Наука . 267 (5194): 90–93. Бибкод : 1995Sci...267...90R . дои : 10.1126/science.7809611 . ПМИД 7809611 .
- ^ Внешние службы EBI (20 января 2010 г.). «Атлас каталитических сайтов Европейского института биоинформатики» . Ebi.ac.uk. Архивировано из оригинала 3 августа 2013 г. Проверено 16 января 2011 г.
- ^ Пикель Б., Шаллер А (октябрь 2013 г.). «Директивные белки: молекулярные характеристики и потенциальные биотехнологические применения». Прикладная микробиология и биотехнология . 97 (19): 8427–8438. дои : 10.1007/s00253-013-5167-4 . ПМИД 23989917 . S2CID 1896003 .
- ^ Рюдигер Х., Зиберт Х.К., Солис Д., Хименес-Барберо Х., Ромеро А., фон дер Лит К.В. и др. (апрель 2000 г.). «Медицинская химия, основанная на коде сахара: основы лектинологии и экспериментальные стратегии с лектинами в качестве мишени». Современная медицинская химия . 7 (4): 389–416. дои : 10.2174/0929867003375164 . ПМИД 10702616 .
- ^ Малдер, Нью-Джерси (28 сентября 2007 г.). «Базы данных по семействам белков». ЭЛС . Чичестер, Великобритания: John Wiley & Sons, Ltd., стр. a0003058.pub2. дои : 10.1002/9780470015902.a0003058.pub2 . ISBN 978-0-470-01617-6 .
- ^ Сису С., Пей Б., Ленг Дж., Франкиш А., Чжан Ю., Баласубраманиан С. и др. (сентябрь 2014 г.). «Сравнительный анализ псевдогенов трех типов» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (37): 13361–13366. Бибкод : 2014PNAS..11113361S . дои : 10.1073/pnas.1407293111 . ПМК 4169933 . ПМИД 25157146 .
- ^ Jump up to: Перейти обратно: а б Гусман Г.И., Сандберг Т.Э., Лакруа Р.А., Ньергес А., Папп Х., де Раад М. и др. (апрель 2019 г.). «Разнообразие ферментов формирует адаптацию к новым субстратам роста» . Молекулярная системная биология . 15 (4): е8462. дои : 10.15252/msb.20188462 . ПМК 6452873 . ПМИД 30962359 .
- ^ Бано К., Куддус М., Захир М.Р., Зия К., Хан М.Ф., Ашраф Г.М. и др. (2017). «Микробная ферментативная деградация биоразлагаемых пластмасс». Современная фармацевтическая биотехнология . 18 (5): 429–440. дои : 10.2174/1389201018666170523165742 . ПМИД 28545359 .
- ^ Эй Дж., Пош А., Коэн А., Лю Н., Харберс А. (2008). «Фракционирование сложных белковых смесей жидкофазным изоэлектрическим фокусированием». 2D-СТРАНИЦА: Подготовка проб и фракционирование . Методы молекулярной биологии. Том. 424. стр. 225–239. дои : 10.1007/978-1-60327-064-9_19 . ISBN 978-1-58829-722-8 . ПМИД 18369866 .
- ^ Терпе К (январь 2003 г.). «Обзор слияний белков-меток: от молекулярных и биохимических основ до коммерческих систем». Прикладная микробиология и биотехнология . 60 (5): 523–533. дои : 10.1007/s00253-002-1158-6 . ПМИД 12536251 . S2CID 206934268 .
- ^ Степаненко О.В., Верхуша В.В., Кузнецова И.М., Уверский В.Н., Туроверов К.К. (август 2008 г.). «Флуоресцентные белки как биомаркеры и биосенсоры: освещают молекулярные и клеточные процессы цветным светом» . Современная наука о белках и пептидах . 9 (4): 338–369. дои : 10.2174/138920308785132668 . ПМК 2904242 . ПМИД 18691124 .
- ^ Юсте Р. (декабрь 2005 г.). «Флуоресцентная микроскопия сегодня». Природные методы . 2 (12): 902–904. дои : 10.1038/nmeth1205-902 . ПМИД 16299474 . S2CID 205418407 .
- ^ Марголин В. (январь 2000 г.). «Зеленый флуоресцентный белок как репортер макромолекулярной локализации в бактериальных клетках». Методы . 20 (1): 62–72. дои : 10.1006/meth.1999.0906 . ПМИД 10610805 .
- ^ Уокер Дж. Х., Уилсон К. (2000). Принципы и методы практической биохимии . Кембридж, Великобритания: Издательство Кембриджского университета. стр. 287–89. ISBN 978-0-521-65873-7 .
- ^ Мэйхью Т.М., Люкок Дж.М. (август 2008 г.). «Разработки клеточной биологии для количественной иммуноэлектронной микроскопии на основе тонких срезов: обзор» . Гистохимия и клеточная биология . 130 (2): 299–313. дои : 10.1007/s00418-008-0451-6 . ПМК 2491712 . ПМИД 18553098 .
- ^ Хосака Т., Сисидо М. (декабрь 2002 г.). «Включение неприродных аминокислот в белки». Современное мнение в области химической биологии . 6 (6): 809–815. дои : 10.1016/S1367-5931(02)00376-9 . ПМИД 12470735 .
- ^ Седрон Ф., Менез А., Кеменёр Э. (август 2000 г.). «Настройка новых функций ферментов путем рационального изменения». Современное мнение в области структурной биологии . 10 (4): 405–410. дои : 10.1016/S0959-440X(00)00106-8 . ПМИД 10981626 .
- ^ Горг А., Вайс В., Данн М.Дж. (декабрь 2004 г.). «Современная технология двумерного электрофореза для протеомики». Протеомика . 4 (12): 3665–3685. дои : 10.1002/pmic.200401031 . ПМИД 15543535 . S2CID 28594824 .
- ^ Конротто П., Сушельницкий С (сентябрь 2008 г.). «Протеомные подходы в биологических и медицинских науках: принципы и приложения». Экспериментальная онкология . 30 (3): 171–180. ПМИД 18806738 .
- ^ Коегль М., Утц П. (декабрь 2007 г.). «Совершенствование двухгибридных систем скрининга дрожжей». Брифинги по функциональной геномике и протеомике . 6 (4): 302–312. дои : 10.1093/bfgp/elm035 . ПМИД 18218650 .
- ^ Плевчинский Д., Гинальский К. (2009). «Интерактом: предсказание белок-белковых взаимодействий в клетках» . Письма по клеточной и молекулярной биологии . 14 (1): 1–22. дои : 10.2478/s11658-008-0024-7 . ПМК 6275871 . ПМИД 18839074 .
- ^ Чжан С., Ким Ш. (февраль 2003 г.). «Обзор структурной геномики: от структуры к функции» . Современное мнение в области химической биологии . 7 (1): 28–32. дои : 10.1016/S1367-5931(02)00015-7 . ПМИД 12547423 .
- ^ Гонен Т., Ченг Ю., Слиз П., Хироаки Ю., Фудзиёси Ю., Харрисон С.С. и др. (декабрь 2005 г.). «Липид-белковые взаимодействия в двухслойных двумерных кристаллах AQP0» . Природа . 438 (7068): 633–638. Бибкод : 2005Natur.438..633G . дои : 10.1038/nature04321 . ПМК 1350984 . ПМИД 16319884 .
- ^ Стэндли Д.М., Киндзё А.Р., Киносита К., Накамура Х (июль 2008 г.). «Базы данных о структурах белков с новыми веб-сервисами для структурной биологии и биомедицинских исследований» . Брифинги по биоинформатике . 9 (4): 276–285. дои : 10.1093/нагрудник/bbn015 . ПМИД 18430752 .
- ^ Валиан П., Кросс Т.А., Япончик Б.К. (2004). «Структурная геномика мембранных белков» . Геномная биология . 5 (4): 215. doi : 10.1186/gb-2004-5-4-215 . ПМЦ 395774 . ПМИД 15059248 .
- ^ Слейтор Р.Д. (2012). «Прогнозирование функций белка». Функциональная геномика . Методы молекулярной биологии. Том. 815. стр. 15–24. дои : 10.1007/978-1-61779-424-7_2 . ISBN 978-1-61779-423-0 . ПМИД 22130980 .
- ^ Чжан Ю (июнь 2008 г.). «Прогресс и проблемы в предсказании структуры белков» . Современное мнение в области структурной биологии . 18 (3): 342–348. дои : 10.1016/j.sbi.2008.02.004 . ПМК 2680823 . ПМИД 18436442 .
- ^ Сян Цзы (июнь 2006 г.). «Достижения в моделировании структуры гомологических белков» . Современная наука о белках и пептидах . 7 (3): 217–227. дои : 10.2174/138920306777452312 . ПМК 1839925 . ПМИД 16787261 .
- ^ Чжан Ю, Сколник Дж (январь 2005 г.). «Проблема предсказания структуры белка может быть решена с использованием текущей библиотеки PDB» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (4): 1029–1034. Бибкод : 2005PNAS..102.1029Z . дои : 10.1073/pnas.0407152101 . ПМЦ 545829 . ПМИД 15653774 .
- ^ Кульман Б., Дантас Дж., Иретон Г.К., Варани Г., Стоддард Б.Л., Бейкер Д. (ноябрь 2003 г.). «Разработка новой складки глобулярного белка с точностью на атомном уровне». Наука . 302 (5649): 1364–1368. Бибкод : 2003Sci...302.1364K . дои : 10.1126/science.1089427 . ПМИД 14631033 . S2CID 1939390 .
- ^ Уорд Дж.Дж., Соди Дж.С., Макгаффин Л.Дж., Бакстон Б.Ф., Джонс Д.Т. (март 2004 г.). «Прогнозирование и функциональный анализ нативных нарушений в белках трех царств жизни». Журнал молекулярной биологии . 337 (3): 635–645. CiteSeerX 10.1.1.120.5605 . дои : 10.1016/j.jmb.2004.02.002 . ПМИД 15019783 .
- ^ Томпа П., Фершт А (2009). Структура и функции внутренне неупорядоченных белков . ЦРК Пресс. ISBN 978-1-4200-7893-0 . [ нужна страница ]
- ^ Ричи Д.В. (февраль 2008 г.). «Последний прогресс и будущие направления в стыковке белков с белками». Современная наука о белках и пептидах . 9 (1): 1–15. CiteSeerX 10.1.1.211.4946 . дои : 10.2174/138920308783565741 . ПМИД 18336319 .
- ^ Загрович Б, Сноу CD, Рубашки MR, Панде VS (ноябрь 2002 г.). «Моделирование сворачивания небольшого альфа-спирального белка в атомистических деталях с использованием распределенных по всему миру вычислений». Журнал молекулярной биологии . 323 (5): 927–937. CiteSeerX 10.1.1.142.8664 . дои : 10.1016/S0022-2836(02)00997-X . ПМИД 12417204 .
- ^ Хергес Т., Венцель В. (январь 2005 г.). «Складывание трехспирального белка in silico и характеристика его ландшафта свободной энергии в полноатомном силовом поле». Письма о физических отзывах . 94 (1): 018101. arXiv : физика/0310146 . Бибкод : 2005PhRvL..94a8101H . doi : 10.1103/PhysRevLett.94.018101 . ПМИД 15698135 . S2CID 1477100 .
- ^ Хоффманн М., Ванко М., Стродел П., Кениг П.Х., Фрауэнхайм Т., Шультен К. и др. (август 2006 г.). «Настройка цвета родопсинов: механизм спектрального сдвига между бактериородопсином и сенсорным родопсином II». Журнал Американского химического общества . 128 (33): 10808–10818. дои : 10.1021/ja062082i . ПМИД 16910676 .
- ^ Мендиве-Тапия Д., Манго Э., Фирмино Т., де ла Ланде А., Десаутер-Лекомт М., Мейер Х.Д. и др. (январь 2018 г.). «Многомерное квантово-механическое моделирование электронного переноса и электронной когерентности в криптохромах растений: роль начальных условий ванны». Журнал физической химии. Б. 122 (1): 126–136. дои : 10.1021/acs.jpcb.7b10412 . ПМИД 29216421 .
- ^ Стрюмпфер Дж., Шультен К. (август 2012 г.). «Открытые расчеты квантовой динамики с помощью иерархических уравнений движения на параллельных компьютерах» . Журнал химической теории и вычислений . 8 (8): 2808–2816. дои : 10.1021/ct3003833 . ПМК 3480185 . ПМИД 23105920 .
- ^ Шерага Х.А., Халили М., Ливо А. (2007). «Динамика сворачивания белков: обзор методов молекулярного моделирования». Ежегодный обзор физической химии . 58 : 57–83. Бибкод : 2007ARPC...58...57S . doi : 10.1146/annurev.physchem.58.032806.104614 . ПМИД 17034338 .
- ^ Муньос-Уэрта Р.Ф., Гевара-Гонсалес Р.Г., Контрерас-Медина Л.М., Торрес-Пачеко И., Прадо-Оливарес Х., Окампо-Веласкес Р.В. (август 2013 г.). «Обзор методов определения азотного статуса растений: преимущества, недостатки и последние достижения» . Датчики . 13 (8): 10823–10843. Бибкод : 2013Senso..1310823M . дои : 10.3390/s130810823 . ПМК 3812630 . ПМИД 23959242 .
- ^ Мартин П.Д., Мэлли Д.Ф., Мэннинг Дж., Фуллер Л. (ноябрь 2002 г.). «Определение органического углерода и азота почвы на полевом уровне методом ближней инфракрасной спектроскопии». Канадский журнал почвоведения . 82 (4): 413–422. дои : 10.4141/S01-054 .
- ^ Броснан Дж.Т. (июнь 2003 г.). «Межорганный транспорт аминокислот и его регуляция» . Журнал питания . 133 (6 Приложение 1): 2068S–2072S. дои : 10.1093/jn/133.6.2068S . ПМИД 12771367 .
- ^ Уотсон Т.Д. (декабрь 1998 г.). «Диета и кожные заболевания у собак и кошек» . Журнал питания . 128 (12 Доп.): 2783S–2789S. дои : 10.1093/jn/128.12.2783S . ПМИД 9868266 .
- ^ Jump up to: Перейти обратно: а б Кейс Л.П., Даристотель Л., Хайек М.Г., Рааш М.Ф. (2010). Электронная книга о питании собак и кошек: ресурс для специалистов по домашним животным . Elsevier Науки о здоровье.
- ^ Гослин Дж., Лилли М., Кэррингтон Э., Геретт П., Ортлепп С., Сэвидж К. (февраль 2002 г.). Бэйли А.Дж., Макмиллан Дж., Шрюри П.Р., Тэтэм А.С. (ред.). «Эластичные белки: биологическая роль и механические свойства» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 357 (1418): 121–132. дои : 10.1098/rstb.2001.1022 . ПМК 1692928 . ПМИД 11911769 .
- ^ Маруяма К., Натори Р., Нономура Ю. (июль 1976 г.). «Новый эластичный белок из мышц». Природа . 262 (5563): 58–60. дои : 10.1038/262058a0 . ПМИД 934326 .
- ^ Цховребова Л., Триник Дж. (февраль 2012 г.). «Делаем мышцы эластичными: структурная основа растяжения миомезина» . ПЛОС Биология . 10 (2): e1001264. дои : 10.1371/journal.pbio.1001264 . ПМЦ 3279349 . ПМИД 22347814 .
- ^ Мизраджи Э., Асеренца Л., Лин Дж. (ноябрь 1987 г.). «Вязкоупругие модели ферментов с множественными конформационными состояниями». Журнал теоретической биологии . 129 (2): 163–175. дои : 10.1016/s0022-5193(87)80010-3 . ПМИД 3455460 .
- ^ Шиллер Т., Шайбель Т. (18 апреля 2024 г.). «Биоинспирированные и биомиметические волокна на белковой основе и их применение» . Коммуникационные материалы . 5 (1): 1–18. дои : 10.1038/s43246-024-00488-2 . ISSN 2662-4443 .
- ^ Сунь Дж., Хэ Х., Чжао К., Ченг В., Ли Ю., Чжан П. и др. (сентябрь 2023 г.). «Белковые волокна с самовосстанавливающимися механическими свойствами с помощью динамической иминной химии» . Природные коммуникации . 14 (1): 5348. doi : 10.1038/s41467-023-41084-1 . ПМЦ 10475138 . ПМИД 37660126 .
- ^ Jump up to: Перейти обратно: а б с д и Гутольд М., Лю В., Спаркс Е.А., Джаверт Л.М., Пэн Л., Фалво М. и др. (02.10.2007). «Сравнение механических и структурных свойств фибриновых волокон с другими белковыми волокнами» . Клеточная биохимия и биофизика . 49 (3): 165–181. дои : 10.1007/s12013-007-9001-4 . ПМК 3010386 . ПМИД 17952642 .
- ^ Ван К., Мэн X, Го Цзы (2021). «Структура эластина, синтез, регуляторный механизм и связь с сердечно-сосудистыми заболеваниями» . Границы клеточной биологии и биологии развития . 9 : 596702. doi : 10.3389/fcell.2021.596702 . ПМЦ 8670233 . ПМИД 34917605 .
- ^ Дебелль Л., Тамбурро А.М. (февраль 1999 г.). «Эластин: молекулярное описание и функция». Международный журнал биохимии и клеточной биологии . 31 (2): 261–272. дои : 10.1016/S1357-2725(98)00098-3 . ПМИД 10216959 .
- ^ Jump up to: Перейти обратно: а б Хури Л.Р., Попа I (ноябрь 2019 г.). «Химическое разворачивание белковых доменов вызывает изменение формы запрограммированных белковых гидрогелей» . Природные коммуникации . 10 (1): 5439. doi : 10.1038/s41467-019-13312-0 . ПМК 6884551 . ПМИД 31784506 .
- ^ Тан Р., Шин Дж., Хо Дж., Коул Б.Д., Хон Дж., Чан Ю (октябрь 2020 г.). «Настройка структурной целостности и механических свойств глобулярных белковых везикул путем смешивания сшиваемых и несшиваемых строительных блоков». Биомакромолекулы . 21 (10): 4336–4344. doi : 10.1021/acs.biomac.0c01147 . ПМИД 32955862 .
- ^ Соуза П.С., Алессандри Р., Барну Дж., Талмайр С., Фаустино И., Грюневальд Ф. и др. (апрель 2021 г.). «Мартини 3: силовое поле общего назначения для крупнозернистой молекулярной динамики» (PDF) . Природные методы . 18 (4): 382–388. дои : 10.1038/s41592-021-01098-3 . ПМИД 33782607 .
- ^ «Домашняя страница Петра Шимчака» . www.fuw.edu.pl. Проверено 13 мая 2024 г.
- ^ Мэпплбек С., Бут Дж., Шалашилин Д. (август 2021 г.). «Моделирование динамики вытягивания белка во втором временном масштабе с помощью молекулярной динамики в штучной упаковке». Журнал химической физики . 155 (8): 085101. дои : 10.1063/5.0059321 . ПМИД 34470356 .
- ^ Каррион-Васкес М., Марсалек П.Е., Оберхаузер А.Ф., Фернандес Х.М. (сентябрь 1999 г.). «Атомно-силовая микроскопия фиксирует фенотипы длины отдельных белков» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (20): 11288–11292. дои : 10.1073/pnas.96.20.11288 . ЧВК 18026 . ПМИД 10500169 .
- ^ МакКиттрик Дж., Чен П.Ю., Бодде С.Г., Ян В., Новицкая Е.Е., Мейерс М.А. (3 апреля 2012 г.). «Структура, функции и механические свойства кератина» . ДЖОМ . 64 (4): 449–468. дои : 10.1007/s11837-012-0302-8 . ISSN 1047-4838 .
- ^ Ян Л., ван дер Верф КО, Фитье КФ, Беннинк МЛ, Дейкстра П.Дж., Фейен Дж. (март 2008 г.). «Механические свойства нативных и сшитых фибрилл коллагена I типа» . Биофизический журнал . 94 (6): 2204–2211. дои : 10.1529/biophysj.107.111013 . ПМК 2257912 . ПМИД 18032556 .
- ^ Лессен Х.Дж., Флеминг П.Дж., Флеминг К.Г., Содт А.Дж. (август 2018 г.). «Строительные блоки внешней мембраны: расчет общей модели упругой энергии для мембранных белков β-цилиндра» . Журнал химической теории и вычислений . 14 (8): 4487–4497. дои : 10.1021/acs.jctc.8b00377 . ПМК 6191857 . ПМИД 29979594 .
- ^ Мишра В., Хит Р.Дж. (август 2021 г.). «Структурные и биохимические особенности человеческого сывороточного альбумина, необходимого для культуры эукариотических клеток» . Международный журнал молекулярных наук . 22 (16): 8411. doi : 10.3390/ijms22168411 . ПМЦ 8395139 . ПМИД 34445120 .
- ^ Спенсер С.Дж., Ранганатан В.Т., Йетирадж А., Эндрюс Г.Т. (март 2024 г.). «Концентрационная зависимость упругих и вязкоупругих свойств водных растворов фиколла и бычьего сывороточного альбумина методом бриллюэновской спектроскопии светорассеяния». Ленгмюр: Журнал поверхностей и коллоидов ACS . 40 (9): 4615–4622. arXiv : 2309.10967 . doi : 10.1021/acs.langmuir.3c02967 . ПМИД 38387073 .
- ^ Монкос К. (февраль 1996 г.). «Вязкость водных растворов бычьего сывороточного альбумина в зависимости от температуры и концентрации». Международный журнал биологических макромолекул . 18 (1–2): 61–68. дои : 10.1016/0141-8130(95)01057-2 . ПМИД 8852754 .
Дальнейшее чтение
- Учебники
- Брэнден С., Туз Дж. (1999). Введение в структуру белка . Нью-Йорк: Паб Garland. ISBN 978-0-8153-2305-1 .
- Мюррей РФ, Харпер Х.В., Граннер Д.К., Мэйес П.А., Родвелл В.В. (2006). Иллюстрированная биохимия Харпера . Нью-Йорк: Lange Medical Books/McGraw-Hill. ISBN 978-0-07-146197-9 .
- Ван Холде К.Е., Мэтьюз К.К. (1996). Биохимия . Менло-Парк, Калифорния: Паб Benjamin/Cummings. компании, Inc. ISBN 978-0-8053-3931-4 .
Внешние ссылки
 Определения из Викисловаря
Определения из Викисловаря  СМИ из Commons
СМИ из Commons
Базы данных и проекты
- База данных белков NCBI Entrez
- База данных структуры белков NCBI
- Справочная база данных по белкам человека
- Протеинпедия человека
- Folding@Home (Стэнфордский университет). Архивировано 8 сентября 2012 г. в Wayback Machine.
- Банк данных белков в Европе (см. также PDBeQuips , короткие статьи и руководства по интересным структурам PDB)
- Исследовательская лаборатория структурной биоинформатики (см. также «Молекулу месяца», заархивированную 24 июля 2020 г. в Wayback Machine , где представлены краткие отчеты об избранных белках из PDB)
- Proteopedia – Жизнь в 3D : вращающаяся, масштабируемая 3D-модель с вики-аннотациями для каждой известной молекулярной структуры белка.
- UniProt — универсальный ресурс белка
Учебники и образовательные сайты
- «Введение в белки» от HOPES (Проект по борьбе с болезнью Хантингтона для образования в Стэнфорде)
- Белки: от биогенеза до деградации - Виртуальная библиотека биохимии и клеточной биологии