Триасовый период
| Триасовый период | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Карта мира, какой она была в позднем триасе, ок. 220 млн лет назад [ нужна ссылка ] | |||||||||||||||||||||||
| Хронология | |||||||||||||||||||||||
| |||||||||||||||||||||||
| Этимология | |||||||||||||||||||||||
| Формальность имени | Формальный | ||||||||||||||||||||||
| Информация об использовании | |||||||||||||||||||||||
| Небесное тело | Земля | ||||||||||||||||||||||
| Региональное использование | Глобальный ( ICS ) | ||||||||||||||||||||||
| Используемая шкала времени | Временная шкала ICS | ||||||||||||||||||||||
| Определение | |||||||||||||||||||||||
| Хронологическая единица | Период | ||||||||||||||||||||||
| Стратиграфическая единица | Система | ||||||||||||||||||||||
| Формальность временного интервала | Формальный | ||||||||||||||||||||||
| Определение нижней границы | Первое появление конодонта Hindeodus parvus. | ||||||||||||||||||||||
| Нижняя граница ГССП | Мэйшань , Чжэцзян , Китай 31 ° 04'47 "N 119 ° 42'21" E / 31,0798 ° N 119,7058 ° E | ||||||||||||||||||||||
| Нижний GSSP ратифицирован | 2001 [6] | ||||||||||||||||||||||
| Определение верхней границы | Первое появление аммонита Psiloceras spelae tirolicum. | ||||||||||||||||||||||
| Верхняя граница ГССП | Участок Кухйох, горы Карвендель , Северные Известняковые Альпы , Австрия 47 ° 29'02 "N 11 ° 31'50" E / 47,4839 ° N 11,5306 ° E | ||||||||||||||||||||||
| Верхний GSSP ратифицирован | 2010 [7] | ||||||||||||||||||||||

Триас ɪ ( / t r aɪ ˈ æ s k ; / try- ASS -ik иногда обозначается 🝈 ) [8] — это геологический период и система , охватывающая 50,5 миллионов лет от конца пермского периода 251,902 миллиона лет назад ( млн лет назад ) до начала юрского периода 201,4 млн лет назад. [9] Триас — первый и самый короткий период мезозойской эры . И начало, и конец периода отмечены крупными событиями вымирания . [10] Триасовый период подразделяется на три эпохи: ранний триас , средний триас и поздний триас .
Триасовый период начался после пермско-триасового вымирания , в результате которого биосфера Земли обеднела; Прошла уже середина триаса, прежде чем жизнь восстановила свое прежнее разнообразие. В триасовой летописи можно выделить три категории организмов: выжившие после вымирания, новые группы, процветавшие на короткое время, и другие новые группы, которые продолжали доминировать в мезозойскую эру. Рептилии , особенно архозавры , были основными наземными позвоночными в это время. Специализированная группа архозавров, называемая динозаврами , впервые появилась в позднем триасе, но не стала доминирующей до следующего юрского периода. [11] Архозавры, ставшие доминирующими в этот период, были прежде всего псевдозухиями , родственниками и предками современных крокодилов , тогда как некоторые архозавры специализировались на полете, впервые среди позвоночных, став птерозаврами .
Терапсиды , доминирующие позвоночные животные предшествующего пермского периода, сократились на протяжении всего этого периода. Первые настоящие млекопитающие , которые сами являются специализированной подгруппой терапсидов, также появились в этот период. Обширный суперконтинент Пангея доминировал на земном шаре в триасовый период, но в последующий юрский период он начал постепенно раскалываться на два отдельных массива суши: Лавразию на севере и Гондвану на юге.
Глобальный климат в триасовом периоде был в основном жарким и сухим. [12] с пустынями, охватывающими большую часть внутренней части Пангеи. Однако климат изменился и стал более влажным, когда Пангея начала расходиться. Конец периода ознаменовался еще одним крупным массовым вымиранием, триасово-юрским вымиранием , которое уничтожило многие группы, в том числе большинство псевдозухий, и позволило динозаврам взять на себя доминирование в юрском периоде.
Этимология
[ редактировать ]Триас был назван в 1834 году Фридрихом Августом фон Альберти в честь последовательности трех отдельных слоев горных пород (по-гречески triás означает «триада»), которые широко распространены на юге Германии : нижний Бунцандштайн (красочный песчаник ) , средний Мушелькальк (ракушечник). известняк ) и верхний Кейпер (цветная глина ). [13]
 раннего триаса Песчаник (Бунцандштайн) недалеко от Штадтроды , Германия
раннего триаса Песчаник (Бунцандштайн) недалеко от Штадтроды , Германия Средний триас Muschelkalk (ракушечник) возле Дёрцбаха , Германия
Средний триас Muschelkalk (ракушечник) возле Дёрцбаха , Германия



Датировки и подразделения
[ редактировать ]В геологическом масштабе времени триас обычно делят на ранний , средний и поздний триас , а соответствующие породы называют нижним, средним или верхним триасом. Этапы фауны от самого молодого к самому старому:
| Серия/Эпоха | Фаунистический этап | Промежуток времени |
|---|---|---|
| Верхний/поздний триас (Tr3) | Ретийский | (208,5–201,4 ± 0,2 млн лет назад) |
| Нориан | (227–208,5 млн лет назад) | |
| Карнийский | (237–227 млн лет назад) | |
| Средний триас (Tr2) | ладинский | (242–237 млн лет назад) |
| Анисийский | (247,2–242 млн лет назад) | |
| Нижний/ранний триас (скифский) | Оленекян | (251,2–247,2 млн лет назад) |
| Индуец | (251,902 ± 0,024–251,2 млн лет назад) |
Палеогеография
[ редактировать ]
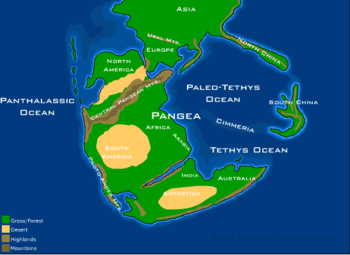
В триасовый период почти вся суша Земли была сосредоточена в едином суперконтиненте , Пангее ( букв. « Вся земля » ). [14] Этот суперконтинент был более или менее сосредоточен на экваторе и простирался между полюсами, хотя с течением времени он все же дрейфовал на север. Южная Пангея, также известная как Гондвана , была составлена из тесно прижатых кратонов, соответствующих современным Южной Америке , Африке , Мадагаскару , Индии , Антарктиде и Австралии . Северная Пангея, также известная как Лавруссия или Лавразия , соответствует современной Северной Америке и фрагментированным предшественникам Евразии . [ нужна ссылка ]
Западный край Пангеи лежал на краю огромного океана Панталасса (буквально «все море»), который примерно соответствует современному Тихому океану . Практически вся глубоководная океаническая кора, существовавшая в триасе, была переработана в результате субдукции океанических плит, поэтому об открытом океане этого периода времени известно очень мало. Большая часть информации о геологии и морской жизни Панталассана получена из островных дуг и редких отложений морского дна, аккумулированных на окружающих участках суши, таких как современная Япония и западная часть Северной Америки. [ нужна ссылка ]
Восточную окраину Пангеи занимала пара обширных океанических бассейнов: океаны Нео-Тетис (или просто Тетис) и Палео-Тетис . Они простирались от Китая до Иберии, и на мелководных тропических перифериях их обитала обильная морская жизнь. Они были отделены друг от друга длинной вереницей микроконтинентов, известных как Киммерийские террейны . Киммерийская кора отделилась от Гондваны в ранней перми и дрейфовала на север в триасе, увеличивая океан Нео-Тетис, образовавшийся в результате этого. В то же время они заставили океан Палео-Тетис сжаться, поскольку он погружался под Азию. К концу триаса океан Палео-Тетис занимал небольшую площадь, и Киммерийские террейны начали сталкиваться с Южной Азией. Это столкновение, известное как Киммерийская складчатость , продолжалось в юрский и меловой период, образуя цепь горных хребтов, простирающуюся от Турции до Малайзии . [15] [16]

Пангея была расколота обширными разломами и рифтовыми бассейнами в триасе, особенно в конце этого периода, но еще не отделилась. Первые неморские отложения в рифте , отмечающем начальный распад Пангеи, отделившей восточную часть Северной Америки от Марокко , относятся к позднему триасу; в Соединенных Штатах эти мощные отложения составляют супергруппу Ньюарка . [18] Рифтовые бассейны также распространены в Южной Америке, Европе и Африке. Наземная среда особенно хорошо представлена в Южной Африке, [19] Россия, Центральная Европа и юго-запад США. триаса Наземная биостратиграфия в основном основана на наземных и пресноводных четвероногих, а также на конхостраканах («моллюски-креветки»), типе быстроразмножающихся ракообразных, обитавших в озерах и гиперсоленой среде.
Поскольку береговая линия суперконтинента меньше, чем у ряда более мелких континентов, морские отложения триаса относительно редки в глобальном масштабе. Серьезным исключением является Западная Европа , где впервые был изучен триас. Северо-восточная окраина Гондваны представляла собой стабильную пассивную окраину вдоль океана Нео-Тетис, а морские отложения сохранились в некоторых частях северной Индии и Аравии . [16] В Северной Америке морские отложения ограничены несколькими обнажениями на западе.
Скандинавия
[ редактировать ]Считается, что в триасовый период пенеплены . на территории нынешней Норвегии и южной Швеции образовались [20] [21] [22] Остатки этого пенеплена можно проследить как наклонную вершину на западном побережье Швеции . [20] В северной Норвегии триасовые пенеплены, возможно, были погребены в отложениях, а затем вновь обнажились в виде прибрежных равнин, называемых береговыми равнинами . [21] Датирование иллитовой глины из береговой равнины Бёмло на юге Норвегии показало, что ландшафт там подвергся выветриванию во времена позднего триаса ( около 210 миллионов лет назад), и ландшафт, вероятно, также сформировался в это время. [23]
Палеоокеанография
[ редактировать ]Эвстатический уровень моря в триасе был постоянно низким по сравнению с другими геологическими периодами. Начало триаса приходилось примерно на современный уровень моря, поднявшись примерно на 10–20 метров (33–66 футов) над современным уровнем моря в раннем и среднем триасе. Повышение уровня моря ускорилось в Ладинском периоде, достигнув кульминации, когда уровень моря поднялся на 50 метров (164 фута) выше современного уровня во время Карнского периода. Уровень моря в Нориане начал снижаться, достигнув минимума на 50 метров (164 фута) ниже современного уровня моря в середине рэтского периода. Низкий глобальный уровень моря сохранялся вплоть до самой ранней юры. На долгосрочный тренд уровня моря накладываются 22 события падения уровня моря, широко распространенные в геологической летописи, в основном незначительные (менее 25 метров (82 фута)) и средние (25–75 метров (82–246 футов)) величины. Отсутствие доказательств существования триасовых континентальных ледниковых щитов позволяет предположить, что ледниковая эвстазия вряд ли может быть причиной этих изменений. [24]
Климат
[ редактировать ]Континентальный внутренний климат триаса в целом был жарким и сухим, поэтому типичными отложениями являются красные пластовые песчаники и эвапориты . Нет никаких свидетельств оледенения ни на одном полюсе, ни вблизи него; на самом деле, полярные регионы, очевидно, были влажными и умеренными , что обеспечивало климат, подходящий для лесов и позвоночных животных, включая рептилий. Большой размер Пангеи ограничивал сдерживающее воздействие мирового океана; его континентальный климат был сильно сезонным, с очень жарким летом и холодной зимой. [25] Сильный контраст между суперконтинентом Пангея и мировым океаном вызвал интенсивные трансэкваториальные муссоны . [25] иногда его называют пангейскими мегамуссонами . [26]
Триас, возможно, в основном был засушливым периодом, но существуют свидетельства того, что он перемежался несколькими эпизодами увеличения количества осадков в тропических и субтропических широтах моря Тетис и окружающей его суши. [27] Отложения и окаменелости, свидетельствующие о более влажном климате, известны от анизийского до ладинского периода Тетисского периода, а также от карнского и ретийского периодов на более обширной территории, включающей также бореальную область (например, Шпицберген острова ), Североамериканский континент, Южно- Китайский блок и Аргентина . Наиболее изученным из таких эпизодов влажного климата и, вероятно, самым интенсивным и широко распространенным было Карнийское плювиальное событие .
Ранний триас
[ редактировать ]Ранний триас был самой горячей частью всего фанерозоя, поскольку происходил во время и сразу после выброса титанических объемов парниковых газов из Сибирских траппов. Ранний триас начался с пермско-триасового термического максимума (PTTM), за которым последовало кратковременное динерское похолодание (DC) с 251 до 249 млн лет назад, за которым, в свою очередь, последовал последний Смитианский термический максимум (LSTT) около 249–248 млн лет назад. . Во время Новейшего оленекского похолодания (LOC), с 248 по 247 млн лет назад, температура понизилась примерно на 6 °C. [28]
Средний триас
[ редактировать ]Средний триас был прохладнее, чем ранний триас, при этом температуры падали на большей части анизийского периода, за исключением резкого потепления во второй части этапа. [29] С 242 по 233 млн лет назад последовало Ладинско-Карнийское похолодание (LCC). [28]
Поздний триас
[ редактировать ]В начале карнского периода глобальная температура продолжала оставаться относительно низкой. [30] Извержение Большой магматической провинции Врангелия около 234 млн лет назад вызвало резкое глобальное потепление, прекратив тенденцию к похолоданию LCC. [31] Это потепление стало причиной Карнийского плювиального события и привело к повсеместному распространению глобальной влажности. [32] CPE положил начало Среднекарнийскому теплому интервалу (MCWI), который длился от 234 до 227 млн лет назад. [28] На границе Карния и Нория произошло положительное δ 13 Считается, что отклонение C означает увеличение захоронения органического углерода. [33] С 227 по 217 млн лет назад был относительно прохладный период, известный как ранненорийский интервал похолодания (ENCI), после которого наступил средненорийский теплый интервал (MNWI) с 217 по 209 млн лет назад. MNWI был ненадолго прерван около 214 млн лет назад из-за похолодания, возможно, связанного с ударом Маникуагана . [28] Около 212 млн лет назад максимум эксцентриситета в 10 млн лет вызвал заболачивание Пангеи и сокращение размеров засушливых климатических зон. [34] Ретийский интервал похолодания (RCI) длился от 209 до 201 млн лет назад. [28] В конце триаса произошло экстремальное потепление, известное как Термальное событие конца триаса (ETTE), которое было ответственным за массовое вымирание в триасово-юрском периоде. [28] Пузырьки углекислого газа в базальтовых породах, датируемые концом триаса, указывают на то, что вулканическая активность из Центрально-Атлантической магматической провинции помогла спровоцировать изменение климата в ETTE. [35]
Флора
[ редактировать ]
Наземные растения
[ редактировать ]В раннем триасе ликофиты , особенно представители отряда Isoetales (который содержит живые иголки ), приобрели широкое распространение из-за нестабильности окружающей среды, последовавшей за пермско-триасовым вымиранием, причем одним особенно ярким примером является род Pleuromeia , который рос в столбчатых формах. мода, иногда достигающая высоты 2 метров (6,6 футов). Актуальность ликофитов снизилась, начиная со среднего триаса, после возвращения более стабильных условий окружающей среды. [36]
Вымершая группа семенных растений Bennettitales , впервые появившаяся в перми, впервые стала заметным элементом мировых флор в позднем триасе, и эту позицию они будут занимать на протяжении большей части мезозоя. [37] На территории Гондваны в южном полушарии дерево Dicroidium , вымерший « семенной папоротник », принадлежащее к отряду Corystospermales, было доминирующим элементом в лесных средах обитания по всему региону в среднем-позднем триасе. [38] В позднем триасе Ginkgoales (которые сегодня представлены только одним видом, Ginkgo biloba ) претерпели значительную диверсификацию. [39] Хвойные деревья были в изобилии в триасовом периоде, включая Voltziales ( которые включают различные линии, вероятно, включая те, которые являются предками современных хвойных деревьев), [40] а также вымершее семейство Cheirolepidiaceae , которое впервые появилось в позднем триасе и будет широко распространено на протяжении большей части остальной части мезозоя. [41]
Уголь
[ редактировать ]
Ни одно известное месторождение угля не относится к началу триасового периода. Это известно как «угольный разрыв» раннего триаса и может рассматриваться как часть пермско-триасового вымирания . [43] Возможные объяснения угольного разрыва включают резкое падение уровня моря во время границы пермо-триаса; [44] кислотные дожди в результате извержений Сибирских траппов или ударного события, захлестнувшего кислые болота; переход климата к тепличному климату, который был слишком жарким и сухим для накопления торфа; эволюция грибов и травоядных животных, которые были более разрушительными для водно-болотных угодий; вымирание всех растений, адаптированных к торфяным болотам, с перерывом в несколько миллионов лет, прежде чем появились новые виды растений, адаптированные к торфяным болотам; [43] или аноксия почвы, когда уровень кислорода резко упал. [45]
Фитопланктон
[ редактировать ]До пермского вымирания архепластиды (красные и зеленые водоросли) были основным морским фитопланктоном примерно 659–645 миллионов лет назад. [46] когда они заменили морские планктонные цианобактерии , впервые появившиеся около 800 миллионов лет назад, в качестве доминирующего фитопланктона в океанах. [47] В триасе вторичные эндосимбиотические водоросли. важнейшим планктоном стали [48]
фауна
[ редактировать ]
Морские беспозвоночные
[ редактировать ]В морской среде новые современные типы кораллов появились в раннем триасе, образуя небольшие участки рифов умеренной протяженности по сравнению с огромными рифовыми системами девона или современности. В конце Карния в Южном Китае произошел рифовый кризис. [49] Серпулиды появились в среднем триасе. [50] Микроконхиды были многочисленными. Головоногие моллюски с панцирем, называемые аммонитами, восстановились, образовавшись из одной линии, пережившей пермское вымирание. Двустворчатые моллюски начали быстро диверсифицироваться в среднем триасе, став очень многочисленными в океанах. [51]
Насекомые
[ редактировать ]Водные насекомые быстро диверсифицировались в течение среднего триаса, причем этот временной интервал представляет собой решающую диверсификацию для Holometabola , клады, содержащей большинство современных видов насекомых. [52]
Рыба
[ редактировать ]
После вымирания ихтиофауна пермско-триасового массового была удивительно однородной, многие семейства и роды демонстрировали космополитическое распространение . [53] Целаканты демонстрируют наибольшее постдевонское разнообразие в раннем триасе . [54] Лучеперые рыбы (актиноптеригии) претерпели значительную диверсификацию в начале триаса, что привело к пику разнообразия в среднем триасе; однако характер этой диверсификации до сих пор не совсем понятен из-за тафономической мегабиастии . [55] Первые стволовой группы костистые кости появились в триасе (костистые кости сегодня являются, безусловно, самой разнообразной группой рыб). [53] Хищные актиноптеригии, такие как заурихтииды и биргерииды , некоторые из которых вырастали более 1,2 м (3,9 фута) в длину, появились в раннем триасе и получили широкое распространение и успех в течение этого периода в целом. [56] Озера и реки были населены двоякодышащими рыбами (Dipnoi), такими как Ceratodus , которые в основном известны по зубным пластинкам, изобилующим в летописи окаменелостей. [57] Гибодонты , группа акулоподобных хрящевых рыб , доминировали как в пресноводной, так и в морской среде на протяжении всего триаса. [58] Последние выжившие представители преимущественно палеозойских Eugeneodontida известны из раннего триаса. [59]
Земноводные
[ редактировать ]
Темноспондиловые амфибии были среди тех групп, которые пережили пермско-триасовое вымирание. Когда-то наземные виды были широко распространены как в наземной, так и в водной среде, но во время вымирания они в основном вымерли. Выжившие в триасе вели водный или полуводный образ жизни и были представлены Tupilakosaurus , Thabanchuia , Branchiosauridae и Micropholis , все из которых вымерли в раннем триасе, а также успешными Stereospondyli , выжившими в меловой период. Самые крупные стереоспондилы триаса, такие как мастодонзавр , имели длину от 4 до 6 метров (от 13 до 20 футов). [60] [61] Некоторые линии (например, трематозавры ) недолго процветали в раннем триасе, в то время как другие (например, капитозавры ) оставались успешными на протяжении всего периода или получили известность только в позднем триасе (например, плагиозавры , метопозавры ).
Первые лиссамфибии (современные амфибии) появляются в триасе, а прародители первых лягушек появились уже в раннем триасе. Однако группа в целом не стала распространенной до юрского периода , когда темноспондилы стали очень редкими.
Большая часть Reptiliomorpha , стеблевых амниот, давших начало амниотам, исчезла в триасе, но выжили две водные группы: Embolomeri , которые дожили только до начала периода, и Chroniosuria , которые дожили до конца триасовый период.
Рептилии
[ редактировать ]Архозавроморфы
[ редактировать ]Пермско-триасовое вымирание опустошило наземную жизнь. Биоразнообразие восстановилось, когда выжившие виды вновь заселили пустующую местность, но это продолжалось недолго. На восстановление разнообразных сообществ со сложной структурой пищевой сети потребовалось 30 миллионов лет. [10] [62] Рептилии -архозавроморфы , которые уже появились и в некоторой степени разнообразились в пермский период, резко возросли в разнообразии в результате адаптивной радиации в ответ на массовое вымирание в пермском и триасовом периоде. К раннему триасу появилось несколько крупных групп архозавроморфов. Ранние архозавроморфы с длинной шеей, похожие на ящериц, были известны как проторозавры , которые, вероятно, являются парафилетической группой, а не настоящей кладой. Танистрофеиды были семейством проторозавров, у которых размер шеи достигал крайних пределов, причем у крупнейшего рода Tanystropheus шея длиннее тела. Семейство проторозавров Sharovipterygidae использовало свои удлиненные задние конечности для планирования. Другие архозавроморфы, такие как ринхозавры и аллокотозавры , были в основном травоядными с коренастым телом и специализированной структурой челюстей.
Ринхозавры, травоядные животные с бочкообразными потрохами, процветали лишь короткий период времени и вымерли около 220 миллионов лет назад. Они были исключительно многочисленны в середине триаса как основные крупные травоядные животные во многих экосистемах карнийского возраста. Они стригли растения с предчелюстными клювами и пластинками вдоль верхней челюсти с несколькими рядами зубов. Аллокотозавры были игуаноподобными рептилиями, в том числе трилофозавром (обычная рептилия позднего триаса с зубами с тремя коронками), тератерпетоном (с длинной клювообразной мордой) и шрингазавром (рогатым травоядным, длина тела которого достигала 3–4 метров). (9,8–13,1 фута)).
Одна группа архозавроморфов, архозавроформ , отличалась активным хищническим образом жизни, зазубренными зубами и вертикальным положением конечностей. Архозавроформы в триасе были разнообразны, включая различных наземных и околоводных хищников всех форм и размеров. Большеголовые и крепкие эритрозухиды были одними из доминирующих хищников в раннем триасе. Фитозавры были особенно распространенной группой, процветавшей в позднем триасе. Эти длиннорылые полуводные хищники напоминают ныне живущих крокодилов и, вероятно, вели аналогичный образ жизни: охотились на рыбу и мелких рептилий у кромки воды. Однако это сходство лишь поверхностное и является ярким примером конвергентной эволюции.
Настоящие архозавры появились в раннем триасе, разделившись на две ветви: Avemetatarsalia (предки птиц) и Pseudosuria (предки крокодилов). Авеметатарзалии были второстепенным компонентом их экосистем, но в конечном итоге произвели самых ранних птерозавров и динозавров в позднем триасе. Ранние длиннохвостые птерозавры появились в Нориане и быстро распространились по всему миру. Триасовые динозавры развились в карнийском периоде и включают ранних зауроподоморфов и теропод. Большинство триасовых динозавров были мелкими хищниками, и лишь немногие из них были обычными, например Coelophys , длина которого составляла от 1 до 2 метров (от 3,3 до 6,6 футов). Триасовые зауроподоморфы в основном населяли более прохладные регионы мира. [63]
Крупный хищник Смок, скорее всего, также был архозавром, но неясно, был ли это примитивный динозавр или псевдозух.
Псевдозухи были гораздо более экологически доминирующими в триасе, включая крупных травоядных (таких как этоозавры ), крупных плотоядных (« рауизухии ») и первых крокодиломорфов (« сфенозухи »). Этозавры были тяжелобронированными рептилиями, которые были обычным явлением в течение последних 30 миллионов лет позднего триаса, пока не вымерли во время триасово-юрского вымирания. Большинство этозавров были травоядными и питались низкорослыми растениями, но некоторые, возможно, ели мясо. « Раузухи » (ранее известные как паракрокодиломорфы ) были ключевыми хищниками большинства наземных экосистем триаса. Было обнаружено более 25 видов, включая гигантских четвероногих охотников, гладких двуногих всеядных и неуклюжих зверей с глубокими парусами на спине. Вероятно, они заняли нишу крупных хищников, позже заполненную тероподами. «Рауизухаи» были предками небольших крокодиломорфов легкого телосложения, единственных псевдозухий, доживших до юрского периода.
 Танистрофей , с длинной шеей. танистрофейд
Танистрофей , с длинной шеей. танистрофейд Proterosuchus , ранний архозавроформ, похожий на крокодила, из раннего триаса.
Proterosuchus , ранний архозавроформ, похожий на крокодила, из раннего триаса. Стаурикозавр , один из самых ранних динозавров, член триасового семейства Herrerasauridae.
Стаурикозавр , один из самых ранних динозавров, член триасового семейства Herrerasauridae. Postosurus , рауизухид , который был высшим хищником в некоторых частях позднего триаса Северной Америки.
Postosurus , рауизухид , который был высшим хищником в некоторых частях позднего триаса Северной Америки. Платеозавр был одним из крупнейших ранних зауроподоморфов , или «просауроподов», позднего триаса.
Платеозавр был одним из крупнейших ранних зауроподоморфов , или «просауроподов», позднего триаса. Целофиз был одним из самых многочисленных динозавров-теропод в позднем триасе.
Целофиз был одним из самых многочисленных динозавров-теропод в позднем триасе.



Морские рептилии
[ редактировать ]
Существовало множество видов морских рептилий. К ним относятся Sauropterygia , среди которых были пахиплеврозавры и нотозавры (оба были распространены в среднем триасе, особенно в районе Тетиса ), плакодонты , самая ранняя известная травоядная морская рептилия Atopodentatus и первые плезиозавры . Первые из ящероподобных талаттозавров ( аскептозавров ) и весьма успешные ихтиозавры , появившиеся в морях раннего триаса, вскоре диверсифицировались, а некоторые в конечном итоге развились до огромных размеров в позднем триасе.
Другие рептилии
[ редактировать ]Среди других рептилий самые ранние черепахи , такие как Proganochelys и Proterochersis , появились в норийский век (этап) позднего триаса. Lepidosauromorpha Sphenodontia , в частности . , впервые были обнаружены в летописи окаменелостей раннего карнийского периода, хотя самые ранние лепидозавроморфы, вероятно, произошли в перми Procolophonidae парарептилии , последние выжившие , были важной группой мелких ящерицеобразных травоядных животных. Дрепанозавры . представляли собой кладу необычных древесных рептилий, похожих на хамелеонов, с птичьими головами и специальными когтями
Синапсиды
[ редактировать ]три группы терапсидов В триасовый период сохранились : дицинодонты , тероцефалы и цинодонты . Цинодонт Cynognathus был характерным высшим хищником оленекского и анизийского периода Гондваны . И каннемейериформные дицинодонты, и гомфодонтные цинодонты оставались важными травоядными животными на протяжении большей части периода. Тероцефалы включали как крупных хищников ( Moschorhinus ), так и травоядных форм ( баурииды ) до их исчезновения в середине периода. Цинодонты Ecteniniid играли роль крупных бегающих хищников в позднем триасе. В карнийском периоде (начало позднего триаса) некоторые продвинутые цинодонты дали начало первым млекопитающим .
В триасе архозавры вытеснили терапсидов как крупнейших и наиболее экологически плодовитых наземных амниот. Это «триасовое поглощение», возможно, способствовало эволюции млекопитающих , заставив выживших терапсидов и их преемников в форме млекопитающих жить как мелкие, в основном ночные насекомоядные животные . Ночной образ жизни , возможно, заставил млекопитающих обзавестись мехом и повысить скорость метаболизма . [65]
 Листрозавр был широко распространенным дицинодонтом и наиболее распространенным наземным позвоночным в раннем триасе, после того как жизнь животных значительно сократилась.
Листрозавр был широко распространенным дицинодонтом и наиболее распространенным наземным позвоночным в раннем триасе, после того как жизнь животных значительно сократилась.


депозиты
[ редактировать ]Два раннего триаса лагерштеттена (высококачественные ископаемые пласты), динерийского возраста . биота Гуйян [66] и самая ранняя спатического возраста парижская биота [67] выделяются своей исключительной сохранностью и разнообразием . Они представляют собой самые ранние лагерштетты мезозойской эры и дают представление о восстановлении биотики после массового вымирания в пермско-триасовом периоде.
Лагерштетте Монте -Сан-Джорджио , ныне расположенное в районе озера Лугано на севере Италии и юге Швейцарии , во среднего триаса времена представляло собой лагуну за рифами с бескислородным нижним слоем, поэтому там не было падальщиков и небольшая турбулентность, которая могла бы нарушить окаменелость, - ситуация, которая может его можно сравнить с более известным юрским из известняка Зольнхофен лагерштеттом . останки рыб и различных морских рептилий (в том числе обыкновенного пахиплеврозавра Neusticosaurus и причудливого длинношеего архозавроморфа Tanystropheus ), а также некоторых наземных форм, таких как Ticinosuchus и Macrocnemus В этом местонахождении были обнаружены . Все эти окаменелости датируются анизийским и ладинским возрастом (около 242 млн лет назад).
Триасово-юрское вымирание
[ редактировать ]
Триасовый период закончился массовым вымиранием, которое было особенно суровым в океанах; конодонты кроме исчезли, как и все морские рептилии, ихтиозавров и плезиозавров . беспозвоночные, такие как брахиоподы и моллюски (например, брюхоногие Серьезно пострадали моллюски). В океанах пропало 22% морских семейств и, возможно, около половины морских родов.
Хотя вымирание в конце триаса не было одинаково разрушительным для всех наземных экосистем, исчезло несколько важных клад круротарсанов (крупных архозавровых рептилий, ранее сгруппированных вместе как текодонты ), а также большинство крупных лабиринтодонтных амфибий, групп мелких рептилий и большинства синапсиды. Некоторые из ранних примитивных динозавров также вымерли, но более адаптивные выжили и эволюционировали в юрский период. Среди сохранившихся растений, которые впоследствии доминировали в мезозойском мире, были современные хвойные деревья и цикадеоиды.
Причина позднетриасового вымирания неясна. Это сопровождалось огромными извержениями вулканов , которые произошли, когда суперконтинент Пангея начал распадаться примерно от 202 до 191 миллиона лет назад (даты 40Ar/39Ar). [68] формирование Центрально-Атлантической магматической провинции (CAMP), [69] одно из крупнейших известных внутренних вулканических событий с тех пор, как планета впервые остыла и стабилизировалась. Другие возможные, но менее вероятные причины вымирания включают глобальное похолодание или даже удар болида ударный кратер, содержащий водохранилище Маникуаган в Квебеке , Канада , для которого был выделен . Однако ударный расплав Маникуагана датируется 214 ± 1 млн лет назад. Дата границы триаса и юры также была недавно установлена более точно - 201,4 млн лет назад. Обе даты становятся точнее благодаря использованию более точных форм радиометрического датирования, в частности, распада урана на свинец в цирконах, образовавшихся во время удара. Таким образом, данные свидетельствуют о том, что воздействие Маникуагана предшествовало концу триаса примерно на 10 ± 2 млн лет назад. Следовательно, это не могло быть непосредственной причиной наблюдаемого массового вымирания. [70]

Количество вымираний в позднем триасе оспаривается. Некоторые исследования предполагают, что в конце триаса существует как минимум два периода вымирания, разделенные промежутком от 12 до 17 миллионов лет. Но против этого можно привести недавнее исследование фауны Северной Америки. В Окаменелом лесу на северо-востоке Аризоны находится уникальная последовательность наземных отложений позднего карния и раннего норийского периода. Анализ, проведенный в 2002 году, не обнаружил существенных изменений в палеосреде. [71] Фитозавры , наиболее распространенные там ископаемые, претерпели смену только на уровне рода, а количество видов осталось прежним. Некоторые этоозавры , следующие по распространенности четвероногие, и ранние динозавры остались неизменными. Однако и фитозавры, и этоозавры были среди групп рептилий-архозавров, полностью уничтоженных в результате вымирания в конце триаса.
Кажется вероятным, что тогда имело место своего рода позднекарнийское вымирание, когда вымерло несколько травоядных групп архозавроморф, в то время как крупные растительноядные терапсиды — дицинодонты каннемейериид и цинодонты траверсодонты — значительно сократились в северной половине Пангеи ( Лавразия ).
Эти вымирания в триасе и в его конце позволили динозаврам занять многие ниши, которые стали незанятыми. Динозавры становились все более доминирующими, многочисленными и разнообразными и оставались такими в течение следующих 150 миллионов лет. Настоящая «Эра динозавров» приходится на юрский и меловой периоды, а не на триасовый период.
См. также
[ редактировать ]- Геологическая шкала времени
- Список мест окаменелостей (с каталогом ссылок)
- Фауны наземных позвоночных триаса
- Филлоцератина
- Динозавры
Примечания
[ редактировать ]- ^ Видманн, Филипп; Бучер, Хьюго; Леу, Марк; и др. (2020). «Динамика крупнейшего выброса изотопов углерода во время биотического восстановления раннего триаса» . Границы в науках о Земле . 8 (196): 196. Бибкод : 2020FrEaS...8..196W . дои : 10.3389/feart.2020.00196 .
- ^ МакЭлвейн, Дж. К.; Пуньясена, Юго-Запад (2007). «Массовые вымирания и летопись окаменелостей растений». Тенденции в экологии и эволюции . 22 (10): 548–557. дои : 10.1016/j.tree.2007.09.003 . ПМИД 17919771 .
- ^ Реталлак, Дж.Дж.; Виверс, Дж .; Моранте, Р. (1996). «Глобальный угольный разрыв между пермско-триасовым вымиранием и восстановлением торфообразующих растений в среднем триасе» . Бюллетень ГСА . 108 (2): 195–207. Бибкод : 1996GSAB..108..195R . doi : 10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2 . Проверено 29 сентября 2007 г.
- ^ Пейн, Дж.Л.; Лерманн, диджей; Вэй, Дж.; Орчард, MJ; Шраг, ДП; Нолл, АХ (2004). «Большие возмущения углеродного цикла во время восстановления после вымирания в конце перми» . Наука . 305 (5683): 506–9. Бибкод : 2004Sci...305..506P . дои : 10.1126/science.1097023 . ПМИД 15273391 . S2CID 35498132 .
- ^ Огг, Джеймс Г.; Огг, Габи М.; Градштейн, Феликс М. (2016). «Триас». Краткая геологическая временная шкала: 2016 г. Эльзевир. стр. 133–149. ISBN 978-0-444-63771-0 .
- ^ Хунфу, Инь; Кэсинь, Чжан; Джиннан, Тонг; Цзуньи, Ян; Шуньбао, Ву (июнь 2001 г.). «Глобальный стратотипический разрез и точка (GSSP) границы перми и триаса» (PDF) . Эпизоды . 24 (2): 102–114. дои : 10.18814/epiiugs/2001/v24i2/004 . Проверено 8 декабря 2020 г.
- ^ Хиллебрандт, Авеню; Кристин, Л.; Кюршнер, ВМ; и др. (сентябрь 2013 г.). «Глобальные стратотипические разрезы и точки (GSSP) основания юрской системы в Куйохе (горы Карвендель, Северные известняковые Альпы, Тироль, Австрия)». Эпизоды . 36 (3): 162–198. CiteSeerX 10.1.1.736.9905 . дои : 10.18814/epiiugs/2013/v36i3/001 . S2CID 128552062 .
- ^ «Триас» . Dictionary.com Полный (онлайн). nd
- ^ «Международная стратиграфическая карта» (PDF) . Международная комиссия по стратиграфии. Июнь 2023 года . Проверено 19 июля 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Сахни, С. и Бентон, М.Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен» . Труды Королевского общества B: Биологические науки . 275 (1636): 759–765. дои : 10.1098/rspb.2007.1370 . ПМЦ 2596898 . ПМИД 18198148 .
- ^ Брусатте, СЛ; Бентон, MJ; Рута, М.; Ллойд, GT (12 сентября 2008 г.). «Превосходство, конкуренция и оппортунизм в эволюционном излучении динозавров» (PDF) . Наука . 321 (5895): 1485–1488. Бибкод : 2008Sci...321.1485B . дои : 10.1126/science.1161833 . hdl : 20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b . ПМИД 18787166 . S2CID 13393888 . Архивировано из оригинала (PDF) 24 июня 2014 г. Проверено 14 января 2012 г.
- ^ « Смертельно горячая» Земля была лишена жизни – могло ли это случиться снова?» . Nationalgeographic.com . 19 октября 2012 г. Архивировано из оригинала 20 октября 2012 г.
- ^ Фридрих фон Альберти, Вклад в монографию о цветном песчанике, ракушечнике и аргиллите и объединении этих структур в одно образование (Штутгарт и Тюбинген (Германия): JG Cotta, 1834). Альберти ввёл термин «триас» на странице 324 :
"...цветной песчаник, Мушелкальк и Кейпер являются результатом периода, их окаменелости, выражаясь словами Э. де Бомона, являются термометрами геологической эпохи,... поэтому разделение этих структур на 3 формации , наблюдавшееся до сих пор, неуместно, и больше соответствовало бы концепции формации Триадой ». объединить их в формацию, которую я условно назову
(… цветной песчаник, ракушечник и аргиллит являются результатом определенного периода; их окаменелости являются, пользуясь словами Э. де Бомона, термометром геологической эпохи; … таким образом, разделение этих структур на 3 формации , которая сохранялась до сих пор, неуместна, и более соответствует понятию «формация» объединить их в одну формацию, которую я пока назову «триас».) - ^ Форте, Джузеппа; Кустачер, Эвелин; Прето, Нерео (1 сентября 2022 г.). «Вариации изотопов углерода (δ13C) указывают на климатические сдвиги и отражают среду обитания растений в последовательности среднего триаса (анизийский, пельсонский период) в Кювизенкопфе / Монте-Пра-делла-Вакка (Доломитовые Альпы, Северо-Восточная Италия)» . Палеогеография, Палеоклиматология, Палеоэкология . 601 : 111098. Бибкод : 2022PPP...60111098F . дои : 10.1016/j.palaeo.2022.111098 . S2CID 249483335 . Проверено 1 декабря 2022 г.
- ^ Мазахери-Джохари, Мина; Роги, Гвидо; Каджати, Марчелло; Кустачер, Эвелин; Гасеми-Неджад, Ибрагим; Занчи, Андреа; Джанолла, Пьеро (15 января 2022 г.). «Распутывание климатического сигнала от тектонических воздействий: триасовый Агдарбандский бассейн (Туранский домен, Иран)» . Палеогеография, Палеоклиматология, Палеоэкология . 586 : 110777. Бибкод : 2022PPP...58610777M . дои : 10.1016/j.palaeo.2021.110777 . S2CID 244696034 . Проверено 10 января 2023 г.
- ↑ Перейти обратно: Перейти обратно: а б Кокс, Л. Робин М. и Торсвик, Тронд Х., ред. (2016). «Триас» . История Земли и палеогеография . Кембридж, Великобритания: Издательство Кембриджского университета. стр. 195–207. дои : 10.1017/9781316225523.012 . ISBN 978-1-316-22552-3 . Проверено 16 мая 2022 г.
- ^ Герберт, Крис; Хелби, Робин (1980). Путеводитель по бассейну Сиднея . Мейтленд, Новый Южный Уэльс: Геологическая служба Нового Южного Уэльса. п. 582. ИСБН 978-0-7240-1250-3 .
- ^ «Лекция 10 – Триас: Ньюарк, Чинл» . Rainbow.ldeo.columbia.edu .
- ^ Джейкобс, Луи Л. (1997). «Африканские динозавры». В Карри, Филипп Дж.; Падиан, Кевин (ред.). Энциклопедия динозавров . Академическая пресса. стр. 2–4.
- ↑ Перейти обратно: Перейти обратно: а б Лидмар-Бергстрем, Карна (1993). «Денудационные поверхности и тектоника в самой южной части Балтийского щита». Докембрийские исследования . 64 (1–4): 337–345. Бибкод : 1993PreR...64..337L . дои : 10.1016/0301-9268(93)90086-h .
- ↑ Перейти обратно: Перейти обратно: а б Олесен, Одлеев; Кирульф, Хальвдан Паскаль; Броннер, Марко; Далсегг, Эйнар; Фредин, Ола; Солбакк, Терье (2013). «Глубокое выветривание, неотектоника и образование пляжных равнин в Нурланде, северная Норвегия». Норвежский геологический журнал . 93 : 189–213.
- ^ Япсен, Питер; Грин, Пол Ф; Боноу, Йохан М; Эрльстрем, Микаэль (2016). «Эпизодические захоронения и эксгумация южной части Балтийского щита: эпейрогенные поднятия во время и после распада Пангеи». Исследования Гондваны . 35 : 357–77. Бибкод : 2016GondR..35..357J . дои : 10.1016/j.gr.2015.06.005 .
- ^ Фредин, Ола; Виола, Джулио; Цвинманн, Хорст; Сёрли, Рональд; Брённер, Марко; Ложь, Ян-Эрик; и др. (2017). «Наследие мезозойского ландшафта западной Скандинавии» . Природа . 8 : 14879. Бибкод : 2017NatCo...814879F . дои : 10.1038/ncomms14879 . ПМЦ 5477494 . ПМИД 28452366 .
- ^ Хак, Билал У. (декабрь 2018 г.). «Пересмотр эвстатических вариаций триаса» . ГСА сегодня . 28 (12). Геологическое общество Америки : 4–9. Бибкод : 2018GSAT...28l...4H . дои : 10.1130/GSATG381A.1 . S2CID 134477691 .
- ↑ Перейти обратно: Перейти обратно: а б Стэнли, 452–53.
- ^ Цзэн, Живэй; Чжу, Хунтао; Ян, Сянхуа; Цзэн, Хунлю; Ху, Сяолинь; Ся, Чэньчэнь (май 2019 г.). «Записи мегамуссонов в Пангее: данные из триасовой формации Мангару, северо-западный шельф Австралии» . Исследования Гондваны . 69 : 1–24. Бибкод : 2019GondR..69....1Z . дои : 10.1016/j.gr.2018.11.015 . S2CID 134145664 . Проверено 9 января 2023 г.
- ^ Прето, Н.; Кустачер, Э.; Виналл, ПБ (2010). «Триасовый климат – современное состояние и перспективы». Палеогеография, Палеоклиматология, Палеоэкология . 290 (1–4): 1–10. Бибкод : 2010PPP...290....1P . дои : 10.1016/j.palaeo.2010.03.015 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Скотезе, Кристофер Роберт; Сун, Хайджун; Миллс, Бенджамин Дж.В.; ван дер Меер, Дау Г. (1 апреля 2021 г.). «Палеотемпературы фанерозоя: изменение климата Земли за последние 540 миллионов лет» . Обзоры наук о Земле . 215 : 103503. Бибкод : 2021ESRv..21503503S . doi : 10.1016/j.earscirev.2021.103503 . ISSN 0012-8252 . S2CID 233579194 . Проверено 22 сентября 2023 г.
- ^ Троттер, Джули А.; Уильямс, Ян С.; Никора, Альда; Мацца, Мишель; Риго, Мануэль (апрель 2015 г.). «Долгосрочные циклы изменения климата в триасе: новый рекорд δ18O из конодонтового апатита» . Письма о Земле и планетологии . 415 : 165–174. Бибкод : 2015E&PSL.415..165T . дои : 10.1016/j.epsl.2015.01.038 . Проверено 22 сентября 2023 г.
- ^ Даль Корсо, Якопо; Миллс, Бенджамин Дж.В.; Чу, Даолян; Ньютон, Роберт Дж.; Сон, Хайджун (15 января 2022 г.). «Фоновое состояние земной системы усилило карнийские (поздний триас) изменения окружающей среды» . Письма о Земле и планетологии . 578 : 117321. Бибкод : 2022E&PSL.57817321D . дои : 10.1016/j.epsl.2021.117321 . S2CID 244847207 . Проверено 22 сентября 2023 г.
- ^ Даль Корсо, Дж.; Миетто, П.; Ньютон, Р.Дж.; Панкост, РД; Прето, Н.; Роги, Г.; Виналл, ПБ (1 января 2012 г.). «Обнаружение крупного отрицательного всплеска 13C в Карнии (поздний триас), связанного с извержением базальтов паводка Врангелии» . Геология . 40 (1): 79–82. Бибкод : 2012Geo....40...79D . дои : 10.1130/G32473.1 . ISSN 0091-7613 . Проверено 22 сентября 2023 г.
- ^ Ли, Лицинь; Кюршнер, Вольфрам М.; Лу, Нин; Чен, Хунъюй; Ань, Пэнчэн; Ван, Юндон (сентябрь 2022 г.). «Палинологическая запись карнийского плювиального эпизода из северо-западной части бассейна Сычуань, юго-запад Китая» . Обзор палеоботаники и палинологии . 304 : 104704. Бибкод : 2022RPaPa.30404704L . дои : 10.1016/j.revpalbo.2022.104704 . hdl : 10852/99190 . S2CID 249528886 . Проверено 22 сентября 2023 г.
- ^ Муттони, Джованни; Мацца, Мишель; Мошер, Дэвид; Кац, Мириам Э.; Кент, Деннис В.; Балини, Марко (1 апреля 2014 г.). «Запись изотопов углерода и кислорода среднего-позднего триаса (ладинский-ретийский период) из Тетического океана» . Палеогеография, Палеоклиматология, Палеоэкология . 399 : 246–259. Бибкод : 2014PPP...399..246M . дои : 10.1016/j.palaeo.2014.01.018 . ISSN 0031-0182 . Проверено 24 ноября 2023 г.
- ^ Икеда, Масаюки; Одзаки, Кадзуми; Легран, Жюльен (23 июля 2020 г.). «Влияние динамики муссонов масштаба 10 млн лет на мезозойский климат и экосистемы» . Научные отчеты . 10 (1): 11984. Бибкод : 2020НатСР..1011984I . doi : 10.1038/s41598-020-68542-w . ISSN 2045-2322 . ПМЦ 7378230 . ПМИД 32704030 .
- ^ Каприоло, Манфредо; Марзоли, Андреа; Аради, Ласло Э.; Каллегаро, Сара; Даль Корсо, Якопо; Ньютон, Роберт Дж.; Миллс, Бенджамин Дж.В.; Виналл, Пол Б.; Бартоли, Омар; Бейкер, Дон Р.; Юби, Насриддин; Ремюза, Лоран; Списс, Ричард; Сабо, Чаба (7 апреля 2020 г.). «Глубинный CO2 в магматической провинции Центральной Атлантики конца триаса» . Природные коммуникации . 11 (1): 1670. Бибкод : 2020NatCo..11.1670C . дои : 10.1038/s41467-020-15325-6 . ISSN 2041-1723 . ПМЦ 7138847 . ПМИД 32265448 .
- ^ Луй, Синди В.; ван Конийненбург-ван Циттерт, Йоханна Х.А.; Дуйнсти, Иво АП (2 марта 2021 г.). «Распространение изоэтальных ликофитов во время пермо-триасовых биотических кризисов: показатель состояния земной биосферы» . Границы в науках о Земле . 9 : 55. Бибкод : 2021FrEaS...9...55L . дои : 10.3389/feart.2021.615370 . ISSN 2296-6463 .
- ^ Бломенкемпер, Патрик; Боймер, Роберт; Бэкер, Мальте; Абу Хамад, Абдалла; Ван, Цзюнь; Керп, Ганс; Бомфлер, Бенджамин (2021). «Беннеттитальные листья из перми экваториальной Пангеи — раннее излучение культовой мезозойской группы голосеменных» . Границы в науках о Земле . 9 : 162. Бибкод : 2021FrEaS...9..162B . дои : 10.3389/feart.2021.652699 .
- ^ Мэйс, Крис; Маклафлин, Стивен (25 февраля 2020 г.). «Оказавшись между двумя массовыми вымираниями: взлет и падение дикроидия» . Депозиты Маг . Проверено 23 сентября 2023 г.
- ^ Чжоу, Чжи-Янь (март 2009 г.). «Обзор ископаемых Ginkgoales» . Палеомир . 18 (1): 1–22. дои : 10.1016/j.palwor.2009.01.001 .
- ^ Форте, Джузеппа; Кустачер, Эвелин; Ван Конийненбург-ван Циттерт, Йоханна Х.А. (1 июля 2021 г.). «Разнообразие хвойных пород в среднем триасе: новые данные из Fossillagerstätte Kühwiesenkopf/Monte Prà della Vacca (пельсонский ярус, анизий) в Доломитовых Альпах (Северо-Восточная Италия)» . Международный журнал наук о растениях . 182 (6): 445–467. дои : 10.1086/714280 . ISSN 1058-5893 .
- ^ Эскапа, Игнасио; Лесли, Эндрю (февраль 2017 г.). «Новые Cheirolepidiaceae (Coniferales) из ранней юры Патагонии (Аргентина): согласование записей отпечатков и перминерализованных окаменелостей» . Американский журнал ботаники . 104 (2): 322–334. дои : 10.3732/ajb.1600321 . hdl : 11336/40738 . ISSN 0002-9122 . ПМИД 28213347 .
- ^ Хошер, В.Т. Магаритц М. Кларк Д. (1987). «События вблизи времени границы перми и триаса». Мод. Геол . 11 : 155–80 [173–74].
- ↑ Перейти обратно: Перейти обратно: а б Реталлак, Дж.Дж.; Виверс, Джей Джей; Моранте, Р. (1996). «Глобальный угольный разрыв между пермско-триасовым вымиранием и восстановлением торфообразующих растений в среднем триасе». Бюллетень Геологического общества Америки . 108 (2): 195–207. Бибкод : 1996GSAB..108..195R . doi : 10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2 .
- ^ Хольсер, WT; Шенлауб, HP; Кляйн, П; Аттреп, М; Бекельманн, Клаус; и др. (1989). «Уникальная геохимическая запись на границе перми и триаса» . Природа . 337 (6202): 39 [42]. Бибкод : 1989Natur.337...39H . дои : 10.1038/337039a0 . S2CID 8035040 . Проверено 24 ноября 2023 г.
- ^ Реталлак, Дж.Дж.; Крулл, ES (2006). «Изотопные данные углерода о выбросах метана в конечном пермском периоде и их роли в вымирании животных, растений, коралловых рифов и торфяных болот» (PDF) . Специальный доклад Геологического общества Америки . 399 : 249. дои : 10.1130/2006.2399(12) . ISBN 978-0-8137-2399-0 . Проверено 14 декабря 2020 г.
- ^ «Как Земля-снежок породила сложную жизнь – журнал «Космос»» . 16 августа 2017 г.
- ^ «Декабрь: Фитопланктон | Новости | Бристольский университет» .
- ^ «Рост водорослей в криогенных океанах и появление животных – ResearchGate» .
- ^ Цзинь, Синь; Ши, Чжицян; Риго, Мануэль; Франчески, Марко; Прето, Нерео (15 сентября 2018 г.). «Кризис карбонатной платформы в карнии (поздний триас) Ханьвана (бассейн Сычуань, Южный Китай): данные о конодонтах и данные о стабильных изотопах» . Журнал азиатских наук о Земле . 164 : 104–124. Бибкод : 2018JAESc.164..104J . дои : 10.1016/j.jseaes.2018.06.021 . S2CID 134733944 . Проверено 22 сентября 2023 г.
- ^ Винн, О.; Мутвей, Х. (2009). «Известковые трубчатые черви фанерозоя» (PDF) . Эстонский журнал наук о Земле . 58 (4): 286–96. дои : 10.3176/earth.2009.4.07 . Проверено 16 сентября 2012 г.
- ^ Комацу, Тосифуми; Чен, Цзинь-хуа; Цао, Мэй-чжэнь; Стиллер, Фрэнк; Нарусэ, Хадзиме (10 июня 2004 г.). «Разнообразные двустворчатые моллюски среднего триаса (анизийского периода): условия отложения и комплексы двустворчатых моллюсков в пачке Лейдапо формации Цинъянь, южный Китай» . Палеогеография, Палеоклиматология, Палеоэкология . 208 (3–4): 227–223. Бибкод : 2004PPP...208..207K . дои : 10.1016/j.palaeo.2004.03.005 . Проверено 31 марта 2023 г.
- ^ Чжэн, Даран; Чанг, Су-Чин; Ван, Хэ; Фанг, Ян; Ван, Цзюнь; Фэн, Чунцин; Се, Гувэй; Яржембовский, Эдмунд А.; Чжан, Хайчунь; Ван, Бо (7 сентября 2018 г.). «Радиация насекомых среднего и позднего триаса, выявленная по разнообразным окаменелостям и изотопному возрасту из Китая» . Достижения науки . 4 (9): eaat1380. Бибкод : 2018SciA....4.1380Z . дои : 10.1126/sciadv.aat1380 . ISSN 2375-2548 . ПМК 6124916 . ПМИД 30191177 .
- ↑ Перейти обратно: Перейти обратно: а б Романо, Карло; Кут, Марта Б.; Коган, Илья; Брайард, Арно; Миних Алла Владимировна; Бринкманн, Винанд; Бучер, Хьюго; Кривет, Юрген (февраль 2016 г.). «Пермско-триасовые Osteichthyes (костистые рыбы): динамика разнообразия и эволюция размеров тела» . Биологические обзоры . 91 (1): 106–47. дои : 10.1111/brv.12161 . ПМИД 25431138 . S2CID 5332637 .
- ^ Кавин, Лайонел; Фуррер, Хайнц; Обрист, Кристиан (2013). «Новый материал целаканта из среднего триаса восточной Швейцарии и комментарии к тактическому разнообразию актинистанов» . Швейцарский журнал геонаук . 106 (2): 161–177. дои : 10.1007/s00015-013-0143-7 .
- ^ Романо, Карло (январь 2021 г.). «Перерыв скрывает раннюю эволюцию современных линий костистых рыб» . Границы в науках о Земле . 8 : 618853. дои : 10.3389/feart.2020.618853 .
- ^ Романо, Карло; Дженкс, Джеймс Ф.; Жаттио, Ромен; Шайер, Торстен М. (2017). «Морские актиноптеригии раннего триаса из округа Элко (Невада, США): последствия Смитовского экваториального затмения позвоночных» . Журнал палеонтологии . 91 (5): 1–22. Бибкод : 2017JPal...91.1025R . дои : 10.1017/jpa.2017.36 .
- ^ Агнолин, Флорида, Матеус О., Милан Дж., Марзола М., Уингс О., Адольфссен Дж.С. и Клемменсен Л.Б. (2018). Ceratodus tunuensis, sp. nov., новая двоякодышащая рыба (Sarcopterygii, Dipnoi) из верхнего триаса центральной части Восточной Гренландии. Журнал палеонтологии позвоночныхЖурнал палеонтологии позвоночных. e1439834
- ^ Кумар, Кришна; Баджпай, Сунил; Пандей, Прагья; Гош, Трипарна; Бхаттачарья, Дебашиш (4 августа 2021 г.). «Гибодонтные акулы из юрского периода Джайсалмера, западная Индия». Историческая биология . 34 (6): 953–963. дои : 10.1080/08912963.2021.1954920 . ISSN 0891-2963 . S2CID 238781606 .
- ^ Муттер, Рауль Дж.; Нойман, Эндрю Г. (2008). «Новые евгенеодонтидные акулы из формации серных гор нижнего триаса в Западной Канаде». Ин Кэвин, Л.; Лонгботтом, А.; Рихтер, М. (ред.). Рыбы и распад Пангеи . Лондонское геологическое общество, специальные публикации. Том. 295. Лондон: Лондонское геологическое общество. стр. 9–41. дои : 10.1144/sp295.3 . S2CID 130268582 .
- ^ Уэллс, Кентвуд Д. (2010). Экология и поведение амфибий . Издательство Чикагского университета . ISBN 978-0-226-89333-4 – через Google Книги.
- ^ Бентон, Майкл (2009). Палеонтология позвоночных . Джон Уайли и сыновья. ISBN 978-1-4051-4449-0 – через Google Книги.
- ^ Дуглас Палмер; Питер Барретт (2009). Эволюция: История жизни . Лондон: Музей естественной истории. ISBN 978-1-84533-339-3 .
- ^ Данн, Эмма М.; Фарнсворт, Александр; Бенсон, Роджер Б.Дж.; Годой, Педро Л.; Грин, Сара Э.; Вальдес, Пол Дж.; Лант, Дэниел Дж.; Батлер, Ричард Дж. (9 января 2023 г.). «Климатический контроль экологического господства динозавров» . Современная биология . 33 (1): 206–214. Бибкод : 2023CBio...33E.206D . дои : 10.1016/j.cub.2022.11.064 . hdl : 1983/aea1ae86-2260-4d4d-a9d5-0fe38a0f470e . ПМИД 36528026 . S2CID 254754419 .
- ^ Шейер и др. (2014): Восстановление морских биотиков в раннем триасе: взгляд хищников. PLoS ONE https://doi.org/10.1371/journal.pone.0088987
- ^ Рубен, Дж. А. и Джонс, Т. Д. (1 августа 2000 г.). «Селективные факторы, связанные с происхождением меха и перьев» . Интегративная и сравнительная биология . 40 (4): 585–96. дои : 10.1093/icb/40.4.585 . Проверено 24 ноября 2023 г.
- ^ Дай, Сюй, Джошуа ХФЛ; Брайард, Овчарова, Мария; Лю, Сяокан; Смит, Кэрри Э.; Цзян, Шоуи, Цао, Иран; Ван, Го, Вэй; Тянь, Ли; Лю, Юйтин; , Хайджун (2023 г.). экосистему современного . морскую » Мезозойская ископаемая лагерштетта , датированная 250,8 миллиона лет назад, Сонг типа ; « демонстрирует : 10.1126 /science.adf1622 PMID 36758082 S2CID 256697946 .
- ^ Брайард, Арно; Круменакер, LJ; Боттинг, Джозеф П.; Дженкс, Джеймс Ф.; Билунд, Кевин Г.; Фара, Эммануэль; Веннин, Эммануэль; Оливье, Николя; Гудеманд, Николя; Сосед, Томас; Шарбонье, Сильвен; Романо, Карло; Догужаева Лариса; Туи, Бен; Хаутманн, Майкл; Стивен, Дэниел А.; Томазо, Кристоф; Эскаргюэль, Жиль (2017). «Неожиданная морская экосистема раннего триаса и появление современной эволюционной фауны» . Достижения науки . 3 (2): e1602159. Бибкод : 2017SciA....3E2159B . дои : 10.1126/sciadv.1602159 . ПМК 5310825 . ПМИД 28246643 .
- ^ Nomade et al., 2007 Палеогеография, Палеоклиматология, Палеоэкология 244, 326–44.
- ^ Marzoli et al., 1999, Science 284. Обширные континентальные паводковые базальты возрастом 200 миллионов лет Центрально-Атлантической магматической провинции, стр. 618–620.
- ^ Ходыч и Даннинг, 1992.
- ^ «Нет значительного неморского карнийско-норианского (позднего триаса) вымирания: данные из национального парка Окаменелый лес» . gsa.confex.com . Архивировано из оригинала 6 ноября 2003 г. Проверено 12 декабря 2003 г.
Ссылки
[ редактировать ]- Эмилиани, Чезаре . (1992). Планета Земля: космология, геология, эволюция жизни и окружающей среды . Издательство Кембриджского университета. (Издание в мягкой обложке ISBN 0-521-40949-7 )
- Огг, Джим; Июнь 2004 г., Обзор разрезов и точек стратотипов глобальной границы (GSSP) Stratigraphy.org , по состоянию на 30 апреля 2006 г.
- Стэнли, Стивен М. История системы Земли. Нью-Йорк: WH Freeman and Company, 1999. ISBN 0-7167-2882-6
- Сьюс, Ханс-Дитер и Фрейзер, Николас К. Триасовая жизнь на суше: Великий переход Нью-Йорк: издательство Колумбийского университета, 2010. Серия: Критические моменты и перспективы в истории Земли и палеобиологии. ISBN 978-0-231-13522-1
- ван Андел, Тьерд, (1985) 1994, Новые взгляды на старую планету: история глобальных изменений , Cambridge University Press