хлорофилл



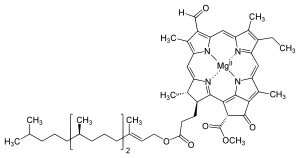
Хлорофилл — это любой из нескольких родственных зеленых пигментов, в цианобактериях и в хлоропластах водорослей обнаруженных и растений . [2] Его название происходит от греческих слов χλωρός , хлорос («бледно-зеленый») и φύλλον , филлон («лист»). [3] Хлорофилл позволяет растениям поглощать энергию света.
Хлорофиллы наиболее сильно поглощают свет в синей части электромагнитного спектра , а также в красной части. [4] И наоборот, он плохо поглощает зеленую и околозеленую части спектра. Следовательно, ткани, содержащие хлорофилл, кажутся зелеными, потому что зеленый свет, диффузно отражаемый такими структурами, как клеточные стенки, меньше поглощается. [1] В фотосистемах зеленых растений существуют два типа хлорофилла: хлорофилл а и б . [5]
История [ править ]
Хлорофилл был впервые выделен и назван Жозефом Бьенеме Каванту и Пьером Жозефом Пеллетье в 1817 году. [6] Присутствие магния в хлорофилле было обнаружено в 1906 г. [7] и было первым обнаружением этого элемента в живой ткани. [8]
После первоначальной работы, проделанной немецким химиком Рихардом Вильштеттером с 1905 по 1915 год, общая структура хлорофилла а была выяснена Гансом Фишером в 1940 году. К 1960 году, когда большая часть стереохимии хлорофилла а была известна, Роберт Бернс Вудворд опубликовал полный синтез. молекулы. [8] [9] В 1967 году последнее оставшееся стереохимическое объяснение было завершено Яном Флемингом . [10] а в 1990 году Вудворд и соавторы опубликовали обновленный синтез. [11] было объявлено, что хлорофилл f присутствует в цианобактериях и других оксигенных микроорганизмах, образующих строматолиты ; В 2010 году [12] [13] молекулярная формула C 55 H 70 O 6 N 4 Mg и строение (2- формил )-хлорофилла а . На основе ЯМР, оптического и масс-спектра установлены [14]
Фотосинтез [ править ]

Хлорофилл жизненно важен для фотосинтеза , который позволяет растениям поглощать энергию света . [15]
Молекулы хлорофилла располагаются внутри и вокруг фотосистем , встроенных в тилакоидные мембраны хлоропластов . [16] В этих комплексах хлорофилл выполняет три функции:
- Функция подавляющего большинства хлорофиллов (до нескольких сотен молекул на фотосистему) — поглощение света.
- При этом те же центры выполняют свою вторую функцию: передачу этой энергии путем резонансной передачи энергии определенной паре хлорофиллов в реакционном центре фотосистем.
- Эта специфическая пара выполняет последнюю функцию хлорофиллов: разделение зарядов, в результате чего образуются несвязанные протоны (H + ) и электроны (e − ), которые по отдельности стимулируют биосинтез.
Двумя принятыми в настоящее время единицами фотосистемы являются фотосистема I и фотосистема II , которые имеют свои собственные реакционные центры, называемые P700 и P680 соответственно. Эти центры названы в честь длины волны (в нанометрах ) их максимума поглощения красного пика. Идентичность, функции и спектральные свойства типов хлорофилла в каждой фотосистеме различны и определяются друг другом и окружающей их белковой структурой.
Функция реакционного центра хлорофилла — поглощать световую энергию и передавать ее другим частям фотосистемы. Поглощенная энергия фотона передается электрону в процессе, называемом разделением зарядов. Отрыв электрона от хлорофилла является реакцией окисления. Хлорофилл отдает электрон высокой энергии ряду молекулярных промежуточных продуктов, называемых цепью переноса электронов . Заряженный реакционный центр хлорофилла (P680 + ) затем восстанавливается обратно в основное состояние, принимая электрон, оторванный от воды. Электрон, восстанавливающий P680 + в конечном итоге происходит в результате окисления воды до O 2 и H + через несколько промежуточных звеньев. Эта реакция — то, как фотосинтезирующие организмы, такие как растения, производят газ O 2 и является источником практически всего O 2 в атмосфере Земли. Фотосистема I обычно работает последовательно с Фотосистемой II; таким образом, P700 + Фотосистемы I обычно восстанавливается, поскольку она принимает электрон через множество промежуточных продуктов в мембране тилакоида электронами, поступающими, в конечном счете, из Фотосистемы II. Однако реакции переноса электрона в тилакоидных мембранах сложны, и источник электронов, используемый для восстановления P700 + может варьироваться.
Поток электронов, создаваемый пигментами хлорофилла реакционного центра, используется для перекачки H. + ионы через тилакоидную мембрану, создавая протондвижущую силу, хемиосмотический потенциал, используемый главным образом для производства АТФ (запасенной химической энергии) или для восстановления НАДФ. + к НАДФН . НАДФН – универсальный агент, используемый для восстановления CO 2 до сахаров, а также в других реакциях биосинтеза.
Комплексы хлорофилл-белок реакционного центра способны напрямую поглощать свет и осуществлять процессы разделения заряда без помощи других пигментов хлорофилла, но вероятность того, что это произойдет при заданной интенсивности света, мала. Таким образом, все остальные хлорофиллы в фотосистеме и белки-пигменты антенн совместно поглощают и направляют световую энергию в реакционный центр. Помимо хлорофилла а и другие пигменты, называемые добавочными пигментами , в этих антенных комплексах пигмент-белок встречаются .
Химическая структура [ править ]

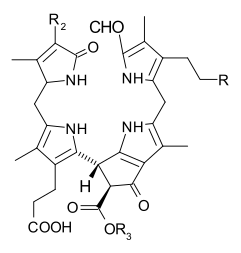
Известно несколько хлорофиллов. Все они определяются как производные исходного хлорина по наличию пятого кетонсодержащего кольца помимо четырех пирролоподобных колец. Большинство хлорофиллов классифицируются как хлорины , которые являются восстановленными родственниками порфиринов (обнаруженных в гемоглобине ). Они имеют общий путь биосинтеза с порфиринами, включая предшественника уропорфириногена III . В отличие от гема, который содержит железо, связанное с центром N4, большинство хлорофиллов связывают магний . Осевые лиганды, присоединенные к Mg 2+ центр часто опускается для ясности. К хлориновому кольцу присоединены различные боковые цепи, обычно включающие длинную фитильную цепь ( С 20 Н 39 О ). Наиболее широко распространенной формой у наземных растений является хлорофилл а . Единственная разница между хлорофиллом а и хлорофиллом b заключается в том, что первый имеет метильную группу, а второй — формильную группу. Эта разница вызывает значительную разницу в спектре поглощения, позволяя растениям поглощать большую часть видимого света.
Структуры хлорофиллов кратко описаны ниже: [17] [18]
| Хлорофилл а | Хлорофилл б | Хлорофилл с 1 | Хлорофилл с 2 | хлорофилл д | Хлорофилл ф [14] | |
|---|---|---|---|---|---|---|
| Молекулярная формула | С 55 Н 72 О 5 Н 4 Мг | С 55 Н 70 О 6 Н 4 Мг | С 35 Н 30 О 5 Н 4 Мг | С 35 Н 28 О 5 Н 4 Мг | С 54 Н 70 О 6 Н 4 Мг | С 55 Н 70 О 6 Н 4 Мг |
| группа С2 | −CH−CH3 | −CH−CH3 | −CH−CH3 | −CH−CH3 | −CH−CH3 | −ЗА |
| группа С3 | −СН=СН 2 | −СН=СН 2 | −СН=СН 2 | −СН=СН 2 | −ЗА | −СН=СН 2 |
| группа С7 | −CH−CH3 | −ЗА | −CH−CH3 | −CH−CH3 | −CH−CH3 | −CH−CH3 |
| группа С8 | −СН 2 СН 3 | −СН 2 СН 3 | −СН 2 СН 3 | −СН=СН 2 | −СН 2 СН 3 | −СН 2 СН 3 |
| группа С17 | −CH 2 CH 2 COO-Фитил | −CH 2 CH 2 COO-Фитил | −CH=CHCOOH | −CH=CHCOOH | −CH 2 CH 2 COO-Фитил | −CH 2 CH 2 COO-Фитил |
| Связь C17-C18 | Одинокий (хлор) | Одинокий (хлор) | Двойной (порфирин) | Двойной (порфирин) | Одинокий (хлор) | Одинокий (хлор) |
| возникновение | Универсальный | В основном растения | Различные водоросли | Различные водоросли | Цианобактерии | Цианобактерии |
- Структуры хлорофиллов
 хлорофилл а
хлорофилл а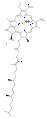 хлорофилл б
хлорофилл б хлорофилл с 1
хлорофилл с 1 хлорофилл с 2
хлорофилл с 2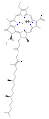 хлорофилл д
хлорофилл д хлорофилл f
хлорофилл f


Хлорофилл е зарезервирован для пигмента, который был извлечен из водорослей в 1966 году, но не описан химически. Помимо буквенных хлорофиллов, в дикой природе известно множество модификаций боковой цепи хлорофилла. Например, Prochromococcus цианобактерия использует 8-винил Chl a и b . [19]
Измерение содержания хлорофилла [ править ]

Хлорофиллы можно экстрагировать из белка органическими растворителями. [20] [21] [22] Таким образом можно оценить концентрацию хлорофилла в листе. [23] Существуют также методы разделения хлорофилла а и хлорофилла b .
В диэтиловом эфире хлорофилл а имеет приблизительные максимумы поглощения 430 и 662 нм, тогда как хлорофилл b имеет приблизительные максимумы 453 и 642 нм. [24] Пики поглощения хлорофилла а находятся при 465 нм и 665 нм. Хлорофилл а флуоресцирует при 673 нм (максимум) и 726 нм. Пиковый молярный коэффициент поглощения хлорофилла а превышает 10 5 М −1 см −1 , что является одним из самых высоких показателей для низкомолекулярных органических соединений. [25] В 90% ацетон-воде пиковые длины волн поглощения хлорофилла а составляют 430 нм и 664 нм; пики хлорофилла b – 460 нм и 647 нм; пики хлорофилла с 1 составляют 442 нм и 630 нм; пики хлорофилла с 2 составляют 444 нм и 630 нм; пики хлорофилла d составляют 401 нм, 455 нм и 696 нм. [26]
Отношение эмиссии флуоресценции можно использовать для измерения содержания хлорофилла. За счет возбуждения флуоресценции хлорофилла на более низкой длине волны соотношение эмиссии флуоресценции хлорофилла при 705 ± 10 нм и 735 ± 10 нм может обеспечить линейную зависимость содержания хлорофилла по сравнению с химическими испытаниями. Соотношение F 735 / F 700 обеспечивало значение корреляции r 2 0,96 по сравнению с химическим тестированием в диапазоне от 41 мг·м. −2 до 675 мг м −2 . Гительсон также разработал формулу для прямого определения содержания хлорофилла в мг·м. −2 . Формула обеспечила надежный метод измерения содержания хлорофилла от 41 мг м. −2 до 675 мг м −2 с корреляцией r 2 значение 0,95. [27]
Биосинтез [ править ]
В некоторых растениях хлорофилл образуется из глутамата и синтезируется по разветвленному пути биосинтеза , который является общим с гемом и сирогемой . [28] [29] [30] Хлорофиллсинтаза [31] Фермент, осуществляющий биосинтез хлорофилла а : [32] [33]
- хлорофиллид а + фитилдифосфат хлорофилл а + дифосфат

В результате этого преобразования образуется сложный эфир группы карбоновой кислоты в хлорофиллиде а с 20-углеродным дитерпеновым спиртом фитолом . Хлорофилл b производится тем же ферментом, который действует на хлорофиллид b . То же самое известно о хлорофиллах d и f , которые оба состоят из соответствующих хлорофиллидов, в конечном итоге полученных из хлорофиллида а . [34]
У покрытосеменных растений более поздние этапы пути биосинтеза зависят от света. Такие растения бледны ( этиолированы ), если выращиваются в темноте. Несосудистые растения и зеленые водоросли имеют дополнительный светонезависимый фермент и растут зелеными даже в темноте. [35]
Хлорофилл связан с белками . Протохлорофиллид , один из промежуточных продуктов биосинтеза, встречается преимущественно в свободной форме и в условиях освещения действует как фотосенсибилизатор , образуя свободные радикалы , которые могут быть токсичными для растения. Следовательно, растения регулируют количество этого предшественника хлорофилла. У покрытосеменных эта регуляция достигается на стадии аминолевулиновой кислоты (АЛК) – одного из промежуточных соединений пути биосинтеза. Растения, получающие АЛК, накапливают высокие и токсичные уровни протохлорофиллида; то же самое делают и мутанты с поврежденной регуляторной системой. [36]
цикл Старение и хлорофилла
Процесс старения растений включает в себя деградацию хлорофилла: например, фермент хлорофиллаза ( EC 3.1.1.14 ) гидролизует фитильную боковую цепь, обращая вспять реакцию, в которой хлорофиллы биосинтезируются из хлорофиллида a или b . Поскольку хлорофиллид а может быть преобразован в хлорофиллид b , а последний может быть повторно этерифицирован в хлорофилл b , эти процессы позволяют совершать циклический цикл между хлорофиллами a и b . Более того, хлорофилл b может быть восстановлен напрямую (через 7 1 -гидроксихлорофилл а ) обратно в хлорофилл а , завершая цикл. [37] [38] На более поздних стадиях старения хлорофиллиды превращаются в группу бесцветных тетрапирролов, известных как нефлуоресцентные катаболиты хлорофилла (NCC), с общей структурой:
Эти соединения также были обнаружены в созревающих плодах и придают характерную осеннюю окраску растениям лиственным . [38] [39]
Распространение [ править ]
Карты хлорофилла показывают миллиграммы хлорофилла на кубический метр морской воды каждый месяц. Места, где количество хлорофилла было очень низким, что указывает на очень низкую численность фитопланктона , отмечены синим цветом. Места, где концентрация хлорофилла была высокой, а это означает, что росло много фитопланктона, отмечены желтым цветом. Наблюдения происходят с помощью спектрорадиометра визуализации среднего разрешения (MODIS) на спутнике НАСА Aqua. Земля окрашена в темно-серый цвет, а места, где MODIS не смог собрать данные из-за морского льда, полярной тьмы или облаков, — светло-серые. Самые высокие концентрации хлорофилла, где процветают крошечные обитающие на поверхности океанские растения , наблюдаются в холодных полярных водах или в местах, где океанские течения выносят холодную воду на поверхность, например, вокруг экватора и вдоль берегов континентов. Не холодная вода сама по себе стимулирует фитопланктон. Вместо этого низкие температуры часто являются признаком того, что вода поднимается на поверхность из более глубоких слоев океана, неся с собой питательные вещества, накопившиеся с течением времени. В полярных водах питательные вещества накапливаются в поверхностных водах в темные зимние месяцы, когда растения не могут расти. Когда весной и летом солнечный свет возвращается, растения процветают в высоких концентрациях. [40]
использование Кулинарное
Синтетический хлорофилл зарегистрирован в качестве красителя пищевой добавки, его номер E — E140 . Повара используют хлорофилл для придания зеленого цвета различным продуктам питания и напиткам, таким как макароны и спиртные напитки. Абсент приобретает свой зеленый цвет естественным образом благодаря хлорофиллу, вносимому в него из большого количества трав, используемых при его производстве. [41] Хлорофилл не растворяется в воде, и его сначала смешивают с небольшим количеством растительного масла, чтобы получить желаемый раствор . [ нужна ссылка ]
использование Биологическое
Исследование 2002 года показало, что «листья, подвергнутые сильному свету, содержат деградированные основные антенные белки , в отличие от тех, которые хранятся в темноте, что согласуется с исследованиями освещения изолированных белков ». Это показалось авторам подтверждением гипотезы о том, что « активные формы кислорода играют роль in vivo » в кратковременном поведении растений. [42]
См. также [ править ]
- Бактериохлорофилл , родственные соединения фототрофных бактерий.
- Хлорофиллин — полусинтетическое производное хлорофилла.
- Глубокий хлорофилловый максимум
- Флуоресценция хлорофилла для измерения стресса растений.
- Гипотеза Пурпурной Земли — научная гипотеза , объясняющая эволюцию красно-синего спектрального сродства хлорофилла.
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б Виртанен О, Константиниду Э, Тюйстъярви Э (2020). «Хлорофилл не отражает зеленый свет – как исправить заблуждение» . Журнал биологического образования . 56 (5): 1–8. дои : 10.1080/00219266.2020.1858930 .
- ^ Мэй П. «Хлорофилл» . Бристольский университет .
- ^ «хлорофилл» . Интернет-словарь этимологии .
- ^ Мунир С., Ким Э.Дж., Пак Дж.С., Ли Дж.Х. (март 2014 г.). «Влияние зеленого, красного и синего светодиодов на белки мультибелкового комплекса и фотосинтетическую активность при различной интенсивности освещения в листьях салата (Lactuca sativa L.)» . Международный журнал молекулярных наук . 15 (3): 4657–70. дои : 10.3390/ijms15034657 . ПМЦ 3975419 . ПМИД 24642884 .
- ^ Шпеер Б.Р. (1997). «Фотосинтетические пигменты» . Глоссарий UCMP (онлайн) . Музей палеонтологии Калифорнийского университета . Проверено 17 июля 2010 г.
- ^ См.:
- Делепин М. [на французском языке] (сентябрь 1951 г.). «Жозеф Пеллетье и Жозеф Кавенту». Журнал химического образования . 28 (9): 454. Бибкод : 1951ЖЧЭд..28..454Д . дои : 10.1021/ed028p454 .
- Пеллетье П.Ж., Кавенту Ж.Б. (1817). «Объявление о зелени в листьях» . Аптечный журнал (на французском языке). 3 : 486–491. Мы п. 490, авторы предлагают новое название хлорофилла. Из стр. 490: «Мы не имеем права называть вещество, давно известное и к истории которого мы добавили лишь несколько фактов; однако мы предложим, не придавая этому никакого значения, название хлорофилл , хлороса , цвет и φύλλον — лист: это название указывает на роль, которую он играет в природе». (Мы не имеем права называть вещество, [которое] давно известно и к рассказу которого мы добавили лишь несколько фактов; однако мы предложим, не придавая этому никакого значения, название хлорофилл , от хлороса , цвет, и φύλλον , лист: это имя указывает на роль, которую он играет в природе.)
- ^ Вильштеттер Р. (1906). « Вклад в знание состава хлорофилла». Анналы химии (на немецком языке). 350 (1–2): 48–82. дои : 10.1002/jlac.19063500103 .
Из стр. 49: «Основным продуктом щелочного гидролиза являются темно-зеленые щелочные соли. Они содержат сложные соединения магния, содержащие металл в связи, которая удивительно устойчива к щелочам даже при высоких температурах». (Темно-зеленые щелочные соли образуют основной продукт щелочного гидролиза. В них присутствуют сложные соединения магния, содержащие металл в связи, необычайно устойчивой к щелочам даже при высокой температуре.)
- ^ Jump up to: Перейти обратно: а б Мотильва М.Ю. (2008). «Хлорофиллы - от функциональности в продуктах питания до значения для здоровья». 5-й конгресс «Пигменты в пищевых продуктах» – за качество и здоровье (Печать). Университет Хельсинки. ISBN 978-952-10-4846-3 .
- ^ Вудворд Р.Б., Айер В.А., Битон Дж.М., Бикельхаупт Ф., Боннетт Р., Бухшахер П. и др. (июль 1960 г.). «Полный синтез хлорофилла» (PDF) . Журнал Американского химического общества . 82 (14): 3800–3802. дои : 10.1021/ja01499a093 . Архивировано (PDF) из оригинала 10 апреля 2011 г.
- ^ Флеминг I (14 октября 1967 г.). «Абсолютная конфигурация и структура хлорофилла». Природа . 216 (5111): 151–152. Бибкод : 1967Natur.216..151F . дои : 10.1038/216151a0 . S2CID 4262313 .
- ^ Вудворд Р.Б., Айер В.А., Битон Дж.М., Бикельхаупт Ф., Боннетт Р., Бухшахер П. и др. (1990). «Полный синтез хлорофилла А ». Тетраэдр . 46 (22): 7599–7659. дои : 10.1016/0040-4020(90)80003-Z .
- ^ Джабр Ф (август 2010 г.). «Новая форма хлорофилла?» . Научный американец .
- ^ «Инфракрасный хлорофилл может усилить солнечные батареи» . Новый учёный . 19 августа 2010 года . Проверено 15 апреля 2012 г.
- ^ Jump up to: Перейти обратно: а б Чен М., Шлип М., Уиллоуз Р.Д., Цай З.Л., Нейлан Б.А., Шеер Х. (сентябрь 2010 г.). «Хлорофилл со смещением в красную сторону». Наука . 329 (5997): 1318–9. Бибкод : 2010Sci...329.1318C . дои : 10.1126/science.1191127 . ПМИД 20724585 . S2CID 206527174 .
- ^ Картер Дж.С. (1996). «Фотосинтез» . Университет Цинциннати . Архивировано из оригинала 29 июня 2013 г.
- ^ «Раздел 1.3. Фотосинтетические клетки» . Основы клеточной биологии . Природа. 5 июля 2013 г.
{{cite book}}:|website=игнорируется ( помогите ) - ^ Шеер Х (2006). «Обзор хлорофиллов и бактериохлорофиллов: биохимия, биофизика, функции и применение». Хлорофиллы и бактериохлорофиллы . Достижения в области фотосинтеза и дыхания. Том. 25. стр. 1–26. дои : 10.1007/1-4020-4516-6_1 . ISBN 978-1-4020-4515-8 .
- ^ Танигучи М., Линдси Дж.С. (январь 2017 г.). «Синтетические хлорины, возможные заменители хлорофиллов, полученные путем дериватизации порфиринов». Химические обзоры . 117 (2): 344–535. doi : 10.1021/acs.chemrev.5b00696 . ОСТИ 1534468 . ПМИД 27498781 .
- ^ Чен М (2019). «Хлорофиллы d и f: синтез, возникновение, сбор света и организация пигментов в хлорофиллсвязывающих белковых комплексах». Достижения в ботанических исследованиях . 90 : 121–139. дои : 10.1016/bs.abr.2019.03.006 . ISBN 9780081027523 . S2CID 149632511 .
- ^ Маркер А.Ф. (1972). «Использование ацетона и метанола для оценки хлорофилла в присутствии феофитина в растении». Пресноводная биология . 2 (4): 361–385. дои : 10.1111/j.1365-2427.1972.tb00377.x .
- ^ Джеффри С.В., Сибата (февраль 1969 г.). «Некоторые спектральные характеристики хлорофилла c Tridacna crocea Zooxanthellae» . Биологический бюллетень . 136 (1): 54–62. дои : 10.2307/1539668 . JSTOR 1539668 .
- ^ Гилпин Л. (21 марта 2001 г.). «Методы анализа донных фотосинтетических пигментов» . Школа естественных наук Университета Нейпира . Архивировано из оригинала 14 апреля 2008 года . Проверено 17 июля 2010 г.
- ^ Кейт ТМ, Перкинс ТД (октябрь 2003 г.). «Мониторинг содержания хлорофилла в клене сахарном (Acer saccharum)» . Физиология дерева . 23 (15): 1077–9. дои : 10.1093/treephys/23.15.1077 . ПМИД 12975132 .
- ^ Гросс Дж (1991). Пигменты овощей: хлорофиллы и каротиноиды . Ван Ностранд Рейнхольд. ISBN 978-0442006570 .
- ^ Порра Р.Дж., Томпсон В.А., Кридеманн П.Е. (1989). «Определение точных коэффициентов экстинкции и одновременных уравнений для анализа хлорофиллов a и b, экстрагированных четырьмя различными растворителями: проверка концентрации стандартов хлорофилла методом атомно-абсорбционной спектроскопии». Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 975 (3): 384–394. дои : 10.1016/S0005-2728(89)80347-0 .
- ^ Ларкум А.В., Дуглас С., Рэйвен Дж.А., ред. (2003). Фотосинтез у водорослей . Лондон: Клювер. ISBN 978-0-7923-6333-0 .
- ^ Гительсон А.А., Бушманн С., Лихтенталер Х.К. (1999). «Коэффициент флуоресценции хлорофилла F 735 / F 700 как точная мера содержания хлорофилла в растениях». Дистанционный датчик окружающей среды . 69 (3): 296–302. Бибкод : 1999RSEnv..69..296G . дои : 10.1016/S0034-4257(99)00023-1 .
- ^ Баттерсби, Арканзас (декабрь 2000 г.). «Тетрапирролы: пигменты жизни». Отчеты о натуральных продуктах . 17 (6): 507–26. дои : 10.1039/B002635M . ПМИД 11152419 .
- ^ Ахтар М (2007). «Модификация боковых цепей ацетата и пропионата во время биосинтеза гема и хлорофиллов: механистические и стереохимические исследования». Симпозиум 180 Фонда Ciba - Биосинтез тетрапиррольных пигментов . Симпозиумы Фонда Новартис. Том. 180. стр. 131–155. дои : 10.1002/9780470514535.ch8 . ISBN 9780470514535 . ПМИД 7842850 .
- ^ Уиллоуз Р.Д. (июнь 2003 г.). «Биосинтез хлорофиллов из протопорфирина IX». Отчеты о натуральных продуктах . 20 (3): 327–41. дои : 10.1039/B110549N . ПМИД 12828371 .
- ^ Шмид Х.К., Рассадина В., Остер У., Шох С., Рюдигер В. (ноябрь 2002 г.). «Предварительная загрузка хлорофиллсинтазы тетрапренилдифосфатом является обязательным этапом биосинтеза хлорофилла» (PDF) . Биологическая химия . 383 (11): 1769–78. дои : 10.1515/BC.2002.198 . ПМИД 12530542 . S2CID 3099209 .
- ^ Экхардт У, Гримм Б, Хёртенштейнер С (сентябрь 2004 г.). «Последние достижения в области биосинтеза хлорофилла и его распада у высших растений» . Молекулярная биология растений . 56 (1): 1–14. дои : 10.1007/s11103-004-2331-3 . ПМИД 15604725 . S2CID 21174896 .
- ^ Болливар Д.В. (ноябрь 2006 г.). «Последние достижения в биосинтезе хлорофилла». Исследования фотосинтеза . 90 (2): 173–94. дои : 10.1007/s11120-006-9076-6 . ПМИД 17370354 . S2CID 23808539 .
- ^ Цузуки Ю, Цукатани Ю, Ямакава Х, Ито С, Фудзита Ю, Ямамото Х (март 2022 г.). «Влияние света и кислорода на биосинтез хлорофилла d у морской цианобактерии Acaryochromis marina » . Растения . 11 (7): 915. doi : 10.3390/plants11070915 . ПМК 9003380 . ПМИД 35406896 .
- ^ Мураки Н., Номата Дж., Эбата К., Мидзогучи Т., Шиба Т., Тамиаки Х., Курису Г., Фудзита Ю. (май 2010 г.). «Рентгеновская кристаллическая структура светонезависимой протохлорофиллидредуктазы». Природа 465 (7294): 110–4. Бибкод : 2010Природа.465..110М . дои : 10.1038/nature08950 . ПМИД 20400946 . S2CID 4427639 .
- ^ Мескаускене Р., Натер М., Гослингс Д., Кесслер Ф., оп ден Кэмп Р., Апель К. (октябрь 2001 г.). «ГРИПП: негативный регулятор биосинтеза хлорофилла у Arabidopsis thaliana» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (22): 12826–31. Бибкод : 2001PNAS...9812826M . дои : 10.1073/pnas.221252798 . JSTOR 3056990 . ПМК 60138 . ПМИД 11606728 .
- ^ «Хлорофилловый цикл» . ИУБМБ. 2011 . Проверено 4 июня 2020 г.
- ^ Jump up to: Перейти обратно: а б Хёртенштейнер С (2006). «Деградация хлорофилла во время старения». Ежегодный обзор биологии растений . 57 : 55–77. doi : 10.1146/annurev.arplant.57.032905.105212 . ПМИД 16669755 .
- ^ Мюллер Т., Ульрих М., Онгания К.Х., Кройтлер Б. (2007). «Бесцветные тетрапиррольные катаболиты хлорофилла, обнаруженные в созревающих фруктах, являются эффективными антиоксидантами» . Ангеванде Хеми . 46 (45): 8699–702. дои : 10.1002/anie.200703587 . ПМК 2912502 . ПМИД 17943948 .
- ^ «Хлорофилл: Карты мира» . Earthobservatory.nasa.gov . Проверено 2 февраля 2014 г.
- ^ Адамс Дж (2004). Отвратительный абсент: история дьявола в бутылке . Великобритания: IBTauris, 2004. с. 22. ISBN 978-1860649202 .
- ^ Золла Л., Ринальдуччи С. (декабрь 2002 г.). «Участие активных форм кислорода в деградации светособирающих белков при световом стрессе». Биохимия . 41 (48): 14391–402. дои : 10.1021/bi0265776 . ПМИД 12450406 .