Цветок

Цветок или , также известный как цветение цветковых цветение , представляет собой репродуктивную структуру растений (растений отдела покрытосеменных ). Цветы состоят из комбинации вегетативных органов — чашелистиков , которые окружают и защищают развивающийся цветок, лепестков, привлекающих опылителей, и репродуктивных органов, вырабатывающих гаметофиты , которые у цветковых растений производят гаметы . Мужские гаметофиты, вырабатывающие сперму, заключены в пыльцевые зерна, образующиеся в пыльниках . Женские гаметофиты содержатся в семязачатках, образующихся в плодолистиках .
Большинство цветковых растений зависят от животных, таких как пчелы , мотыльки и бабочки , которые переносят пыльцу между разными цветами, и эволюционировали, чтобы привлечь этих опылителей с помощью различных стратегий, включая ярко окрашенные, заметные лепестки, привлекательные ароматы и производство нектара. , источник пищи для опылителей. [1] Таким образом, многие цветковые растения эволюционировали совместно с опылителями и стали взаимозависимыми от услуг, которые они оказывают друг другу — в случае растения — от средств размножения; в случае опылителя - источник пищи. [2]
Когда пыльца пыльника рыльце цветка откладывается на пестика , это называется опылением . Некоторые цветы могут самоопыляться , образуя семена с использованием пыльцы другого цветка одного и того же растения, но другие имеют механизмы предотвращения самоопыления и полагаются на перекрестное опыление , когда пыльца переносится с пыльника одного цветка на рыльце пестика другого цветка. другой цветок на другой особи того же вида . Самоопыление происходит в цветах, где тычинки и плодолистики созревают одновременно и расположены так, что пыльца может попасть на рыльце цветка. Это опыление не требует от растения вложений в обеспечение нектара и пыльцы в качестве пищи для опылителей. [3] Некоторые цветки образуют диаспоры без оплодотворения ( партенокарпия ). После оплодотворения из завязи цветка развивается плод , содержащий семена .
Цветы издавна ценились людьми за их красоту и приятный аромат, а также имеют культурное значение как религиозные, ритуальные или символические объекты, а также источники лекарств и продуктов питания .
Этимология
Цветок происходит от среднеанглийского слова « мука» , которое относилось как к молотому зерну , так и к репродуктивной структуре растений, прежде чем оно отделилось в 17 веке. Первоначально оно происходит от латинского имени итальянской богини цветов Флоры . Первое слово, обозначающее цветок в английском языке, было « цветок» . [4] хотя сейчас это относится только к цветам фруктовых деревьев . [5]
Морфология

Морфология , цветка, или его форма и строение [6] можно рассматривать как две части: вегетативную часть, состоящую из нерепродуктивных структур, таких как лепестки ; и репродуктивная или сексуальная часть. Стереотипный цветок состоит из четырех видов структур, прикрепленных к кончику короткого стебля или оси, называемых цветоложем . Каждая из этих частей или цветочных органов расположена по спирали, называемой мутовкой . [7] Четыре основных оборота (начинающиеся от основания цветка или самого нижнего узла и идущие вверх) — это чашечка , венчик , андроцей и гинецей . Чашечка и венчик вместе составляют нерепродуктивную часть цветка, называемую околоцветником , и в некоторых случаях могут не дифференцироваться. Если это так, то их называют листочками околоцветника . [8]
Околоцветник
Чашечка
Чашелистики листья , называемые чашечкой, представляют собой видоизмененные , расположенные на внешнем мутовке цветка. Они похожи на листья, имеют широкое основание, устьица и хлорофилл. [9] и может иметь прилистники . Чашелистики часто восковые и жесткие, быстро растут, защищая цветок по мере его развития. [9] [10] Они могут быть листопадными , но чаще всего растут, помогая распространению плодов. Если чашечка срослась, ее называют гамосепалой. [9]
Королла
Лепестки . вместе с венчиком представляют собой листовидные структуры, почти или полностью лишенные волокон, образующие самый внутренний оборот околоцветника Они часто нежные и тонкие, обычно имеют цвет, форму или аромат, способствующие опылению. [11] Хотя по форме они похожи на листья, они больше похожи на тычинки в том, что образуются почти одновременно друг с другом, но их последующий рост задерживается. Если венчик сросшийся, его называют симпетальным. [12]
репродуктивный

Андроцей
Андроцей . , или тычинки, представляет собой мутовку мужских частей, образующих пыльцу Тычинки обычно состоят из пыльника, состоящего из четырех пыльцевых мешочков, расположенных в две теки , соединенных с нитью или стеблем. Пыльник содержит микроспороциты, которые становятся пыльцой , мужским гаметофитом после мейоза . Хотя они демонстрируют самое широкое разнообразие среди цветочных органов, андроцей обычно ограничивается одним оборотом и только в редких случаях двумя оборотами. Тычинки различаются по количеству, размеру, форме, ориентации и месту соединения с цветком. [11] [12]
Вообще существует только один тип тычинок, но есть виды растений, у которых цветки имеют два типа; «нормальный» и один с пыльниками, производящими стерильную пыльцу, предназначенную для привлечения опылителей. [13]
Гинецей
Гинецей . , или плодолистики, — это женская часть цветка, расположенная на самом внутреннем мутовке Каждый плодолистик состоит из рыльца , на которое попадает пыльца, столбика, выполняющего роль стебля, и завязи , в которой находятся семязачатки. Плодолистики могут состоять из одного или нескольких оборотов, а при слиянии их часто называют пестиком. Внутри яичника яйцеклетки прикрепляются к плаценте с помощью структур, называемых канатиками . [14] [15]
Вариация
Хотя такое расположение считается «типичным», виды растений демонстрируют большие различия в цветочной структуре. [16] Четыре основные части цветка обычно определяются их положением на цветоложе, а не их функцией. У многих цветов отсутствуют некоторые части, или части могут быть модифицированы для выполнения других функций или выглядеть как обычно другая часть. [17] У некоторых семейств, например у злаковых , лепестки сильно редуцированы; у многих видов чашелистики красочные и похожи на лепестки. У других цветов есть видоизмененные тычинки, похожие на лепестки; махровые цветки пионов и роз представляют собой преимущественно лепестковидные тычинки. [18]
Многие цветы обладают симметрией . Когда околоцветник делится пополам по центральной оси в любой точке и образуются симметричные половинки, цветок называется актиноморфным или правильным. Это пример радиальной симметрии . Когда цветы разделены пополам и образуют только одну линию, образующую симметричные половинки, цветок называется неправильным или зигоморфным . Если в редких случаях они вообще не имеют симметрии, их называют асимметричными. [19] [20]
Цветы могут прикрепляться непосредственно к растению у основания ( сидячие — опорный стебель или стебель сильно редуцирован или отсутствует). [21] Стебель или стебель, на котором расположен цветок, или соцветие цветов, называется цветоносом . Если цветонос поддерживает более одного цветка, стебли, соединяющие каждый цветок с главной осью, называются цветоножками . [22] Вершина цветущего стебля образует терминальное вздутие, которое называется тором или цветоложем. [20]
У большинства видов отдельные цветки имеют как пестики , так и тычинки. Ботаники описывают эти цветы как совершенные, двуполые или гермафродитные . У некоторых видов растений цветки несовершенные или однополые: имеющие либо мужскую (тычинки), либо женскую (пестик) части. Если на одном растении появляются однополые мужские и женские цветки, вид называют однодомным . [23] Однако если отдельное растение женского или мужского пола, вид называется двудомным . У многих цветов есть нектарники — железы, вырабатывающие сладкую жидкость, используемую для привлечения опылителей. Они не рассматриваются как отдельный орган. [24]
Соцветие

У тех видов, которые имеют на оси более одного цветка, собирательная гроздь цветков называется соцветием . Некоторые соцветия состоят из множества мелких цветков, расположенных в форме, напоминающей один цветок. Типичным примером этого является большинство представителей очень большой сложной группы ( Asteraceae ). одиночная маргаритка или подсолнух Например, — это не цветок, а цветочная головка — соцветие, состоящее из множества цветков (или соцветий). [26] Соцветие может включать в себя специализированные стебли и видоизмененные листья, известные как прицветники . [27]
Цветочные схемы и формулы
Цветочная формула — это способ представить структуру цветка с помощью определенных букв, цифр и символов, представляя существенную информацию о цветке в компактной форме. Он может представлять собой таксон , обычно указывая диапазоны количества различных органов или отдельных видов. Цветочные формулы были разработаны в начале 19 века, и с тех пор их использование сократилось. Преннер и др. (2010) разработали расширение существующей модели, чтобы расширить описательные возможности формулы. [28] Формат цветочных формул различается в разных частях мира, но они передают одну и ту же информацию. [29] [30] [31] [32]
Строение цветка можно выразить и с помощью цветочных схем . Использование схематических диаграмм может заменить длинные описания или сложные рисунки в качестве инструмента для понимания как структуры, так и эволюции цветка. На таких диаграммах могут быть показаны важные особенности цветков, в том числе взаимное расположение различных органов, в том числе наличие слияния и симметрии, а также детали строения. [33]
Разработка
Цветок развивается на модифицированном побеге или оси из детерминированной апикальной меристемы ( детерминантность означает, что ось вырастает до заданного размера). У него сжатые междоузлия, несущие структуры, которые в классической морфологии растений интерпретируются как сильно модифицированные листья . [34] Однако детальные исследования развития показали, что тычинки часто зарождаются более или менее как модифицированные стебли (кауломы), которые в некоторых случаях могут даже напоминать веточки . [35] [16] Принимая во внимание все разнообразие развития андроцея цветковых растений, мы обнаруживаем континуум между видоизмененными листьями (филломами), видоизмененными стеблями (кауломами) и видоизмененными веточками (побегами). [36] [37]
Переход
Переход к цветению — одно из основных фазовых изменений, которые растение совершает в течение своего жизненного цикла. Переход должен происходить в то время, которое благоприятно для оплодотворения и образования семян , обеспечивая тем самым максимальный репродуктивный успех. Чтобы удовлетворить эти потребности, растение способно интерпретировать важные эндогенные сигналы и сигналы окружающей среды, такие как изменения уровня растительных гормонов , а также сезонные изменения температуры и фотопериода . [38] Многие многолетние и большинство двулетних растений требуют яровизации для цветения . Молекулярная интерпретация этих сигналов осуществляется посредством передачи сложного сигнала, известного как флориген , который включает в себя множество генов , включая Constans, локус цветения C и локус цветения T. Флориген вырабатывается в листьях в репродуктивно благоприятных условиях и действует в почки и кончики роста вызывают ряд различных физиологических и морфологических изменений. [39]

Первым этапом перехода является трансформация вегетативных стеблевых зачатков в цветочные зачатки. Это происходит, когда происходят биохимические изменения, приводящие к изменению клеточной дифференциации тканей листьев, почек и стебля в ткани, которые впоследствии перерастают в репродуктивные органы. Рост центральной части кончика стебля прекращается или уплощается, а по бокам вокруг внешней части конца стебля появляются мутовчатые или спиральные выступы. Эти выступы развиваются в чашелистики, лепестки, тычинки и плодолистики . Как только этот процесс начинается, у большинства растений его невозможно повернуть вспять, и на стеблях появляются цветы, даже если первоначальное начало формирования цветка зависело от какого-либо фактора окружающей среды. [40]
Развитие органов
Модель ABC — это простая модель, описывающая гены, ответственные за развитие цветов. Три генные активности взаимодействуют комбинаторным образом, определяя идентичность развития зачаткового цветка органа внутри апикальной меристемы . Эти функции генов называются A, B и C. Гены A экспрессируются только во внешней и самой нижней части апикальной меристемы, которая становится мутовкой чашелистиков. Во втором обороте экспрессируются гены A и B, что приводит к образованию лепестков. В третьем обороте гены B и C взаимодействуют, образуя тычинки, а в центре цветка только гены C дают начало плодолистикам. Модель основана на исследованиях аберрантных цветков и мутаций Arabidopsis thaliana и львиного зева Antirhinum majus . Например, при утрате функции гена B мутантные цветы производятся с чашелистиками в первом мутовке, как обычно, но также и во втором мутовке вместо нормального формирования лепестков. В третьем обороте отсутствие функции B, но наличие функции C имитирует четвертый оборот, что приводит к образованию плодолистиков и в третьем обороте. [41]
Function
The principal purpose of a flower is the reproduction[42] of the individual and the species. All flowering plants are heterosporous, that is, every individual plant produces two types of spores. Microspores are produced by meiosis inside anthers and megaspores are produced inside ovules that are within an ovary. Anthers typically consist of four microsporangia and an ovule is an integumented megasporangium. Both types of spores develop into gametophytes inside sporangia. As with all heterosporous plants, the gametophytes also develop inside the spores, i. e., they are endosporic.
Pollination

Since the flowers are the reproductive organs of the plant, they mediate the joining of the sperm, contained within pollen, to the ovules — contained in the ovary.[10] Pollination is the movement of pollen from the anthers to the stigma.[43] Normally pollen is moved from one plant to another, known as cross-pollination, but many plants are able to self-pollinate. Cross-pollination is preferred because it allows for genetic variation, which contributes to the survival of the species.[44] Many flowers depend on external factors for pollination, such as: the wind, water, animals, and especially insects. Larger animals such as birds, bats, and even some pygmy possums,[45] however, can also be employed.[46][47] To accomplish this, flowers have specific designs which encourage the transfer of pollen from one plant to another of the same species. The period of time during which this process can take place (when the flower is fully expanded and functional) is called anthesis,[48] hence the study of pollination biology is called anthecology.[49]
Flowering plants usually face evolutionary pressure to optimize the transfer of their pollen, and this is typically reflected in the morphology of the flowers and the behavior of the plants.[50] Pollen may be transferred between plants via a number of 'vectors,' or methods. Around 80% of flowering plants make use of biotic, or living vectors. Others use abiotic, or non-living, vectors and some plants make use of multiple vectors, but most are highly specialised.[51]
Though some fit between or outside of these groups,[52] most flowers can be divided between the following two broad groups of pollination methods:
Biotic pollination
Flowers that use biotic vectors attract and use insects, bats, birds, or other animals to transfer pollen from one flower to the next. Often they are specialized in shape and have an arrangement of the stamens that ensures that pollen grains are transferred to the bodies of the pollinator when it lands in search of its attractant (such as nectar, pollen, or a mate).[53] In pursuing this attractant from many flowers of the same species, the pollinator transfers pollen to the stigmas—arranged with equally pointed precision—of all of the flowers it visits.[54] Many flowers rely on simple proximity between flower parts to ensure pollination, while others have elaborate designs to ensure pollination and prevent self-pollination.[44] Flowers use animals including: insects (entomophily), birds (ornithophily), bats (chiropterophily), lizards,[47] and even snails and slugs (malacophilae).[55]
Attraction methods

Plants cannot move from one location to another, thus many flowers have evolved to attract animals to transfer pollen between individuals in dispersed populations. Most commonly, flowers are insect-pollinated, known as entomophilous; literally "insect-loving" in Greek.[57] To attract these insects flowers commonly have glands called nectaries on various parts that attract animals looking for nutritious nectar.[58] Some flowers have glands called elaiophores, which produce oils rather than nectar.[59] Birds and bees have color vision, enabling them to seek out colorful flowers.[60] Some flowers have patterns, called nectar guides, that show pollinators where to look for nectar; they may be visible only under ultraviolet light, which is visible to bees and some other insects.[61]
Flowers also attract pollinators by scent, though not all flower scents are appealing to humans; a number of flowers are pollinated by insects that are attracted to rotten flesh and have flowers that smell like dead animals. These are often called Carrion flowers, including plants in the genus Rafflesia, and the titan arum.[60] Flowers pollinated by night visitors, including bats and moths, are likely to concentrate on scent to attract pollinators and so most such flowers are white.[62] Some plants pollinated by bats have a sonar-reflecting petal above its flowers, which helps the bat find them,[63] and one species, the cactus Espostoa frutescens, has flowers that are surrounded by an area of sound-absorbent and wooly hairs called the cephalium, which absorbs the bat's ultrasound instead.[64]
Flowers are also specialized in shape and have an arrangement of the stamens that ensures that pollen grains are transferred to the bodies of the pollinator when it lands in search of its attractant. Other flowers use mimicry or pseudocopulation to attract pollinators. Many orchids for example, produce flowers resembling female bees or wasps in colour, shape, and scent. Males move from one flower to the next in search of a mate, pollinating the flowers.[65][66]
Pollinator relationships
Many flowers have close relationships with one or a few specific pollinating organisms. Many flowers, for example, attract only one specific species of insect, and therefore rely on that insect for successful reproduction. This close relationship an example of coevolution, as the flower and pollinator have developed together over a long period of time to match each other's needs.[67] This close relationship compounds the negative effects of extinction, however, since the extinction of either member in such a relationship would almost certainly mean the extinction of the other member as well.[68]
Abiotic pollination


Flowers that use abiotic, or non-living, vectors use the wind or, much less commonly, water, to move pollen from one flower to the next.[51] In wind-dispersed (anemophilous) species, the tiny pollen grains are carried, sometimes many thousands of kilometres,[69] by the wind to other flowers. Common examples include the grasses, birch trees, along with many other species in the order Fagales,[70] ragweeds, and many sedges. They have no need to attract pollinators and therefore tend not to grow large, showy, or colorful flowers, and do not have nectaries, nor a noticeable scent. Because of this, plants typically have many thousands of tiny flowers which have comparatively large, feathery stigmas; to increase the chance of pollen being received.[65] Whereas the pollen of entomophilous flowers is usually large, sticky, and rich in protein (to act as a "reward" for pollinators), anemophilous flower pollen is typically small-grained, very light, smooth, and of little nutritional value to insects.[71][72] In order for the wind to effectively pick up and transport the pollen, the flowers typically have anthers loosely attached to the end of long thin filaments, or pollen forms around a catkin which moves in the wind. Rarer forms of this involve individual flowers being moveable by the wind (Pendulous), or even less commonly; the anthers exploding to release the pollen into the wind.[71]
Pollination through water (hydrophily) is a much rarer method, occurring in only around 2% of abiotically pollinated flowers.[51] Common examples of this include Calitriche autumnalis, Vallisneria spiralis and some sea-grasses. One characteristic which most species in this group share is a lack of an exine, or protective layer, around the pollen grain.[73] Paul Knuth identified two types of hydrophilous pollination in 1906 and Ernst Schwarzenbach added a third in 1944. Knuth named his two groups 'Hyphydrogamy' and the more common 'Ephydrogamy'.[74] In hyphydrogamy pollination occurs below the surface of the water and so the pollen grains are typically negatively buoyant. For marine plants that exhibit this method the stigmas are usually stiff, while freshwater species have small and feathery stigmas.[75] In ephydrogamy pollination occurs on the surface of the water and so the pollen has a low density to enable floating, though many also use rafts, and are hydrophobic. Marine flowers have floating thread-like stigmas and may have adaptations for the tide, while freshwater species create indentations in the water.[75] The third category, set out by Schwarzenbach, is those flowers which transport pollen above the water through conveyance. This ranges from floating plants, (Lemnoideae), to staminate flowers (Vallisneria). Most species in this group have dry, spherical pollen which sometimes forms into larger masses, and female flowers which form depressions in the water; the method of transport varies.[75]
Mechanisms
Flowers can be pollinated by two mechanisms; cross-pollination and self-pollination. No mechanism is indisputably better than the other as they each have their advantages and disadvantages. Plants use one or both of these mechanisms depending on their habitat and ecological niche.[76]
Cross-pollination
Cross-pollination is the pollination of the carpel by pollen from a different plant of the same species. Because the genetic make-up of the sperm contained within the pollen from the other plant is different, their combination will result in a new, genetically distinct, plant, through the process of sexual reproduction. Since each new plant is genetically distinct, the different plants show variation in their physiological and structural adaptations and so the population as a whole is better prepared for an adverse occurrence in the environment. Cross-pollination, therefore, increases the survival of the species and is usually preferred by flowers for this reason.[44][77]
The principal adaptive function of flowers is the promotion of cross-pollination or outcrossing, a process that allows the masking of deleterious mutations in the genome of progeny. The masking effect of outcrossing sexual reproduction is known as “genetic complementation”.[78] This beneficial effect of outcrossing on progeny is also recognized as hybrid vigor or heterosis. Once outcrossing is established due to the benefits of genetic complementation, subsequent switching to inbreeding becomes disadvantageous because it allows expression of the previously masked deleterious recessive mutations, usually referred to as inbreeding depression. Charles Darwin in his 1889 book The Effects of Cross and Self-Fertilization in the Vegetable Kingdom[79] at the beginning of chapter XII noted “The first and most important of the conclusions which may be drawn from the observations given in this volume, is that generally cross-fertilisation is beneficial and self-fertilisation often injurious, at least with the plants on which I experimented.”
Self-pollination

Self-pollination is the pollination of the carpel of a flower by pollen from either the same flower or another flower on the same plant,[44] leading to the creation of a genetic clone through asexual reproduction. This increases the reliability of producing seeds, the rate at which they can be produced, and lowers the amount energy needed.[80] But, most importantly, it limits genetic variation. In addition, self-pollination causes inbreeding depression, due largely to the expression of recessive deleterious mutations.[81][82]
The extreme case of self-fertilization, when the ovule is fertilized by pollen from the same flower or plant, occurs in flowers that always self-fertilize, such as many dandelions.[83] Some flowers are self-pollinated and have flowers that never open or are self-pollinated before the flowers open; these flowers are called cleistogamous; many species in the genus Viola exhibit this, for example.[84]
Conversely, many species of plants have ways of preventing self-pollination and hence, self-fertilization. Unisexual male and female flowers on the same plant may not appear or mature at the same time, or pollen from the same plant may be incapable of fertilizing its ovules. The latter flower types, which have chemical barriers to their own pollen, are referred to as self-incompatible.[23][85] In Clianthus puniceus, self-pollination is used strategically as an "insurance policy". When a pollinator, in this case a bird, visits C. puniceus, it rubs off the stigmatic covering and allows for pollen from the bird to enter the stigma. If no pollinators visit, however, then the stigmatic covering falls off naturally to allow for the flower's own anthers to pollinate the flower through self-pollination.[80]
Allergies
Pollen is a large contributor to asthma and other respiratory allergies which combined affect between 10 and 50% of people worldwide. This number appears to be growing, as the temperature increases due to climate change mean that plants are producing more pollen[citation needed], which is also more allergenic. Pollen is difficult to avoid, however, because of its small size and prevalence in the natural environment. Most of the pollen which causes allergies is that produced by wind-dispersed pollinators such as the grasses, birch trees, oak trees, and ragweeds; the allergens in pollen are proteins which are thought to be necessary in the process of pollination.[86][87]
Fertilization

Fertilization, also called Synagmy, occurs following pollination, which is the movement of pollen from the stamen to the carpel. It encompasses both plasmogamy, the fusion of the protoplasts, and karyogamy, the fusion of the nuclei. When pollen lands on the stigma of the flower it begins creating a pollen tube which runs down through the style and into the ovary. After penetrating the centre-most part of the ovary it enters the egg apparatus and into one synergid. At this point the end of the pollen tube bursts and releases the two sperm cells, one of which makes its way to an egg, while also losing its cell membrane and much of its protoplasm. The sperm's nucleus then fuses with the egg's nucleus, resulting in the formation of a zygote, a diploid (two copies of each chromosome) cell.[88]
Whereas in fertilization only plasmogamy, or the fusion of the whole sex cells, results, in Angiosperms (flowering plants) a process known as double fertilization, which involves both karyogamy and plasmogamy, occurs. In double fertilization the second sperm cell subsequently also enters the synergid and fuses with the two polar nuclei of the central cell. Since all three nuclei are haploid, they result in a large endosperm nucleus which is triploid.[88]
Seed development

Following the formation of zygote it begins to grow through nuclear and cellular divisions, called mitosis, eventually becoming a small group of cells. One section of it becomes the embryo, while the other becomes the suspensor; a structure which forces the embryo into the endosperm and is later undetectable. Two small primordia also form at this time, that later become the cotyledon, which is used as an energy store. Plants which grow out one of these primordia are called monocotyledons, while those that grow out two are dicotyledons. The next stage is called the Torpedo stage and involves the growth of several key structures, including: the radicle (embryotic root), the epicotyl (embryotic stem), and the hypocotyl, (the root/shoot junction). In the final step vascular tissue develops around the seed.[89]
Fruit development
The ovary, inside which the seed is forming from the ovule, grows into a fruit. All the other main floral parts die during this development, including: the style, stigma, sepals, stamens, and petals. The fruit contains three structures: the exocarp, or outer layer, the mesocarp, or the fleshy part, and the endocarp, or innermost layer, while the fruit wall is called the pericarp. The size, shape, toughness, and thickness varies among different fruit. This is because it is directly connected to the method of seed dispersal; that being the purpose of fruit - to encourage or enable the seed's dispersal and protect the seed while doing so.[89]
Seed dispersal



Following the pollination of a flower, fertilization, and finally the development of a seed and fruit, a mechanism is typically used to disperse the fruit away from the plant.[93] In Angiosperms (flowering plants) seeds are dispersed away from the plant so as to not force competition between the mother and the daughter plants,[94] as well as to enable the colonisation of new areas. They are often divided into two categories, though many plants fall in between or in one or more of these:[95]
Allochory
In allochory, plants use an external vector, or carrier, to transport their seeds away from them. These can be either biotic (living), such as by birds and ants, or abiotic (non-living), such as by the wind or water.[95][96][97]
Biotic vectors
Many plants use biotic vectors to disperse their seeds away from them. This method falls under the umbrella term zoochory, while endozoochory, also known as fruigivory, refers specifically to plants adapted to grow fruit in order to attract animals to eat them. Once eaten they go through typically go through animal's digestive system and are dispersed away from the plant.[97] Some seeds are specially adapted either to last in the gizzard of animals or even to germinate better after passing through them.[98][99] They can be eaten by birds (ornithochory), bats (chiropterochory), rodents, primates, ants (myrmecochory),[100] non-bird sauropsids (saurochory), mammals in general (mammaliochory),[98] and even fish.[101] Typically their fruit are fleshy, have a high nutritional value, and may have chemical attractants as an additional "reward" for dispersers. This is reflected morphologically in the presence of more pulp, an aril, and sometimes an elaiosome (primarily for ants), which are other fleshy structures.[102]
Epizoochory occurs in plants whose seeds are adapted to cling on to animals and be dispersed that way, such as many species in the genus Acaena.[103] Typically these plants seed's have hooks or a viscous surface to easier grip to animals, which include birds and animals with fur. Some plants use mimesis, or imitation, to trick animals into dispersing the seeds and these often have specially adapted colors.[102][104]
The final type of zoochory is called synzoochory, which involves neither the digestion of the seeds, nor the unintentional carrying of the seed on the body, but the deliberate carrying of the seeds by the animals. This is usually in the mouth or beak of the animal (called Stomatochory), which is what is used for many birds and all ants.[105]


Abiotic vectors
In abiotic dispersal plants use the vectors of the wind, water, or a mechanism of their own to transport their seeds away from them.[97][96] Anemochory involves using the wind as a vector to disperse plant's seeds. Because these seeds have to travel in the wind, they are almost always small — sometimes even dust-like, have a high surface-area-to-volume ratio, and are produced in a large number — sometimes up to a million. Plants such as tumbleweeds detach the entire shoot to let the seeds roll away with the wind. Another common adaptation are wings, plumes or balloon-like structures that let the seeds stay in the air for longer and hence travel farther.
In hydrochory plants are adapted to disperse their seeds through bodies of water and so typically are buoyant and have a low relative density with regards to the water. Commonly seeds are adapted morphologically with hydrophobic surfaces, small size, hairs, slime, oil, and sometimes air spaces within the seeds.[102] These plants fall into three categories: ones where seeds are dispersed on the surface of water currents, under the surface of water currents, and by rain landing on a plant.[106]
Автохория
При автохории растения создают свои собственные переносчики для транспортировки семян от них. выталкивают семена Приспособления для этого обычно заключаются в том, что плоды взрываются и баллистически , как, например, у Hura crepitans . [107] а иногда и в создании ползучих диаспор . [102] Из-за относительно небольших расстояний, на которые эти методы могут распространять семена, их часто сочетают с внешним переносчиком. [104]
Эволюция
Хотя наземные растения существуют уже около 425 миллионов лет, первые из них размножились путем простой адаптации своих водных аналогов: спор . В море растения – и некоторые животные – могут просто разбрасывать свои генетические клоны , уплывать и расти в другом месте. Так размножались ранние растения. Но вскоре растения разработали методы защиты этих экземпляров от высыхания и других повреждений, которые даже более вероятны на суше, чем в море. Защита стала семенем , хотя цветок еще не развился. К ранним семенным растениям относятся гинкго и хвойные деревья .

Несколько групп вымерших голосеменных растений , особенно семенных папоротников , были предложены в качестве предков цветковых растений, но не существует убедительных ископаемых свидетельств, показывающих, как именно развивались цветы. назвал это «отвратительной загадкой» Очевидно внезапное появление относительно современных цветов в летописи окаменелостей поставило такую проблему для теории эволюции, что Чарльз Дарвин .
Недавно обнаруженные окаменелости покрытосеменных, такие как Archaefructus , наряду с дальнейшими открытиями ископаемых голосеменных, позволяют предположить, что характеристики покрытосеменных могли быть приобретены в несколько этапов. Возраст раннего ископаемого цветкового растения Archaefructus liaoningensis из Китая составляет около 125 миллионов лет. [108] [109] возрастом 125–130 миллионов лет Еще раньше из Китая встречается Archaefructus sinensis . В 2015 году возраст растения ( Montsechia vidalii , обнаруженного в Испании, возрастом 130 миллионов лет) был заявлен как 130 миллионов лет. [110] В 2018 году ученые сообщили, что самые ранние цветы появились около 180 миллионов лет назад. [111]

Недавний ДНК анализ ( молекулярная систематика ) [112] показывает, что Amborella trichopoda , обнаруженная на тихоокеанском острове Новая Каледония, является единственным видом в сестринской группе по отношению к остальным цветковым растениям, а морфологические исследования показывают, что она имеет черты, которые могли быть характерны для самых ранних цветковых растений. [113]
Помимо убедительных доказательств существования цветов в меловом периоде или незадолго до него , [114] [115] есть косвенные свидетельства существования цветов 250 миллионов лет назад. Химическое вещество, используемое растениями для защиты своих цветов, олеанан , было обнаружено в древних ископаемых растениях, в том числе в гигантоптеридах . [116] которые развились в то время и несут многие черты современных цветковых растений, хотя неизвестно, что они сами являются цветковыми растениями, поскольку детально сохранились только их стебли и колючки; один из самых ранних примеров окаменения .
Сходство в структуре листьев и стеблей может быть очень важным, поскольку цветы генетически представляют собой всего лишь адаптацию нормальных компонентов листьев и стеблей растений, комбинации генов, обычно отвечающих за образование новых побегов. [117] Считается, что самые примитивные цветы имели разное количество частей цветка, часто отделенных друг от друга (но соприкасающихся друг с другом). Цветы имели тенденцию расти по спирали, быть двуполыми (у растений это означает, что в одном цветке есть как мужская, так и женская части), и в них доминировала завязь ( женская часть). По мере того, как цветы становились более продвинутыми, в некоторых вариациях появлялись части, слитые вместе, с гораздо более конкретным количеством и дизайном, а также с определенным полом цветка или растения или, по крайней мере, с «нижней завязью».
Общее предположение состоит в том, что функция цветов с самого начала заключалась в вовлечении животных в процесс размножения. Пыльца может быть рассеяна без ярких цветов и очевидных форм, что, следовательно, будет помехой при использовании ресурсов растения, если только они не принесут какой-либо другой пользы. Одна из предполагаемых причин внезапного, полностью развитого появления цветов заключается в том, что они развивались в изолированной среде, такой как остров или цепочка островов, где растения, несущие их, смогли развить высокоспециализированные отношения с каким-то конкретным животным (оса, например), так же сегодня развиваются многие островные виды. Эти симбиотические отношения, при которых гипотетическая оса переносит пыльцу с одного растения на другое, подобно тому, как это делают сегодня инжирные осы , могли в конечном итоге привести к тому, что и растение(а), и их партнеры развили высокую степень специализации. островная генетика Считается, что является распространенным источником видообразования , особенно когда речь идет о радикальных адаптациях, которые, по-видимому, требуют низших переходных форм. Обратите внимание, что пример с осами не случаен; пчелы, по-видимому, эволюционировавшие специально для симбиотических отношений с растениями, произошли от ос.
Точно так же большая часть плодов , используемых при размножении растений, образуется в результате увеличения частей цветка. Этот плод часто является орудием, которое зависит от животных, желающих его съесть и таким образом разбрасывающих содержащиеся в нем семена.
Хотя многие подобные симбиотические отношения остаются слишком хрупкими, чтобы выдержать конкуренцию с материковыми организмами, цветы оказались необычайно эффективным средством производства, распространившись (каким бы ни было их фактическое происхождение) и став доминирующей формой наземной растительной жизни.
Эволюция цветов продолжается и по сей день; Современные цветы подверглись столь глубокому влиянию человека, что многие из них невозможно опылить в природе. Многие современные одомашненные цветы раньше были простыми сорняками, которые прорастали только тогда, когда почва была потревожена. Некоторые из них имели тенденцию расти вместе с человеческими культурами, а самые красивые не собирались из-за их красоты, у них развивалась зависимость и особая адаптация к человеческой привязанности. [118]
Цвет
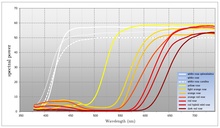
Многие цветущие растения отражают как можно больше света в диапазоне видимых длин волн опылителя, которого растение намеревается привлечь. Цветы, отражающие весь спектр видимого света, обычно воспринимаются как белые человеком-наблюдателем . Важной особенностью белых цветов является то, что они одинаково отражают свет во всем видимом спектре. Хотя многие цветковые растения используют белый цвет для привлечения опылителей, использование цвета также широко распространено (даже внутри одного вида). Цвет позволяет цветковому растению более точно определить опылителя, которого оно стремится привлечь. [ нужна ссылка ] Цветовая модель, используемая в технологии воспроизведения человеческого цвета ( CMYK ), основана на модуляции пигментов, которые делят спектр на широкие области поглощения. Цветущие растения, напротив, способны смещать длину волны точки перехода между поглощением и отражением. Если предположить, что зрительные системы большинства опылителей рассматривают видимый спектр как круговой , то можно сказать, что цветковые растения производят цвет, поглощая свет в одной области спектра и отражая свет в другой области. При использовании CMYK цвет создается в зависимости от амплитуды широких областей поглощения. Цветущие растения, напротив, производят цвет, изменяя частоту (или, скорее, длину волны) отраженного света. Большинство цветов поглощают свет в области спектра от синего до желтого и отражают свет от области спектра от зеленого до красного. Для многих видов цветковых растений именно точка перехода характеризует цвет, который они производят. Цвет можно модулировать, сдвигая точку перехода между поглощением и отражением, и таким образом цветущее растение может указать, какого опылителя оно стремится привлечь. [ нужна ссылка ] Некоторые цветущие растения также обладают ограниченной способностью модулировать области поглощения. Обычно это не так точно, как контроль длины волны. Люди-наблюдатели будут воспринимать это как степень насыщенности (количество белого в цвете).
Классическая таксономия
В таксономии растений , которая изучает классификацию и идентификацию растений, морфология цветков растений широко используется – и использовалась на протяжении тысячелетий. Хотя история систематики растений восходит, по крайней мере, к 300 г. до н.э., начиная с сочинений Теофраста , [120] Фундамент современной науки основан на работах XVIII и XIX веков. [121]
Карл Линней (1707–1778) был шведским ботаником, который большую часть своей трудовой жизни проработал профессором естествознания. В его знаковой книге 1757 года «Виды растений» изложена его система классификации, а также концепция биномиальной номенклатуры , последняя из которых до сих пор используется во всем мире. [121] [примечание 1] Он выделил 24 класса, основываясь главным образом на количестве, длине и расположении тычинок . Первые десять классов напрямую следуют за количеством тычинок ( у Октандрии 8 тычинок и т. д.), [119] в то время как в одиннадцатом классе 11–20 тычинок, а в двенадцатом и тринадцатом классах - 20 тычинок; различаются только местом крепления. Следующие пять классов относятся к длине тычинок, а последние пять - к характеру репродуктивной способности растения; где растет тычинка; и если цветок скрыт или существует вообще (например, у папоротников ). Этот метод классификации, несмотря на свою искусственность, [119] широко использовалась в течение следующих семи десятилетий, прежде чем была заменена системой другого ботаника. [122]
Антуан Лоран де Жюссье (1748–1836) был французским ботаником, чья работа 1787 года Genera plantarum: secundum ordines naturales disposita изложила новый метод классификации растений; Вместо этого они основаны на природных характеристиках. Растения разделяли по количеству семядолей , если таковые имеются , и расположению тычинок. [122] Следующая по значимости система классификации появилась в конце 19 века от ботаников Джозефа Далтона Хукера (1817–1911) и Джорджа Бентама (1800–1884). Они опирались на более ранние работы де Жюссье и Огюстена Пирама де Кандоля мира и разработали систему, которая до сих пор используется во многих гербариях . Растения разделялись на высшем уровне по числу семядолей и характеру цветков, прежде чем разделить на отряды ( семейства ), роды и виды . Эта система классификации была опубликована в трех томах Genera plantarum между 1862 и 1883 годами. [123] В некоторых случаях это наиболее уважаемая и считающаяся «лучшей системой классификации». [124]
После развития научной мысли после работы Дарвина «Происхождение видов » многие ботаники стали использовать больше филогенетических методов, а использование генетического секвенирования , цитологии и палинологии становится все более распространенным. Несмотря на это, морфологические характеристики, такие как характер цветка и соцветия, по-прежнему составляют основу систематики растений. [124] [125]
Символизм

Многие цветы имеют важное символическое значение в западной культуре. [126] Практика присвоения значения цветам известна как флориография . Некоторые из наиболее распространенных примеров включают в себя:
- Красные розы дарят как символ любви, красоты и страсти. [127]
- Маки – символ утешения во время смерти. В Великобритании, Новой Зеландии, Австралии и Канаде красные маки носят в память о солдатах, погибших во время войны.
- Ирисы / лилии используются в погребениях как символ «воскресения/жизни». Он также ассоциируется со звездами (солнцем) и его цветущими/сияющими лепестками.
- Ромашки – символ невинности.

Из-за своего разнообразного и красочного внешнего вида цветы уже давно стали любимым объектом художников. Некоторые из самых знаменитых картин известных художников изображают цветы, например, » Ван Гога или серия «Подсолнухи « Моне Кувшинки» . Цветы также сушат, сублимируют и прессуют для создания долговечных трехмерных произведений цветочного искусства .
Цветы в искусстве также символизируют женские гениталии . [128] как это видно в работах таких художников, как Джорджия О'Киф , Имоджен Каннингем , Вероника Руис де Веласко и Джуди Чикаго , а также в азиатском и западном классическом искусстве. Многие культуры по всему миру имеют заметную тенденцию ассоциировать цветы с женственностью .
Большое разнообразие нежных и красивых цветов вдохновило творчество многих поэтов, особенно эпохи романтизма XVIII–XIX веков . Известные примеры включают Уильяма Вордсворта « Я бродил одиноко как облако» и Уильяма Блейка . «Ах!» Солнце-Цветок .
Их символика во сне также обсуждалась с возможными интерпретациями, включая «потенциал цветения». [129]
Римская богиня цветов, садов и весны — Флора . Греческая богиня весны, цветов и природы — Хлорис .
В индуистской мифологии цветы имеют значительный статус. Вишну, один из трех главных богов индуистской системы, часто изображается стоящим прямо на цветке лотоса . [130] Помимо ассоциации с Вишну, индуистская традиция также считает, что лотос имеет духовное значение. [131] Например, оно фигурирует в индуистских историях творения. [132]
Использование человека
Этот раздел нуждается в дополнительных цитатах для проверки . ( Ноябрь 2018 г. ) |

История показывает, что цветы использовались людьми на протяжении тысячелетий для самых разных целей. Ранний пример этого произошел примерно 4500 лет назад в Древнем Египте , где цветы использовались для украшения женских волос. Цветы также снова и снова вдохновляют искусство, например, в Моне «Кувшинках» или в стихотворении Уильяма Вордсворта о нарциссах , озаглавленном: « Я бродил одиноко, как облако ». [133]

В наше время люди искали способы выращивать, покупать, носить или иным образом находиться рядом с цветами и цветущими растениями, отчасти из-за их приятного внешнего вида и запаха . Во всем мире люди используют цветы, чтобы отметить важные события в своей жизни:
- Для новорожденных или крещений
- В качестве корсажа или бутоньерки, которую носят на общественных мероприятиях или на праздниках.
- В знак любви или уважения
- Для свадебных цветов для свадебного банкета и в качестве украшения свадебных площадок.
- В качестве яркого украшения дома.
- В качестве подарка на память о вечеринках в честь путешествия , вечеринках в честь возвращения домой и подарках «думаю о тебе».
- За похоронные цветы и выражения сочувствия скорбящим
- Для поклонения. В христианстве . цветы алтаря часто украшают церкви [134] В индуистской культуре приверженцы обычно приносят цветы в дар храмам. [135]

Цветы, такие как жасмин, на протяжении веков использовались в качестве замены традиционному чаю в Китае. В последнее время многие другие травы и цветы, традиционно используемые во всем мире, приобретают все большее значение для приготовления различных цветочных чаев . [ нужна ссылка ]
Поэтому люди выращивают цветы вокруг своих домов, посвящают часть своего жизненного пространства цветникам , собирают полевые цветы или покупают коммерчески выращенные цветы у флористов . Производство и торговля цветами поддерживают развивающиеся экономики благодаря их доступности в качестве продукта справедливой торговли . [136]

Цветы дают меньше пищи, чем другие основные части растения ( семена , плоды , корни , стебли и листья ), но все же дают несколько важных овощей и специй . Цветочные овощи включают брокколи , цветную капусту и артишок . Самая дорогая пряность — шафран — состоит из высушенных рылец крокуса . Другие цветочные специи — гвоздика и каперсы . Цветки хмеля используются для ароматизации пива . Цветы бархатцев скармливают цыплятам, чтобы придать их яичным желткам золотисто-желтый цвет, который потребители считают более желательным; как пряность и краситель Сушеные и молотые цветки бархатцев также используются в грузинской кухне . Из цветков одуванчика и бузины часто делают вино. Пчелиная пыльца , собранная у пчел, некоторыми людьми считается здоровой пищей. Мед состоит из цветочного нектара, обработанного пчелами, и его часто называют по типу цветка, например, апельсина мед из цветов , клеверный мед и мед тупело .
Сотни свежих цветов съедобны , но лишь немногие широко продаются в качестве продуктов питания. Их часто добавляют в салаты в качестве гарнира . Цветки тыквы обваливают в сухарях и обжаривают. Некоторые съедобные цветы включают настурцию , хризантему , гвоздику , рогоз , японскую жимолость , цикорий , василек , канну и подсолнечник . [137] Съедобные цветы, такие как маргаритка , роза и фиалка , иногда засахаривают. [138]
Такие цветы, как хризантема, роза, жасмин, жимолость японская и ромашка , выбранные из-за их аромата и лечебных свойств, используются в качестве отваров , смешанных с чаем или сами по себе. [139]
Цветы использовались в погребальных ритуалах с доисторических времен: следы пыльцы были найдены на женской могиле в пещере Эль-Мирон в Испании. [140] Многие культуры связывают цветы с жизнью и смертью, а из-за сезонного возвращения цветы также символизируют возрождение, что может объяснить, почему многие люди возлагают цветы на могилы. Древние греки , как записано в Еврипида пьесе «Финикийки » , возлагали на голову умершего венок из цветов; [141] они также покрывали могилы венками и лепестками цветов. Цветы широко использовались в древнеегипетских захоронениях. [142] и мексиканцы по сей день широко используют цветы во время Дня мертвых. празднования [143] точно так же, как это делали их предки ацтеки .

предоставление

Традиция дарения цветов восходит к доисторическим временам, когда цветы часто имели лечебные и травяные свойства. Археологи обнаружили в нескольких могилах остатки лепестков цветов. Цветы впервые использовались как жертвенные и погребальные предметы. Древние египтяне , а затем греки и римляне использовали цветы. В Египте погребальные предметы примерно 1540 г. до н.э. [ нужна ссылка ] были найдены, на которых были изображены красный мак , желтый Араун , василек и лилии . Сведения о дарении цветов встречаются в китайских писаниях и египетских иероглифах, а также в греческой и римской мифологии . Практика дарения цветов процветала в средние века, когда пары выражали свою привязанность с помощью цветов.
Традиция дарения цветов существует во многих формах. Это важная часть русской культуры и фольклора. Учащиеся часто дарят цветы своим учителям. Дарить желтые цветы в романтических отношениях означает расставание в России. В наши дни цветы часто дарят в виде цветочного букета . [144] [145] [146]
См. также
- Изменение цвета цветка
- Консервация цветов
- Сад
- Список садовых растений
- Эволюционная биология развития растений
- Репродуктивная морфология растений
- Посев
Примечания
- ^ Его ранние работы: Systema Naturae (1735) и Genera plantarum (1737) также оказали влияние в этой области. [119]
Ссылки
Цитаты
- ^ «Стратегии опыления растений» . Лесная служба США . 12 декабря 2022 года. Архивировано из оригинала 16 февраля 2023 года . Проверено 16 февраля 2023 г.
- ^ Байер, Мэнди (9 сентября 2015 г.). «Опылители в ландшафте II: Растения и опылители» . Центр сельского хозяйства, продовольствия и окружающей среды Массачусетского университета в Амхерсте. Архивировано из оригинала 7 января 2024 года.
- ^ «Самоопыление и перекрестное опыление | Биология для специальностей II» . Люмен Обучение . Архивировано из оригинала 21 июля 2020 г. Проверено 21 июля 2020 г.
- ^ Крессвелл, Джулия (2010). Оксфордский словарь происхождения слов . Интернет-архив. Издательство Оксфордского университета . стр. 165 и 172. ISBN. 978-0-19-954793-7 .
- ^ «Определение ЦВЕТЕНИЯ» . www.merriam-webster.com . Архивировано из оригинала 24 июня 2021 г. Проверено 16 июня 2021 г.
- ^ «Морфология | Определение морфологии в Оксфордском словаре на Lexico.com, а также значение морфологии» . Лексико-словари | Английский . Архивировано из оригинала 5 марта 2020 года . Проверено 8 июля 2021 г.
- ^ Де Крен и П. 2010 , с. 4.
- ^ Де Крейн и П. 2010 , стр. 5–6.
- ^ Jump up to: Перейти обратно: а б с Де Крен и П. 2010 , с. 7.
- ^ Jump up to: Перейти обратно: а б Д. Маусет 2016 , стр. 225.
- ^ Jump up to: Перейти обратно: а б Д. Маусет 2016 , стр. 226.
- ^ Jump up to: Перейти обратно: а б Де Крен и П. 2010 , с. 8.
- ^ Биология насекомых и растений
- ^ Д. Маусет, 2016 , стр. 229.
- ^ Де Крен и П. 2010 , с. 14.
- ^ Jump up to: Перейти обратно: а б Саттлер, Р. (1973). Органогенез цветов. Фотографический текстовый атлас . Университет Торонто Пресс. ISBN 978-0-8020-1864-9 .
- ^ Эндресс, Питер К. (25 июля 1996 г.). Разнообразие и эволюционная биология тропических цветов . Издательство Кембриджского университета. п. 11. ISBN 978-0-521-56510-3 .
- ^ Рейнольдс, Джоан; Тампион, Джон (1983). Махровые цветки: научное исследование . Лондон: [Опубликовано для] Политехнического института Центрального Лондона Press [Pembridge Press]. п. 41. ИСБН 978-0-86206-004-6 .
- ^ Де Крен и П. 2010 , с. 25.
- ^ Jump up to: Перейти обратно: а б Веберлинг, Фокко (1992). Морфология цветков и соцветий . Издательство Кембриджского университета. стр. 17–19. ISBN 0-521-25134-6 .
- ^ Д. Маусет, 2016 , стр. 243.
- ^ Де Крен и П. 2010 , с. 410.
- ^ Jump up to: Перейти обратно: а б Д. Маусет 2016 , стр. 239.
- ^ Де Крен и П. 2010 , с. 21.
- ^ «Zantedeschia aethiopica» . Сеть охраны растений Новой Зеландии . Проверено 8 мая 2022 г.
- ^ Д. Маусет, 2016 , стр. 228.
- ^ Де Крейн и П. 2010 , стр. 22–24.
- ^ Преннер, Джернард (февраль 2010 г.). «Цветочные формулы обновлены для регулярного включения в формальные таксономические описания» . Таксон . 59 (1): 241–250. дои : 10.1002/tax.591022 . Архивировано из оригинала 29 марта 2018 г.
- ^ Де Крен и П. 2010 , с. 38.
- ^ Стивен Дауни; Кен Робертсон. «Цифровые цветы: цветочные формулы» . Университет Иллинойса. Архивировано из оригинала 4 марта 2016 года . Проверено 28 января 2014 г.
- ^ «Таксономия растений – Биология 308» . .csbsju.edu . Архивировано из оригинала 24 июня 2014 г.
- ^ Шарма 2009 , стр. 165–166.
- ^ Де Крен и П. 2010 , с. 36.
- ^ Имс, Эй Джей (1961). Морфология покрытосеменных растений . Нью-Йорк: McGraw-Hill Book Co.
- ^ Лейнс, Питер (2010). Цветок и плод: морфология, онтогенез, филогения, функции и экология . Клаудия Эрбар. Штутгарт: Швейцербарт. ISBN 978-3-510-65261-7 . OCLC 678542669 .
- ^ Саттлер, Р. (1988). «Динамический многомерный подход к развитию цветов». В Лейнсе, П.; Такер, С.С. и Эндресс, П.К. (ред.). Аспекты цветочного развития . Берлин: Дж. Крамер/Борнтрегер. стр. 1–6.
- ^ Саттлер Р. и Жен Б. (1992). «Многомерный анализ подтверждает непрерывный взгляд на форму растений». Анналы ботаники . 69 (3): 249–262. doi : 10.1093/oxfordjournals.aob.a088338 .
- ^ Аусин, И.; и др. (2005). «Экологическая регламентация цветения» . Int J Dev Biol . 49 (5–6): 689–705. doi : 10.1387/ijdb.052022ia . ПМИД 16096975 .
- ^ Терк, Ф.; Форнара, Ф.; Купленд, Г. (2008). «Регулирование и идентичность Флоригена: цветущий локус Т выходит в центр внимания» . Ежегодный обзор биологии растений . 59 : 573–594. doi : 10.1146/annurev.arplant.59.032607.092755 . hdl : 11858/00-001M-0000-0012-374F-8 . ПМИД 18444908 . S2CID 39798675 . Архивировано из оригинала 15 декабря 2019 г. Проверено 23 июля 2019 г.
- ^ Сирл, И.; и др. (2006). «Фактор транскрипции FLC обеспечивает реакцию цветения на яровизацию, подавляя компетентность меристемы и системную передачу сигналов у Arabidopsis» . Генс Дев . 20 (7): 898–912. дои : 10.1101/gad.373506 . ПМЦ 1472290 . ПМИД 16600915 .
- ^ Д. Маусет, 2016 , стр. 392–395.
- ^ Бикман, Мадлен; Ньювенхейс, Барт; Ортис-Барриентос, Даниэль; Эванс, Джонатан П. (2016). «Половой отбор у гермафродитов, сперматозоидов и производителей, растений и грибов» . Философские труды: Биологические науки . 371 (1706): 5. doi : 10.1098/rstb.2015.0541 . ISSN 0962-8436 . JSTOR 26143395 . ПМК 5031625 . ПМИД 27619704 .
Единственная цель цветов – секс.
- ^ Уокер 2020 , с. 9.
- ^ Jump up to: Перейти обратно: а б с д Д. Маусет 2016 , стр. 238.
- ^ Тернер, Вивьен (1984). «Пыльца банксии как источник белка в рационе двух австралийских сумчатых Cercartetus nanus и Tarsipes rostratus» . Ойкос . 43 (1): 53–61. Бибкод : 1984Ойкос..43...53Т . дои : 10.2307/3544245 . ISSN 0030-1299 . JSTOR 3544245 . Архивировано из оригинала 24 июня 2021 г. Получено 20 июня 2021 г. - через JSTOR .
[T] медовый опоссум, Tarsipes rostratus (Turner 1983). Это сумчатое животное узкоспециализировано в питании цветами и, как известно, посещает несколько видов Banksia.
- ^ Уокер 2020 , с. 65.
- ^ Jump up to: Перейти обратно: а б Уокер, 2020 , стр. 69–83.
- ^ Уокер 2020 , с. 120.
- ^ Бейкер, Герберт Г. (1 января 1983 г.). «ГЛАВА 2 - Очерк истории антэкологии, или биологии опыления ** На основе, но расширенной и обновленной статьи, опубликованной в NZJ Bot. (Baker, 1979), с разрешения редактора» . Очерк истории антекологии, или биологии опыления . п. 8. дои : 10.1016/B978-0-12-583980-8.50009-0 . ISBN 9780125839808 . Архивировано из оригинала 24 июня 2021 г. Получено 20 июня 2021 г. - через Elsevier Science Direct .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Фридман, Дженнис (2011). «Унесенные ветром: понимание эволюционных переходов между ветром и опылением животными у покрытосеменных» . Новый фитолог . 191 (4): 911–913. дои : 10.1111/j.1469-8137.2011.03845.x . ISSN 0028-646X . JSTOR 20869225 . ПМИД 21834912 .
- ^ Jump up to: Перейти обратно: а б с Акерман, доктор медицинских наук (1 марта 2000 г.). «Абиотическая пыльца и опыление: экологические, функциональные и эволюционные перспективы» . Систематика и эволюция растений . 222 (1): 167–185. Бибкод : 2000PSyEv.222..167A . дои : 10.1007/BF00984101 . ISSN 1615-6110 . S2CID 36015720 . Архивировано из оригинала 21 февраля 2022 г. Проверено 20 июня 2021 г.
- ^ Уокер 2020 , с. 46.
- ^ Д. Маусет, 2016 , стр. 239–240.
- ^ Уокер 2020 , с. 68.
- ^ Кнут, Мюллер и Эйнсворт Дэвис 1906 , стр. 72–80.
- ^ Верикекен, Николас Дж.; Уилсон, Кэрол А.; Хетлинг, Сюзанн; Шульц, Стефан; Банкетов Сергей А.; Мардулин, Патрик (7 декабря 2012 г.). «Преадаптация и эволюция опыления путем сексуального обмана: новый взгляд на правило специализации Коупа» . Труды Королевского общества B: Биологические науки . 279 (1748): 4786–4794. дои : 10.1098/rspb.2012.1804 . ПМК 3497092 . ПМИД 23055065 .
- ^ Уокер 2020 , с. 81.
- ^ Уокер 2020 , стр. 112–113.
- ^ Шеффлер, Ирмгард; Дёттерль, Стефан (2011). «Один день из жизни масличной пчелы: фенология, гнездование и кормовое поведение» (PDF) . Апидология . 42 (3): 409–424. дои : 10.1007/s13592-011-0010-3 . S2CID 29392750 .
- ^ Jump up to: Перейти обратно: а б Уокер, 2020 , стр. 107–108.
- ^ Уокер 2020 , с. 121.
- ^ Уокер 2020 , с. 173.
- ^ Святое говорящее растение! Цветок общается с летучими мышами
- ^ Эквадорский кактус поглощает ультразвук, привлекая летучих мышей к цветам
- ^ Jump up to: Перейти обратно: а б Д. Маусет 2016 , стр. 241.
- ^ Пикалл, Род; Эберт, Дэниел; Полди, Жаклин; Барроу, Рассел А.; Франке, Виттко; Бауэр, Колин С.; Шистль, Флориан П. (2010). «Специфика опылителей, химия цветочного запаха и филогения австралийских сексуально обманчивых орхидей Chiloglottis: последствия для видообразования, управляемого опылителями» . Новый фитолог . 188 (2): 437–450. дои : 10.1111/j.1469-8137.2010.03308.x . ISSN 1469-8137 . ПМИД 20561345 .
- ^ Д. Маусет, 2016 , стр. 240.
- ^ Бава, Канзас (1990). «Взаимодействие растений и опылителей в тропических лесах» . Ежегодный обзор экологии и систематики . 21 : 415. doi : 10.1146/annurev.es.21.110190.002151 . ISSN 0066-4162 . JSTOR 2097031 . Архивировано из оригинала 25 июня 2021 г. Получено 20 июня 2021 г. - через JSTOR.
Во-первых, эффект может быть прямым, так как, например, потеря одного из взаимодействующих партнеров в видоспецифичных взаимодействиях может привести к вымиранию другого.
- ^ Робледо-Арнунсио, Хуан Хосе (апрель 2011 г.). «Ветровое опыление на мезомасштабных расстояниях: исследование сосны обыкновенной» . Новый фитолог . 190 (1): 222–233. дои : 10.1111/j.1469-8137.2010.03588.x . ISSN 0028-646X . ПМИД 21175640 .
- ^ Софиев, М.; Сильямо, П.; Ранта, Х.; Линкосало, Т.; Джагер, С.; Расмуссен, А.; Рантио-Лехтимаки, А.; Северова Е.; Кукконен, Дж. (01 января 2013 г.). «Численная модель выброса и рассеивания пыльцы березы в атмосфере. Описание эмиссионного модуля» . Международный журнал биометеорологии . 57 (1): 45–58. Бибкод : 2013IJBm...57...45S . дои : 10.1007/s00484-012-0532-z . ISSN 1432-1254 . ПМЦ 3527742 . ПМИД 22410824 . Архивировано из оригинала 21 февраля 2022 г. Проверено 15 июня 2021 г.
- ^ Jump up to: Перейти обратно: а б Кнут, Мюллер и Эйнсворт Дэвис 1906 , стр. 68–72.
- ^ Хёхерль, Николь; Сиде, Рейнхольд; Иллис, Ингрид; Гетшенбергер, Хайке; Тауц, Юрген (01 февраля 2012 г.). «Оценка пищевой ценности кукурузы для медоносных пчел» . Журнал физиологии насекомых . 58 (2): 278–285. дои : 10.1016/j.jinsphys.2011.12.001 . ISSN 0022-1910 . ПМИД 22172382 . Архивировано из оригинала 24 июня 2021 г. Проверено 20 июня 2021 г.
- ^ Уокер 2020 , с. 51.
- ^ Кнут, Мюллер и Эйнсворт Дэвис 1906 , стр. 68–69.
- ^ Jump up to: Перейти обратно: а б с Кокс, Пол Алан (1 ноября 1988 г.). «Гидрофильное опыление» . Ежегодный обзор экологии и систематики . 19 (1): 261–279. doi : 10.1146/annurev.es.19.110188.001401 . ISSN 0066-4162 . Архивировано из оригинала 24 июня 2021 г. Проверено 20 июня 2021 г.
- ^ Уокер 2020 , с. 36–37.
- ^ Д. Маусет, 2016 , стр. 222.
- ^ Бернштейн Х., Байерли Х.К., Хопф Ф.А., Мишод Р.Э. Генетические повреждения, мутации и эволюция пола. Наука. 1985, 20 сентября; 229 (4719): 1277-81. doi: 10.1126/science.3898363. ПМИД 3898363
- ^ Дарвин, CR 1876. Эффекты перекрестного и самооплодотворения в растительном царстве. Лондон: Джон Мюррей». darwin-online.org.uk.
- ^ Jump up to: Перейти обратно: а б Уокер 2020 , с. 42.
- ^ Чарльзуорт, Дебора; Уиллис, Джон Х. (ноябрь 2009 г.). «Генетика инбредной депрессии» . Обзоры природы Генетика . 10 (11): 783–796. дои : 10.1038/nrg2664 . ISSN 1471-0056 . ПМИД 19834483 .
- ^ Бернштейн, Х.; Хопф, ФА; Мишод, Р.Э. (1987), Молекулярная основа эволюции пола , Достижения в генетике, том. 24, Elsevier, стр. 323–370, doi : 10.1016/s0065-2660(08)60012-7 , ISBN. 978-0-12-017624-3 , PMID 3324702
- ^ МакПик, Тамара М.; Ван, Сяньчжун (2007). «Размножение одуванчика (Taraxacum officinale) в среде с повышенным содержанием CO2» . Наука о сорняках . 55 (4): 334. doi : 10.1614/WS-07-021 . ISSN 0043-1745 . JSTOR 4539580 . S2CID 86250272 . Архивировано из оригинала 25 июня 2021 г. Получено 20 июня 2021 г. - через JSTOR .
- ^ Гилмартин, AJ; Браун, Грегори К. (1985). «Клейстогамия у Tillandsia capillaris (Bromeliaceae)» . Биотропика . 17 (3): 256. Бибкод : 1985Биотр..17..256Г . дои : 10.2307/2388227 . ISSN 0006-3606 . JSTOR 2388227 . Архивировано из оригинала 25 июня 2021 г. Проверено 20 июня 2021 г.
- ^ Уокер 2020 , с. 34.
- ^ Чен, Мяолинь; Сюй, Цзе; Дэвис, Дебора; Ши, Цзяньсинь; Рен, Канг; Сирл, Иэн; Чжан, Дабин (2016). «Происхождение и функциональное предсказание пыльцевых аллергенов растений» . Физиология растений . 172 (1): 341–357. дои : 10.1104/стр.16.00625 . ISSN 0032-0889 . ПМК 5074609 . ПМИД 27436829 .
- ^ Констибль, Хуанита; Сандро, Люк; Ли, Ричард Э. (2008). «Ага! Повышенный риск аллергии на пыльцу в северном полушарии» . Журнал преподавания естественных наук в колледже . 37 (4): 82–89. ISSN 0047-231X . JSTOR 42992782 . Архивировано из оригинала 25 июня 2021 г. Проверено 20 июня 2021 г.
- ^ Jump up to: Перейти обратно: а б Д. Маусет 2016 , стр. 234.
- ^ Jump up to: Перейти обратно: а б Д. Маусет, 2016 , стр. 235–237.
- ^ Вайман, Таррин; Келли, Дэйв (2017). «Количественная оценка распространения семян птицами и опоссумами в низинных лесах Новой Зеландии» . Новозеландский журнал экологии . 41 (1). дои : 10.20417/nzjecol.41.4 . Архивировано из оригинала 30 июня 2021 г. Проверено 1 июля 2021 г.
- ^ Сайп, Тимоти В.; Линнерут, Эми Р. (1995). «Внутривидовая изменчивость самарской морфологии и летного поведения Acer saccharinum (Aceraceae)» . Американский журнал ботаники . 82 (11): 1412–1419. дои : 10.2307/2445868 . ISSN 0002-9122 . JSTOR 2445868 . Архивировано из оригинала 9 июля 2021 г. Проверено 1 июля 2021 г.
- ^ Маунт, Энн; Пикеринг, Кэтрин Марина (01 октября 2009 г.). «Проверка способности одежды выступать в качестве переносчика чужеродных семян на охраняемых территориях» . Журнал экологического менеджмента . 91 (1): 168–179. дои : 10.1016/j.jenvman.2009.08.002 . hdl : 10072/29656 . ISSN 0301-4797 . ПМИД 19717222 . Архивировано из оригинала 29 июня 2021 г. Проверено 1 июля 2021 г.
- ^ Д. Маусет, 2016 , стр. 248.
- ^ Боулер Д.Э., Бентон Т.Г. (май 2005 г.). «Причины и последствия стратегий расселения животных: связь индивидуального поведения с пространственной динамикой». Биологические обзоры Кембриджского философского общества . 80 (2): 205–25. дои : 10.1017/S1464793104006645 . ПМИД 15921049 . S2CID 39351147 .
- ^ Jump up to: Перейти обратно: а б Стрелка 1972 , с. 71.
- ^ Jump up to: Перейти обратно: а б Забудьте, Пьер-Мишель (2005). Судьба семян: хищничество, расселение и формирование рассады . Уоллингфорд, Оксфордшир, Великобритания: Паб CABI. п. 21. ISBN 0-85199-806-2 . OCLC 54803650 . Архивировано из оригинала 21 февраля 2022 г. Проверено 1 июля 2021 г.
Семена на растениях могут распространяться с помощью различных первичных механизмов распространения, включая абиотические факторы, такие как ветер или баллистическая проекция, или биотические факторы, такие как птицы, питающиеся фруктами.
- ^ Jump up to: Перейти обратно: а б с Норсуорси, Джейсон К.; Гриффит, Грифф; Гриффин, Терри; Багаватианнан, Мутхукумар; Гбур, Эдвард Э. (2014). «Перемещение устойчивого к глифосату амаранта Палмера (Amaranthus Palmen) в полевых условиях и его влияние на урожайность хлопкового ворса: данные, подтверждающие стратегию нулевого порога» . Наука о сорняках . 62 (2): 237–249. дои : 10.1614/WS-D-13-00145.1 . ISSN 0043-1745 . JSTOR 43700653 . S2CID 86013249 . Архивировано из оригинала 9 июля 2021 г. Проверено 1 июля 2021 г. - через JSTOR .
Ветер и вода являются распространенными механизмами распространения абиотических семян, но существует несколько биотических механизмов распространения, таких как перемещение через животных путем адгезии (эпизоохия) или проглатывания (эндозоохория) и даже движение в результате деятельности человека (антропохория).
- ^ Jump up to: Перейти обратно: а б Д. Маусет, 2016 , стр. 244–247.
- ^ Травесет, А.; Риера, Н.; Мас, Р.Э. (2001). «Прохождение через кишки птиц вызывает межвидовые различия в характеристиках прорастания семян» . Функциональная экология . 15 (5): 669–675. Бибкод : 2001FuEco..15..669T . дои : 10.1046/j.0269-8463.2001.00561.x . ISSN 1365-2435 . Архивировано из оригинала 9 июля 2021 г. Проверено 1 июля 2021 г.
Многие исследования показывают, что прорастание происходит более успешно после того, как семена проходят через пищеварительный тракт плодоядных животных.
- ^ Лендьель, Сабольч; Гоув, Аарон Д.; Латимер, Эндрю М.; Майер, Джонатан Д.; Данн, Роберт Р. (20 февраля 2010 г.). «Конвергентная эволюция распространения семян муравьями, а также филогения и биогеография цветковых растений: глобальный обзор» . Перспективы экологии, эволюции и систематики растений . 12 (1): 43–55. дои : 10.1016/j.ppees.2009.08.001 . ISSN 1433-8319 . Архивировано из оригинала 2 июля 2021 г. Проверено 1 июля 2021 г.
- ^ Хямяляйнен, Анни; Бродли, Кейт; Дрогини, Аманда; Хейнс, Джессика А.; Лэмб, Клейтон Т.; Бутин, Стэн; Гилберт, Софи (2017). «Экологическое значение вторичного распространения семян хищниками» . Экосфера . 8 (2): e01685. Бибкод : 2017Ecosp...8E1685H . дои : 10.1002/ecs2.1685 . ISSN 2150-8925 .
- ^ Jump up to: Перейти обратно: а б с д Хоу, Х.Ф. и Смоллвуд Дж. (1982). «Экология распространения семян» (PDF) . Ежегодный обзор экологии и систематики . 13 : 201–228. doi : 10.1146/annurev.es.13.110182.001221 . Архивировано из оригинала (PDF) 13 мая 2006 г.
- ^ Саймон, Делавэр; Уэлен, Массачусетс; Маккей, Д.А. (2000). «Разнообразие Acaena (Rosaceae) в Южной Австралии» . Журнал Ботанического сада Аделаиды . 19 : 55–73. ISSN 0313-4083 . JSTOR 23874441 . Архивировано из оригинала 28 июня 2021 г. Проверено 1 июля 2021 г. - через JSTOR .
- ^ Jump up to: Перейти обратно: а б Виттоз, Паскаль; Энглер, Робин (7 февраля 2008 г.). «Расстояния распространения семян: типология, основанная на способах распространения и особенностях растений» (PDF) . Ботаника Гельветика . 117 (2): 109–124. дои : 10.1007/s00035-007-0797-8 . S2CID 2339616 . Архивировано (PDF) из оригинала 27 июня 2021 года . Проверено 1 июля 2021 г.
- ^ Стрелка 1972 , стр. 24–25.
- ^ Jump up to: Перейти обратно: а б Хислоп, Джейд; Троусдейл, Сэм (2012). «Обзор гидрохории (распространения семян водой) с последствиями для восстановления прибрежных территорий» . Журнал гидрологии (Новая Зеландия) . 51 (2): 137–152. ISSN 0022-1708 . JSTOR 43945038 . Архивировано из оригинала 9 июля 2021 г. Проверено 1 июля 2021 г.
- ^ Jump up to: Перейти обратно: а б Фельдкамп, Сьюзен (2006). Современная биология . США: Холт, Райнхарт и Уинстон . п. 618.
- ^ Льюис, Сьюзен К. (17 апреля 2007 г.). «Цветы современные и древние» . PBS онлайн . Архивировано из оригинала 7 сентября 2009 года . Проверено 30 августа 2010 г.
- ^ НОВА . 17 апреля 2007 г. ПБС. ВГБХ. Первый цветок .
- ^ Томсон, Хелен (17 августа 2015 г.). «Окаменелые останки старейшего в мире цветка обнаружены в Испании» . Хранитель . Архивировано из оригинала 3 марта 2017 г.
- ^ Китайская академия наук (18 декабря 2018 г.). «Цветы возникли на 50 миллионов лет раньше, чем считалось ранее» . ЭврекАлерт! . Архивировано из оригинала 18 декабря 2018 года . Проверено 18 декабря 2018 г.
- ^ Солтис, Дуглас Э.; Солтис, Памела С. (2004). «Амборелла — не «базальное покрытосеменное растение»? Не так быстро» . Американский журнал ботаники . 91 (6): 997–1001. дои : 10.3732/ajb.91.6.997 . ПМИД 21653455 .
- ^ «Растения южной части Тихого океана могут быть недостающим звеном в эволюции цветковых растений» . Eurekalert.org. 17 мая 2006 г. Архивировано из оригинала 14 мая 2011 г. Проверено 30 августа 2010 г.
- ^ Габботт, Сара (1 августа 2017 г.). «Первый цветок выглядел так?» . Новости Би-би-си . Архивировано из оригинала 1 августа 2017 года . Проверено 1 августа 2017 г.
- ^ Соке, Эрве; и др. (1 августа 2017 г.). «Предковый цветок покрытосеменных и его раннее разнообразие» . Природные коммуникации . 8 : 16047. Бибкод : 2017NatCo...816047S . дои : 10.1038/ncomms16047 . ПМЦ 5543309 . ПМИД 28763051 .
- ^ «Нефтяные окаменелости дают ключ к разгадке эволюции цветов» . Sciencedaily.com. 05 апреля 2001 г. Архивировано из оригинала 19 августа 2010 г. Проверено 30 августа 2010 г.
- ^ «Ответ на извечный вопрос об эволюции цветов» . Unisci.com. 15 июня 2001 г. Архивировано из оригинала 10 июня 2010 г. Проверено 30 августа 2010 г.
- ^ Май 2005 г., Роберт Рой Бритт, 26 лет (26 мая 2005 г.). «Человеческая привязанность изменила эволюцию цветов» . www.livscience.com . Архивировано из оригинала 06 сентября 2021 г. Проверено 06 сентября 2021 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Jump up to: Перейти обратно: а б с д Шарма 2009 , с. 21.
- ^ Шарма 2009 , с. 8.
- ^ Jump up to: Перейти обратно: а б Шарма 2009 , с. 10.
- ^ Jump up to: Перейти обратно: а б Шарма 2009 , с. 22.
- ^ Шарма 2009 , стр. 24–27.
- ^ Jump up to: Перейти обратно: а б Шарма 2009 , с. 11.
- ^ Шарма 2009 , с. 96.
- ^ «Значения цветов» . Цветочный журнал . 28 мая 2018 г. Архивировано из оригинала 31 июля 2020 г. Проверено 12 декабря 2018 г.
- ^ Одет, Мэри. «Розы и их значение» . Архивировано из оригинала 26 августа 2014 года . Проверено 22 августа 2014 г.
- ^ Фраунфелтер, Андреа. Цветочный символизм как женская сексуальная метафора (Тезис). Университет Восточного Мичигана. Архивировано из оригинала 10 августа 2014 г.
- ^ Ди, Нерис (1995). Ваши сны и что они означают: как понять тайный язык сна . Лондон: Торнсоны. п. 142. ИСБН 978-0-7225-3218-8 . ОСЛК 33208041 . Архивировано из оригинала 21 февраля 2022 г. Проверено 15 июня 2021 г.
- ^ «Вишну» . BBC.co.uk. 24 августа 2009 г. Архивировано из оригинала 28 октября 2010 г. Проверено 30 августа 2010 г.
- ^ «Любимый цветок Бога» . Индуизм сегодня. Архивировано из оригинала 13 апреля 2009 г. Проверено 30 августа 2010 г.
- ^ «Лотос» . Theosociety.org. Архивировано из оригинала 10 сентября 2017 г. Проверено 30 августа 2010 г.
- ^ Фогден, Майкл; Фогден, Патрисия (2018). Естественная история цветов . Издательство Техасского университета A&M. п. 1. ISBN 978-1623496449 .
- ^ Уилсон, Аделаида Б (1967). Цветы для вашей церкви . Место издания не указано: Barrows & Co. p. 26. OCLC 500430357 . Архивировано из оригинала 28 июня 2021 г. Проверено 15 июня 2021 г.
- ^ «ПОСЕЩЕНИЕ ИНДУССКОГО ХРАМА; РУКОВОДСТВО ДЛЯ НАЧИНАЮЩИХ» . Индуизм сегодня . Апрель 1991 г. Архивировано из оригинала 16 апреля 2021 г. Проверено 27 ноября 2018 г.
- ↑ Fairtrade Foundation , Цветочные фермеры и рабочие , по состоянию на 23 марта 2023 г.
- ^ Вуд, Зои (30 июня 2017 г.). «Вкусно цветут – съедобные цветы – самая горячая тенденция лета в еде» . Хранитель . Архивировано из оригинала 27 ноября 2018 года . Проверено 27 ноября 2018 г.
- ^ Брэдли, Сью (8 мая 2015 г.). «Как кристаллизовать цветы, как в викторианском стиле» . Телеграф . Архивировано из оригинала 11 января 2022 г.
- ^ Вонг, Джеймс (7 мая 2017 г.). «Вырастите свои собственные травяные чаи» . Хранитель . Архивировано из оригинала 27 ноября 2018 года . Проверено 27 ноября 2018 г.
- ^ «Скорбящие каменного века возлагали цветы на могилы » . Телеграф . 10 мая 2015 г. Архивировано из оригинала 11 января 2022 г.
- ^ «Погребальные обряды» . Хелленика Мир. Архивировано из оригинала 29 ноября 2018 года . Проверено 28 ноября 2018 г.
- ^ Хейс, Кристофер Б. (2011). Смерть в Железном веке II и в Первой книге Исайи . Мор Зибек. п. 302. ИСБН 978-3-16-150785-4 . Архивировано из оригинала 28 декабря 2019 г. Проверено 28 ноября 2018 г.
- ^ Дэй, Фрэнсис Энн (2003). Латиноамериканские и латиноамериканские голоса в литературе . Гринвуд. п. 72 . ISBN 978-0-313-32394-2 .
- ^ Автор (11 августа 2015 г.). «Увлекательная традиция дарить цветы» . Полевые цветы Лас-Вегас . Архивировано из оригинала 16 марта 2019 г. Проверено 15 марта 2019 г.
- ^ «Блог о межкультурной риторике: Цветы в России» . веб-сайт Stanford.edu . Архивировано из оригинала 13 апреля 2019 г. Проверено 15 марта 2019 г.
- ^ «Народное искусство в России и Украине | Новости и информация» . Архивировано из оригинала 24 марта 2019 г. Проверено 15 марта 2019 г.
Источники
- Де Крен, Ронс; П., Луи (2010). Цветочные схемы . Кембридж: Издательство Кембриджского университета. дои : 10.1017/cbo9780511806711 . ISBN 978-0-511-80671-1 .
- Кнут, Пол; Мюллер, Герман; Эйнсворт Дэвис-младший (1906). Справочник по опылению цветов: на основе работы Германа Мюллера «Оплодотворение цветов насекомыми» . Том. 1. Кларендон Пресс. дои : 10.5962/bhl.title.54973 . OCLC 1841036 . Архивировано из оригинала 21 февраля 2022 г. Проверено 15 июня 2021 г.
- Фрич, Феликс Юджин; Солсбери, EJ (Эдвард Джеймс) (1920). Знакомство со строением и размножением растений . Библиотека Корнелльского университета. Лондон: G. Bell and Sons Ltd.
- Уокер, Тимоти (2020). Опыление: прочная связь между растением и опылителем . Издательство Принстонского университета . ISBN 978-0-691-20375-1 .
- Д. Маусет, Джеймс (2016). Ботаника: Введение в биологию растений (6-е изд.). Джонс и Бартлетт Обучение. ISBN 978-1-284-07753-7 .
- Пийл, Л. ван дер (1972). Принципы распространения у высших растений (2-е изд.). Берлин Гейдельберг: Springer-Verlag. ISBN 978-3-642-96108-3 . Архивировано из оригинала 9 июля 2021 г. Проверено 1 июля 2021 г.
- Шарма, ОП (2009). Таксономия растений (2-е изд.). Нью-Дели, Индия: Tata McGraw Hill Education Private Limited . ISBN 978-0070141599 .
Дальнейшее чтение
- Бухманн, Стивен (2016). Причина появления цветов: их история, культура, биология и как они меняют нашу жизнь . Скрибнер. ISBN 978-1-4767-5553-3 .
- Исав, Кэтрин (1965). Анатомия растений (2-е изд.). Нью-Йорк: Джон Уайли и сыновья. ISBN 978-0-471-24455-4 .
- Грейсон, Род-Айленд (1994). Развитие цветов . Издательство Оксфордского университета. ISBN 978-0-19-506688-3 .
- Лейнс П. и Эрбар К. (2010). Цветок и фрукты . Штутгарт: Издательство Schweizerbart Science. ISBN 978-3-510-65261-7 .
- Саттлер, Р. (1973). Органогенез цветов. Фотографический текстовый атлас . Университет Торонто Пресс. ISBN 978-0-8020-1864-9 .
Внешние ссылки
 Цитаты, связанные с цветами, в Wikiquote
Цитаты, связанные с цветами, в Wikiquote - Информационная сеть местных растений
| Субдисциплины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растений Группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Воспроизведение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||