Гемоглобин
| Гемоглобин | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (гетеротетрамер, ( α β ) 2 ) | |||||||||||||
 Структура гемоглобина человека. Субъединицы α- и β- глобина выделены красным и синим цветом соответственно, а железосодержащие гемовые группы — зеленым. Из PDB : 1GZX Протеопедия Гемоглобин | |||||||||||||
| Тип белка | металлопротеин , хромопротеин , глобулин | ||||||||||||
| Функция | кислорода транспорт | ||||||||||||
| Кофактор(ы) | гем (4) | ||||||||||||
| |||||||||||||
Гемоглобин ( гемоглобин , [а] Hb или Hgb ) — белок, содержащий железо , облегчающий транспорт кислорода в эритроцитах . Почти все позвоночные содержат гемоглобин. [3] за исключением рыб семейства Channichthyidae . [4] Гемоглобин в крови переносит кислород от органов дыхания ( легких или жабр ) к другим тканям тела, где он высвобождает кислород, обеспечивая аэробное дыхание веществ животного , которое обеспечивает обмен . У здорового человека приходится от 12 до 20 граммов гемоглобина на каждые 100 мл крови . Гемоглобин — металлопротеин , хромопротеин и глобулин .
У млекопитающих эритроцитов гемоглобин составляет около 96% сухой массы (без воды) и около 35% от общей массы (включая воду). [5] Гемоглобин имеет кислородсвязывающую способность 1,34 мл О 2 на грамм. [6] что увеличивает общую кислородную емкость крови в семьдесят раз по сравнению с растворенным кислородом только в плазме крови. [7] Молекула гемоглобина млекопитающих может связывать и транспортировать до четырех молекул кислорода. [8]
Гемоглобин переносит и другие газы. из организма Он уносит часть дыхательного углекислого газа (около 20–25% от общего количества). [9] как карбаминогемоглобин , в котором CO 2 связывается с белком гема . Молекула также несет важную регуляторную молекулу оксида азота, связанную с тиоловой группой в глобиновом белке, высвобождая ее одновременно с кислородом. [10]
Гемоглобин обнаруживается также в других клетках, в том числе в дофаминергических нейронах А9 , черной субстанции макрофагах , альвеолярных клетках , легких, пигментном эпителии сетчатки, гепатоцитах, мезангиальных клетках почек, клетках эндометрия, клетках шейки матки и эпителиальных клетках влагалища. [11] В этих тканях гемоглобин поглощает ненужный кислород в качестве антиоксиданта и регулирует метаболизм железа . [12] Избыток глюкозы в крови может прикрепиться к гемоглобину и повысить уровень гемоглобина A1c. [13]
Гемоглобин и гемоглобиноподобные молекулы также обнаружены у многих беспозвоночных, грибов и растений. [14] В этих организмах гемоглобины могут переносить кислород или транспортировать и регулировать другие небольшие молекулы и ионы, такие как углекислый газ, оксид азота, сероводород и сульфид. Вариант, называемый легоглобином, служит для удаления кислорода из анаэробных систем, таких как азотфиксирующие клубеньки бобовых растений, предотвращая кислородное отравление.
Заболевание гемоглобинемия , форма анемии , вызвано внутрисосудистым гемолизом , при котором гемоглобин просачивается из эритроцитов в плазму крови .
История исследований [ править ]

В 1825 году Иоганн Фридрих Энгельхарт обнаружил, что соотношение железа и белка одинаково в гемоглобинах нескольких видов. [16] [17] На основании известной атомной массы железа он рассчитал молекулярную массу гемоглобина до n × 16000 ( n = количество атомов железа на молекулу гемоглобина, теперь известно, что оно равно 4), что стало первым определением молекулярной массы белка. Этот «поспешный вывод» вызвал насмешки со стороны коллег, которые не могли поверить, что какая-либо молекула может быть такой большой. Однако Гилберт Смитсон Адэр подтвердил результаты Энгельхарта в 1925 году, измерив осмотическое давление растворов гемоглобина. [18]
Хотя кровь была известна как переносчик кислорода, по крайней мере, с 1794 года. [19] [20] Свойство гемоглобина переносить кислород было описано Хюнефельдом в 1840 году. [21] В 1851 году немецкий физиолог Отто Функе опубликовал серию статей, в которых описал выращивание кристаллов гемоглобина путем последовательного разбавления эритроцитов растворителем, таким как чистая вода, спирт или эфир, с последующим медленным испарением растворителя из полученного раствора белка. [22] [23] Обратимая оксигенация гемоглобина была описана несколько лет спустя Феликсом Хоппе-Сейлером . [24]
С развитием рентгеновской кристаллографии стало возможным секвенировать белковые структуры. [25] В 1959 году Макс Перуц определил молекулярную структуру гемоглобина. [26] [27] За эту работу он разделил Нобелевскую премию по химии 1962 года с Джоном Кендрю , который секвенировал глобулярный белок миоглобин . [25] [28]
Роль гемоглобина в крови выяснил французский физиолог Клод Бернар .
Название гемоглобин происходит от слов «гем» и «глобин» , что отражает тот факт, что каждая субъединица гемоглобина представляет собой глобулярный белок со встроенной гемовой группой. Каждая гемовая группа содержит один атом железа, который может связывать одну молекулу кислорода посредством дипольных сил, индуцированных ионами. Наиболее распространенный тип гемоглобина млекопитающих содержит четыре таких субъединицы. [29]
Генетика [ править ]
Гемоглобин состоит из белковых субъединиц ( молекул глобина ), которые представляют собой полипептиды , длинные свернутые цепи определенных аминокислот , которые определяют химические свойства и функцию белка. Аминокислотная последовательность любого полипептида транслируется с участка ДНК соответствующего гена .
Существует более одного гена гемоглобина. У человека гемоглобин А (основная форма гемоглобина у взрослых) кодируется генами HBA1 , HBA2 и HBB . [30] Субъединицы альфа-1 и альфа-2 соответственно кодируются генами HBA1 и HBA2, расположенными близко друг к другу на хромосоме 16, тогда как бета-субъединица кодируется геном HBB на хромосоме 11. Аминокислотные последовательности субъединиц глобина обычно различаются между видами, причем разница увеличивается. с эволюционной дистанцией. Например, наиболее распространенные последовательности гемоглобина у людей, бонобо и шимпанзе полностью идентичны и содержат одинаковые белковые цепи альфа- и бета-глобина. [31] [32] [33] Гемоглобин человека и гориллы различается по одной аминокислоте как в альфа-, так и в бета-цепях, и эти различия становятся больше между менее близкими видами. [ нужна ссылка ]
Мутации в генах гемоглобина могут привести к появлению вариантов гемоглобина в пределах одного вида, хотя одна последовательность обычно «наиболее распространена» у каждого вида. [34] [35] Многие из этих мутаций не вызывают заболеваний, но некоторые вызывают группу наследственных заболеваний, называемых гемоглобинопатиями . Наиболее известной гемоглобинопатией является серповидно-клеточная анемия , которая была первым заболеванием человека, механизм которого был понятен на молекулярном уровне. В основном отдельный набор заболеваний, называемых талассемией, включает в себя недостаточное производство нормального, а иногда и аномального гемоглобина из-за проблем и мутаций в регуляции генов глобина . Все эти заболевания вызывают анемию . [36]

Вариации последовательностей гемоглобина, как и других белков, могут быть адаптивными. Например, было обнаружено, что гемоглобин по-разному адаптируется к разреженному воздуху на больших высотах, где более низкое парциальное давление кислорода уменьшает его связывание с гемоглобином по сравнению с более высоким давлением на уровне моря. Недавние исследования на мышах-оленях обнаружили мутации в четырех генах, которые могут объяснять различия между популяциями, живущими на высоких и низких высотах. Было обнаружено, что гены обеих пород «практически идентичны, за исключением тех, которые регулируют способность их гемоглобина переносить кислород... Генетическая разница позволяет горным мышам более эффективно использовать кислород». [37] Гемоглобин мамонта имел мутации, которые позволяли доставлять кислород при более низких температурах, что позволило мамонтам мигрировать в более высокие широты во время плейстоцена . [38] Это также было обнаружено у колибри, населяющих Анды. Колибри уже расходуют много энергии и, следовательно, имеют высокую потребность в кислороде, однако было обнаружено, что андские колибри прекрасно себя чувствуют на больших высотах. Несинонимические мутации в гене гемоглобина у нескольких видов, обитающих на больших высотах ( Oreotrochilus, A. castelnaudii, C. violifer, P. gigas и A. viridicuada ), привели к тому, что белок имел меньшее сродство к инозитолгексафосфату (IHP). ), молекула, обнаруженная у птиц и выполняющая ту же роль, что и 2,3-BPG у людей; это приводит к способности связывать кислород при более низких парциальных давлениях. [39]
птиц Уникальная система кровообращения в легких также способствует эффективному использованию кислорода при низком парциальном давлении O 2 . Эти две адаптации усиливают друг друга и объясняют замечательные способности птиц на высоте. [ нужна ссылка ]
Адаптация гемоглобина распространяется и на человека. Среди тибетских женщин с генотипами с высоким насыщением кислородом, проживающих на высоте 4000 м над уровнем моря, выше выживаемость потомства. [40] Естественный отбор, по-видимому, является основной силой, воздействующей на этот ген, поскольку уровень смертности потомства значительно ниже у женщин с более высоким сродством к гемоглобину и кислороду по сравнению с уровнем смертности потомков у женщин с низким сродством к гемоглобину и кислороду. Хотя точный генотип и механизм, посредством которого это происходит, еще не ясен, отбор воздействует на способность этих женщин связывать кислород при низком парциальном давлении, что в целом позволяет им лучше поддерживать важнейшие метаболические процессы. [ нужна ссылка ]
Синтез [ править ]
Гемоглобин (Hb) синтезируется в сложную серию стадий. Гемовая часть синтезируется в ряд стадий в митохондриях и цитозоле незрелых эритроцитов, тогда как глобиновые белковые части синтезируются рибосомами в цитозоле. [41] Производство Hb продолжается в клетке на протяжении всего ее раннего развития от проэритробласта до ретикулоцита в костном мозге . На этом этапе ядро теряется в эритроцитах млекопитающих, но не у птиц и многих других видов. Даже после потери ядра у млекопитающих остаточная рибосомальная РНК позволяет осуществлять дальнейший синтез Hb до тех пор, пока ретикулоцит не потеряет свою РНК вскоре после попадания в сосудистую сеть (эта РНК, синтезирующая гемоглобин, фактически придает ретикулоциту сетчатый вид и название). [42]
Структура гема [ править ]

Гемоглобин имеет четвертичную структуру, характерную для многих многосубъединичных глобулярных белков. [43] Большинство аминокислот в гемоглобине образуют альфа-спирали , и эти спирали соединены короткими неспиральными сегментами. Водородные связи стабилизируют спиральные участки внутри этого белка, вызывая притяжение внутри молекулы, что затем заставляет каждую полипептидную цепь сворачиваться в определенную форму. [44] Четвертичная структура гемоглобина состоит из четырех его субъединиц, расположенных примерно в тетраэдрическом порядке. [43]
У большинства позвоночных молекула гемоглобина представляет собой совокупность четырех субъединиц глобулярного белка . Каждая субъединица состоит из белковой цепи, тесно связанной с небелковой простетической группой гема . Каждая белковая цепь образует набор структурных сегментов альфа-спирали, соединенных вместе в глобиновую складку . Такое название дано потому, что это расположение представляет собой тот же мотив сворачивания, который используется в других белках гема/глобина, таких как миоглобин . [45] [46] Этот складчатый узор содержит карман, который прочно связывает гемовую группу. [ нужна ссылка ]
железа (Fe), Гемовая группа состоит из иона удерживаемого в гетероциклическом кольце, известном как порфирин . Это порфириновое кольцо состоит из четырех молекул пиррола , циклически связанных между собой ( метиновыми мостиками) с ионом железа, связанным в центре. [47] Ион железа, являющийся местом связывания кислорода, координируется с четырьмя атомами азота в центре кольца, которые все лежат в одной плоскости. Гем прочно (ковалентно) связан с глобулярным белком через атомы N имидазольного кольца остатка гистидина F8 (также известного как проксимальный гистидин) под порфириновым кольцом. Шестая позиция может обратимо связывать кислород координатной ковалентной связью . [48] завершает октаэдрическую группу из шести лигандов. Именно эта обратимая связь с кислородом является причиной того, почему гемоглобин так полезен для транспортировки кислорода по организму. [49] Кислород связывается в геометрии «изогнутого конца», когда один атом кислорода связывается с Fe, а другой выступает под углом. Когда кислород не связан, место заполняет очень слабо связанная молекула воды, образуя искаженный октаэдр .
Хотя углекислый газ переносится гемоглобином, он не конкурирует с кислородом за позиции, связывающие железо, а связан с аминогруппами белковых цепей, присоединенными к гемовым группам.
Ион железа может находиться либо в двухвалентном Fe 2+ или в трехвалентном Fe 3+ состоянии, но ферригемоглобин ( метгемоглобин ) (Fe 3+ ) не может связывать кислород. [50] При связывании кислород временно и обратимо окисляется (Fe 2+ ) до (Fe 3+ ), в то время как кислород временно превращается в ион супероксида , поэтому железо должно существовать в степени окисления +2, чтобы связывать кислород. Если супероксид-ион связан с Fe 3+ протонируется, железо гемоглобина останется окисленным и неспособным связывать кислород. В таких случаях фермент метгемоглобинредуктаза сможет в конечном итоге реактивировать метгемоглобин за счет восстановления железного центра.
У взрослых людей наиболее распространенным типом гемоглобина является тетрамер (который содержит четыре субъединицы белка), называемый гемоглобином А , состоящий из двух нековалентно связанных альфа- и двух бета-субъединиц, каждая из которых состоит из 141 и 146 аминокислотных остатков соответственно. Это обозначается как α 2 β 2 . Субъединицы структурно схожи и примерно одинакового размера. Каждая субъединица имеет молекулярную массу около 16 000 дальтон . [51] при общей молекулярной массе тетрамера около 64000 дальтон (64458 г/моль). [52] Таким образом, 1 г/дл=0,1551 ммоль/л. Гемоглобин А — наиболее интенсивно изучаемая из молекул гемоглобина. [ нужна ссылка ]
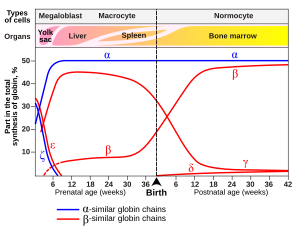
У младенцев молекула фетального гемоглобина состоит из 2 α-цепей и 2 γ-цепей. По мере роста ребенка γ-цепи постепенно заменяются β-цепями. [53]
Четыре полипептидные цепи связаны друг с другом солевыми мостиками , водородными связями и гидрофобным эффектом .
Насыщение кислородом [ править ]
В целом гемоглобин может быть насыщен молекулами кислорода (оксигемоглобин) или денасыщен молекулами кислорода (дезоксигемоглобин). [54]
Оксигемоглобин [ править ]
Оксигемоглобин образуется во время физиологического дыхания , когда кислород связывается с гемовым компонентом белка гемоглобина в эритроцитах. Этот процесс происходит в легочных капиллярах, прилегающих к альвеолам легких. Затем кислород проходит через кровоток и попадает в клетки, где он используется в качестве конечного акцептора электронов при производстве АТФ в процессе окислительного фосфорилирования . Однако это не помогает противодействовать снижению pH крови. Вентиляция или дыхание могут обратить вспять это состояние за счет удаления углекислого газа , вызывая тем самым повышение pH. [55]
Гемоглобин существует в двух формах: тугой (напряженной) форме (Т) и расслабленной форме (R). Различные факторы, такие как низкий уровень pH, высокий уровень CO 2 и высокий уровень 2,3 BPG на уровне тканей, благоприятствуют тугой форме, которая имеет низкое сродство к кислороду и выделяет кислород в тканях. И наоборот, высокий уровень pH, низкий уровень CO 2 или низкий уровень 2,3 BPG благоприятствуют расслабленной форме, которая может лучше связывать кислород. [56] Парциальное давление системы также влияет на сродство к O 2 , где при высоких парциальных давлениях кислорода (например, присутствующих в альвеолах) предпочтительнее расслабленное (высокое сродство, R) состояние. И наоборот, при низком парциальном давлении (например, в дышащих тканях) предпочтительнее напряженное состояние (низкое сродство, Т). [57] Кроме того, связывание кислорода с гемом железа (II) втягивает железо в плоскость порфиринового кольца, вызывая небольшой конформационный сдвиг. Этот сдвиг способствует связыванию кислорода с тремя оставшимися единицами гема в гемоглобине (таким образом, связывание кислорода является кооперативным). [ нужна ссылка ]
Классически считается, что железо в оксигемоглобине существует в степени окисления железа (II). Однако комплекс кислорода с гемовым железом диамагнитен , тогда как и кислород, и высокоспиновое железо(II) парамагнитны . Экспериментальные данные убедительно свидетельствуют о том, что гем-железо находится в степени окисления железа (III) в оксигемоглобине, при этом кислород существует в виде супероксид-аниона (O 2 •− ) или в ковалентном комплексе с переносом заряда. [58]
Дезоксигенированный гемоглобин [ править ]
Дезоксигенированный гемоглобин (дезоксигемоглобин) — это форма гемоглобина без связанного кислорода. Спектры поглощения оксигемоглобина и дезоксигемоглобина различаются. Оксигемоглобин имеет значительно более низкое поглощение при длине волны 660 нм , чем дезоксигемоглобин, а при длине волны 940 нм его поглощение немного выше. Эта разница используется для измерения количества кислорода в крови пациента с помощью прибора, называемого пульсоксиметром . Это различие также объясняет появление цианоза – синего или пурпурного цвета, который появляется в тканях во время гипоксии . [59]
Дезоксигенированный гемоглобин парамагнитен ; он слабо притягивается магнитными полями . [60] [61] Напротив, насыщенный кислородом гемоглобин проявляет диамагнетизм — слабое отталкивание от магнитного поля. [61]
гемоглобина Эволюция позвоночных
Ученые сходятся во мнении, что событие, отделившее миоглобин от гемоглобина, произошло после того, как миноги отделились от челюстных позвоночных . [62] Такое разделение миоглобина и гемоглобина позволило возникнуть и развить различные функции двух молекул: миоглобин больше связан с хранением кислорода, а гемоглобин отвечает за транспортировку кислорода. [63] Гены α- и β-подобного глобина кодируют отдельные субъединицы белка. [30] Предшественники этих генов возникли в результате другого события дупликации, также после того, как общий предок гнатосом произошел от бесчелюстных рыб, примерно 450–500 миллионов лет назад. [62] Исследования реконструкции предков показывают, что предок преддупликации генов α и β представлял собой димер, состоящий из идентичных субъединиц глобина, которые затем эволюционировали, чтобы собраться в тетрамерную архитектуру после дупликации. [64] Развитие генов α и β создало возможность того, что гемоглобин будет состоять из множества отдельных субъединиц, и этот физический состав имеет решающее значение для способности гемоглобина переносить кислород. Наличие нескольких субъединиц способствует способности гемоглобина кооперативно связывать кислород, а также регулировать аллостерически. [63] [64] Впоследствии ген α также подвергся дупликации с образованием генов HBA1 и HBA2 . [65] Эти дальнейшие дупликации и расхождения привели к созданию разнообразного спектра α- и β-подобных глобиновых генов, которые регулируются так, что определенные формы возникают на разных стадиях развития. [63]
Большинство ледяных рыб семейства Channichthyidae утратили гены гемоглобина в результате адаптации к холодной воде. [4]
Кооперативность [ править ]

Когда кислород связывается с комплексом железа, он заставляет атом железа двигаться обратно к центру плоскости порфиринового кольца (см. движущуюся диаграмму). В то же время имидазольная боковая цепь гистидинового остатка, взаимодействующего на другом полюсе железа, притягивается к порфириновому кольцу. Это взаимодействие сдвигает плоскость кольца в сторону внешней стороны тетрамера, а также вызывает растяжение белковой спирали, содержащей гистидин, по мере ее приближения к атому железа. Этот штамм передается остальным трем мономерам тетрамера, где он вызывает аналогичные конформационные изменения в других сайтах гема, так что связывание кислорода с этими сайтами становится легче.
Когда кислород связывается с одним мономером гемоглобина, конформация тетрамера смещается из состояния T (напряженное) в состояние R (расслабленное). Этот сдвиг способствует связыванию кислорода с гемовыми группами остальных трех мономеров, тем самым насыщая молекулу гемоглобина кислородом. [66]
Таким образом, в тетрамерной форме нормального взрослого гемоглобина связывание кислорода представляет собой совместный процесс . Сродство связывания гемоглобина с кислородом увеличивается за счет насыщения молекулы кислородом, при этом первые связанные молекулы кислорода влияют на форму мест связывания последующих молекул, что благоприятно для связывания. Это положительное кооперативное связывание достигается за счет стерических конформационных изменений белкового комплекса гемоглобина, как обсуждалось выше; т.е. когда одна субъединица белка в гемоглобине становится оксигенированной, инициируются конформационные или структурные изменения во всем комплексе, в результате чего другие субъединицы приобретают повышенное сродство к кислороду. Как следствие, кривая связывания кислорода гемоглобином имеет сигмоидальную или S -образную форму, в отличие от нормальной гиперболической кривой, связанной с некооперативным связыванием.
динамический механизм кооперативности в гемоглобине и его связь с низкочастотным резонансом . Обсужден [67]
Связывание лигандов, кислорода от отличных
Помимо кислородного лиганда , который кооперативно связывается с гемоглобином, лиганды гемоглобина также включают конкурентные ингибиторы, такие как окись углерода (CO) и аллостерические лиганды, такие как диоксид углерода (CO 2 ) и оксид азота (NO). Углекислый газ связывается с аминогруппами белков глобина с образованием карбаминогемоглобина ; Считается, что на этот механизм приходится около 10% транспорта углекислого газа у млекопитающих. Оксид азота также может переноситься гемоглобином; он связан со специфическими тиоловыми группами в белке глобина с образованием S-нитрозотиола, который снова диссоциирует на свободный оксид азота и тиол, поскольку гемоглобин высвобождает кислород из своего гема. Предполагается, что этот транспорт оксида азота к периферическим тканям способствует транспортировке кислорода в тканях за счет высвобождения сосудорасширяющего оксида азота в ткани, в которых уровень кислорода низкий. [68]
Конкурсный [ править ]
На связывание кислорода влияют такие молекулы, как окись углерода (например, от курения табака , выхлопных газов и неполного сгорания в печах). CO конкурирует с кислородом в месте связывания гема. Сродство гемоглобина к CO в 250 раз превышает его сродство к кислороду. [69] [70] Поскольку угарный газ представляет собой бесцветный газ без запаха и вкуса и представляет собой потенциально смертельную угрозу, детекторы угарного газа, на рынке появились предупреждающие об опасных уровнях в жилых домах. Когда гемоглобин соединяется с CO, он образует очень ярко-красное соединение, называемое карбоксигемоглобином , из-за которого кожа жертв отравления CO после смерти может выглядеть розовой, а не белой или синей. Когда вдыхаемый воздух содержит уровень CO всего 0,02%, головная боль и тошнота возникают ; если концентрация CO увеличится до 0,1%, наступит потеря сознания. У заядлых курильщиков до 20% кислород-активных участков могут быть заблокированы CO.
Аналогичным образом гемоглобин также обладает конкурентным сродством связывания с цианидом (CN − ), оксид серы (SO) и сульфид (S 2− ), включая сероводород (H 2 S). Все они связываются с железом в геме, не меняя степени его окисления, но, тем не менее, ингибируют связывание кислорода, вызывая серьезную токсичность.
Атом железа в гемовой группе изначально должен находиться в двухвалентной (Fe 2+ ) степень окисления для поддержки связывания и транспорта кислорода и других газов (она временно переключается на железо во время связывания кислорода, как объяснено выше). Первоначальное окисление до трехвалентного железа (Fe 3+ ) состояние без кислорода превращает гемоглобин в «гем и глобин» или метгемоглобин , который не может связывать кислород. Гемоглобин в нормальных эритроцитах защищен системой восстановления, чтобы этого не произошло. Оксид азота способен превращать небольшую часть гемоглобина в метгемоглобин в эритроцитах. Последняя реакция представляет собой остаточную активность более древней диоксигеназной функции оксида азота глобинов.
Аллостерический [ править ]
углерода Оксид занимает другое место связывания на гемоглобине. В тканях, где концентрация углекислого газа выше, углекислый газ связывается с аллостерическим участком гемоглобина, способствуя выгрузке кислорода из гемоглобина и, в конечном итоге, его удалению из организма после высвобождения кислорода в тканях, подвергающихся метаболизму. Это повышенное сродство венозной крови к углекислому газу известно как эффект Бора . Через фермент карбоангидразу углекислый газ реагирует с водой с образованием угольной кислоты , которая распадается на бикарбонат и протоны :
- CO 2 + H 2 O → H 2 CO 3 → HCO 3 − + Ч +
Следовательно, кровь с высоким уровнем углекислого газа также имеет более низкий pH (более кислая ). Гемоглобин способен связывать протоны и углекислый газ, что вызывает конформационные изменения белка и облегчает выделение кислорода. Протоны связываются в различных местах белка, а углекислый газ связывается с α-аминогруппой. [71] Углекислый газ связывается с гемоглобином и образует карбаминогемоглобин . [72] Это снижение сродства гемоглобина к кислороду за счет связывания углекислого газа и кислоты известно как эффект Бора . Эффект Бора благоприятствует состоянию T, а не состоянию R. O 2 (смещает кривую насыщения вправо ). И наоборот, когда уровень углекислого газа в крови снижается (т. е. в легочных капиллярах), углекислый газ и протоны высвобождаются из гемоглобина, увеличивая сродство белка к кислороду. Снижение общей способности гемоглобина связывать кислород (т.е. смещение кривой вниз, а не только вправо) из-за снижения pH называется корневым эффектом . Это наблюдается у костистых рыб.
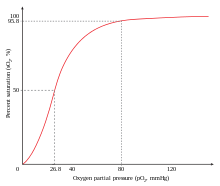
Гемоглобину необходимо высвободить кислород, который он связывает; в противном случае нет смысла связывать его. Сигмоидальная кривая гемоглобина делает его эффективным в связывании (захват О 2 в легких) и эффективным в разгрузке (выгрузка О 2 в тканях). [73]
У людей, акклиматизированных к большой высоте, в крови повышена концентрация 2,3-бисфосфоглицерата (2,3-БФГ), что позволяет этим лицам доставлять к тканям большее количество кислорода в условиях пониженного напряжения кислорода . Это явление, когда молекула Y влияет на связывание молекулы X с транспортной молекулой Z, называется гетеротропным аллостерическим эффектом. Гемоглобин в организмах, живущих на больших высотах, также адаптировался таким образом, что он имеет меньшее сродство к 2,3-BPG, и поэтому белок будет больше сдвинут в сторону своего R-состояния. В состоянии R гемоглобин будет легче связывать кислород, что позволяет организмам выполнять необходимые метаболические процессы, когда кислород присутствует при низких парциальных давлениях. [74]
Животные, кроме человека, используют разные молекулы для связывания с гемоглобином и изменения его сродства к О 2 при неблагоприятных условиях. Рыбы используют как АТФ , так и ГТФ . Они связываются с фосфатным «карманом» на молекуле гемоглобина рыбы, что стабилизирует напряженное состояние и, следовательно, снижает сродство к кислороду. [75] ГТФ снижает сродство гемоглобина к кислороду в гораздо большей степени, чем АТФ, что, как полагают, происходит из-за образования дополнительной водородной связи , которая дополнительно стабилизирует напряженное состояние. [76] В условиях гипоксии концентрация АТФ и ГТФ в эритроцитах рыб снижается, что увеличивает сродство к кислороду. [77]
Вариант гемоглобина, называемый фетальным гемоглобином (HbF, α 2 γ 2 ), обнаруживается у развивающегося плода и связывает кислород с большим сродством, чем гемоглобин взрослого человека. Это означает, что кривая связывания кислорода для фетального гемоглобина сдвинута влево (т.е. более высокий процент гемоглобина имеет связанный с ним кислород при более низком напряжении кислорода) по сравнению с кривой гемоглобина взрослого человека. В результате кровь плода в плаценте способна забирать кислород из материнской крови.
Гемоглобин также несет оксид азота (NO) в глобиновой части молекулы. Это улучшает доставку кислорода на периферию и способствует контролю дыхания. NO обратимо связывается со специфическим остатком цистеина в глобине; связывание зависит от состояния (R или T) гемоглобина. Образующийся S-нитрозилированный гемоглобин влияет на различные действия, связанные с NO, такие как контроль сосудистого сопротивления, артериальное давление и дыхание. NO не высвобождается в цитоплазму эритроцитов, а транспортируется из них анионообменником, называемым AE1 . [78]
Виды гемоглобина у человека [ править ]
Варианты гемоглобина являются частью нормального развития эмбриона и плода . Они также могут быть патологическими мутантными формами гемоглобина в популяции , вызванными генетическими вариациями. Некоторые хорошо известные варианты гемоглобина, такие как серповидно-клеточная анемия , ответственны за заболевания и считаются гемоглобинопатиями . Другие варианты не вызывают обнаруживаемой патологии и поэтому считаются непатологическими вариантами. [34] [79]
У эмбрионов :
- Гауэр 1 (ζ 2 ε 2 ).
- Гауэр 2 (α 2 ε 2 ) ( PDB : 1A9W ).
- Гемоглобин Портланд I (ζ 2 γ 2 ).
- Гемоглобин Портланд II (ζ 2 β 2 ).
У плодов:
- Гемоглобин F (α 2 γ 2 ) ( PDB : 1FDH ).
У новорожденных (новорожденных сразу после рождения):
- Гемоглобин А (взрослый гемоглобин) (α 2 β 2 ) ( PDB : 1BZ0 ) – наиболее распространенный с нормальным количеством более 95%.
- Гемоглобин A 2 (α 2 δ 2 ) – синтез δ-цепи начинается в конце третьего триместра, и у взрослых его нормальный уровень составляет 1,5–3,5%.
- Гемоглобин F (фетальный гемоглобин) (α 2 γ 2 ). У взрослых гемоглобин F ограничен ограниченной популяцией эритроцитов, называемых F-клетками. Однако уровень Hb F может быть повышен у лиц с серповидноклеточной анемией и бета-талассемией .
Аномальные формы, встречающиеся при заболеваниях:
- Гемоглобин D – (α 2 β Д 2 ) – вариантная форма гемоглобина.
- Гемоглобин H (β 4 ) – вариантная форма гемоглобина, образованная тетрамером β-цепей, которая может присутствовать при вариантах α-талассемии .
- Гемоглобин Бартс (γ 4 ) – вариант формы гемоглобина, образованный тетрамером γ-цепей, который может присутствовать при вариантах α-талассемии.
- Гемоглобин S (α 2 β С 2 ) – Вариант формы гемоглобина, обнаруженный у людей с серповидноклеточной анемией. Существует вариация гена β-цепи, вызывающая изменение свойств гемоглобина, что приводит к серповидному образованию эритроцитов.
- Гемоглобин С (α 2 β С 2 ) – Другой вариант, обусловленный вариацией гена β-цепи. Этот вариант вызывает легкую хроническую гемолитическую анемию .
- Гемоглобин E (α 2 β И 2 ) – Другой вариант, обусловленный вариацией гена β-цепи. Этот вариант вызывает легкую хроническую гемолитическую анемию.
- Гемоглобин AS - гетерозиготная форма, вызывающая серповидноклеточный признак с одним взрослым геном и одним геном серповидноклеточной анемии.
- Гемоглобиновая болезнь SC – сложная гетерозиготная форма с одним серповидным геном и другим, кодирующим гемоглобин C.
- Гемоглобин Хопкинса-2 – вариант формы гемоглобина, который иногда рассматривают в сочетании с гемоглобином S для возникновения серповидноклеточной анемии.
позвоночных животных Деградация
Когда эритроциты достигают конца своей жизни из-за старения или дефектов, они удаляются из кровообращения за счет фагоцитарной активности макрофагов в селезенке или печени или гемолизируются в кровообращении. Затем свободный гемоглобин выводится из кровообращения посредством переносчика гемоглобина CD163 , который экспрессируется исключительно на моноцитах или макрофагах. Внутри этих клеток молекула гемоглобина расщепляется, и железо перерабатывается. Этот процесс также производит одну молекулу окиси углерода на каждую молекулу деградировавшего гема. [80] Деградация гема является единственным естественным источником окиси углерода в организме человека и отвечает за нормальный уровень окиси углерода в крови у людей, дышащих обычным воздухом. [81]
Другим основным конечным продуктом деградации гема является билирубин . Повышенные уровни этого химического вещества обнаруживаются в крови, если эритроциты разрушаются быстрее, чем обычно. Неправильно расщепленный белок гемоглобина или гемоглобин, который слишком быстро высвобождается из клеток крови, может закупорить мелкие кровеносные сосуды, особенно чувствительные сосуды почек, фильтрующие кровь , вызывая повреждение почек. Железо удаляется из гема и сохраняется для дальнейшего использования. Оно сохраняется в тканях в виде гемосидерина или ферритина и транспортируется в плазму бета-глобулинами в виде трансферринов . Когда порфириновое кольцо разрушается, его фрагменты обычно выделяются в виде желтого пигмента, называемого билирубином, который выделяется в кишечник в виде желчи. В кишечнике билирубин метаболизируется в уробилиноген. Уробилиноген покидает организм с фекалиями в виде пигмента стеркобилина. Глобулин метаболизируется до аминокислот, которые затем высвобождаются в кровоток.
[ править ]
Дефицит гемоглобина может быть вызван либо уменьшением количества молекул гемоглобина, как при анемии , либо снижением способности каждой молекулы связывать кислород при том же парциальном давлении кислорода. Гемоглобинопатии (генетические дефекты, приводящие к аномальной структуре молекулы гемоглобина) [82] может вызвать и то, и другое. В любом случае дефицит гемоглобина снижает способность крови переносить кислород . Дефицит гемоглобина обычно строго отличают от гипоксемии , определяемой как снижение парциального давления кислорода в крови. [83] [84] [85] [86] хотя оба являются причинами гипоксии (недостаточного снабжения тканей кислородом).
Другие распространенные причины низкого гемоглобина включают потерю крови, дефицит питательных веществ, проблемы с костным мозгом, химиотерапию, почечную недостаточность или аномальный гемоглобин (например, серповидно-клеточную анемию).
Способность каждой молекулы гемоглобина переносить кислород обычно модифицируется изменением pH крови или CO 2 , что приводит к изменению кривой диссоциации кислорода и гемоглобина . Однако он также может быть патологически изменен, например, при отравлении угарным газом .
Снижение гемоглобина с абсолютным уменьшением эритроцитов или без него приводит к симптомам анемии. Анемия имеет множество различных причин, хотя дефицит железа и возникающая из него железодефицитная анемия являются наиболее распространенными причинами в западном мире. Поскольку отсутствие железа снижает синтез гема, эритроциты при железодефицитной анемии являются гипохромными (без красного пигмента гемоглобина) и микроцитарными (меньше нормального размера). Другие анемии встречаются реже. При гемолизе (ускоренном распаде эритроцитов) желтуха вызывается метаболитом гемоглобина билирубином, а циркулирующий гемоглобин может вызвать почечную недостаточность .
Некоторые мутации в цепи глобина связаны с гемоглобинопатиями , такими как серповидноклеточная анемия и талассемия . Другие мутации, как обсуждалось в начале статьи, являются доброкачественными и называются просто вариантами гемоглобина .
Существует группа генетических нарушений, известных как порфирии , которые характеризуются нарушениями метаболических путей синтеза гема. Король Соединенного Королевства Георг III был, вероятно, самым известным больным порфирией.
В небольшой степени гемоглобин А медленно соединяется с глюкозой на концевом валине (альфа-аминокислоте) каждой β-цепи. Полученную молекулу часто называют Hb A 1c , гликированный гемоглобин . Связывание глюкозы с аминокислотами в гемоглобине происходит спонтанно (без помощи фермента) во многих белках и, как известно, не служит полезной цели. Однако по мере увеличения концентрации глюкозы в крови увеличивается процент Hb A, который превращается в Hb A 1c . У диабетиков , у которых уровень глюкозы обычно высокий, процент Hb A 1c также повышается. Из-за медленной скорости сочетания Hb A с глюкозой процент Hb A 1c отражает средневзвешенное значение уровня глюкозы в крови за время жизни эритроцитов, которое составляет примерно 120 дней. [87] Поэтому уровни гликированного гемоглобина измеряют для мониторинга долгосрочного контроля хронического заболевания сахарного диабета 2 типа (СД2). Плохой контроль СД2 приводит к высокому уровню гликированного гемоглобина в эритроцитах. Нормальный референтный диапазон составляет примерно 4,0–5,9%. Хотя получить данные трудно, людям с СД2 рекомендуются значения менее 7%. Уровни более 9% связаны с плохим контролем гликированного гемоглобина, а уровни более 12% связаны с очень плохим контролем. Диабетики, у которых уровень гликированного гемоглобина близок к 7%, имеют гораздо больше шансов избежать осложнений, которые могут сопровождать диабет (чем те, у кого уровень гликированного гемоглобина составляет 8% или выше). [88] Кроме того, повышенное гликирование гемоглобина увеличивает его сродство к кислороду, тем самым предотвращая его высвобождение в тканях и вызывая в крайних случаях уровень гипоксии. [89]
Повышенный уровень гемоглобина связан с увеличением количества или размера эритроцитов, что называется полицитемией . Это повышение может быть вызвано врожденным пороком сердца , легочным сердцем , фиброзом легких , избытком эритропоэтина или истинной полицитемией . [90] Высокий уровень гемоглобина также может быть вызван пребыванием на большой высоте, курением, обезвоживанием (искусственно путем концентрации гемоглобина), прогрессирующим заболеванием легких и некоторыми опухолями. [53]
Недавнее исследование, проведенное в Пондичерри, Индия, показывает его важность при ишемической болезни сердца. [91]
Диагностическое использование [ править ]

Измерение концентрации гемоглобина является одним из наиболее часто выполняемых анализов крови , обычно как часть общего анализа крови . Например, его обычно тестируют до или после сдачи крови . Результаты сообщаются в г / л , г/ дл или моль /л. 1 г/дл равен примерно 0,6206 ммоль/л, хотя последние единицы используются не так часто из-за неопределенности относительно полимерного состояния молекулы. [92] Этот коэффициент преобразования, использующий молекулярную массу одной единицы глобина 16 000 Да , чаще встречается для концентрации гемоглобина в крови. Для MCHC (средняя концентрация корпускулярного гемоглобина) более распространенным является коэффициент преобразования 0,155, в котором используется масса тетрамера 64 500 Да. [93] Нормальные уровни:
- Мужчины: от 13,8 до 18,0 г/дл (от 138 до 180 г/л или от 8,56 до 11,17 ммоль/л).
- Женщины: от 12,1 до 15,1 г/дл (от 121 до 151 г/л или от 7,51 до 9,37 ммоль/л)
- Дети: от 11 до 16 г/дл (от 110 до 160 г/л или от 6,83 до 9,93 ммоль/л)
- Беременные женщины: от 11 до 14 г/дл (от 110 до 140 г/л или от 6,83 до 8,69 ммоль/л) (обычное значение во время беременности от 9,5 до 15). [94] [б]
Нормальные значения гемоглобина в 1-м и 3-м триместрах у беременных должны составлять не менее 11 г/дл и не менее 10,5 г/дл во 2-м триместре. [96]
Обезвоживание или гипергидратация могут сильно влиять на измеряемый уровень гемоглобина. Альбумин может указывать на состояние гидратации.
Если концентрация ниже нормы, это называется анемией. Анемии классифицируются по размеру эритроцитов — клеток, содержащих гемоглобин у позвоночных. Анемия называется «микроцитарной», если эритроциты маленькие, «макроцитарной», если они большие, и «нормоцитарной» в противном случае.
Гематокрит , доля объема крови, занимаемая эритроцитами, обычно примерно в три раза превышает концентрацию гемоглобина, измеренную в г/дл. Например, если уровень гемоглобина составляет 17 г/дл, это соответствует гематокриту 51%. [97]
Лабораторные методы определения гемоглобина требуют забора крови (артериальной, венозной или капиллярной) и проведения анализа на гематологическом анализаторе и СО-оксиметре. Кроме того, доступен новый метод неинвазивного определения гемоглобина (SpHb) под названием «Пульсовая CO-оксиметрия», точность которого сравнима с инвазивными методами. [98]
Концентрации окси- и дезоксигемоглобина можно измерять непрерывно, регионально и неинвазивно с помощью NIRS . [99] [100] [101] [102] [103] БИКС можно использовать как на голове, так и на мышцах. Этот метод часто используется для исследований, например, в спортивной подготовке, эргономике, реабилитации, мониторинге пациентов, неонатальных исследованиях, функциональном мониторинге мозга, интерфейсе мозг-компьютер , урологии (сокращение мочевого пузыря), неврологии (нейроваскулярная связь) и многом другом.
Массу гемоглобина у людей можно измерить с помощью нерадиоактивного метода повторного дыхания угарным газом (CO), который используется уже более 100 лет. При использовании этого метода небольшой объем чистого газа CO вдыхается и вдыхается повторно в течение нескольких минут. Во время повторного дыхания CO связывается с гемоглобином, присутствующим в эритроцитах. По увеличению содержания CO в крови после периода повторного дыхания можно определить массу гемоглобина по принципу разведения. Хотя газ CO в больших объемах токсичен для человека, объем CO, используемый для оценки объемов крови, соответствует тому, который можно вдыхать при курении сигареты. В то время как исследователи обычно используют специальные дыхательные контуры, Detalo Performance от Detalo Health автоматизировала процедуру и сделала измерения доступными для более широкой группы пользователей. [104]
Долгосрочный контроль концентрации сахара в крови можно измерить по концентрации Hb A 1c . Для его непосредственного измерения потребуется много образцов, поскольку уровень сахара в крови сильно варьируется в течение дня. Hb A 1c – продукт необратимой реакции гемоглобина А с глюкозой. глюкозы Более высокая концентрация приводит к увеличению количества Hb A 1c . Поскольку реакция протекает медленно, доля Hb A 1c представляет собой уровень глюкозы в крови, усредненный за период полураспада эритроцитов, который обычно составляет ~ 120 дней. Доля Hb A 1c 6,0% или менее свидетельствует о хорошем долгосрочном контроле уровня глюкозы, тогда как значения выше 7,0% являются повышенными. Этот тест особенно полезен диабетикам. [с]
Аппарат функциональной магнитно-резонансной томографии (фМРТ) использует сигнал дезоксигемоглобина, который чувствителен к магнитным полям, поскольку является парамагнитным. Комбинированное измерение с помощью NIRS показывает хорошую корреляцию как с сигналом окси-, так и с дезоксигемоглобином по сравнению с сигналом, выделенным ЖИРНЫМ шрифтом . [105]
Использование спортивного отслеживания и самоотслеживания [ править ]
Гемоглобин можно отслеживать неинвазивно, чтобы создать индивидуальный набор данных, отслеживающий эффекты гемоконцентрации и гемодилюции в повседневной деятельности, для лучшего понимания спортивных результатов и тренировок. Спортсмены часто обеспокоены выносливостью и интенсивностью тренировок. Датчик использует светоизлучающие диоды, которые излучают красный и инфракрасный свет через ткань на фотодетектор, который затем отправляет сигнал на процессор для расчета поглощения света белком гемоглобина. [106] Этот датчик похож на пульсоксиметр , который состоит из небольшого сенсорного устройства, прикрепляемого к пальцу.
Аналоги у беспозвоночных организмов [ править ]
В организмах животного и растительного царств существует множество белков, переносящих и связывающих кислород. Организмы, включая бактерии , простейшие и грибы, содержат гемоглобиноподобные белки, известная и предсказанная роль которых включает обратимое связывание газообразных лигандов . Поскольку многие из этих белков содержат глобины и гемовую часть (железо в плоской порфириновой подложке), их часто называют гемоглобинами, даже если их общая третичная структура сильно отличается от структуры гемоглобина позвоночных. В частности, различение «миоглобина» и гемоглобина у низших животных часто невозможно, так как некоторые из этих организмов не содержат мышц . Или у них может быть узнаваемая отдельная система кровообращения , но не та, которая занимается транспортировкой кислорода (например, у многих насекомых и других членистоногих ). Во всех этих группах гем/глобинсодержащие молекулы (даже мономерные глобиновые), которые связывают газы, называются оксигемоглобинами. Помимо транспорта и определения кислорода, они также могут иметь дело с NO, CO. 2 , сульфидные соединения и даже улавливание O 2 в средах, которые должны быть анаэробными. [107] Они могут даже заниматься детоксикацией хлорированных материалов аналогично гемсодержащим ферментам P450 и пероксидазам.

Структура гемоглобина варьируется у разных видов. Гемоглобин встречается во всех царствах организмов, но не во всех организмах. Примитивные виды, такие как бактерии, простейшие, водоросли и растения, часто содержат одноглобиновые гемоглобины. Многие нематоды , моллюски и ракообразные содержат очень крупные многосубъединичные молекулы, намного крупнее, чем у позвоночных. В частности, химерные гемоглобины, обнаруженные у грибов и гигантских кольчатых червей, могут содержать как глобин, так и другие типы белков. [14]
Одно из наиболее ярких проявлений и применений гемоглобина в организмах произошло у гигантского трубчатого червя ( Riftia pachyptila , также называемого Vestimentifera), который может достигать 2,4 метра в длину и населяет жерла океанских вулканов . Вместо пищеварительного тракта эти черви содержат популяцию бактерий, составляющую половину веса организма. Бактерии окисляют H 2 S из вентиляционного отверстия с помощью O 2 из воды, чтобы произвести энергию для приготовления пищи из H 2 O и CO 2 . Верхний конец червей представляет собой темно-красную веерообразную структуру («шлейф»), которая простирается в воду и поглощает H 2 S и O 2 для бактерий, а также CO 2 для использования в качестве синтетического сырья, аналогичного фотосинтезирующим растениям. . Структуры ярко-красного цвета из-за содержания в них нескольких чрезвычайно сложных гемоглобинов, которые имеют до 144 глобиновых цепей, каждая из которых включает связанные с ними гемовые структуры. Эти гемоглобины замечательны тем, что способны переносить кислород в присутствии сульфида и даже переносить сульфид, не подвергаясь при этом полному «отравлению» или ингибированию им, как это происходит с гемоглобинами большинства других видов. [108] [109]
кислородсвязывающие белки Другие
- Миоглобин
- Обнаруженный в мышечной ткани многих позвоночных, включая человека, он придает мышечной ткани отчетливый красный или темно-серый цвет. По структуре и последовательности он очень похож на гемоглобин, но не является тетрамером; вместо этого это мономер, у которого отсутствует кооперативное связывание. Он используется для хранения кислорода, а не для его транспортировки.
- гемоцианин
- Второй по распространенности белок, переносящий кислород, обнаруженный в природе, он содержится в крови многих членистоногих и моллюсков. Использует простетические группы меди вместо гемовых групп железа и имеет синий цвет при насыщении кислородом.
- Гемеритрин
- Некоторые морские беспозвоночные и некоторые виды кольчатых червей используют этот железосодержащий негемовый белок для переноса кислорода в крови. При насыщении кислородом становится розовым/фиолетовым, в отсутствие кислорода — прозрачным.
- хлорокруорин
- Обнаруженный у многих кольчатых червей, он очень похож на эритрокруорин, но гемовая группа существенно отличается по структуре. Имеет зеленый цвет при деоксигенации и красный при насыщении кислородом.
- Привычный
- Также известные как ванадия хромагены , они обнаружены в крови асцидий . Когда-то была выдвинута гипотеза об использовании металлического ванадия в качестве простетической группы, связывающей кислород. Однако, хотя они преимущественно содержат ванадий, они, по-видимому, связывают мало кислорода и, таким образом, выполняют какую-то другую функцию, которая не выяснена (морские асцидии также содержат некоторое количество гемоглобина). Они могут действовать как токсины.
- Эритрокруорин
- Обнаруженный у многих кольчатых червей, включая дождевых червей , это гигантский свободно плавающий белок крови, содержащий многие десятки — возможно, сотни — железо- и гемосодержащих белковых субъединиц, связанных вместе в единый белковый комплекс с молекулярной массой более 3,5 миллионов дальтон.
- Леггемоглобин
- У бобовых растений, таких как люцерна или соевые бобы, азотфиксирующие бактерии в корнях защищены от кислорода с помощью гема железа, содержащего кислородсвязывающий белок. Специфическим защищенным ферментом является нитрогеназа , которая не способна восстанавливать газообразный азот в присутствии свободного кислорода.
- Кобоглобин
- Синтетический порфирин на основе кобальта. Кобопротеин будет бесцветным при насыщении кислородом, но желтым в венах.
в неэритроидных Присутствие клетках
Некоторые неэритроидные клетки (т.е. клетки, отличные от линии эритроцитов) содержат гемоглобин. В головном мозге к ним относятся дофаминергические нейроны А9 в черной субстанции , астроциты в коре головного мозга и гиппокампе , а также во всех зрелых олигодендроцитах . [12] Было высказано предположение, что гемоглобин мозга в этих клетках может обеспечивать «накопление кислорода для обеспечения гомеостатического механизма в аноксических условиях, что особенно важно для нейронов A9 DA, которые имеют повышенный метаболизм с высокой потребностью в производстве энергии». [12] Далее было отмечено, что « дофаминергические нейроны А9 могут подвергаться особому риску аноксической дегенерации, поскольку в дополнение к их высокой митохондриальной активности они испытывают интенсивный окислительный стресс, вызванный выработкой перекиси водорода посредством автоокисления и/или моноаминоксидазы (МАО)». опосредованное дезаминирование дофамина и последующая реакция доступного двухвалентного железа с образованием высокотоксичных гидроксильных радикалов». [12] Это может объяснить риск дегенерации этих клеток при болезни Паркинсона . [12] Железо, полученное из гемоглобина в этих клетках, не является причиной посмертного потемнения этих клеток (происхождение латинского названия, черная субстанция ), а скорее обусловлено нейромеланином .
Вне мозга гемоглобин выполняет функции, не связанные с переносом кислорода, в качестве антиоксиданта и регулятора метаболизма железа в макрофагах . [110] альвеолярные клетки , [111] и мезангиальные клетки почки. [112]
В истории, искусстве и музыке [ править ]

Исторически ассоциация между цветом крови и ржавчины возникает при ассоциации планеты Марс с римским богом войны, поскольку планета оранжево-красного цвета, что напоминало древним о крови. Хотя цвет планеты обусловлен соединениями железа в сочетании с кислородом в марсианской почве, распространено заблуждение, что железо в гемоглобине и его оксидах придает крови красный цвет. Цвет на самом деле обусловлен порфириновой частью гемоглобина, с которой связано железо, а не самим железом. [113] хотя лигирование и окислительно-восстановительное состояние железа могут влиять на электронные переходы pi-pi* или n-pi* порфирина и, следовательно, на его оптические характеристики.
Художник Джулиан Восс-Андрее в 2005 году создал скульптуру под названием «Стальное сердце (гемоглобин)» , основанную на белковом остове. Скульптура была изготовлена из стекла и атмосферостойкой стали . Намеренное ржавление первоначально блестящего произведения искусства отражает фундаментальную химическую реакцию гемоглобина, связанную с связыванием кислорода с железом. [114] [115]
Монреальский художник Николя Байер создал Lustre (Hémoglobine) — скульптуру из нержавеющей стали, показывающую структуру молекулы гемоглобина. Он выставлен в атриуме исследовательского центра Центра здоровья Университета Макгилла в Монреале. Размеры скульптуры составляют примерно 10 × 10 × 10 метров. [116] [117]
См. также [ править ]
- Карбаминогемоглобин (Hb, связанный с CO 2 )
- Карбоксигемоглобин (Hb, связанный с CO)
- Хлорофилл (гем магния)
- Общий анализ крови
- Дельта-глобус
- Гемоглобинометр
- гемопротеин
- Метгемоглобин (железо Hb или ферригемоглобин)
- Оксигемоглобин (с двухатомным кислородом , окрашенный в кроваво-красный цвет)
- Tegillarca granosa – «кровавый моллюск».
- Комплекс Васьки – металлоорганический комплекс иридия, отличающийся способностью связываться с О 2 . обратимо
Ссылки [ править ]
Примечания [ править ]
- ^ от греческого слова αἷμα, haîma «кровь» + латинская глобула «шар, сфера» + -in ; / ˌ h iː m ə ˈ ɡ l oʊ b ɪ n , ˈ h ɛ m oʊ ˌ -/ [1] [2]
- ^ Хотя другие источники могут иметь немного отличающиеся значения, например, Британский «Справочник общей практики». [95]
- ^ Этот уровень Hb A 1c полезен только для людей, у которых эритроциты (эритроциты) имеют нормальную выживаемость (т. е. нормальный период полураспада). У людей с аномальными эритроцитами, будь то из-за аномальных молекул гемоглобина (например, гемоглобина S при серповидно-клеточной анемии), дефектов мембраны эритроцитов или других проблем, период полувыведения эритроцитов часто сокращается. У этих людей можно использовать альтернативный тест, называемый «уровень фруктозамина». Он измеряет степень гликирования (связывания глюкозы) с альбумином, наиболее распространенным белком крови, и отражает средний уровень глюкозы в крови за предыдущие 18–21 день, что представляет собой период полураспада молекул альбумина в кровообращении.
Источники [ править ]
- ^ Уэллс, Джон К. (2008). Словарь произношения Лонгмана (3-е изд.). Лонгман. ISBN 978-1-4058-8118-0 .
- ^ Джонс, Дэниел (2011). Роуч, Питер ; Сеттер, Джейн ; Эслинг, Джон (ред.). Кембриджский словарь английского произношения (18-е изд.). Издательство Кембриджского университета. ISBN 978-0-521-15255-6 .
- ^ Матон, Антея; Джин Хопкинс; Чарльз Уильям Маклафлин; Сьюзан Джонсон; Марианна Куон Уорнер; Дэвид ЛаХарт; Джилл Д. Райт (1993). Биология человека и здоровье . Энглвуд Клиффс, Нью-Джерси, США: Прентис Холл. ISBN 978-0-13-981176-0 .
- ↑ Перейти обратно: Перейти обратно: а б Сиделл, Брюс; Кристин О'Брайен (2006). «Когда с хорошей рыбой случаются плохие вещи: потеря экспрессии гемоглобина и миоглобина у антарктических ледяных рыб». Журнал экспериментальной биологии . 209 (Часть 10): 1791–802. дои : 10.1242/jeb.02091 . ПМИД 16651546 . S2CID 29978182 .
- ^ Виид, Роберт И.; Рид, Клод Ф.; Берг, Джордж (1963). «Является ли гемоглобин важным структурным компонентом мембран эритроцитов человека?» . Джей Клин Инвест . 42 (4): 581–88. дои : 10.1172/JCI104747 . ПМК 289318 . ПМИД 13999462 .
- ^ Домингес де Вильота ЭД, Руис Кармона МТ, Рубио Х.Дж., де Андрес С. (1981). «Равенство кислородсвязывающей способности гемоглобина in vivo и in vitro у больных с тяжелыми респираторными заболеваниями» . Бр Джей Анест . 53 (12): 1325–28. дои : 10.1093/бья/53.12.1325 . ПМИД 7317251 . S2CID 10029560 .
- ^ Роудс, Карл Э.; Дено, Дина; Варакалло, Мэтью (04 марта 2019 г.). Физиология, Транспорт кислорода . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 30855920 . Архивировано из оригинала 27 августа 2021 г. Проверено 4 мая 2019 г. - через NCBI Bookshelf.
- ^ Костанцо, Линда С. (2007). Физиология . Хагерствон, доктор медицины: Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-7311-9 .
- ^ Паттон, Кевин Т. (10 февраля 2015 г.). Анатомия и физиология . Elsevier Науки о здоровье. ISBN 978-0-323-31687-3 . Архивировано из оригинала 26 апреля 2016 г. Проверено 9 января 2016 г.
- ^ Эпштейн, Ф.Х.; Ся, CCW (1998). «Дыхательная функция гемоглобина». Медицинский журнал Новой Англии . 338 (4): 239–47. дои : 10.1056/NEJM199801223380407 . ПМИД 9435331 .
- ^ Саха, Д.; Редди, КВР; Патгаонкар, М.; Айяр, К.; Башир, Т.; Шрофф, А. (2014). «Экспрессия гемоглобина в неэритроидных клетках: новая или повсеместная?» . Международный журнал воспаления . 2014 (803237): 1–8. дои : 10.1155/2014/803237 . ПМК 4241286 . ПМИД 25431740 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Бьяджоли М., Пинто М., Чеселли Д. и др. (2009). «Неожиданная экспрессия альфа- и бета-глобина в мезэнцефалических дофаминергических нейронах и глиальных клетках» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (36): 15454–59. Бибкод : 2009PNAS..10615454B . дои : 10.1073/pnas.0813216106 . ПМЦ 2732704 . ПМИД 19717439 .
- ^ «Анализ крови» . Национальный институт сердца, легких и крови (NHLBI) . Архивировано из оригинала 9 апреля 2019 г. Проверено 27 апреля 2019 г.
- ↑ Перейти обратно: Перейти обратно: а б Вебер Р.Э., Виноградов С.Н. (2001). «Гемоглобины беспозвоночных: функции и молекулярные адаптации». Физиол. Преподобный . 81 (2): 569–628. дои : 10.1152/physrev.2001.81.2.569 . ПМИД 11274340 . S2CID 10863037 .
- ^ «Макс Перуц, отец молекулярной биологии, умер в возрасте 87 лет» . Нью-Йорк Таймс . 8 февраля 2002 г. Архивировано из оригинала 23 апреля 2016 г.
- ^ Энгельхарт, Иоганн Фридрих (1825). Комментарий о природе истинного вещества крови, придающего ей малиновый цвет (на латыни). Геттинген: Дитрих. Архивировано из оригинала 16 июня 2020 г. Проверено 16 июня 2020 г.
- ^ «Энгельгард и Роуз о красящем веществе крови» . Эдинбургский медицинский и хирургический журнал . 27 (90): 95–102. 1827. ЧВК 5763191 . ПМИД 30330061 .
- ^ Адэр, Гилберт Смитсон (1925). «Критическое исследование прямого метода измерения осмотического давления гемоглобина» . Учеб. Р. Сок. Лонд . 108 (750): 292–300. Бибкод : 1925RSPSA.109..292A . дои : 10.1098/rspa.1925.0126 .
- ^ Парри, CH (1794). Письма доктора Уизеринга,... доктора Юарта,... доктора Тортона... и доктора Биггса... вместе с некоторыми другими статьями, дополняющими две публикации об астме, чахотке, лихорадке и других заболеваниях, Т. Беддос . п. 43. Архивировано из оригинала 31 января 2022 г. Проверено 30 ноября 2021 г.
- ^ Беддос, Т. (1796). Соображения о медицинском использовании и производстве искусственных эфиров: Часть I. Томас Беддоес, доктор медицинских наук. Часть II. Джеймс Ватт, инженер; «Часть 1, раздел 2, «О дыхании человека и знакомых животных» . Балгин и Россер. Часть 1, стр. 9–13. Архивировано из оригинала 31 января 2022 г. Проверено 30 ноября 2021 г.
- ^ Хюнефельд, Фридрих Людвиг (1840). Химизм в организации животных (на немецком языке). Лейпциг: Ф.А. Брокгауз. Архивировано из оригинала 14 апреля 2021 года . Проверено 26 февраля 2021 г.
- ^ Искра О (1851). «О крови селезеночной вены». З Крыса Мед . 1 : 172-218.
- ^ «Рецепт НАСА для кристаллографии белков» (PDF) . Образовательная записка . Национальное управление по аэронавтике и исследованию космического пространства. Архивировано из оригинала (PDF) 10 апреля 2008 г. Проверено 12 октября 2008 г.
- ^ Хоппе-Зейлер Ф (1866). «Об окислении в живой крови». Лаборатория медико-химических исследований . 1 :133–40.
- ↑ Перейти обратно: Перейти обратно: а б Стоддарт, Шарлотта (1 марта 2022 г.). «Структурная биология: как белки оказались крупным планом» . Знающий журнал . doi : 10.1146/knowable-022822-1 . Проверено 25 марта 2022 г.
- ^ Перуц, МФ; Россманн, МГ; Каллис, А.Ф.; Мюрхед, Х.; Уилл, Г.; Норт, АКТ (1960). «Структура гемоглобина: трехмерный синтез Фурье с разрешением 5,5 А, полученный с помощью рентгеновского анализа». Природа . 185 (4711): 416–22. Бибкод : 1960Natur.185..416P . дои : 10.1038/185416a0 . ПМИД 18990801 . S2CID 4208282 .
- ^ Перуц М.Ф. (1960). «Структура гемоглобина». Брукхейвенские симпозиумы по биологии . 13 : 165–83. ПМИД 13734651 .
- ^ де Чадаревян, Сорая (июнь 2018 г.). «Джон Кендрю и миоглобин: определение структуры белка в 1950-х годах: Джон Кендрю и миоглобин» . Белковая наука . 27 (6): 1136–1143. дои : 10.1002/pro.3417 . ПМК 5980623 . ПМИД 29607556 .
- ^ Ниси, Хироши; Инаги, Рэйко; Като, Хидеки; Танемото, Масаюки; Кодзима, Ичиро; Сын Дайсуке; Фудзита, Тосиро; Нангаку, Масаоми (август 2008 г.). «Гемоглобин экспрессируется мезангиальными клетками и снижает окислительный стресс» . Журнал Американского общества нефрологов: JASN . 19 (8): 1500–1508. дои : 10.1681/ASN.2007101085 . ISSN 1046-6673 . ПМЦ 2488266 . ПМИД 18448584 .
- ↑ Перейти обратно: Перейти обратно: а б Хардисон, Росс К. (1 декабря 2012 г.). «Эволюция гемоглобина и его генов» . Перспективы Колд-Спринг-Харбора в медицине . 2 (12): а011627. doi : 10.1101/cshperspect.a011627 . ISSN 2157-1422 . ПМК 3543078 . ПМИД 23209182 .
- ^ Оффнер, Сьюзен (01 апреля 2010 г.). «Использование баз данных генома NCBI для сравнения генов бета-гемоглобина человека и шимпанзе» . Американский учитель биологии . 72 (4): 252–56. дои : 10.1525/ок.2010.72.4.10 . ISSN 0002-7685 . S2CID 84499907 . Архивировано из оригинала 26 декабря 2019 г. Проверено 26 декабря 2019 г.
- ^ «HBB - бета-субъединица гемоглобина - Pan paniscus (карликовый шимпанзе) - ген и белок HBB» . uniprot.org . Архивировано из оригинала 01 августа 2020 г. Проверено 10 марта 2020 г.
- ^ «HBA1 - альфа-субъединица гемоглобина - пантроглодиты (шимпанзе) - ген и белок HBA1» . uniprot.org . Архивировано из оригинала 01 августа 2020 г. Проверено 10 марта 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б Хьюсман THJ (1996). «Программа вариантов человеческого гемоглобина» . Генный сервер Глобина . Государственный университет Пенсильвании. Архивировано из оригинала 11 декабря 2008 г. Проверено 12 октября 2008 г.
- ^ Варианты гемоглобина. Архивировано 5 ноября 2006 г. в Wayback Machine . Labtestsonline.org. Проверено 5 сентября 2013 г.
- ^ Усман, Эд. «Гемоглобинопатии и талассемии» . Архивировано из оригинала 15 декабря 2007 г. Проверено 26 декабря 2007 г.
- ^ Рид, Лесли. «Адаптация обнаружена в генах мыши». Omaha World-Herald , 11 августа 2009 г.: EBSCO. [ нужна страница ]
- ^ «У мамонтов была «незамерзающая» кровь» . Би-би-си. 2 мая 2010 г. Архивировано из оригинала 4 мая 2010 г. Проверено 2 мая 2010 г.
- ^ Прожекто-Гарсия, Джоана; Натараджан, Чандрасекхар; Морияма, Хидеаки; Вебер, Рой Э.; Фаго, Анжела; Чевирон, Закари А.; Дадли, Роберт; Макгуайр, Джимми А.; Витт, Кристофер К. (17 декабря 2013 г.). «Повторяющиеся переходы по высоте в функции гемоглобина в ходе эволюции андских колибри» . Труды Национальной академии наук . 110 (51): 20669–74. Бибкод : 2013PNAS..11020669P . дои : 10.1073/pnas.1315456110 . ISSN 0027-8424 . ПМЦ 3870697 . ПМИД 24297909 .
- ^ Билл, Синтия М.; Сон, Киджон; Элстон, Роберт С.; Гольдштейн, Мелвин К. (28 сентября 2004 г.). «Более высокая выживаемость потомства у тибетских женщин с генотипами с высоким насыщением кислородом, проживающих на высоте 4000 м» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (39): 14300–04. Бибкод : 2004PNAS..10114300B . дои : 10.1073/pnas.0405949101 . ISSN 0027-8424 . ПМК 521103 . ПМИД 15353580 .
- ^ «Синтез гемоглобина» . 14 апреля 2002 года. Архивировано из оригинала 26 декабря 2007 года . Проверено 26 декабря 2007 г.
- ^ Бурка, Эдвард (1969). «Особенности деградации РНК в эритроидной клетке» . Журнал клинических исследований . 48 (7): 1266–72. дои : 10.1172/jci106092 . ПМЦ 322349 . ПМИД 5794250 .
- ↑ Перейти обратно: Перейти обратно: а б Ван Кессель, Ганс (2002). «Белки – природные полиамиды». Нельсон Химия 12 . Торонто: Томсон. п. 122. ИСБН 978-0-17-625986-0 .
- ^ «Учебник по гемоглобину». Архивировано 26 ноября 2009 г. в Массачусетском университете Wayback Machine в Амхерсте. Веб. 23 октября 2009 г.
- ^ Стейнберг, МЗ (2001). Нарушения гемоглобина: генетика, патофизиология и клиническое лечение . Издательство Кембриджского университета. п. 95. ИСБН 978-0-521-63266-9 . Архивировано из оригинала 17 ноября 2016 г. Проверено 18 февраля 2016 г.
- ^ Хардисон, Р.К. (1996). «Краткая история гемоглобинов: растений, животных, простейших и бактерий» . Proc Natl Acad Sci США . 93 (12): 5675–79. Бибкод : 1996PNAS...93.5675H . дои : 10.1073/pnas.93.12.5675 . ПМК 39118 . ПМИД 8650150 .
- ^ «Гемоглобин». Архивировано 13 ноября 2009 г. в Wayback Machine Школе химии Бристольского университета, Великобритания. Веб. 12 октября 2009 г.
- ^ WikiPremed> Координационная химия. Архивировано 23 августа 2009 г. в Wayback Machine . Проверено 2 июля 2009 г.
- ^ Фундаментальная биология (2015). «Клетки крови» . Архивировано из оригинала 18 июля 2021 г. Проверено 27 марта 2020 г.
- ^ Линберг Р., Коновер К.Д., Шум К.Л., Шорр Р.Г. (1998). «Переносчики кислорода на основе гемоглобина: сколько метгемоглобина — это слишком много?» . Кровозаменитель Artif Cells Immobil Biotechnol . 26 (2): 133–48. дои : 10.3109/10731199809119772 . ПМИД 9564432 .
- ^ Гемоглобин. Архивировано 15 марта 2017 г. в Wayback Machine . Worthington-biochem.com. Проверено 5 сентября 2013 г.
- ^ Ван Биквелт MC, Кольер В.Н., Веверс Р.А., Ван Энгелен Б.Г. (2001). «Эффективность ближней инфракрасной спектроскопии для измерения местного потребления O2 и кровотока в скелетных мышцах». J Appl Physiol . 90 (2): 511–19. дои : 10.1152/яп.2001.90.2.511 . ПМИД 11160049 . S2CID 15468862 .
- ↑ Перейти обратно: Перейти обратно: а б «Гемоглобин». Архивировано 24 января 2012 г. в Wayback Machine MedicineNet. Веб. 12 октября 2009 г.
- ^ "Дом гемоглобина". Архивировано 1 декабря 2009 г. в Wayback Machine Biology @ Davidson. Веб. 12 октября 2009 г.
- ^ «График насыщения гемоглобином» . высота.орг. Архивировано из оригинала 31 августа 2010 г. Проверено 6 июля 2010 г.
- ^ Кинг, Майкл В. «Страница медицинской биохимии - гемоглобин» . Архивировано из оригинала 4 марта 2012 г. Проверено 20 марта 2012 г.
- ^ Воэт, Д. (2008) Основы биохимии , 3-е. ред., рис. 07_06, John Wiley & Sons. ISBN 0470129301
- ^ Шикама К. (2006). «Природа связи FeO2 в миоглобине и гемоглобине: новая молекулярная парадигма» . Прога Биофиз Мол Биол . 91 (1–2): 83–162. doi : 10.1016/j.pbiomolbio.2005.04.001 . ПМИД 16005052 .
- ^ Аренс; Кимберли, Бэшам (1993). Основы оксигенации: значение для клинической практики . Джонс и Бартлетт Обучение. п. 194. ИСБН 978-0-86720-332-5 .
- ^ Огава, С; Менон, RS; Танк, ДВ; Ким, СГ; Меркл, Х; Эллерманн, Дж. М.; Угурбил, К. (1993). «Функциональное картирование мозга с помощью контрастной магнитно-резонансной томографии, зависящей от уровня оксигенации крови. Сравнение характеристик сигнала с биофизической моделью» . Биофизический журнал . 64 (3): 803–12. Бибкод : 1993BpJ....64..803O . дои : 10.1016/S0006-3495(93)81441-3 . ПМЦ 1262394 . ПМИД 8386018 .
- ↑ Перейти обратно: Перейти обратно: а б Брен К.Л., Айзенберг Р., Грей Х.Б. (2015). «Открытие магнитного поведения гемоглобина: начало бионеорганической химии» . Proc Natl Acad Sci США . 112 (43): 13123–27. Бибкод : 2015PNAS..11213123B . дои : 10.1073/pnas.1515704112 . ПМЦ 4629386 . ПМИД 26508205 .
- ↑ Перейти обратно: Перейти обратно: а б Гудман, Моррис; Мур, Дж. Уильям; Мацуда, Гэндзи (20 февраля 1975 г.). «Дарвиновская эволюция в генеалогии гемоглобина». Природа . 253 (5493): 603–08. Бибкод : 1975Natur.253..603G . дои : 10.1038/253603a0 . ПМИД 1089897 . S2CID 2979887 .
- ↑ Перейти обратно: Перейти обратно: а б с Сторц, Джей Ф.; Опазо, Хуан К.; Хоффманн, Федерико Г. (01 февраля 2013 г.). «Дупликация генов, дупликация генома и функциональное разнообразие глобинов позвоночных» . Молекулярная филогенетика и эволюция . 66 (2): 469–78. Бибкод : 2013МОЛПЭ..66..469С . дои : 10.1016/j.ympev.2012.07.013 . ISSN 1095-9513 . ПМК 4306229 . ПМИД 22846683 .
- ↑ Перейти обратно: Перейти обратно: а б Пиллаи, Арвинд С.; Чендлер, Шейн А.; Лю, Ян; Синьор, Энтони В.; Кортес-Ромеро, Карлос Р.; Бенеш, Джастин Л.П.; Лагановский, Артур; Сторц, Джей Ф.; Хохберг, Георг К.А.; Торнтон, Джозеф В. (май 2020 г.). «Происхождение сложности эволюции гемоглобина» . Природа . 581 (7809): 480–85. Бибкод : 2020Natur.581..480P . дои : 10.1038/s41586-020-2292-y . ISSN 1476-4687 . ПМЦ 8259614 . ПМИД 32461643 . S2CID 218761566 .
- ^ Циммер Э.А., Мартин С.Л., Беверли С.М., Кан Ю.В., Уилсон AC (1 апреля 1980 г.). «Быстрое дупликация и потеря генов, кодирующих альфа-цепи гемоглобина» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (4): 2158–62. Бибкод : 1980PNAS...77.2158Z . дои : 10.1073/pnas.77.4.2158 . ISSN 0027-8424 . ПМЦ 348671 . ПМИД 6929543 .
- ^ Михайлеску, Михаэла-Рита; Руссу, Ирина М. (27 марта 2001 г.). «Признак перехода T → R в гемоглобине человека» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (7): 3773–77. Бибкод : 2001PNAS...98.3773M . дои : 10.1073/pnas.071493598 . ISSN 0027-8424 . ПМК 31128 . ПМИД 11259676 .
- ^ Чжоу К.С. (1989). «Низкочастотный резонанс и кооперативность гемоглобина». Тенденции биохимии. Наука . 14 (6): 212–13. дои : 10.1016/0968-0004(89)90026-1 . ПМИД 2763333 .
- ^ Дженсен, Фрэнк Б. (2009). «Двойная роль эритроцитов в доставке кислорода к тканям: переносчики кислорода и регуляторы местного кровотока». Журнал экспериментальной биологии . 212 (Часть 21): 3387–93. дои : 10.1242/jeb.023697 . ПМИД 19837879 . S2CID 906177 .
- ^ Холл, Джон Э. (2010). Учебник Гайтона и Холла по медицинской физиологии (12-е изд.). Филадельфия, Пенсильвания: Сондерс/Эльзевир. п. 502. ИСБН 978-1-4160-4574-8 .
- ^ Роудс, Карл Э.; Дено, Дина; Варакалло, Мэтью (04 марта 2019 г.). Физиология, Транспорт кислорода . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 30855920 . Архивировано из оригинала 27 августа 2021 г. Проверено 4 мая 2019 г. - через NCBI Bookshelf.
Важно отметить, что на фоне карбоксигемоглобинемии причиной патологии является не снижение кислородной способности, а нарушение доставки связанного кислорода к тканям-мишеням.
- ^ Нельсон, DL; Кокс, ММ (2000). Ленингерские принципы биохимии , 3-е изд. Нью-Йорк: Издательство Worth. п. 217, ISBN 1572599316 .
- ^ Гайтон, Артур К.; Джон Э. Холл (2006). Учебник медицинской физиологии (11-е изд.). Филадельфия: Эльзевир Сондерс. п. 511. ИСБН 978-0-7216-0240-0 .
- ^ Лекция - 12 миоглобина и гемоглобина на YouTube.
- ^ Биохимия (Восьмое изд.). Нью-Йорк: WH Freeman. 2015. ISBN 978-1-4641-2610-9 . [ нужна страница ]
- ^ Рутьес, штат Ха; Нивин, MC; Вебер, Р.Э.; Витте, Ф.; Ван ден Тилларт, GEEJM (20 июня 2007 г.). «Множественные стратегии цихлид озера Виктория по борьбе с пожизненной гипоксией включают переключение гемоглобина». AJP: Регуляторная, интегративная и сравнительная физиология . 293 (3): R1376–83. дои : 10.1152/ajpregu.00536.2006 . ПМИД 17626121 .
- ^ Гроненборн, Анджела М.; Клор, Г.Мариус; Брунори, Маурицио; Джардина, Бруно; Фальчиони, Джанкарло; Перуц, Макс Ф. (1984). «Стереохимия АТФ и ГТФ, связанных с гемоглобинами рыб». Журнал молекулярной биологии . 178 (3): 731–42. дои : 10.1016/0022-2836(84)90249-3 . ПМИД 6492161 .
- ^ Вебер, Рой Э.; Фрэнк Б. Дженсен (1988). «Функциональные адаптации гемоглобинов экзотермных позвоночных». Ежегодный обзор физиологии . 50 : 161–79. дои : 10.1146/annurev.ph.50.030188.001113 . ПМИД 3288089 .
- ^ Ранг, HP; Дейл ММ; Риттер Дж.М.; Мур ПК (2003). Фармакология, пятое издание . Эльзевир. ISBN 978-0-443-07202-4 . [ нужна страница ]
- ^ «Варианты гемоглобина» . Лабораторные тесты онлайн . Американская ассоциация клинической химии. 10 ноября 2007 г. Архивировано из оригинала 20 сентября 2008 г. Проверено 12 октября 2008 г.
- ^ Кикучи, Г.; Ёсида, Т.; Ногучи, М. (2005). «Гемоксигеназа и деградация гема». Связь с биохимическими и биофизическими исследованиями . 338 (1): 558–67. дои : 10.1016/j.bbrc.2005.08.020 . ПМИД 16115609 .
- ^ Кумс, Маргарита В. (2011). «Аминокислоты и метаболизм гема». В Девлине, Томас М. (ред.). Учебник биохимии: с клиническими корреляциями (7-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. п. 797. ИСБН 978-0-470-28173-4 .
- ^ «гемоглобинопатия» в Медицинском словаре Дорланда.
- ^ гипоксемия. Архивировано 2 февраля 2009 г. в Wayback Machine . Британская энциклопедия , констатирующая гипоксемию (снижение напряжения кислорода в крови) .
- ^ Biology-Online.org --> Словарь » H » Гипоксемия. Архивировано 21 ноября 2009 г. в Wayback Machine , последнее изменение - 29 декабря 2008 г.
- ^ Уильям, К. Уилсон; Гранде, Кристофер М.; Хойт, Дэвид Б. (2007). «Патофизиология острой дыхательной недостаточности» . Травма, Том II: Критическая терапия . Тейлор и Фрэнсис. п. 430. ИСБН 978-1-4200-1684-0 . Архивировано из оригинала 17 ноября 2016 г. Проверено 18 февраля 2016 г.
- ^ Макгаффиган, Пенсильвания (1996). «Опасности гипоксемии: как защитить пациента от низкого уровня кислорода» . Уход . 26 (5): 41–46, викторина 46. doi : 10.1097/00152193-199626050-00013 . ПМИД 8710285 .
- ^ «НГСП: HbA1c и eAG» . ngsp.org . Архивировано из оригинала 15 октября 2015 г. Проверено 28 октября 2015 г.
- ^ «Определение гликозилированного гемоглобина». Архивировано 23 января 2014 г. в Wayback Machine Medicine Net. Веб. 12 октября 2009 г.
- ^ Мэдсен, Х; Дитцель, Дж (1984). «Транспорт кислорода в крови в первом триместре диабетической беременности». Acta Obstetricia et Gynecologica Scandinavica . 63 (4): 317–20. дои : 10.3109/00016348409155523 . ПМИД 6741458 . S2CID 12771673 .
- ^ Гемоглобин. Архивировано 10 июня 2016 г. в Wayback Machine в Medline Plus.
- ^ Падманабан, П.; Тоора, Б. (2011). «Гемоглобин: новый маркер стабильной ишемической болезни сердца» . Хроники молодых ученых . 2 (2): 109. дои : 10.4103/2229-5186.82971 .
- ^ Общество биомедицинских исследований диабета. Калькулятор преобразования единиц СИ. Архивировано 9 марта 2013 г. на Wayback Machine .
- ^ Хандин, Роберт И.; Люкс, Сэмюэл Э. и СтоссельБлад, Томас П. (2003). Кровь: принципы и практика гематологии . Липпинкотт Уильямс и Уилкинс, ISBN 0781719933
- ^ «Тест уровня гемоглобина» . Ibdcrohns.about.com . 16 августа 2013 г. Архивировано из оригинала 29 января 2007 г. Проверено 5 сентября 2013 г.
- ^ «гемоглобин (референтный диапазон)» . Тетрадь общей практики . Архивировано из оригинала 25 сентября 2009 г.
- ^ Мюррей СС и МакКинни ES (2006). Основы ухода за матерями и новорожденными . 4-е изд., с. 919. Филадельфия: Сондерс Эльзевир. ISBN 1416001417 .
- ^ «Гематокрит (HCT) или объем упакованных клеток (PCV)» . DoctorsLounge.com. Архивировано из оригинала 02 января 2008 г. Проверено 26 декабря 2007 г.
- ^ Фраска, Д.; Дайо-Физелье, К.; Кэтрин, К.; Леврат, К.; Дебен, Б.; Мимоз, О. (2011). «Точность непрерывного неинвазивного мониторирования гемоглобина у пациентов отделений интенсивной терапии*». Медицина критических состояний . 39 (10): 2277–82. дои : 10.1097/CCM.0b013e3182227e2d . ПМИД 21666449 . S2CID 205541592 .
- ^ Феррари, М.; Бинцони, Т.; Куарезима, В. (1997). «Окислительный метаболизм в мышцах» . Философские труды Королевского общества B: Биологические науки . 352 (1354): 677–83. Бибкод : 1997RSPTB.352..677F . дои : 10.1098/rstb.1997.0049 . ПМК 1691965 . ПМИД 9232855 .
- ^ Мэдсен, Польша; Сечер, Нью-Хэмпшир (1999). «Ближняя инфракрасная оксиметрия головного мозга». Прогресс нейробиологии . 58 (6): 541–60. дои : 10.1016/S0301-0082(98)00093-8 . ПМИД 10408656 . S2CID 1092056 .
- ^ Маккалли, КК; Хамаока, Т. (2000). «Спектроскопия ближнего инфракрасного диапазона: что она может сказать нам о насыщении кислородом скелетных мышц?». Обзоры физических упражнений и спортивных наук . 28 (3): 123–27. ПМИД 10916704 .
- ^ Перри, СП (2008). «Неинвазивная БИК-спектроскопия функции мозга человека во время тренировки». Методы . 45 (4): 289–99. дои : 10.1016/j.ymeth.2008.04.005 . ПМИД 18539160 .
- ^ Рольф, П. (2000). «Инвивонар-Инфракрасная спектроскопия». Ежегодный обзор биомедицинской инженерии . 2 : 715–54. doi : 10.1146/annurev.bioeng.2.1.715 . ПМИД 11701529 .
- ^ Зибенманн, Кристоф; Кайзер, Стефани; Робах, Пол; Лундбю, Карстен (29 июня 2017 г.). «CORP: Оценка общей массы гемоглобина по повторному дыханию угарного газа» . Журнал прикладной физиологии . 123 (3): 645–654. doi : 10.1152/japplphysicalol.00185.2017 . ISSN 8750-7587 . ПМИД 28663373 .
- ^ Механьул-Шиппер DJ, ван дер Каллен Б.Ф., Кольер В.Н., ван дер Слейс MC, ван Эрнинг Л.Дж., Тийссен Х.О., Озебург Б., Хофнагельс В.Х., Янсен Р.В. (2002). «Одновременные измерения изменений оксигенации головного мозга во время активации мозга с помощью ближней инфракрасной спектроскопии и функциональной магнитно-резонансной томографии у здоровых молодых и пожилых людей» . Карта мозга Hum . 16 (1): 14–23. дои : 10.1002/hbm.10026 . ПМК 6871837 . ПМИД 11870923 .
- ^ «Cercacor - Как работает неинвазивная технология гемоглобина Ember» . Technology.cercacor.com . Архивировано из оригинала 4 ноября 2016 г. Проверено 3 ноября 2016 г.
- ^ Л. Инт Панис; Б. Годдерис; Р. Верхейен (1995). «Концентрация гемоглобина личинок Chironomus cf.Plumosus L. (Diptera: Chironomidae) из двух сточных местообитаний» . Нидерландский журнал водной экологии . 29 (1): 1–4. Бибкод : 1995NJAqE..29....1P . дои : 10.1007/BF02061785 . S2CID 34214741 . Архивировано из оригинала 5 сентября 2018 г. Проверено 10 ноября 2013 г.
- ^ Зал Ф., Лалье Ф.Х., Грин Б.Н., Виноградов С.Н., Тулмонд А. (1996). «Мультигемоглобиновая система гидротермального жерлового червя Riftia pachyptila. II. Полный состав полипептидной цепи исследован методом анализа максимальной энтропии масс-спектров» . Ж. Биол. Хим . 271 (15): 8875–81. дои : 10.1074/jbc.271.15.8875 . ПМИД 8621529 .
- ^ Миник З., Эрве Г. (2004). «Биохимические и ферментологические аспекты симбиоза глубоководного трубчатого червя Riftia pachyptila и его бактериального эндосимбионта» . Евро. Дж. Биохим . 271 (15): 3093–102. дои : 10.1111/j.1432-1033.2004.04248.x . ПМИД 15265029 .
- ^ Лю Л., Цзэн М., Стамлер Дж.С. (1999). «Индукция гемоглобина в макрофагах мыши» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (12): 6643–47. Бибкод : 1999PNAS...96.6643L . дои : 10.1073/pnas.96.12.6643 . ЧВК 21968 . ПМИД 10359765 .
- ^ Ньютон Д.А., Рао К.М., Длуги Р.А., Баатц Дж.Э. (2006). «Гемоглобин экспрессируется альвеолярными эпителиальными клетками» . Журнал биологической химии . 281 (9): 5668–76. дои : 10.1074/jbc.M509314200 . ПМИД 16407281 .
- ^ Ниши, Х.; Инаги, Р.; Като, Х.; Танемото, М.; Кодзима, И.; Сон, Д.; Фудзита, Т.; Нангаку, М. (2008). «Гемоглобин экспрессируется мезангиальными клетками и снижает окислительный стресс» . Журнал Американского общества нефрологов . 19 (8): 1500–08. дои : 10.1681/ASN.2007101085 . ПМЦ 2488266 . ПМИД 18448584 .
- ^ Бо, Ларри (2001). Руководство по фармацевтической практике: Руководство по клиническому опыту . Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-2541-5 .
- ^ Холден, Констанс (2005). «Кровь и сталь». Наука . 309 (5744): 2160. doi : 10.1126/science.309.5744.2160d . S2CID 190178048 .
- ^ Моран Л., Хортон Р.А., Скримджер Дж., Перри М. (2011). Принципы биохимии . Бостон, Массачусетс: Пирсон. п. 127. ИСБН 978-0-321-70733-8 .
- ^ Генри, Шон (7 августа 2014 г.). «Взгляните на коллекцию произведений искусства MUHC» . Новости Си-Би-Си. Архивировано из оригинала 5 февраля 2016 года . Проверено 1 февраля 2016 г.
- ^ «Блеск (Гемоглобин) 2014» . Арт Паблик Монреаль . Монреаль. Архивировано из оригинала 1 февраля 2016 года . Проверено 1 февраля 2016 г.
Дальнейшее чтение [ править ]
- Кэмпбелл, МК (1999). Биохимия (третье изд.). Харкорт. ISBN 978-0-03-024426-1 .
- Эшагян, С; Хорвич, ТБ; Фонаров, GC (2006). «Неожиданная обратная зависимость между уровнями HbA1c и смертностью у пациентов с диабетом и прогрессирующей систолической сердечной недостаточностью». Ам Харт Дж . 151 (1): 91.e1–91.e6. дои : 10.1016/j.ahj.2005.10.008 . ПМИД 16368297 .
- Ганонг, ВФ (2003). Обзор медицинской физиологии (21-е изд.). Ланге. ISBN 978-0-07-140236-1 .
- Хагер, Т. (1995). Сила природы: Жизнь Лайнуса Полинга . Саймон и Шустер. ISBN 978-0-684-80909-0 .
- Хейзелвуд, Лорен (2001) Не могу жить без него: история гемоглобина в болезни и в здоровье , Nova Science Publishers ISBN 1-56072-907-4
- Кнайпп Дж., Балакришнан Г., Чен Р., Шен Т.Дж., Саху С.К., Хо НТ, Джованнелли Дж.Л., Симплачану В., Хо С., Спиро Т. (2005). «Динамика аллостерии в гемоглобине: роль предпоследних H-связей тирозина». Дж Мол Биол . 356 (2): 335–53. дои : 10.1016/j.jmb.2005.11.006 . ПМИД 16368110 .
- Хардисон, Росс К. (2012). «Эволюция гемоглобина и его генов» . Перспективы Колд-Спринг-Харбора в медицине . 2 (12): а011627. doi : 10.1101/cshperspect.a011627 . ISSN 2157-1422 . ПМК 3543078 . ПМИД 23209182 .
Внешние ссылки [ править ]
- Протеопедия Гемоглобин
- Национальный совет по действиям по анемии на сайте anemia.org
- Новый тип гемоглобина вызывает ложный диагноз с помощью пульсоксиметров. Архивировано 9 марта 2016 г. на сайте Wayback Machine на www.life-of-science.net. Архивировано 7 марта 2017 г. на Wayback Machine.
- Анимация гемоглобина: от дезокси-к окси-форме на vimeo.com