морская трава
| Морские травы Временной диапазон: | |
|---|---|
 | |
| Zostera marina – самый распространенный вид морских водорослей в Северном полушарии. | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Трахеофиты |
| Клэйд : | покрытосеменные растения |
| Клэйд : | Однодольные |
| Заказ: | Алисмалес Р.Бр. бывший Берхт. и Дж.Пресл |
| Семьи | |
См. Таксономию | |
Морские травы — единственные цветковые растения , произрастающие в морской среде. Существует около 60 видов полностью морских морских трав, принадлежащих к четырем семействам ( Posidoniaceae , Zosteraceae , Hydrocharitaceae и Cymodoceaceae ), все они относятся к порядку Alismatales (в кладе однодольных ). [1] Морские травы произошли от наземных растений , которые повторно заселили океан 70–100 миллионов лет назад.
Название морская трава происходит от многих видов с длинными и узкими листьями , которые растут за счет корневища и часто распространяются по большим « лугам », напоминающим луга ; многие виды внешне напоминают наземные травы семейства Poaceae .
Как и все автотрофные растения, морские травы фотосинтезируют в подводной фотической зоне , и большинство из них встречается в мелких и защищенных прибрежных водах, закрепленных на песчаном или илистом дне. Большинство видов подвергаются подводному опылению и завершают свой жизненный цикл под водой. Хотя ранее считалось, что это опыление осуществлялось без опылителей и исключительно за счет дрейфа морского течения, оказалось, что это неверно, по крайней мере, для одного вида, Thalassia testudinum , который реализует смешанную биотико-абиотическую стратегию. Ракообразные (такие как крабы, Majidae zoae , Thalassinidea zoea ) и личинки силлидных многощетинковых червей были обнаружены с пыльцевыми зернами, причем растение производит питательные слизистые комки пыльцы, которые привлекают и прилипают к ним вместо нектара, как это делают наземные цветы. [2]
Морские травы образуют густые подводные луга из морских водорослей , которые являются одними из самых продуктивных экосистем в мире. Они функционируют как важные поглотители углерода. [3] и обеспечивают среду обитания и пищу для разнообразных морских обитателей, сравнимых с разнообразием коралловых рифов .
Обзор
[ редактировать ]Морские травы представляют собой парафилетическую группу морских покрытосеменных растений эволюционировали , которые параллельно три-четыре раза от наземных растений обратно в море. Для определения вида морской травы можно использовать следующие характеристики:
- Он обитает в устье реки или в морской среде и больше нигде.
- Опыление . происходит под водой специальной пыльцой
- Семена, которые распространяются как биотическими , так и абиотическими агентами, производятся под водой. [4]
- Виды морских водорослей имеют специализированные листья с уменьшенной кутикулой и эпидермисом без устьиц , который является основной фотосинтетической тканью.
- Корневище в или подземный стебель играют важную роль закреплении .
- Корни могут жить в бескислородной среде и зависеть от транспорта кислорода от листьев и корневищ, но также играют важную роль в процессах переноса питательных веществ . [4] [5]
Морские травы оказывают глубокое влияние на физическую, химическую и биологическую среду прибрежных вод. [4] Хотя морские травы оказывают неоценимые экосистемные услуги , выступая в качестве питательной среды для различных организмов и стимулируя коммерческое рыболовство , многие аспекты их физиологии недостаточно изучены. В прибрежных водах Северной Америки встречается 26 видов морских трав. [6] Несколько исследований показали, что среда обитания морских водорослей сокращается во всем мире. [7] [8] Десять видов морских водорослей находятся под повышенным риском исчезновения (14% всех видов морских водорослей), при этом три вида относятся к категории находящихся под угрозой исчезновения . Утрата морских водорослей и деградация биоразнообразия морских водорослей будут иметь серьезные последствия для морского биоразнообразия и населения, которое зависит от ресурсов и экосистемных услуг, которые обеспечивают морские водоросли. [9] [4]
Морские травы образуют важные прибрежные экосистемы . [10] Нахождение под угрозой исчезновения этих морских лугов во всем мире, которые обеспечивают пищу и среду обитания для многих морских видов , вызывает необходимость защиты и понимания этих ценных ресурсов. [11]
Эволюция
[ редактировать ]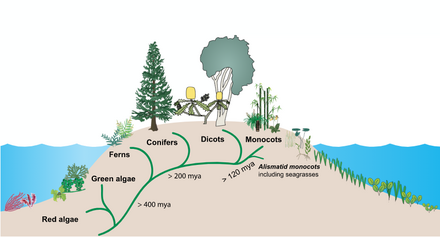
Около 140 миллионов лет назад морские травы произошли от ранних однодольных растений, которым удалось покорить морскую среду. [11] Однодольные — травянистые и травянистоцветковые растения (покрытосеменные), семена которых обычно содержат только один зародышевый лист или семядолю . [12]
Наземные растения произошли, возможно, уже 450 миллионов лет назад из группы зеленых водорослей . [13] Затем морские травы произошли от наземных растений, которые мигрировали обратно в океан. [14] [15] Примерно между 70 и 100 миллионами лет назад три независимые линии морских водорослей ( Hydrocharitaceae , Cymodoceaceae complex и Zosteraceae ) произошли от одной линии однодольных цветковых растений. [16]
Другие растения, колонизировавшие море, такие как растения солончаков , мангровые заросли и морские водоросли , имеют более разнообразные эволюционные линии. Несмотря на небольшое видовое разнообразие, морским травам удалось колонизировать континентальные шельфы всех континентов, кроме Антарктиды. [17]
Недавнее секвенирование геномов Zostera marina и Zostera muelleri позволило лучше понять покрытосеменных растений адаптацию к морю. [18] [19] Во время эволюционного возвращения в океан различные гены были потеряны (например, устьичные гены) или уменьшены (например, гены, участвующие в синтезе терпеноидов ), а другие были восстановлены, например, гены, участвующие в сульфатировании . [19] [11]
Геномная информация показала, что адаптация к морской среде обитания была достигнута за счет радикальных изменений в клеточной стенки . составе [18] [19] Однако клеточные стенки морских трав изучены недостаточно. В дополнение к наследственным чертам наземных растений можно было бы ожидать, что процесс адаптации к новой среде, обусловленный средой обитания, характеризуется множеством абиотических (высокое количество соли) и биотических (различные травоядные растения и бактериальная колонизация) стрессоров. [11] Клеточные стенки морских трав кажутся сложной комбинацией особенностей, известных как у покрытосеменных наземных растений, так и у морских макроводорослей, с новыми структурными элементами. [11]
Таксономия
[ редактировать ]Сегодня морские травы представляют собой полифилетическую группу морских покрытосеменных растений, насчитывающую около 60 видов в пяти семействах ( Zosteraceae , Hydrocharitaceae , Posidoniaceae , Cymodoceaceae и Ruppiaceae ), которые принадлежат к порядку Alismatales в соответствии с системой IV группы филогении покрытосеменных . [20] Род Ruppia , который встречается в солоноватой воде, не рассматривается всеми авторами как «настоящие» морские травы, а некоторые авторы относят его к Cymodoceaceae. [21] Система APG IV и веб-страница со списком растений [22] не разделяйте это семейное задание. [11]
| Семья | Изображение | Роды | Описание |
|---|---|---|---|
| Зостеровые | Семейство Zosteraceae , также известное как семейство морских водорослей , включает два рода, содержащие 14 морских видов. Он встречается в умеренных и субтропических прибрежных водах, наибольшее разнообразие наблюдается вокруг Кореи и Японии. Итого по видам: | ||
 | Филлоспадикс | ||
 | Опоясывающий лишай | ||
| Гидрохаритовые | Семейство Hydrocharitaceae , также известное как ленточные травы , включает канадские водоросли и лягушачьи укусы. Семейство включает как пресноводных, так и морских обитателей, хотя из шестнадцати известных в настоящее время родов только три являются морскими. [23] Они встречаются по всему миру в самых разных средах обитания, но в основном это тропические виды. Итого по видам: | ||
 | Энгалус | ||
 | Галофила | ||
 | Талассия | ||
| Посидониевые | Семейство Posidoniaceae включает один род с двумя-девятью морскими видами, встречающимися в морях Средиземного моря и у южного побережья Австралии . Всего видов: от 2 до 9 | ||
 | Посидония | ||
| Цимодоцеевые | Семейство Cymodoceaceae , также известное как ламантиновая трава , включает только морские виды. [24] Некоторые систематики не признают это семейство. Итого по видам: | ||
 | Амфиболис | ||
 | Цимодокея | ||
 | Халодул | ||
 | Сирингий | ||
 | Талассодендрон | ||
| Всего видов: | |||
Сексуальная вербовка
[ редактировать ]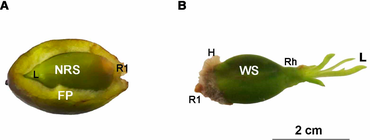
дисперсия, адгезия и расселение
Популяциям морских водорослей в настоящее время угрожают различные антропогенные стрессоры . [26] [8] Способность морских трав справляться с возмущениями окружающей среды в некоторой степени зависит от генетической изменчивости , которая достигается посредством полового рекрутирования . [27] [28] [29] Формируя новых особей, морские травы увеличивают свое генетическое разнообразие и, следовательно, способность колонизировать новые территории и адаптироваться к изменениям окружающей среды. [30] [31] [32] [33] [34] [25] [ чрезмерное цитирование ]
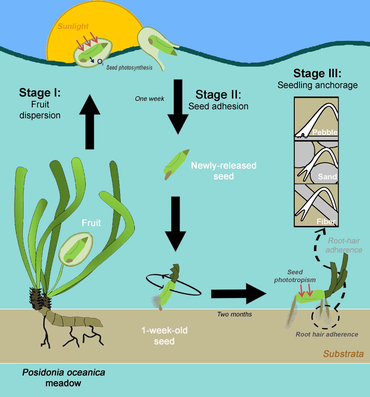
Морские травы имеют противоположные стратегии колонизации . [35] Некоторые морские травы образуют семенные банки из мелких семян с твердыми околоплодниками , которые могут оставаться в стадии покоя в течение нескольких месяцев. Эти морские травы, как правило, недолговечны и могут быстро восстанавливаться после нарушений, не прорастая далеко от материнских лугов (например, виды Halophila , Halodule виды Cymodocea , виды Zostera , виды и виды Heterozostera ). [35] [36] Напротив, другие морские травы образуют дисперсные побеги . Такая стратегия типична для долгоживущих морских трав, которые могут образовывать плавучие плоды с крупными неспящими внутри семенами, например, роды Posidonia sp., Enhalus sp. и Thalassia sp. [35] [37] Соответственно, семена долгоживущих морских трав обладают большей рассеивающей способностью по сравнению с семенами короткоживущих видов. [38] что позволяет вывести виды за пределы неблагоприятных световых условий за счет рассадного развития родительских лугов. [25]
Морская трава Posidonia Oceanica (L.) Delile — один из старейших и крупнейших видов на Земле. Особь может образовывать луга шириной около 15 км и иметь возраст от сотен до тысяч лет. [39] P. Oceanica Луга играют важную роль в поддержании геоморфологии средиземноморского побережья, что, среди прочего, делает эту морскую траву приоритетной средой обитания для сохранения. [40] В настоящее время цветение и пополнение P. Oceanica происходят чаще, чем ожидалось в прошлом. [41] [42] [43] [44] [45] Кроме того, эта морская трава обладает уникальными приспособлениями, повышающими ее выживаемость во время пополнения. Большое количество питательных веществ, содержащихся в семенах этой морской травы, поддерживает рост побегов и корней даже до первого года развития рассады. [39] В первые месяцы прорастания , когда развитие листьев слабое, семена P. Oceanica проявляют фотосинтетическую активность, что увеличивает скорость их фотосинтеза и, таким образом, максимизирует успех приживления рассады. [46] [47] Сеянцы также проявляют высокую морфологическую пластичность в процессе корневой системы. развития [48] [49] образуя липкие корневые волоски , которые помогают закрепиться на каменистых отложениях. [41] [50] [51] Однако многие факторы, связанные с половым привлечением P. Oceanica, остаются неизвестными, например, когда активен фотосинтез в семенах или как семена могут оставаться прикрепленными к субстрату и сохраняться на нем до тех пор, пока их корневая система не разовьется полностью. [25]
Приливно-отливная и сублиторальная
[ редактировать ]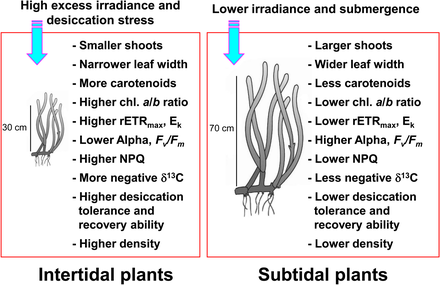
Морские травы, встречающиеся в приливных и сублиторальных зонах, подвергаются сильно меняющимся условиям окружающей среды из-за приливных изменений. [53] [54] Морские травы сублиторальной зоны чаще подвергаются воздействию низкой освещенности из-за множества природных и антропогенных воздействий, которые уменьшают проникновение света за счет увеличения плотности взвешенных непрозрачных материалов. Условия субприливной освещенности можно оценить с высокой точностью с помощью искусственного интеллекта, что позволяет более быстро смягчить последствия, чем это было возможно при использовании методов на месте . [55] Морские травы в приливной зоне регулярно подвергаются воздействию воздуха и, следовательно, испытывают экстремально высокие и низкие температуры, высокую фотоингибирующую радиацию и стресс от высыхания по сравнению с морскими водорослями сублиторальной зоны. [54] [56] [57] Такие экстремальные температуры могут привести к значительному вымиранию морских водорослей, когда морские травы подвергаются воздействию воздуха во время отлива. [58] [59] [60] Стресс от высыхания во время отлива считается основным фактором, ограничивающим распространение морских водорослей в верхней приливной зоне. [61] Морские травы, обитающие в приливной зоне, обычно меньше, чем в сублиторальной зоне, чтобы свести к минимуму последствия стресса, вызванного вылетом. [62] [59] Приливные морские травы также демонстрируют светозависимые реакции, такие как снижение эффективности фотосинтеза и усиление фотозащиты в периоды высокой освещенности и воздействия воздуха. [63] [64]

Напротив, морские травы в сублиторальной зоне адаптируются к условиям пониженной освещенности, вызванной ослаблением и рассеянием света из-за наложения водного столба и взвешенных частиц. [66] [67] Морские травы в глубокой сублиторальной зоне обычно имеют более длинные листья и более широкие листовые пластинки, чем водоросли в мелководной сублиторальной или литоральной зоне, что обеспечивает больший фотосинтез, что, в свою очередь, приводит к большему росту. [57] Морские травы также реагируют на пониженное освещение, увеличивая содержание хлорофилла и уменьшая соотношение хлорофиллов a/b, чтобы повысить эффективность поглощения света за счет эффективного использования большого количества длин волн. [68] [69] [70] Поскольку морские травы в приливной и сублиторальной зонах находятся в совершенно разных условиях освещения, они демонстрируют совершенно разные фотоакклиматические реакции, направленные на максимизацию фотосинтетической активности и фотозащиты от избыточного облучения. [ нужна ссылка ]
Морские травы усваивают большое количество неорганического углерода для достижения высокого уровня производства. [71] [72] Морские макрофиты , включая морские водоросли, используют как CO2 , так и CO2. HCO - 3 ( бикарбонат ) для фотосинтетического восстановления углерода. [73] [74] [75] Несмотря на воздействие воздуха во время отлива, морские травы в приливной зоне могут продолжать фотосинтезировать, используя CO 2 в воздухе. [76] Таким образом, состав неорганических источников углерода для фотосинтеза морских водорослей, вероятно, различается у растений приливной и сублиторальной зоны. Поскольку соотношение стабильных изотопов углерода в тканях растений меняется в зависимости от неорганических источников углерода для фотосинтеза, [77] [78] Морские травы в приливной и сублиторальной зонах могут иметь разные диапазоны соотношения стабильных изотопов углерода.
Луга с водорослями
[ редактировать ]

Заросли /луга морских водорослей могут быть как моновидовыми (состоящими из одного вида), так и смешанными зарослями. В районах с умеренным климатом обычно доминируют один или несколько видов (например, угри Zostera marina в Северной Атлантике), тогда как тропические зарегистрировано до тринадцати видов гряды обычно более разнообразны: на Филиппинах . [ нужна ссылка ]
Заросли морских водорослей представляют собой разнообразные и продуктивные экосистемы и могут содержать сотни связанных видов всех типов , например, молодь и взрослые рыбы , эпифитные и свободноживущие макроводоросли и микроводоросли , моллюски , щетинистые черви и нематоды . Первоначально считалось, что лишь немногие виды питаются непосредственно листьями морских водорослей (отчасти из-за их низкого содержания питательных веществ), но научные обзоры и усовершенствованные методы работы показали, что травоядные водоросли являются важным звеном в пищевой цепи, кормящим сотни видов, включая зеленых черепах. , дюгони , ламантины , рыбы , гуси , лебеди , морские ежи и крабы . Некоторые виды рыб, которые посещают морские травы и питаются ими, выращивают молодь в соседних мангровых зарослях или коралловых рифах .
Морские травы задерживают осадок и замедляют движение воды, вызывая осаждение взвешенных отложений. Улавливание осадков приносит пользу кораллам , поскольку уменьшает нагрузку на них и улучшает фотосинтез как кораллов, так и морских водорослей. [79]
Морские травы, хотя их часто упускают из виду, предоставляют ряд экосистемных услуг . [80] [81] Морские травы считаются инженерами экосистем . [82] [15] [14] Это означает, что растения изменяют экосистему вокруг себя. Эта корректировка происходит как в физической, так и в химической форме. Многие виды морских водорослей образуют обширную подземную сеть корней и корневищ , которые стабилизируют отложения и уменьшают береговую эрозию . [83] Эта система также помогает насыщать осадки кислородом, создавая благоприятную среду для обитающих в отложениях организмов . [82] Морские травы также улучшают качество воды , стабилизируя тяжелые металлы, загрязняющие вещества и избыток питательных веществ. [84] [15] [14] Длинные стебли морских трав замедляют движение воды, что снижает энергию волн и обеспечивает дополнительную защиту от береговой эрозии и штормовых нагонов . Кроме того, поскольку морские травы являются подводными растениями, они производят значительное количество кислорода, который насыщает кислородом толщу воды. На эти луга приходится более 10% общего объема запасов углерода в океане. На гектар он удерживает вдвое больше углекислого газа, чем тропические леса, и может улавливать около 27,4 миллиона тонн CO 2 ежегодно. [85]
Луга с водорослями служат пищей для многих морских травоядных. Морские черепахи, ламантины, рыбы-попугаи, рыбы-хирурги, морские ежи и острицы питаются морскими травами. Многие другие более мелкие животные питаются эпифитами и беспозвоночными, обитающими на стеблях морских водорослей и среди них. [86] Луга с водорослями также обеспечивают физическую среду обитания в районах, которые в противном случае были бы лишены какой-либо растительности. Из-за этой трехмерной структуры толщи воды многие виды занимают места обитания морских водорослей в качестве убежища и корма. Подсчитано, что 17 видов коралловых рифовых рыб проводят всю свою молодую стадию жизни исключительно на равнинах, покрытых водорослями. [87] Эти места обитания также служат местом нагула ценных для коммерческого и рекреационного рыболовства видов, в том числе острого окуня ( Mycteroperca microlepis ), красного барабана, обыкновенного снука и многих других. [88] [89] Некоторые виды рыб используют луга с водорослями и проходят различные стадии жизненного цикла. В недавней публикации доктор Росс Боучек и его коллеги обнаружили, что две очень популярные плоские рыбы, обыкновенный снук и пятнистая морская форель, обеспечивают необходимую среду обитания для кормления во время размножения. [90] Половое размножение чрезвычайно затратно с энергетической точки зрения, чтобы его можно было осуществлять за счет накопленной энергии; поэтому для полного воспроизводства им требуются луга с водорослями в непосредственной близости. [90] многие коммерчески важные беспозвоночные Кроме того, в местах обитания морских водорослей также обитают , включая заливных гребешков ( Argopecten irradians ), мечехвостов и креветок . Харизматическую фауну также можно увидеть, посетив места обитания морских водорослей. К этим видам относятся ламантины Вест-Индии , зеленые морские черепахи и различные виды акул. Высокое разнообразие морских организмов, обитающих в местах обитания морских водорослей, делает их туристической достопримечательностью и важным источником дохода для многих прибрежных стран вдоль Мексиканского залива и Карибского бассейна.

 Белопятнистые иглобрюхи , часто встречаются в районах с водорослями.
Белопятнистые иглобрюхи , часто встречаются в районах с водорослями.- Подводные кадры луга с водорослями, бычьего гуся и морского угря


Микробиом морских водорослей
[ редактировать ]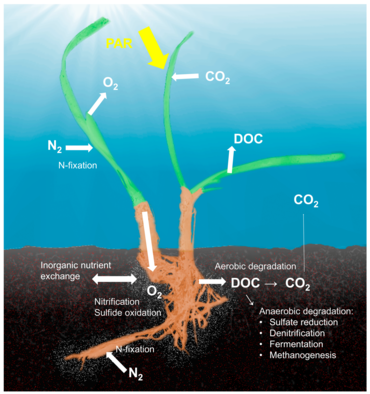
Голобионт морских водорослей
[ редактировать ]Концепция холобионта , подчеркивающая важность и взаимодействие микробного хозяина с ассоциированными микроорганизмами и вирусами и описывающая их функционирование как единой биологической единицы, [93] был исследован и обсужден для многих модельных систем, хотя существует серьезная критика концепции, которая определяет разнообразные симбиозы хозяина и микроба как единую биологическую единицу. [94] Концепции холобионта и гологенома эволюционировали со времени первоначального определения. [95] и нет никаких сомнений в том, что симбиотические микроорганизмы имеют решающее значение для биологии и экологии хозяина, обеспечивая витаминами, энергией и неорганическими или органическими питательными веществами, участвуя в защитных механизмах или управляя эволюцией хозяина. [96]
Хотя большая часть работ по взаимодействиям хозяина и микроба была сосредоточена на животных системах, таких как кораллы, губки или человек, существует значительный объем литературы по голобионтам растений . [97] Связанные с растениями микробные сообщества влияют на оба ключевых компонента приспособленности растений: рост и выживание. [98] и формируются наличием питательных веществ и защитными механизмами растений. [99] Было описано, что в нескольких средах обитания обитают микробы, связанные с растениями, включая ризоплану (поверхность корневой ткани), ризосферу (периферию корней), эндосферу (внутри ткани растения) и филлосферу (общая площадь надземной поверхности). . [91] Микробное сообщество в ризосфере P. Oceanica демонстрирует такую же сложность, как и наземные среды обитания, которые содержат тысячи таксонов на грамм почвы. Напротив, в химическом составе ризосферы P. Oceanica доминировало присутствие сахаров, таких как сахароза и фенольные соединения. [100]
Клеточные стенки
[ редактировать ]
морских водорослей Клеточные стенки содержат те же полисахариды , что и наземные растения покрытосеменных , такие как целлюлоза. [101] Однако клеточные стенки некоторых морских трав характеризуются сульфатированными полисахаридами. [102] [103] что является общим признаком макроводорослей из групп красных , бурых , а также зеленых водорослей . В 2005 году было высказано предположение, что морские покрытосеменные растения восстановили способность синтезировать сульфатированные полисахариды. [102] Еще одной уникальной особенностью клеточных стенок морских трав является наличие необычных пектиновых полисахаридов, называемых апиогалактуронанами . [104] [105] [11]
Помимо полисахаридов, гликопротеины семейства гликопротеинов, богатых гидроксипролином , [106] являются важными компонентами клеточных стенок наземных растений. Высокогликозилированные белки арабиногалактана представляют интерес из-за их участия как в архитектуре стенок, так и в процессах клеточной регуляции. [107] [108] Белки арабиногалактана широко распространены в семенных растениях. [108] а также были обнаружены у папоротников , плаунов и мхов . [109] Структурно они характеризуются крупными полисахаридными фрагментами , состоящими из арабиногалактанов (обычно более 90% молекулы), которые ковалентно связаны через гидроксипролин с относительно небольшими белково-пептидными остовами (обычно менее 10% молекулы). [108] Различные модификации гликанов были идентифицированы у разных видов и тканей, и было высказано предположение, что они влияют на физические свойства и функции. В 2020 году AGP были впервые выделены и структурно охарактеризованы из морской травы. [110] Хотя общая структура основной цепи арабиногалактановых белков наземных растений сохраняется, гликановые структуры демонстрируют уникальные особенности, указывающие на роль арабиногалактановых белков морских водорослей в осморегуляции . [111] [11]
Другими компонентами вторичных стенок растений являются сшитые фенольные полимеры, называемые лигнином , которые отвечают за механическое укрепление стенок. В морских травах этот полимер также был обнаружен, но часто в меньших количествах по сравнению с наземными растениями покрытосеменными. [112] [113] [114] [115] [11] Таким образом, клеточные стенки морских трав, по-видимому, содержат комбинации особенностей, известных как у покрытосеменных наземных растений, так и у морских макроводорослей, а также новые структурные элементы. Сушеные листья морских водорослей могут быть полезны для изготовления бумаги или в качестве изоляционного материала, поэтому знание состава клеточной стенки имеет определенную технологическую значимость. [11]
Угрозы и сохранение
[ редактировать ]Несмотря на то, что морские травы покрывают лишь 0,1–0,2% поверхности океана, они образуют критически важные экосистемы. Как и многие другие регионы океана, морские травы столкнулись с ускоряющимся глобальным сокращением численности. [116] С конца 19-го века было потеряно более 20% мировой площади морских водорослей, при этом потеря зарослей морских водорослей происходит со скоростью 1,5% каждый год. [117] Из 72 видов морских водорослей, встречающихся в мире, примерно четверть (15 видов) можно считать находящимися под угрозой исчезновения или в состоянии, близком к угрожаемому, в Красном списке видов, находящихся под угрозой исчезновения МСОП . [118] Угрозы включают сочетание природных факторов, таких как ураганы и болезни, и антропогенного происхождения, включая разрушение среды обитания , загрязнение окружающей среды и изменение климата . [116]
Безусловно, наиболее распространенной угрозой для морских водорослей является деятельность человека. [119] [120] В прибрежных районах в результате деятельности человека пострадало до 67 видов (93%) морских трав. [118] Такие виды деятельности, как освоение прибрежных земель, катание на моторных лодках и методы рыболовства, такие как траление , либо физически уничтожают заросли морских водорослей, либо увеличивают мутность воды, вызывая их отмирание. Поскольку морские травы предъявляют одни из самых высоких требований к освещению среди покрытосеменных растений, на них сильно влияют условия окружающей среды, которые изменяют прозрачность воды и блокируют свет. [121]
На морские травы также негативно влияет изменение глобальных климатических условий. Участившиеся погодные явления, повышение уровня моря и повышение температуры в результате глобального потепления — все это может привести к массовой потере морских водорослей. Дополнительной угрозой для зарослей морских водорослей является внедрение чужеродных видов. Среди зарослей морских водорослей во всем мире зафиксировано как минимум 28 неместных видов. из этих инвазивных видов большинство (64%) оказывают негативное воздействие на экосистему. Документально подтверждено, что [121]
Другой важной причиной исчезновения морских водорослей является эвтрофикация прибрежных районов . Быстро растущая плотность населения вдоль береговой линии привела к высокой нагрузке биогенными веществами в прибрежных водах из-за сточных вод и других последствий развития. Увеличение биогенной нагрузки создает ускоряющийся каскад прямых и косвенных эффектов, которые приводят к сокращению численности морских водорослей. Хотя некоторое воздействие высоких концентраций питательных веществ, особенно азота и фосфора , может привести к увеличению продуктивности морских водорослей, высокие уровни питательных веществ могут также стимулировать быстрый рост макроводорослей и эпифитов на мелководье, а также фитопланктона на большей глубине. В ответ на высокий уровень питательных веществ макроводоросли образуют плотные пологи на поверхности воды, ограничивая доступ света к донным морским травам. [122] Цветение водорослей , вызванное эвтрофикацией, также приводит к гипоксическим условиям, к которым морские травы также очень восприимчивы. Поскольку прибрежные отложения, как правило, бескислородны , морские травы должны снабжать кислородом свои подземные корни либо посредством фотосинтеза , либо путем диффузии кислорода в толще воды. Когда вода, окружающая водоросли, становится гипоксичной, то же самое происходит и с тканями морских водорослей. Гипоксические условия отрицательно влияют на рост и выживаемость морских трав: показано, что морские травы, подвергшиеся воздействию гипоксических условий, имеют снижение скорости фотосинтеза, усиление дыхания и замедление роста. Гипоксические условия могут в конечном итоге привести к отмиранию морских водорослей, что создает цикл положительной обратной связи , при котором разложение органического вещества еще больше снижает количество кислорода, присутствующего в толще воды. [122]
Возможные траектории популяций морских водорослей были изучены в Средиземном море . Эти исследования показывают, что присутствие морских водорослей зависит от физических факторов, таких как температура, соленость, глубина и мутность, а также от природных явлений, таких как изменение климата и антропогенное давление. Хотя есть исключения, регресс был общей тенденцией во многих районах Средиземного моря. По оценкам, сокращение на 27,7% вдоль южного побережья Лацио , сокращение на 18–38% в бассейне Северного Средиземноморья, сокращение на 19–30% на лигурийском побережье с 1960-х годов и сокращение на 23% во Франции за последние 50 лет. В Испании основной причиной регресса стала человеческая деятельность, такая как незаконный траловый промысел и аквакультура . Было обнаружено, что районы со средней и высокой степенью антропогенного воздействия пострадали от более серьезного сокращения. В целом было высказано предположение, что с 1879 года исчезло 29% известных площадных популяций морских водорослей. Сокращение этих площадей предполагает, что, если потепление в Средиземноморском бассейне продолжится, это может привести к функциональному исчезновению видов морских водорослей. Posidonia Oceanica в Средиземноморье к 2050 году. Ученые предположили, что выявленные ими тенденции являются частью широкомасштабной тенденции во всем мире. [123]
Усилия по сохранению необходимы для выживания видов морских водорослей. Несмотря на то, что в области сохранения морских водорослей необходимо преодолеть множество проблем, есть некоторые серьезные, которые можно решить. Осведомленность общества о том, что такое морские травы и их значение для благополучия человека, невероятно важна. По мере того как большинство людей становятся более урбанизированными, они все больше оторваны от мира природы. Это допускает заблуждения и непонимание экологии морских водорослей и ее важности. Кроме того, сложно получать и хранить информацию о статусе и состоянии популяций морских водорослей. Учитывая большое количество популяций по всему миру, сложно составить карту нынешних популяций. Еще одна проблема, с которой приходится сталкиваться при сохранении морских водорослей, — это способность выявлять угрожающие виды деятельности в местном масштабе. Кроме того, в условиях постоянно растущего населения человечества необходимо сбалансировать потребности людей, а также потребности планеты. Наконец, сложно проводить научные исследования в поддержку сохранения морских водорослей. На изучение морских трав тратятся ограниченные усилия и ресурсы. [124] Это наблюдается в таких регионах, как Индия и Китай , где практически нет планов по сохранению популяций морских водорослей. Однако сохранение и восстановление морских водорослей может способствовать достижению 16 из 17 целей ООН в области устойчивого развития . [125]
В ходе исследования по сохранению морских водорослей в Китае ученые сделали несколько предложений о том, как лучше сохранять морские водоросли. Они предложили включить заросли морских водорослей в программу охраны природы Китая, как это сделано в других странах. Они призвали китайское правительство запретить мелиорацию земель в районах рядом с зарослями морских водорослей или в них, сократить количество и размер культурных прудов, контролировать плотовую аквакультуру и улучшить качество отложений, создать заповедники морских водорослей, повысить осведомленность рыбаков о зарослях морских водорослей. и политикам, а также провести восстановление морских водорослей. [126] Аналогичные предложения были сделаны в Индии, где ученые предположили, что участие общественности имеет важное значение. Кроме того, ученые, общественность и правительственные чиновники должны работать в тандеме над интеграцией традиционных экологических знаний и социокультурных практик для разработки природоохранной политики. [127]
Всемирный день морских водорослей — ежегодное мероприятие, проводимое 1 марта с целью повышения осведомленности о морских водорослях и их важных функциях в морской экосистеме. [128] [129]
См. также
[ редактировать ]- Алисмалес
- Синий углерод
- Соленое болото
- Мангровые заросли
- Морское возвращение
- Питомник среды обитания
- Средство просмотра океанских данных : содержит набор данных о глобальном распределении морских трав.
Ссылки
[ редактировать ]- ^ Томлинсон и Варго (1966). «О морфологии и анатомии черепаховой травы Thalassia testudinum (Hydrocharitaceae). I. Вегетативная морфология» . Бюллетень морской науки . 16 : 748–761.
- ^ ван Туссенбрук, Бригитта И.; Вильямиль, Нора; Маркес-Гусман, Юдит; Вонг, Рикардо; Монрой-Веласкес, Л. Вероника; Солис-Вайс, Вивианна (29 сентября 2016 г.). «Экспериментальные доказательства опыления морских цветов фауной беспозвоночных» . Природные коммуникации . 7 (1): 12980. Бибкод : 2016NatCo...712980V . дои : 10.1038/ncomms12980 . ISSN 2041-1723 . ПМК 5056424 . ПМИД 27680661 .
- ^ «39 способов спасти планету — возвышенные водоросли» . Радио Би-би-си 4 . Би-би-си . Проверено 12 февраля 2022 г.
- ^ Перейти обратно: а б с д Папенброк, Ютта (2012). «Основные особенности филогении, физиологии и метаболизма морских трав: что делает их особенными?» . ISRN Ботаника . 2012 : 1–15. дои : 10.5402/2012/103892 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 . - ^ Ларкум AWD, Р. Дж. Орт и К. М. Дуарте (2006) Морская трава: биология, экология и охрана , Спрингер, Нидерланды.
- ^ «Морские травы» .
- ^ Орт, Роберт Дж.; Каррутерс, ТИМ Дж.Б.; Деннисон, Уильям К.; Дуарте, Карлос М.; Фуркурин, Джеймс В.; Черт возьми, Кеннет Л.; Хьюз, А. Рэндалл; Кендрик, Гэри А.; Кенворти, В. Джадсон; Олярник, Сюзанна; Шорт, Фредерик Т.; Уэйкотт, Мишель; Уильямс, Сьюзен Л. (2006). «Глобальный кризис экосистем морских водорослей» . Бионаука . 56 (12): 987. doi : 10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2 . ISSN 0006-3568 . S2CID 4936412 .
- ^ Перейти обратно: а б Уэйкотт, М.; Дуарте, CM; Каррутерс, TJB; Орт, Р.Дж.; Деннисон, WC; Олярник С.; Калладин, А.; Фуркурен, Дж.В.; Черт возьми, КЛ; Хьюз, Арканзас; Кендрик, Джорджия; Кенворти, WJ; Короткий, FT; Уильямс, СЛ (2009). «Ускорение исчезновения морских трав по всему миру угрожает прибрежным экосистемам» . Труды Национальной академии наук . 106 (30): 12377–12381. Бибкод : 2009PNAS..10612377W . дои : 10.1073/pnas.0905620106 . ПМК 2707273 . ПМИД 19587236 .
- ^ Шорт, Фредерик Т.; Полидоро, Бет; Ливингстон, Сюзанна Р.; Карпентер, Кент Э.; Бандейра, Саломан; Буджанг, Джапар Сидик; Калумпонг, Хилконида П.; Каррутерс, Тим Дж.Б.; Коулз, Роберт Г.; Деннисон, Уильям К.; Эрфтемейер, Пол Л.А.; Фортес, Мигель Д.; Фриман, Аарен С.; Джагтап, Т.Г.; Камаль, Абу Хена М.; Кендрик, Гэри А.; Джадсон Кенворти, В.; Ла Нафи, Яю А.; Насутион, Ичван М.; Орт, Роберт Дж.; Пратеп, Анчана; Санчиангко, Джоннелл К.; Туссенбрук, Бригитта ван; Вергара, Шейла Г.; Уэйкотт, Мишель; Зиман, Джозеф К. (2011). «Оценка риска исчезновения видов морских водорослей в мире» (PDF) . Биологическая консервация . 144 (7): 1961–1971. Бибкод : 2011BCons.144.1961S . дои : 10.1016/j.biocon.2011.04.010 . S2CID 32533417 .
- ^ Хемминга, Массачусетс, и Дуарте, ред. CM (2000). «Морские травы в среде обитания человека», в журнале «Экология морских водорослей» (Кембридж: издательство Кембриджского университета), 248–291.
- ^ Перейти обратно: а б с д и ж г час я дж к Пфайфер, Лукас; Классен, Биргит (2020). «Клеточная стенка морских трав: захватывающая, необычная и чистый холст для будущих исследований» . Границы в науке о растениях . 11 : 588754. doi : 10.3389/fpls.2020.588754 . ПМЦ 7644952 . ПМИД 33193541 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Рудалл, Паула Дж .; Бузго, Матьяс (2002). «Эволюционная история однодольного листа» . Генетика развития и эволюция растений . Специальные тома Ассоциации систематики. Том. 20020544. стр. 431–458. дои : 10.1201/9781420024982.ch23 . ISBN 978-0-415-25790-9 . , в Cronk, Bateman & Hawkins (2002)
- ^ Кнаут, Л. Пол; Кеннеди, Мартин Дж. (2009). «Позднедокембрийское позеленение Земли». Природа . 460 (7256): 728–732. Бибкод : 2009Natur.460..728K . дои : 10.1038/nature08213 . ПМИД 19587681 . S2CID 4398942 .
- ^ Перейти обратно: а б с Орт; и др. (2006). «Глобальный кризис экосистем морских водорослей» . Бионаука . 56 (12): 987–996. doi : 10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2 . hdl : 10261/88476 . S2CID 4936412 .
- ^ Перейти обратно: а б с Папенброк, Дж (2012). «Основные особенности филогении, физиологии и метаболизма морских водорослей: что делает их такими видами?». Международная сеть научных исследований : 1–15.
- ^ Лес, Д.Х., Клеланд, М.А. и Уэйкотт, М. (1997) «Филогенетические исследования Alismatidae, II: эволюция морских покрытосеменных (морских трав) и гидрофилия». Систематическая ботаника 22 (3): 443–463.
- ^ Орт, Роберт Дж.; Каррутерс, ТИМ Дж.Б.; Деннисон, Уильям К.; Дуарте, Карлос М.; Фуркурин, Джеймс В.; Черт возьми, Кеннет Л.; Хьюз, А. Рэндалл; Кендрик, Гэри А.; Кенворти, В. Джадсон; Олярник, Сюзанна; Шорт, Фредерик Т.; Уэйкотт, Мишель; Уильямс, Сьюзен Л. (2006). «Глобальный кризис экосистем морских водорослей» . Бионаука . 56 (12): 987. doi : 10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2 . ISSN 0006-3568 . S2CID 4936412 .
- ^ Перейти обратно: а б Ли, Хуэйтинг; Голич, Агнешка А.; Байер, Филипп Э.; Цзяо, Юаньнянь; Тан, Хайбао; Патерсон, Эндрю Х.; Саблок, Гаурав; Кришнарадж, Рахул Р.; Чан, Чон-Кит Кеннет; Бэтли, Жаклин; Кендрик, Гэри А.; Ларкум, Энтони В.Д.; Ральф, Питер Дж.; Эдвардс, Дэвид (2016). «Геном вида морских водорослей южного полушария (Zostera muelleri)» . Физиология растений . 172 (1): 272–283. дои : 10.1104/стр.16.00868 . ПМК 5074622 . ПМИД 27373688 .
- ^ Перейти обратно: а б с Олсен, Джанин Л.; и др. (2016). «Геном морской травы Zostera marina демонстрирует адаптацию покрытосеменных растений к морю» . Природа . 530 (7590): 331–335. Бибкод : 2016Natur.530..331O . дои : 10.1038/nature16548 . ПМИД 26814964 . S2CID 3713147 .
- ^ «Обновление классификации филогенетических групп покрытосеменных для порядков и семейств цветковых растений: APG IV» . Ботанический журнал Линнеевского общества . 181 : 1–20. 2016. дои : 10.1111/boj.12385 . S2CID 7498637 .
- ^ Уилкин, Пол; Мэйо, Саймон Дж. (30 мая 2013 г.). Ранние события в эволюции однодольных растений . Издательство Кембриджского университета. ISBN 9781107244603 .
- ^ Список растений (2020). Руппия. Доступно онлайн по адресу: http://www.theplantlist.org/1.1/browse/A/Ruppiaceae/Ruppia/ (по состоянию на 22 сентября 2020 г.).
- ^ Кристенхус, Мартен Дж. М.; Бинг, Джеймс В. (20 мая 2016 г.). «Число известных видов растений в мире и его ежегодное увеличение» . Фитотаксы . 261 (3): 201. doi : 10.11646/phytotaxa.261.3.1 . ISSN 1179-3163 .
- ^ Уэйкотт, Мишель; МакМахон, Кэтрин; Лавери, Пол (2014). Путеводитель по морским травам южного умеренного пояса . Издательство CSIRO. ISBN 9781486300150 .
- ^ Перейти обратно: а б с д и Герреро-Месегер, Лаура; Санс-Ласаро, Карлос; Марин, Арнальдо (2018). «Понимание сексуального вовлечения одного из старейших и крупнейших организмов на Земле, морской травы Posidonia Oceanica» . ПЛОС ОДИН . 13 (11): e0207345. Бибкод : 2018PLoSO..1307345G . дои : 10.1371/journal.pone.0207345 . ПМК 6239318 . ПМИД 30444902 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Коллиер, CJ; Уэйкотт, М. (2014). «Крайние температуры замедляют рост морских водорослей и вызывают смертность». Бюллетень о загрязнении морской среды . 83 (2): 483–490. Бибкод : 2014MarPB..83..483C . doi : 10.1016/j.marpolbul.2014.03.050 . ПМИД 24793782 .
- ^ Ройш, ТБХ; Элерс, А.; Хаммерли, А.; Червь, Б. (2005). «Восстановление экосистемы после экстремальных климатических явлений, усиленное генотипическим разнообразием» . Труды Национальной академии наук . 102 (8): 2826–2831. Бибкод : 2005PNAS..102.2826R . дои : 10.1073/pnas.0500008102 . ПМК 549506 . ПМИД 15710890 .
- ^ Герреро-Месегер, Лаура; Санс-Ласаро, Карлос; Марин, Арнальдо (2018). «Понимание сексуального вовлечения одного из старейших и крупнейших организмов на Земле, морской травы Posidonia Oceanica» . ПЛОС ОДИН . 13 (11): e0207345. Бибкод : 2018PLoSO..1307345G . дои : 10.1371/journal.pone.0207345 . ПМК 6239318 . ПМИД 30444902 .
- ^ Элерс, А.; Червь, Б.; Ройш, ТБХ (2008). «Важность генетического разнообразия угрей Zostera marina для ее устойчивости к глобальному потеплению» . Серия «Прогресс в области морской экологии» . 355 : 1–7. Бибкод : 2008MEPS..355....1E . дои : 10.3354/meps07369 .
- ^ Орт, Роберт Дж.; Харвелл, Мэтью С.; Инглис, Грэм Дж. (2006). «Экология семян морских водорослей и процессов распространения морских водорослей». Морские травы: биология, экология и охрана . стр. 111–133. дои : 10.1007/978-1-4020-2983-7_5 . ISBN 978-1-4020-2942-4 .
- ^ Ван Дейк, Дж. К.; Ван Туссенбрук, BI; Хименес-Дюран, К.; Маркес-Гусман, Дж.Дж.; Оуборг, Дж. (2009). «Высокий уровень потока генов и низкая популяционная генетическая структура связаны с высоким потенциалом распространения тропических морских покрытосеменных растений» . Серия «Прогресс в области морской экологии» . 390 : 67–77. Бибкод : 2009MEPS..390...67В . дои : 10.3354/meps08190 . hdl : 2066/75120 .
- ^ Межправительственная группа экспертов по изменению климата, под ред. (2014). «Резюме для политиков». Изменение климата 2013 – Физическая научная основа . стр. 1–30. дои : 10.1017/CBO9781107415324.004 . hdl : 10818/26263 . ISBN 9781107415324 .
- ^ МакМахон, Кэтрин; Ван Дейк, Кор-Джент; Руис-Монтойя, Леонардо; Кендрик, Гэри А.; Краусс, Зигфрид Л.; Уэйкотт, Мишель; Верден, Дженнифер; Лоу, Райан; Стэттон, Джон; Браун, Элоиза; Дуарте, Карлос (2014). «Экология движения морских трав» . Труды Королевского общества B: Биологические науки . 281 (1795). дои : 10.1098/rspb.2014.0878 . ПМК 4213608 . ПМИД 25297859 .
- ^ Смит, Тимоти М.; Йорк, Пол Х.; Макриди, Питер I.; Кио, Майкл Дж.; Росс, Д. Джефф; Шерман, Крейг Д.Х. (2016). «Пространственные вариации репродуктивных усилий морской травы южной Австралии». Морские экологические исследования . 120 : 214–224. Бибкод : 2016MarER.120..214S . дои : 10.1016/j.marenvres.2016.08.010 . ПМИД 27592387 .
- ^ Перейти обратно: а б с Дж. Инглис, Грэм (1999). «Изменение в пополнении семян морских водорослей: последствия для динамики популяции и управления ресурсами». Тихоокеанская природоохранная биология . 5 (4): 251. дои : 10.1071/PC000251 .
- ^ Куо, Джон; Иидзуми, Хитоши; Нильсен, Бьорг Э.; Айой, Кейко (1990). «Анатомия плодов, прорастание семян и развитие рассады японской морской травы Phyllospadix (Zosteraceae)». Водная ботаника . 37 (3): 229–245. Бибкод : 1990AqBot..37..229K . дои : 10.1016/0304-3770(90)90072-С .
- ^ Куо Дж., Ден Хартог К. (2006) «Морфология, анатомия и ультраструктура водорослей». В: Ларкум AWD, Орт Р.Дж., Дуарте К.М. (редакторы), Морские травы: биология, экология и охрана , Springer, страницы 51–87.
- ^ Фонсека, Марк С.; Кенворти, В. Джадсон (1987). «Влияние тока на фотосинтез и распространение морских трав». Водная ботаника . 27 (1): 59–78. Бибкод : 1987AqBot..27...59F . дои : 10.1016/0304-3770(87)90086-6 .
- ^ Перейти обратно: а б Арно-Хаон, Софи; Дуарте, Карлос М.; Диас-Алмела, Елена; Марба, Нурия; Синтес, Томас; Серран, Эстер А. (2012). «Последствия чрезвычайной продолжительности жизни клональных организмов: тысячелетние клоны на лугах находящихся под угрозой исчезновения морских водорослей Posidonia Oceanica» . ПЛОС ОДИН . 7 (2): e30454. Бибкод : 2012PLoSO...730454A . дои : 10.1371/journal.pone.0030454 . ПМК 3270012 . ПМИД 22312426 .
- ^ Вакки, Маттео; Де Фалько, Джованни; Симеоне, Симона; Монтефальконе, Моника; Морри, Карла; Феррари, Марко; Бьянки, Карло Найк (2017). «Биогеоморфология средиземноморских посидоний океанических травяных лугов» . Процессы на поверхности Земли и формы рельефа . 42 (1): 42–54. Бибкод : 2017ESPL...42...42В . дои : 10.1002/особенно 3932 . S2CID 130872337 .
- ^ Перейти обратно: а б Трипати, Дургеш К.; Мишра, Рохит К.; Сингх, Свати; Сингх, Самикша; Вишвакарма, Канчан; Шарма, Шивеш; Сингх, Виджай П.; Сингх, Прашант К.; Прасад, Шео М.; Дубей, Наваль К.; Панди, Авинаш К.; Сахи, Шивендра; Чаухан, Девендра К. (2017). «Оксид азота снижает фитотоксичность наночастиц оксида цинка в проростках пшеницы: влияние цикла аскорбат-глутатион» . Границы в науке о растениях . 8 : 1. дои : 10.3389/fpls.2017.00001 . ПМК 5292406 . ПМИД 28220127 .
- ^ Монтефальконе, М.; Джованнетти, Э.; Морри, К.; Пейрано, А.; Бьянки, Китай (2013). «Цветение морской травы Posidonia Oceanica на северо-западе Средиземноморья: есть ли связь с солнечной активностью?» . Средиземноморская морская наука . 14 (2): 416. дои : 10.12681/ммс.529 . S2CID 85362624 .
- ^ Руис, Дж. М.; Марин-Гуйрао, Л.; Гарсиа-Муньос, Р.; Рамос-Сегура, А.; Бернардо-Эстеллер, Дж.; Перес, М.; Санмарти, Н.; Онтория, Ю.; Ромеро, Дж.; Артур, Р.; Альковеро, Т.; Прокаччини, Г. (2018). «Экспериментальные доказательства вызванного потеплением цветения средиземноморской морской травы Posidonia Oceanica». Бюллетень о загрязнении морской среды . 134 : 49–54. Бибкод : 2018МартПБ.134...49Р . дои : 10.1016/j.marpolbul.2017.10.037 . hdl : 10508/15205 . ПМИД 29102072 . S2CID 11523686 .
- ^ Диас-Алмела, Елена; Марба, Нурия; Дуарте, Карлос М. (2007). «Последствия потепления в Средиземном море для записей цветения морской травы (Posidonia Oceanica)». Биология глобальных изменений . 13 (1): 224–235. Бибкод : 2007GCBio..13..224D . дои : 10.1111/j.1365-2486.2006.01260.x . S2CID 84055440 .
- ^ Балестри, Э.; Гобер, С.; Лепойнт, Г.; Лардиччи, К. (2009). «Содержание питательных веществ в семенах и пищевой статус сеянцев Posidonia Oceanica в северо-западной части Средиземного моря» . Серия «Прогресс в области морской экологии» . 388 : 99–109. Бибкод : 2009MEPS..388...99B . дои : 10.3354/meps08104 . hdl : 11568/132307 .
- ^ Селдран, Дэвид; Марин, Арнальдо (2011). «Фотосинтетическая активность неспящих семян Posidonia Oceanica». Морская биология . 158 (4): 853–858. Бибкод : 2011МарБи.158..853C . дои : 10.1007/s00227-010-1612-4 . S2CID 84357626 .
- ^ Селдран, Дэвид; Марин, Арнальдо (2013). «Фотосинтез семян усиливает рост проростков Posidonia Oceanica» . Экосфера . 4 (12): ст.149. дои : 10.1890/ES13-00104.1 .
- ^ Балестри, Елена; Де Баттисти, Давиде; Валлерини, Флавия; Лардиччи, Клаудио (2015). «Первые свидетельства морфологических и архитектурных изменений корней у молодых растений Posidonia Oceanica, колонизирующих различные типологии субстрата». Устьевые, прибрежные и шельфовые науки . 154 : 205–213. Бибкод : 2015ECSS..154..205B . дои : 10.1016/j.ecss.2015.01.002 . hdl : 11568/755031 .
- ^ Герреро-Месегер, Лаура; Санс-Ласаро, Карлос; Сук-Уенг, Криттавит; Марин, Арнальдо (2017). «Влияние субстрата и захоронения на развитие Posidonia Oceanica: Значение для восстановления». Реставрационная экология . 25 (3): 453–458. Бибкод : 2017ResEc..25..453G . дои : 10.1111/рек.12438 . hdl : 10045/66474 . S2CID 88876962 .
- ^ Бадаламенти, Фабио; Аланья, Адриана; Фичи, Сильвио (2015). «Свидетельства адаптивных свойств к каменистым субстратам подрывают парадигму предпочтения среды обитания средиземноморской морской травы Posidonia Oceanica» . Научные отчеты . 5 : 8804. Бибкод : 2015NatSR...5E8804B . дои : 10.1038/srep08804 . ПМК 4350093 . ПМИД 25740176 .
- ^ Аланья, Адриана; Фернандес, Томас Вега; Терлицци, Антонио; Бадаламенти, Фабио (2013). «Влияние микросреды обитания на выживаемость и рост рассады средиземноморской морской травы posidonia Oceanica (L.) Delile». Устьевые, прибрежные и шельфовые науки . 119 : 119–125. Бибкод : 2013ECSS..119..119A . дои : 10.1016/j.ecss.2013.01.009 .
- ^ Пак, Санг Рул; Ким, Сангил; Ким, Ён Гюн; Канг, Чанг-Гын; Ли, Кун Соп (2016). «Фотоакклиматические реакции Zostera marina в приливной и сублиторальной зонах» . ПЛОС ОДИН . 11 (5): e0156214. Бибкод : 2016PLoSO..1156214P . дои : 10.1371/journal.pone.0156214 . ПМК 4881947 . ПМИД 27227327 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Сильва, Дж.; Сантос, Р. (2003). «Суточные изменения фотосинтеза морских водорослей по вертикальному градиенту» . Серия «Прогресс в области морской экологии» . 257 : 37–44. Бибкод : 2003MEPS..257...37S . дои : 10.3354/meps257037 . hdl : 10400.1/12140 .
- ^ Перейти обратно: а б Бозе, Брюс Л.; Роббинс, Брэдли Д.; Терсби, Глен (2005). «Высыхание является ограничивающим фактором для распространения взморника (Zostera marina L.) в приливной зоне устья северо-восточной части Тихого океана (США)». Ботаника Марина . 48 (4). дои : 10.1515/БОТ.2005.037 . S2CID 85105171 .
- ^ Пирсон, Райан М.; Коллиер, Кэтрин Дж.; Браун, Кристофер Дж.; Рашид, Майкл А.; Бурнер, Джессика; Тершвелл, Миша П.; Сиверс, Майкл; Коннолли, Род М. (15 августа 2021 г.). «Удаленная оценка водной световой среды с использованием машинного обучения: новый инструмент управления подводной водной растительностью» . Наука об общей окружающей среде . 782 : 146886. Бибкод : 2021ScTEn.78246886P . doi : 10.1016/j.scitotenv.2021.146886 . hdl : 10072/403729 . ISSN 0048-9697 . S2CID 233519731 .
- ^ Дурако, MJ; Кунцельман, Дж.И.; Кенворти, WJ; Хаммерстрем, К.К. (2003). «Глубинная изменчивость фотобиологии двух популяций Halophila johnsonii и Halophila decipiens». Морская биология . 142 (6): 1219–1228. Бибкод : 2003MarBi.142.1219D . дои : 10.1007/s00227-003-1038-3 . S2CID 85627116 .
- ^ Перейти обратно: а б Оливе, И.; Вергара, Джей Джей; Перес-Льоренс, ХЛ (2013). «Фотосинтетическая и морфологическая фотоакклиматизация морской травы Cymodocea nodosa к сезону, глубине и положению листьев». Морская биология . 160 (2): 285–297. Бибкод : 2013МарБи.160..285О . дои : 10.1007/s00227-012-2087-2 . S2CID 86386210 .
- ^ Hemminga MA и Durate CM (2000) Экология морских водорослей . Издательство Кембриджского университета.
- ^ Перейти обратно: а б Седдон, С.; Чешир, AC (2001). «Фотосинтетическая реакция Amphibolis antarctica и Posidonia australis на температуру и высыхание с использованием флуоресценции хлорофилла» . Серия «Прогресс в области морской экологии» . 220 : 119–130. Бибкод : 2001MEPS..220..119S . дои : 10.3354/meps220119 . hdl : 2440/12249 .
- ^ Херст А., Болл Д., Хейслерс С., Янг П., Блейк С., Кутс А. Программа мониторинга морских водорослей в районе залива, Промежуточный отчет № 2 (2008). Технический отчет Fisheries Victoria № 29, январь 2009 г.
- ^ Кох, Эвамария В. (2001). «За пределами света: физические, геологические и геохимические параметры как возможные требования к среде обитания подводной водной растительности». Эстуарии . 24 (1): 1–17. дои : 10.2307/1352808 . JSTOR 1352808 . S2CID 85287808 .
- ^ Танака, Ю.; Накаока, М. (2004). «Эмерджентный стресс и морфологические ограничения влияют на распределение видов и рост субтропических приливных морских трав» . Серия «Прогресс в области морской экологии» . 284 : 117–131. Бибкод : 2004MEPS..284..117T . дои : 10.3354/meps284117 .
- ^ Бьорк, М.; Уку, Дж.; Вейль, А.; Бир, С. (1999). «Фотосинтетическая устойчивость тропических приливных морских трав к высыханию» . Серия «Прогресс в области морской экологии» . 191 : 121–126. Бибкод : 1999MEPS..191..121B . дои : 10.3354/meps191121 .
- ^ Петру, К.; Хименес-Деннесс, И.; Шартран, К.; МакКормак, К.; Рашид, М.; Ральф, Пи Джей (2013). «Сезонная гетерогенность фотофизиологической реакции на воздействие воздуха у двух видов тропических приливных водорослей» (PDF) . Серия «Прогресс в области морской экологии» . 482 : 93–106. Бибкод : 2013MEPS..482...93P . дои : 10.3354/meps10229 . hdl : 10453/23914 .
- ^ Сюй, Шаочунь; Чжоу, И; Ван, Пэнмей; Ван, Фэн; Чжан, Сяомэй; Гу, Руитинг (2016). «Соленость и температура существенно влияют на прорастание семян, приживаемость и рост рассады зостеры марины » . ПерДж . 4 : е2697. дои : 10.7717/peerj.2697 . ПМК 5119234 . ПМИД 27896031 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Шварц, А.-М.; Бьорк, М.; Булуда, Т.; Мтолера, М.; Бир, С. (2000). «Фотосинтетическое использование углерода и света двумя тропическими видами морских водорослей, измеренное in situ». Морская биология . 137 (5–6): 755–761. Бибкод : 2000МарБи.137..755С . дои : 10.1007/s002270000433 . S2CID 86384408 .
- ^ Кэмпбелл, Стюарт Дж.; Маккензи, Лен Дж.; Кервилл, Саймон П.; Бите, Хуанита С. (2007). «Закономерности фотосинтеза тропических морских водорослей в зависимости от света, глубины и среды обитания». Устьевые, прибрежные и шельфовые науки . 73 (3–4): 551–562. Бибкод : 2007ECSS...73..551C . дои : 10.1016/j.ecss.2007.02.014 .
- ^ Ли, Кун Соп; Дантон, Кеннет Х. (1997). «Влияние снижения освещенности in situ на поддержание, рост и распределение углеродных ресурсов в банках Thalassia testudinum ex König». Журнал экспериментальной морской биологии и экологии . 210 (1): 53–73. Бибкод : 1997JEMBE.210...53L . дои : 10.1016/S0022-0981(96)02720-7 .
- ^ Лонгстафф, Би Джей; Деннисон, WC (1999). «Выживание морских трав во время импульсной мутности: влияние недостатка света на морские травы Halodule pinifolia и Halophila ovalis». Водная ботаника . 65 (1–4): 105–121. Бибкод : 1999AqBot..65..105L . дои : 10.1016/S0304-3770(99)00035-2 .
- ^ Коллиер, CJ; Лавери, PS; Ральф, Пи Джей; Масини, Р.Дж. (2008). «Физиологические характеристики морской травы Posidonia sinuosa в зависимости от глубины градиента доступности света» . Серия «Прогресс в области морской экологии» . 353 : 65–79. Бибкод : 2008MEPS..353...65C . дои : 10.3354/meps07171 . hdl : 10453/8694 .
- ^ Ли, Кун Соп; Пак, Санг Рул; Ким, Ён Кюн (2007). «Влияние освещенности, температуры и питательных веществ на динамику роста морских трав: обзор». Журнал экспериментальной морской биологии и экологии . 350 (1–2): 144–175. Бибкод : 2007JEMBE.350..144L . дои : 10.1016/j.jembe.2007.06.016 .
- ^ Наяр, С.; Коллингс, Дж.Дж.; Миллер, диджей; Брайарс, С.; Чешир, AC (2009). «Поглощение и распределение ресурсов неорганического углерода морскими травами умеренного пояса Posidonia и Amphibolis». Журнал экспериментальной морской биологии и экологии . 373 (2): 87–95. Бибкод : 2009JEMBE.373...87N . дои : 10.1016/j.jembe.2009.03.010 .
- ^ Пиво, Свен (1989). «Фотосинтез и фотодыхание морских покрытосеменных». Водная ботаника . 34 (1–3): 153–166. Бибкод : 1989AqBot..34..153B . дои : 10.1016/0304-3770(89)90054-5 .
- ^ Ларкум AWD, Джеймс П.Л. К модели поглощения неорганического углерода морскими травами с участием карбоангидразы. Куо Дж., Филлипс Р.К., Уокер Д.И., Киркман Х., редакторы. Биология морских водорослей: материалы международного семинара. Недлендс: Университет Западной Австралии; 1996. стр. 191–196.
- ^ Пиво, Свен; Ренберг, Джон (1997). «Приобретение неорганического углерода морской травой Zostera marina». Водная ботаника . 56 (3–4): 277–283. Бибкод : 1997AqBot..56..277B . дои : 10.1016/S0304-3770(96)01109-6 .
- ^ Силва, Жуан; Сантос, Руи; Каллеха, Мария Лл.; Дуарте, Карлос М. (2005). «Продуктивность макрофитов в приливной зоне по сравнению с воздействием воздуха: от физиологических оценок до оценок на уровне сообщества». Журнал экспериментальной морской биологии и экологии . 317 (1): 87–95. Бибкод : 2005JEMBE.317...87S . дои : 10.1016/j.jembe.2004.11.010 .
- ^ О'Лири, Мэрион Х. (1988). «Изотопы углерода в фотосинтезе». Бионаука . 38 (5): 328–336. дои : 10.2307/1310735 . JSTOR 1310735 .
- ^ Рэйвен, Джон А.; Джонстон, Эндрю М.; Кюблер, Джанет Э.; Корб, Ребекка; Макинрой, Шона Г.; Хэндли, Линда Л.; Скримджер, Чарли М.; Уокер, Диана И.; Бердалл, Джон; Вандерклифт, Мэтью; Фредриксен, Штейн; Дантон, Кеннет Х. (2002). «Механистическая интерпретация дискриминации изотопов углерода морскими макроводорослями и морскими травами». Функциональная биология растений . 29 (3): 355–378. дои : 10.1071/PP01201 . ПМИД 32689482 .
- ^ Seagrass-Watch: Что такое морская трава? Проверено 16 ноября 2012 г.
- ^ Нордлунд, Лина; Кох, Евамария В.; Барбье, Эдвард Б.; Крид, Джоэл К. (12 октября 2016 г.). Рейнхарт, Курт О. (ред.). «Экосистемные услуги морских водорослей и их изменчивость в зависимости от рода и географических регионов» . ПЛОС ОДИН . 11 (10): e0163091. Бибкод : 2016PLoSO..1163091M . дои : 10.1371/journal.pone.0163091 . ISSN 1932-6203 . ПМК 5061329 . ПМИД 27732600 .
- ^ Программа ООН по окружающей среде (2020). Совершенно неожиданно: ценность морских трав для окружающей среды и людей. ЮНЕП, Найроби. https://www.unenvironment.org/resources/report/out-blue-value-seagrasses-environment-and-people
- ^ Перейти обратно: а б Джонс, Клайв Г.; Лоутон, Джон Х.; Шачак, Моше (1994). «Организмы как инженеры экосистем». Ойкос . 69 (3): 373–386. Бибкод : 1994Oikos..69..373J . дои : 10.2307/3545850 . JSTOR 3545850 .
- ^ Грей, Уильям; Моффлер, Марк (1987). «Цветение морской травы Thalassia testudinum (Hydrocharitacea) в районе залива Тампа, Флорида». Водная ботаника . 5 : 251–259. дои : 10.1016/0304-3770(78)90068-2 .
- ^ Дарнелл, Келли; Дантон, Кеннет (2016). «Репродуктивная фенология субтропических морских трав Thalassia testudinum (черепашья трава) и Halodule wrightii (мелкая трава) в северо-западной части Мексиканского залива». Ботаника Марина . 59 (6): 473–483. дои : 10.1515/bot-2016-0080 . S2CID 88685282 .
- ^ Макриди, ИП; Бэрд, Мэн; Треватан-Тэкетт, С.М.; Ларкум, полный привод; Ральф, Пи Джей (2013). «Количественная оценка и моделирование способности лугов с морскими водорослями улавливать углерод». Бюллетень о загрязнении морской среды . 83 (2): 430–439. doi : 10.1016/j.marpolbul.2013.07.038 . ПМИД 23948090 .
- ^ «Часто задаваемые вопросы о морских водорослях» . Комиссия Флориды по охране рыбы и дикой природы .
- ^ Нагелькеркен, И.; Робертс, CM; ван дер Вельде, Г.; Доренбош, М.; ван Риель, MC; Кошере де ла Мориньер, Э.; Ниенхейс, PH (2002). «Насколько важны мангровые заросли и заросли морских водорослей для коралловых рифов? Гипотеза о питомнике проверена в масштабе острова» . Серия «Прогресс в области морской экологии» . 244 : 299–305. Бибкод : 2002MEPS..244..299N . дои : 10.3354/meps244299 . hdl : 2066/126674 .
- ^ Нордлунд, LM; Ансворт, РКФ; Галлстрем, М.; Каллен-Ансворт, LC (2018). «Глобальное значение промысла морских водорослей» . Рыба и рыболовство . 19 (3): 399–412. Бибкод : 2018AqFF...19..399N . дои : 10.1111/faf.12259 .
- ^ Ансворт, РКФ; Нордлунд, LM; Каллен-Ансворт, LC (2019). «Луга из морских водорослей поддерживают глобальное рыболовство» . Сохрани Летт . е12566 (1): е12566. Бибкод : 2019ConL...12E2566U . дои : 10.1111/conl.12566 .
- ^ Перейти обратно: а б Бучек, RE; Леоне, Э.; Бикфорд, Дж.; Уолтерс-Бернсед, С.; Лоуэр-Барбьери, С. (2017). «Больше, чем просто место нереста: изучение мелкомасштабного космического использования двух эстуарных видов рыб в месте нереста». Границы морской науки (4): 1–9.
- ^ Перейти обратно: а б Угарелли К., Чакрабарти С., Лаас П. и Стингл У. (2017) «Голобионт морской травы и его микробиом». Микроорганизмы , 5 (4): 81. doi : 10.3390/microorganisms5040081 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Тарквинио Ф., Хайндс Г.А., Лаверок Б., Коендерс А. и Савстрем К. (2019) «Голобионт морских водорослей: понимание взаимодействия водорослей и бактерий и их роли в функционировании экосистемы морских водорослей». Письма FEMS по микробиологии , 366 (6): fnz057. два : 10.1093/femsle/fnz057 .
- ^ Маргулис, Линн (1991) «Симбиогенез и симбионтизм» . В: Симбиоз как источник эволюционных инноваций ; Маргулис Л., Фестер Р. (ред.), Cambridge MIT Press. ISBN 9780262132695 .
- ^ Дуглас, AE; Веррен, Дж. Х. (2016) «Дыры в гологеноме: почему симбиозы хозяина и микроба не являются холобионтами». mBio , 7 : e02099-15. дои : 10.1128/mBio.02099-15 .
- ^ Тайс, КР; Дейли, Нью-Мексико; Классен, Дж.Л.; Брукер, Р.М.; Бейнс, Дж. Ф.; Бош, ТКГ; Крайан, Дж. Ф.; Гилберт, Сан-Франциско; Спокойной ночи, СиДжей; Ллойд, Э.А.; и др. Правильное понимание концепции гологенома: эко-эволюционная основа хозяев и их микробиомов. mSystems 2016, 1, e00028-16. дои : 10.1128/mSystems.00028-16 .
- ^ Розенберг, Э. и Зильбер-Розенберг, И. (2016) «Микробы стимулируют эволюцию животных и растений: концепция гологенома». МБио , 7 (2). дои : 10.1128/mBio.01395-15 .
- ^ Зильбер-Розенберг И. и Розенберг Э. (2008) «Роль микроорганизмов в эволюции животных и растений: гологеномная теория эволюции». Обзоры микробиологии FEMS , 32 (5): 723–735. дои : 10.1111/j.1574-6976.2008.00123.x .
- ^ Ванденкокорнхейз П., Квайзер А., Дюамель М., Ле Ван А. и Дюфрен А. (2015) «Важность микробиома холобионта растений». Новый фитолог , 206 (4): 1196-1206. дои : 10.1111/nph.13312 .
- ^ Санчес-Канисарес, К., Хоррин, Б., Пул, П.С. и Ткач, А. (2017) «Понимание холобионта: взаимозависимость растений и их микробиома». Текущее мнение по микробиологии , 38 : 188–196. дои : 10.1016/j.mib.2017.07.001 .
- ^ Согин, Э. Мэгги; Мишель, Долма; Грубер-Водика, Харальд Р.; Бурсо, Патрик; Гейер, Бенедикт; Мейер, Дмитрий В.; Зейдель, Майкл; Ахмеркамп, Серен; Шорн, Сина; Д'Анджело, Грейс; Прокаччини, Габриэле (2 мая 2022 г.). «Сахар доминирует в ризосфере морских водорослей» . Экология и эволюция природы . 6 (7): 866–877. Бибкод : 2022NatEE...6..866S . дои : 10.1038/s41559-022-01740-z . ISSN 2397-334X . ПМЦ 9262712 . ПМИД 35501482 .
- ^ Сайед, Нурул Фарахин Нур; Закария, Мута Хара; Буджанг, Джапар Сидик (2016). «Характеристики волокон и изготовление бумаги из морских водорослей с использованием взбитой вручную и смешанной целлюлозы» . Биоресурсы . 11 (2). дои : 10.15376/biores.11.2.5358-5380 .
- ^ Перейти обратно: а б Акино, РС; Ландейра-Фернандес, AM; Валенте, АП; Андраде, ЛР; Мурао, Пенсильвания (2004). «Появление сульфатированных галактанов у морских покрытосеменных: эволюционные последствия» . Гликобиология . 15 (1): 11–20. дои : 10.1093/гликоб/cwh138 . ПМИД 15317737 .
- ^ Сильва, Джулиана MC; Дантас-Сантос, Неднальдо; Гомес, Даянн Л.; Коста, Леандро С.; Кордейро, Сара Л.; Коста, Марианский SSP; Сильва, Наисандра Б.; Фрейтас, Мария Л.; Скортеччи, Катя Кастаньо; Лейте, Эдда Л.; Роча, Хьюго АО (2012). «Биологическая активность сульфатированного полисахарида сосудистого растения Halodule wrightii» . Бразильский журнал фармакогнозии . 22 : 94–101. дои : 10.1590/S0102-695X2011005000199 .
- ^ Глоаген, Винсент; Брудье, Вероник; Клосс, Бриджит; Барбат, Алин; Крауш, Пьер; Сент-Катрин, Одиль; Кремер, Мишель; Мэйс, Эммануэль; Герардель, Янн (2010). «Структурная характеристика и цитотоксические свойства богатого апиозой пектинового полисахарида, полученного из клеточной стенки морского Phanerogam Zostera marina». Журнал натуральных продуктов . 73 (6): 1087–1092. дои : 10.1021/np100092c . ПМИД 20465284 .
- ^ Лев, Юцзин; Шан, Зинди; Чжао, Ся; Цай, Чао; Чжао, Сяолян; Ланг, Иньчжи; Чжу, Хэ; Ю, Гуанли (2015). «Экстракция, выделение, структурная характеристика и противоопухолевые свойства богатого апигалактуронаном полисахарида из морской травы Zostera caespitosa Miki» . Морские наркотики . 13 (6): 3710–3731. дои : 10.3390/md13063710 . ПМЦ 4483652 . ПМИД 26110894 .
- ^ Джонсон, Ким Л.; Джонс, Брайан Дж.; Бачич, Антоний; Шульц, Кэролайн Дж. (2003). «Фаскиклин-подобные арабиногалактановые белки арабидопсиса. Мультигенное семейство предполагаемых молекул клеточной адгезии» . Физиология растений . 133 (4): 1911–1925. дои : 10.1104/стр.103.031237 . ПМК 300743 . ПМИД 14645732 .
- ^ Эллис, Мириам; Эгелунд, Джек; Шульц, Кэролайн Дж.; Бачич, Энтони (2010). «Арабиногалактан-белки: ключевые регуляторы на поверхности клетки?» . Физиология растений . 153 (2): 403–419. дои : 10.1104/стр.110.156000 . ПМЦ 2879789 . ПМИД 20388666 .
- ^ Перейти обратно: а б с Ма, Инсюань; Цзэн, Вэй; Бачич, Антоний; Джонсон, Ким (2018). «AGP сквозь время и пространство». Ежегодные обзоры растений онлайн . стр. 767–804. дои : 10.1002/9781119312994.apr0608 . ISBN 9781119312994 . S2CID 104384164 .
- ^ Классен, Биргит; Бауманн, Александр; Утермолен, Джон (2019). «Арабиногалактан-белки в споровых наземных растениях». Углеводные полимеры . 210 : 215–224. дои : 10.1016/j.carbpol.2019.01.077 . ПМИД 30732757 . S2CID 73426733 .
- ^ Пфайфер, Лукас; Шафи, Томас; Джонсон, Ким Л.; Бачич, Антоний; Классен, Биргит (2020). «Арабиногалактан-белки Zostera marina L. Содержат уникальные гликановые структуры и дают представление о процессах адаптации к соленой среде» . Научные отчеты . 10 (1): 8232. Бибкод : 2020NatSR..10.8232P . дои : 10.1038/s41598-020-65135-5 . ПМЦ 7237498 . ПМИД 32427862 .
- ^ Лэмпорт, Дерек Т.А.; Келишевский, Марсия Дж.; Шоуолтер, Аллан М. (2006). «Солевой стресс усиливает периплазматические белки арабиногалактана: использование солевого стресса для анализа функции AGP». Новый фитолог . 169 (3): 479–492. дои : 10.1111/j.1469-8137.2005.01591.x . ПМИД 16411951 .
- ^ Опсал, С.; Беннер, Р. (1993). «Разложение стареющих стеблей морской травы Halodule wrightii в субтропической лагуне» . Серия «Прогресс в области морской экологии» . 94 : 191–205. Бибкод : 1993MEPS...94..191O . дои : 10.3354/meps094191 .
- ^ Клап, Вирджиния; Хемминга, Массачусетс; Бун, Джей-Джей (2000). «Удержание лигнина в морских травах: покрытосеменные растения, вернувшиеся в море» . Серия «Прогресс в области морской экологии» . 194 : 1–11. Бибкод : 2000MEPS..194....1K . дои : 10.3354/meps194001 .
- ^ Мартоне, Патрик Т.; Эстевес, Хосе М.; Лу, Фачуанг; Руэль, Катя; Денни, Марк В.; Сомервилл, Крис; Ральф, Джон (2009). «Открытие лигнина в морских водорослях свидетельствует о конвергентной эволюции архитектуры клеточной стенки» . Современная биология . 19 (2): 169–175. Бибкод : 2009CBio...19..169M . дои : 10.1016/j.cub.2008.12.031 . ПМИД 19167225 . S2CID 17409200 .
- ^ Каал, Джоэри; Серрано, Оскар; Дель Рио, Хосе К.; Ренкорет, Хорхе (2018). «Радикально отличающийся состав лигнина у видов Posidonia может быть связан с различиями в способности связывать органический углерод» . Органическая геохимия . 124 : 247–256. Бибкод : 2018OrGeo.124..247K . doi : 10.1016/j.orggeochem.2018.07.017 . hdl : 10261/169361 . S2CID 105104424 .
- ^ Перейти обратно: а б Дуарте, Карлос М. (июнь 2002 г.). «Будущее лугов с водорослями» . Охрана окружающей среды . 29 (2): 192–206. Бибкод : 2002EnvCo..29..192D . дои : 10.1017/S0376892902000127 . hdl : 10261/89840 . ISSN 1469-4387 . S2CID 31971900 .
- ^ Уэйкотт, Мишель; Дуарте, Карлос М.; Каррутерс, Тим Дж.Б.; Орт, Роберт Дж.; Деннисон, Уильям К.; Олярник, Сюзанна; Калладайн, Эйнсли; Фуркурин, Джеймс В.; Черт возьми, Кеннет Л.; Хьюз, А. Рэндалл; Кендрик, Гэри А. (28 июля 2009 г.). «Ускорение исчезновения морских трав по всему миру угрожает прибрежным экосистемам» . Труды Национальной академии наук . 106 (30): 12377–12381. Бибкод : 2009PNAS..10612377W . дои : 10.1073/pnas.0905620106 . ISSN 0027-8424 . ПМК 2707273 . ПМИД 19587236 .
- ^ Перейти обратно: а б Шорт, Фредерик Т.; Полидоро, Бет; Ливингстон, Сюзанна Р.; Карпентер, Кент Э.; Бандейра, Саломан; Буджанг, Джапар Сидик; Калумпонг, Хилконида П.; Каррутерс, Тим Дж.Б.; Коулз, Роберт Г.; Деннисон, Уильям К.; Эрфтемейер, Пол Л.А. (1 июля 2011 г.). «Оценка риска исчезновения видов морских водорослей в мире» . Биологическая консервация . 144 (7): 1961–1971. Бибкод : 2011BCons.144.1961S . дои : 10.1016/j.biocon.2011.04.010 . ISSN 0006-3207 . S2CID 32533417 .
- ^ Хевел, Майкл Р.; Хичкок, Джесси К.; Гроб, Майкл Р.С.; Патер, Кристина С.; Куртенэ, Саймон К. (8 мая 2019 г.). «Неорганический азот оказывает доминирующее влияние на распространение эстуарного взморника в южном заливе Св. Лаврентия, Канада» . Лимнология и океанография . 64 (6): 2313–2327. Бибкод : 2019LimOc..64.2313H . дои : 10.1002/lno.11185 . ISSN 0024-3590 .
- ^ Хемминга, Мартен А.; Дуарте, Карлос М. (2000). Экология морских водорослей . Кембридж: Издательство Кембриджского университета. дои : 10.1017/cbo9780511525551 . ISBN 978-0-521-66184-3 .
- ^ Перейти обратно: а б Орт, Роберт Дж.; Каррутерс, Тим Дж.Б.; Деннисон, Уильям К.; Дуарте, Карлос М.; Фуркурин, Джеймс В.; Черт возьми, Кеннет Л.; Хьюз, А. Рэндалл; Кендрик, Гэри А.; Кенворти, В. Джадсон; Олярник, Сюзанна; Шорт, Фредерик Т. (1 декабря 2006 г.). «Глобальный кризис экосистем морских водорослей» . Бионаука . 56 (12): 987–996. doi : 10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2 . ISSN 0006-3568 . S2CID 4936412 .
- ^ Перейти обратно: а б Беркхолдер, Джоанн М.; Томаско, Дэвид А.; Тушетт, Брант В. (9 ноября 2007 г.). «Морские травы и эвтрофикация» . Журнал экспериментальной морской биологии и экологии . Биология и экология морских трав. 350 (1): 46–72. Бибкод : 2007JEMBE.350...46B . дои : 10.1016/j.jembe.2007.06.024 . ISSN 0022-0981 .
- ^ Телеска, Лука; Беллускио, Андреа; Крисколи, Алессандро; Ардиццоне, Джандоменико; Апостолаки Евгения Т.; Фраскетти, Симонетта; Гристина, Мишель; Книтвайс, Лейла; Мартин, Корин С.; Пержан, Жерар; Аланья, Адриана (28 июля 2015 г.). «Распространение и траектории изменений лугов с морскими травами (Posidonia Oceanica)» . Научные отчеты . 5 (1): 12505. Бибкод : 2015NatSR...512505T . дои : 10.1038/srep12505 . ISSN 2045-2322 . ПМК 4516961 . ПМИД 26216526 .
- ^ Ансворт, Ричард К.Ф.; Маккензи, Лен Дж.; Коллиер, Кэтрин Дж.; Каллен-Ансворт, Линн К.; Дуарте, Карлос М.; Эклёф, Йохан С.; Джарвис, Джесси С.; Джонс, Бенджамин Л.; Нордлунд, Лина М. (1 августа 2019 г.). «Глобальные проблемы сохранения морских водорослей» . Амбио . 48 (8): 801–815. Бибкод : 2019Амбио..48..801U . дои : 10.1007/s13280-018-1115-y . ISSN 1654-7209 . ПМК 6541581 . ПМИД 30456457 .
- ^ Ансворт, Ричард К.Ф.; Каллен-Ансворт, Линн К.; Джонс, Бенджамин Л.; Лилли, Ричард Дж. (5 августа 2022 г.). «Планетарная роль сохранения морских водорослей» . Наука . 377 (6606): 609–613. Бибкод : 2022Sci...377..609U . дои : 10.1126/science.abq6923 . ПМИД 35926055 . S2CID 251347987 .
- ^ Сюй, Шаочунь; Цяо, Юнлян; Сюй, Шуай; Юэ, Шидун; Чжан, Ю; Лю, Минцзе; Чжан, Сяомэй; Чжоу, И (1 июня 2021 г.). «Разнообразие, распространение и сохранение морских водорослей в прибрежных водах Ляодунского полуострова, северная часть Желтого моря, северный Китай: значение для сохранения морских водорослей» . Бюллетень о загрязнении морской среды . 167 : 112261. Бибкод : 2021MarPB.16712261X . doi : 10.1016/j.marpolbul.2021.112261 . ISSN 0025-326X . ПМИД 33799145 . S2CID 232775373 .
- ^ Ньюмастер, А.Ф.; Берг, К.Дж.; Рагупати, С.; Паланисами, М.; Самбандан, К.; Ньюмастер, SG (23 ноября 2011 г.). «Местные знания и сохранение морских трав в штате Тамилнад, Индия» . Журнал этнобиологии и этномедицины . 7 (1): 37. дои : 10.1186/1746-4269-7-37 . ISSN 1746-4269 . ПМК 3269989 . ПМИД 22112297 .
- ^ Мохсин, Харун (24 июня 2022 г.). «Всемирный день морских водорослей» . Национальный сегодня .
- ^ «Всемирный день морских водорослей» . Всемирная ассоциация морских водорослей . 10 июня 2018 года . Проверено 14 июля 2022 г.
Дальнейшие ссылки
[ редактировать ]- ден Хартог, К. 1970. Морские травы мира . Трактат Королевской Нидерландской академии искусств и наук, кафедра. Физика , нет. 59(1).
- Дуарте, Карлос М. и Карина Л. Чискано «Биомасса и производство морских водорослей: переоценка», Водная ботаника, том 65, выпуски 1–4, ноябрь 1999 г., страницы 159–174.
- Грин, EP и Short, FT (ред.). 2003. Мировой атлас морских трав . Калифорнийский университет Press, Беркли, Калифорния. 298 стр.
- Хемминга, Массачусетс и Дуарте, К. 2000. Экология морских водорослей . Издательство Кембриджского университета, Кембридж. 298 стр.
- Хогарт, Питер Биология мангровых зарослей и морских трав (Oxford University Press, 2007)
- Ларкум, Энтони В.Д., Роберт Дж. Орт и Карлос М. Дуарте (редакторы) Морские травы: биология, экология и охрана (Springer, 2006)
- Орт, Роберт Дж. и др. «Глобальный кризис экосистем морских водорослей», BioScience , декабрь 2006 г. / Том. 56 № 12, страницы 987–996.
- Шорт, FT и Коулз, RG (ред.). 2001. Глобальные методы исследования морских водорослей . Elsevier Science, Амстердам. 473 стр.
- AWD Ларкум, Р. Дж. Орт и К. М. Дуарте (редакторы). Биология морских водорослей: трактат. CRC Press, Бока-Ратон, Флорида, в печати.
- А. Шварц; М. Моррисон; И. Хоуз; Дж. Холлидей. 2006. Физические и биологические характеристики редкой морской среды обитания: субприливные заросли морских водорослей прибрежных островов. Наука в защиту природы 269. 39 стр. [1]
- Уэйкотт М., МакМэхон К. и Лавери П. 2014, Путеводитель по морским травам южного умеренного пояса, CSIRO Publishing, Мельбурн.
Внешние ссылки
[ редактировать ]- Каллен-Ансуорт, Линн С .; Ансворт, Ричард (3 августа 2018 г.). «Призыв к защите морских водорослей» . Наука . 361 (6401): 446–448. Бибкод : 2018Sci...361..446C . дои : 10.1126/science.aat7318 . ISSN 0036-8075 . ПМИД 30072524 . S2CID 51908021 .
- Project Seagrass - Благотворительная организация, способствующая сохранению морских водорослей посредством сообщества, исследований и действий.
- Проект «Восстановление морских трав ».
- SeagrassSpotter — проект гражданской науки, направленный на повышение осведомленности о лугах с водорослями и картографирование их местонахождения.
- Обзор водорослей и зарослей морских водорослей на Смитсоновском океаническом портале
- Статья в журнале Nature Geoscience, описывающая расположение лугов с водорослями по всему миру.
- Seagrass-Watch — крупнейшая в мире научная программа неразрушающего анализа и мониторинга морских водорослей.
- Restore-A-Scar - некоммерческая кампания по восстановлению лугов с водорослями, поврежденных опорами лодок.
- SeagrassNet - глобальная программа мониторинга морских водорослей
- Фонд морских водорослей при Фонде океана
- Систематика морских трав
- Всемирная ассоциация морских водорослей
- Морская траваLI
- Наука и управление морскими водорослями в Южно-Китайском море и Сиамском заливе
- Морская экология (декабрь 2006 г.) - специальный выпуск о морских травах.
- Камбоджийские морские травы
- Производительность морских водорослей — действие COST ES0906
- Рыболовство в Западной Австралии – информационный бюллетень о морских водорослях

