Плиоцен
Эта статья нуждается в дополнительных цитатах для проверки . ( январь 2019 г. ) |
| Плиоцен | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Хронология | |||||||||||
| |||||||||||
| Этимология | |||||||||||
| Формальность имени | Формальный | ||||||||||
| Информация об использовании | |||||||||||
| Небесное тело | Земля | ||||||||||
| Региональное использование | Глобальный ( ICS ) | ||||||||||
| Используемая шкала времени | Временная шкала ICS | ||||||||||
| Определение | |||||||||||
| Хронологическая единица | Эпоха | ||||||||||
| Стратиграфическая единица | Ряд | ||||||||||
| Формальность временного интервала | Формальный | ||||||||||
| Определение нижней границы | База магнитного события Твера (C3n.4n), которое всего на 96 тыс. лет назад (5 циклов прецессии) моложе GSSP. | ||||||||||
| Нижняя граница ГССП | Раздел Гераклея Минойская, Гераклея Минойская , Каттолика Эраклея , Сицилия , Италия 37 ° 23'30 "N 13 ° 16'50" E / 37,3917 ° N 13,2806 ° E | ||||||||||
| Нижний GSSP ратифицирован | 2000 [4] | ||||||||||
| Определение верхней границы |
| ||||||||||
| Верхняя граница ГССП | Секция Монте-Сан-Никола, Джела , Сицилия, Италия 37 ° 08'49 "N 14 ° 12'13" E / 37,1469 ° N 14,2035 ° E | ||||||||||
| Верхний GSSP ратифицирован | 2009 г. (основа четвертичного периода и плейстоцена) [5] | ||||||||||

| Часть серии о |
| История человечества и предыстория |
|---|
| ↑ до Homo ( эпоха плиоцена ) |
| ↓ Будущее ( эпоха голоцена ) |
Плиоцен ( / ˈ p l aɪ . ə s iː n , ˈ p l aɪ . oʊ- / PLY -ə-seen, PLY -oh- ; [6] [7] также плейоцен ) [8] это эпоха в геологической шкале времени , которая простирается от 5,333 миллиона до 2,58 года. [9] миллион лет назад. Это вторая и самая поздняя эпоха неогенового периода кайнозойской эры . Плиоцен следует за эпохой миоцена , а за ней следует эпоха плейстоцена . До пересмотра геологической шкалы времени в 2009 году, согласно которому четыре последних крупных оледенения полностью относились к плейстоцену, плиоцен также включал гелазийский этап, который длился от 2,588 до 1,806 миллиона лет назад и теперь включен в плейстоцен. [10]
Как и в случае с другими более древними геологическими периодами, геологические пласты , определяющие начало и конец эпохи, хорошо идентифицированы, но точные даты начала и конца эпохи немного неопределенны. Границы, определяющие плиоцен, установлены не на каком-то легко определяемом всемирном событии, а скорее на региональных границах между более теплым миоценом и относительно более холодным плейстоценом. Верхняя граница была установлена в начале плейстоценовых оледенений.
Этимология [ править ]
Чарльз Лайель (позже сэр Чарльз) дал название плиоцену в «Принципах геологии» (том 3, 1833 г.). [11]
Слово плиоцен происходит от греческих слов πλεῖον ( плейон , «больше») и καινός ( каинос , «новый» или «недавний»). [12] и означает примерно «продолжение недавнего», имея в виду по сути современную фауну морских моллюсков .
Подразделения [ править ]
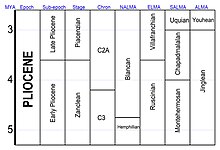
В официальной шкале времени ICS плиоцен подразделяется на два этапа . От младшего к старшему это:
Пьяченциан иногда называют поздним плиоценом, тогда как занклинский период называют ранним плиоценом.
В системе
- Возраст наземных млекопитающих Северной Америки (NALMA) включает гемфиллиан (9–4,75 млн лет назад), [14] [15] и Бланкан (4,75–1,6 млн лет назад). [16] Бланкан простирается вперед в плейстоцене .
- Возраст наземных млекопитающих Южной Америки (SALMA) включает монтегермосанский (6,8–4,0 млн лет назад), чападмалаланский (4,0–3,0 млн лет назад) и укианский (3,0–1,2 млн лет назад). [17]
В районе Паратетиса (центральная Европа и некоторые части западной Азии) плиоцен содержит дакийский (примерно равный занклинскому) и румынский (примерно равный пьяченцианскому и геласскому вместе взятым) этапам. Как обычно в стратиграфии, используется множество других региональных и местных подразделений.
В Британии плиоцен делится на следующие этапы (от старого к молодому): гедгравийский, вальтонский , прелудхемский, лудхемский, турнский, брамертонский или антианский, препастонский или бавентский, пастонский и бистонский . В Нидерландах плиоцен делится на следующие этапы (от старого к молодому): брюнсумский C, ревверский A, ревверский B, ревверский C, претильский , тильянский A, тильянский B, тильянский C1-4b, тильянский C4c, тильянский C5, тильянский C6 и Эбуронский . Точные корреляции между этими локальными этапами и этапами Международной комиссии по стратиграфии (ICS) все еще остаются вопросом деталей. [18]
Климат [ править ]

В эпоху плиоцена (от 5,3 до 2,6 миллионов лет назад (млн лет назад) климат Земли стал прохладнее и суше, а также стал более сезонным, отмечая переход от относительно теплого миоцена к более прохладному плейстоцену . [19] Начало плиоцена ознаменовалось повышением глобальных температур по сравнению с более холодным мессинским периодом, 1,2 миллиона лет что связано с циклом модуляции амплитуды наклона продолжительностью . [20] Средняя глобальная температура в середине плиоцена (3,3–3 млн лет назад) была на 2–3 ° C выше, чем сегодня. [21] уровень углекислого газа был таким же, как сегодня (400 частей на миллион), [22] а глобальный уровень моря был на 25 м выше. [23] Ледниковый щит северного полушария был эфемерным до начала обширного оледенения над Гренландией , которое произошло в позднем плиоцене около 3 млн лет назад. [24] Об образовании арктической ледяной шапки сигнализирует резкое изменение кислорода соотношения изотопов и наплавленные льдом булыжники в слоях Северной Атлантики и Северной части Тихого океана . [25] Оледенение в средних широтах, вероятно, продолжалось еще до конца эпохи. Глобальное похолодание, произошедшее в плиоцене, возможно, ускорилось из-за исчезновения лесов и распространения лугов и саванн. [26]
В течение плиоцена реакция климатической системы Земли изменилась от периода высокочастотных колебаний с низкой амплитудой, в котором преобладал 41 000-летний период наклона Земли , к периоду низкочастотных колебаний с высокой амплитудой, в котором преобладал 100 000-летний период орбитального движения . эксцентричность, характерная для плейстоценовых ледниково-межледниковых циклов. [27]
Во время позднеплиоценовой и ранней плейстоценовой серии кайнозойской эры, от 3,6 до 2,6 млн лет назад, Арктика была намного теплее, чем сейчас (летние температуры примерно на 8 ° C выше, чем сегодня). Это ключевой вывод исследования керна озерных отложений, полученного в Восточной Сибири, который имеет исключительное значение, поскольку на данный момент он предоставил самую длинную непрерывную наземную летопись позднекайнозойских осадочных пород. [28]
В плиоцене Центральная Азия стала более сезонной, с более холодной и сухой зимой и более влажным летом, что способствовало увеличению численности растений C 4 по всему региону. [29] На плато Лёссовом значения δ13C окклюдированного органического вещества увеличились на 2,5%, а значения педогенного карбоната увеличились на 5% в течение позднего миоцена и плиоцена, что указывает на усиление аридизации. [30] Дальнейшее засушивание Центральной Азии было вызвано развитием оледенения Северного полушария в позднем плиоцене. [31] Керн отложений из северной части Южно-Китайского моря показывает увеличение активности пыльных бурь в среднем плиоцене. [32]
В южно-центральных Андах засушливый период длился от 6,1 до 5,2 млн лет назад, а другой - от 3,6 до 3,3 млн лет назад. Эти засушливые периоды совпадают с глобальными холодными периодами, во время которых положение западных ветров Южного полушария сместилось к северу и нарушило южноамериканскую струю низкого уровня, которая приносит влагу на юго-восток Южной Америки. [33]
На северо-западе Африки во время Занклина тропические леса простирались до мыса Блан примерно до 3,5 млн лет назад. В период пьяченца, примерно с 3,5 до 2,6 млн лет назад, регион был засажен деревьями через неравные промежутки времени и содержал значительную сахарскую палеорику до 3,35 млн лет назад, когда пассаты начали доминировать над речным переносом пыльцы. Около 3,26 млн лет назад произошла сильная аридификация, за которой последовал возврат к более влажным условиям, за которым последовала еще одна аридификация около 2,7 млн лет назад. С 2,6 по 2,4 млн лет назад растительные зоны начали неоднократно смещаться по широте в ответ на ледниково-межледниковые циклы. [34]
Климат Восточной Африки был очень похож на современный. Неожиданно расширение лугов в Восточной Африке в эту эпоху, по-видимому, было связано с засушиванием, а не было вызвано им, о чем свидетельствует их асинхронность. [35]
В юго-западной Австралии в среднем и позднем плиоцене были пустоши , кустарники и леса с большим видовым разнообразием по сравнению с сегодняшним днем. Три различных события аридификации произошли около 2,90, 2,59 и 2,56 млн лет назад и, возможно, были связаны с началом континентального оледенения в Арктике, что позволяет предположить, что изменения растительности в Австралии в плиоцене вели себя так же, как и во время позднего плейстоцена, и, вероятно, характеризовались сопоставимые циклы засушливости и влажности. [36]
в экваториальной части Тихого океана Градиент температуры поверхности моря был значительно ниже, чем сегодня. Средние температуры поверхности моря на востоке были существенно выше, чем сегодня, но на западе были такими же. Это состояние было описано как постоянное состояние Эль-Ниньо или «Эль-Падре». [37] Было предложено несколько механизмов этой закономерности, включая усиление активности тропических циклонов . [38]
Протяженность Западно-Антарктического ледникового щита колебалась в течение 40- тысячного периода наклона Земли. Обрушение ледникового покрова произошло, когда средняя глобальная температура была на 3 °C выше, чем сегодня, а концентрация углекислого газа составляла 400 ppmv. Это привело к появлению открытых вод в море Росса . [39] Глобальное колебание уровня моря, связанное с разрушением ледникового покрова, вероятно, достигало 7 метров для западной Антарктики и 3 метров для восточной Антарктики. Модельные симуляции согласуются с реконструированными колебаниями ледникового покрова и предполагают переход от меньшего к большему ледниковому щиту Западной Антарктики за последние 5 миллионов лет. Интервалы разрушения ледникового покрова были гораздо более распространены в начале-среднем плиоцене (5–3 млн лет назад), после трехмиллионных интервалов с объемом современного или ледникового льда стали длиннее, и коллапс происходит только в те моменты, когда более высокие глобальные температуры совпадают с сильные аномалии инсоляции южного лета. [40]
Палеогеография [ править ]

Континенты продолжали дрейфовать , перемещаясь от позиций, находящихся, возможно, на расстоянии до 250 км от их нынешнего местоположения, к позициям всего в 70 км от их нынешнего местоположения. Южная Америка стала связана с Северной Америкой через Панамский перешеек во время плиоцена, что сделало возможным Великий американский обмен Южной Америки и почти полностью положило конец самобытной местной фауне копытных . [41] хотя другие южноамериканские линии, такие как хищные млекопитающие, к этому моменту уже вымерли, а другие, такие как ксенартраны, продолжали преуспевать и после этого. Образование перешейка имело серьезные последствия для глобальной температуры, поскольку теплые экваториальные океанские течения были прерваны и начался цикл атлантического охлаждения, при этом холодные арктические и антарктические воды снизили температуру в теперь отделенном Атлантическом океане. [42]
Африки В результате столкновения с Европой образовалось Средиземное море , отрезавшее остатки океана Тетис . Граница между миоценом и плиоценом является также временем Мессинского кризиса солености . [43] [44]
В позднем плиоцене Гималаи стали менее активными в своем поднятии, о чем свидетельствуют изменения осадконакопления в Бенгальском конусе . [45]
Сухопутный мост между Аляской и Сибирью ( Берингией ) впервые был затоплен в начале плиоцена, что позволило морским организмам распространиться между Северным Ледовитым и Тихим океанами. После этого мост будет периодически затапливать и восстанавливать. [46]
Плиоценовые морские образования обнажены на северо-востоке Испании . [47] южная Калифорния , [48] Новая Зеландия , [49] и Италия . [50]
В плиоцене части южной Норвегии и южной Швеции, которые находились вблизи уровня моря, поднялись. В Норвегии это возвышение подняло плато Хардангервидда на высоту 1200 м в раннем плиоцене. [51] В Южной Швеции аналогичные движения подняли южношведское нагорье , что привело к отклонению древней реки Эриданос от ее первоначального пути через юго-центральную Швецию к югу от Швеции. [52]
Окружающая среда и эволюция предков человека [ править ]
Плиоцен отмечен двумя важными событиями в эволюции предков человека. Первое — появление гоминина Australopithecus anamensis в раннем плиоцене, около 4,2 миллиона лет назад. [53] [54] [55] Второе — появление Homo , рода, включающего современных людей и их ближайших вымерших родственников, ближе к концу плиоцена, 2,6 миллиона лет назад. [56] Ключевые черты, которые развились среди гомининов в плиоцене, включают наземное двуногость и, к концу плиоцена, энцефализованный мозг (мозг с большим неокортексом по сравнению с массой тела). [57] [а] и производство каменных орудий. [58]
Усовершенствования в методах датирования и использовании климатических показателей предоставили ученым возможность проверить гипотезы эволюции предков человека. [58] [59] Ранние гипотезы эволюции человеческих качеств подчеркивали избирательное давление, создаваемое конкретными средами обитания. Например, многие учёные долгое время поддерживали гипотезу саванны . Это предполагает, что эволюция наземного двуногости и других черт была адаптивной реакцией на изменение климата в плиоцене, которое превратило леса в более открытые саванны . Эту теорию отстаивал Графтон Эллиот Смит в своей книге 1924 года « Эволюция человека разработал ее » как «неизвестный мир за деревьями», а Рэймонд Дарт как теорию обезьян-убийц . [60] Другие ученые, такие как Шервуд Л. Уошберн , подчеркивали внутреннюю модель эволюции гомининов. Согласно этой модели, раннее эволюционное развитие вызвало более позднее развитие. В модели мало внимания уделялось окружающей среде. [61] Антропологи, как правило, сосредотачивались на внутренних моделях, в то время как геологи и палеонтологи позвоночных, как правило, уделяли больше внимания местам обитания. [62]
Альтернативы гипотезе саванны включают гипотезу лесистости/леса, которая подчеркивает эволюцию гоминидов в закрытых средах обитания, или гипотезы, подчеркивающие влияние более холодных мест обитания в более высоких широтах или влияние сезонных колебаний. Более поздние исследования подчеркнули гипотезу отбора изменчивости, которая предполагает, что изменчивость климата способствовала развитию черт гомининов. [58] Улучшенные климатические прогнозы показывают, что плиоценовый климат Восточной Африки был сильно изменчивым, что позволяет предположить, что адаптация к изменяющимся условиям была более важна для эволюции гомининов, чем постоянное давление конкретной среды обитания. [57]
Flora[editФлора
Переход к более прохладному, сухому и сезонному климату оказал значительное воздействие на растительность плиоцена, сократив количество тропических видов во всем мире. Распространились лиственные леса, хвойные леса и тундра покрыли большую часть севера, а луга распространились на всех континентах (кроме Антарктиды). Тропические леса ограничились узкой полосой вокруг экватора, а помимо сухих в саванн пустыни . Азии и Африке появились [63] [ не удалось пройти проверку ]
Fauna[editфауна
И морская, и континентальная фауна были по существу современными, хотя континентальная фауна была немного более примитивной, чем сегодня.
Столкновения суши означали большую миграцию и смешение ранее изолированных видов, как, например, в Великом американском обмене . Травоядные животные стали крупнее, как и специализированные хищники.
Галерея изображений [ править ]


 Брюхоногие моллюски и прикрепленная червячная трубка серпулид из плиоцена Кипра.
Брюхоногие моллюски и прикрепленная червячная трубка серпулид из плиоцена Кипра. Брюхоногий моллюск Turritella carinata из плиоцена Кипра.
Брюхоногий моллюск Turritella carinata из плиоцена Кипра. Колючая устрица Spondylus, внутренняя часть правой и левой створки из плиоцена Кипра.
Колючая устрица Spondylus, внутренняя часть правой и левой створки из плиоцена Кипра. Блюдечко . Diodora italica из плиоцена Кипра
Блюдечко . Diodora italica из плиоцена Кипра
 Брюхоногий моллюск Aporrhais из плиоцена Кипра.
Брюхоногий моллюск Aporrhais из плиоцена Кипра. Дуговой двустворчатый моллюск Анадара из плиоцена Кипра.
Дуговой двустворчатый моллюск Анадара из плиоцена Кипра. Пектенидный двустворчатый моллюск Ammusium cristatum из плиоцена Кипра.
Пектенидный двустворчатый моллюск Ammusium cristatum из плиоцена Кипра. Верметидный брюхоногий моллюск Petaloconchus intortus, прикрепленный к ветке коралла Cladocora из плиоцена Кипра.
Верметидный брюхоногий моллюск Petaloconchus intortus, прикрепленный к ветке коралла Cladocora из плиоцена Кипра.












Млекопитающие [ править ]
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||

В Северной Америке численность грызунов , крупных мастодонтов , гомфотерий и опоссумов успешно сохранялась, в то время как численность копытных ( копытных ) сокращалась, а верблюдов , оленей и лошадей популяции трехпалые лошади ( Nannippus ), ореодонты , протоцератиды и халикотерии сокращались. Вымерли . Собаки-борофаги и агриотерии вымерли, но другие хищники , включая семейство ласки, диверсифицировались, а собаки и короткомордые медведи преуспели. Наземные ленивцы , огромные глиптодонты и броненосцы пришли на север с образованием Панамского перешейка.
В Евразии грызуны преуспели, тогда как распространение приматов сократилось. В Азии успехами пользовались слоны , гомфотерии и стегодонты (крупнейшими наземными млекопитающими плиоцена были такие хоботные, как Deinotherium , Anancus и Mammut borsoni) . [64] ), а даманы мигрировали на север из Африки. Разнообразие лошадей сократилось, в то время как тапиры и носороги преуспели. Крупный рогатый скот и антилопы добились успеха; некоторые виды верблюдов проникли в Азию из Северной Америки. Появились гиены и ранние саблезубые кошки , присоединившиеся к другим хищникам, включая собак, медведей и ласк.
 |
В Африке доминировали копытные животные, и приматы продолжили свою эволюцию: австралопитеки (некоторые из первых гомининов ) и павионоподобные обезьяны, такие как динопитек в позднем плиоцене появились . Грызуны добились успеха, и популяция слонов увеличилась. Коровы и антилопы продолжили диверсификацию и обогнали свиней по количеству видов. Появились первые жирафы . На сцену вышли лошади и современные носороги. Медведи, собаки и ласки (родом из Северной Америки) присоединились к кошкам, гиенам и циветам в качестве африканских хищников, вынудив гиен адаптироваться в качестве специализированных падальщиков. Число куньих в Африке сократилось в результате усиления конкуренции со стороны новых хищников, хотя Enhydriodon omoensis оставался необычайно успешным наземным хищником.
Южная Америка была заселена североамериканскими видами впервые со времен мелового периода , при этом североамериканские грызуны и приматы смешались с южными формами. Литоптерны и нотонгулаты , аборигены Южной Америки, были в основном истреблены, за исключением макраухенид и токсодонтов , которым удалось выжить. мелкие ласкообразные плотоядные куньи , коати и короткомордые медведи С севера мигрировали . Пасущиеся глиптодонты , гигантские наземные ленивцы и более мелкие кавиоморфные грызуны , пампатеры и броненосцы делали противоположное, мигрируя на север и процветая там.
Сумчатые оставались доминирующими австралийскими млекопитающими, с травоядными формами, включая вомбатов и кенгуру , а также огромного дипротодона . В плиоцене продолжали охотиться плотоядные сумчатые, в том числе дасюриды , собачьи тилацины и кошачьи тилаколео . Первые грызуны прибыли в Австралию. Появился современный утконос однопроходной — .
Птицы [ править ]

Хищные южноамериканские форусрациды в то время были редкостью; Среди последних был Titanis , крупный форусрацид, который мигрировал в Северную Америку и соперничал с млекопитающими в качестве главного хищника. В это время, вероятно, появились и другие птицы: некоторые современные (такие как роды Cygnus , Bubo , Struthio и Corvus ), некоторые ныне вымершие.
Рептилии и земноводные [ править ]
Аллигаторы и крокодилы вымерли в Европе из-за похолодания климата. Роды ядовитых змей продолжали увеличиваться по мере развития большего количества грызунов и птиц. Гремучие змеи впервые появились в плиоцене. Современный вид Alligator Mississippiensis , возникший в миоцене, продолжил свое существование и в плиоцене, за исключением более северного ареала; образцы были найдены в отложениях очень позднего миоцена в Теннесси . Гигантские черепахи все еще процветали в Северной Америке, и их роды были такими, как Hesperotestudo . Мадцоид Змеи все еще обитали в Австралии. Отряд земноводных Allocaudata вымер.
Двустворчатые моллюски [ править ]
В Западной Атлантике сообщества двустворчатых моллюсков демонстрировали удивительный застой в отношении скорости основного метаболизма на протяжении различных климатических изменений плиоцена. [65]
Кораллы [ править ]
Плиоцен был высшей точкой видового разнообразия карибских кораллов. В период с 5 по 2 млн лет назад темпы возникновения видов кораллов в Карибском бассейне были относительно высокими, хотя в конце этого интервала произошло заметное вымирание и снижение разнообразия. [66]
Океаны [ править ]
Эта статья нуждается в дополнительных цитатах для проверки . ( май 2021 г. ) |
В плиоцене океаны продолжали быть относительно теплыми, хотя и продолжали охлаждаться. , Образовалась арктическая ледяная шапка высушившая климат и увеличившая прохладные мелкие течения в Северной Атлантике. Глубокие холодные течения текли из Антарктики.
Образование Панамского перешейка около 3,5 миллионов лет назад. [67] отрезали последний остаток того, что когда-то было по сути околоэкваториальным течением, существовавшим с мелового периода и раннего кайнозоя . Возможно, это способствовало дальнейшему охлаждению океанов во всем мире.
Моря плиоцена были полны морских коров , тюленей , морских львов , акул и китов .
См. также [ править ]
- Список мест окаменелостей (с каталогом ссылок)
Примечания [ править ]
- ^ Из-за переноса границы плиоцена и плейстоцена в 2009 году с 1,8 на 2,6 миллиона лет назад, более старые статьи об эволюции плиоценовых гомининов иногда включают события, которые теперь можно было бы рассматривать как происходящие в раннем плейстоцене.
Ссылки [ править ]
- ^ Крийгсман, В.; Гарсес, М.; Ланжерайс, К.Г.; Даамс, Р.; Ван Дам, Дж.; Ван дер Мюлен, AJ; Агусти, Дж.; Кабрера, Л. (1996). «Новая хронология континентальных рекордов среднего и позднего миоцена в Испании». Письма о Земле и планетологии . 142 (3–4): 367–380. Бибкод : 1996E&PSL.142..367K . дои : 10.1016/0012-821X(96)00109-4 .
- ^ Реталлак, Дж.Дж. (1997). «Неогеновое расширение североамериканских прерий» . ПАЛЕОС . 12 (4): 380–390. дои : 10.2307/3515337 . JSTOR 3515337 . Проверено 11 февраля 2008 г.
- ^ «Временная шкала ICS» (PDF) . www.stratigraphy.org .
- ^ Перейти обратно: а б Ван Куверинг, Джон; Кастрадори, Давиде; Сита, Мария; Хильген, Фредерик; Рио, Доменико (сентябрь 2000 г.). «Основание занклинского яруса и плиоценовой серии» (PDF) . Эпизоды . 23 (3): 179–187. дои : 10.18814/epiiugs/2000/v23i3/005 .
- ^ Гиббард, Филип; Руководитель Мартин (сентябрь 2010 г.). «Недавно ратифицированное определение четвертичной системы/периода и новое определение плейстоценовой серии/эпохи, а также сравнение предложений, выдвинутых до официальной ратификации» (PDF) . Эпизоды . 33 (3): 152–158. дои : 10.18814/epiiugs/2010/v33i3/002 . Проверено 8 декабря 2020 г.
- ^ «Плиоцен» . Словарь Merriam-Webster.com .
- ^ «Плиоцен» . Dictionary.com Полный (онлайн). nd
- ^ «Плейоцен» . Dictionary.com Полный (онлайн). nd
- ^ См. версию геологической шкалы времени ICS за 2014 г. Архивировано 30 мая 2014 г. на Wayback Machine.
- ^ Огг, Джеймс Джордж; Огг, Габи; Градштейн Ф.М. (2008). Краткая геологическая шкала времени . Издательство Кембриджского университета. стр. 150–1. ISBN 9780521898492 .
- ^ См.:
- Письмо Уильяма Уэвелла Чарльзу Лайелю от 31 января 1831 г.: Тодхантер, Исаак, изд. (1876). Уильям Уэвелл, доктор медицинских наук, магистр Тринити-колледжа, Кембридж: отчет о его произведениях с отрывками из его литературной и научной переписки . Том. 2. Лондон, Англия: Macmillan and Co. p. 111.
- Лайель, Чарльз (1833). Принципы геологии, … . Том. 3. Лондон, Англия: Джон Мюррей. п. 53. Со с. 53: «Мы получаем термин «плиоцен» от πλειων, «мажор», и χαινος, «recens», поскольку большая часть ископаемых панцирей этой эпохи относится к современным видам*.
- ^ «Плиоцен» . Интернет-словарь этимологии .
- ^ Кастрадори, Д.; Рио, Д.; Хильген, Ф.Дж.; Лоренс, ЖЖ (1998). «Глобальный стандартный стратотип-разрез и точка (GSSP) пиаченцианского яруса (средний плиоцен)» . Эпизоды . 21 (2): 88–93. дои : 10.18814/epiiugs/1998/v21i2/003 .
- ^ Тедфорд, Ричард Х.; Олбрайт, Л. Барри; Барноски, Энтони Д.; Феррускиа-Виллафранка, Исмаэль; Хант, Роберт М.; Сторер, Джон Э.; Свишер, Карл С.; Вурхис, Майкл Р.; Уэбб, С. Дэвид; Уистлер, Дэвид П. (31 декабря 2004 г.). «6. Биохронология млекопитающих от арикарея до гемфиллийского интервала (от позднего олигоцена до раннего плиоцена)». Позднемеловые и кайнозойские млекопитающие Северной Америки : 169–231. дои : 10.7312/wood13040-008 . ISBN 9780231130400 .
- ^ Халберт, Ричард С. младший (2 августа 2016 г.). «Гемфиллианский возраст наземных млекопитающих Северной Америки» . Ископаемые виды Флориды . Музей Флориды . Проверено 7 июня 2021 г.
- ^ Халберт, Ричард С. младший (2 августа 2016 г.). «Бланканский возраст наземных млекопитающих Северной Америки» . Ископаемые виды Флориды . Музей Флориды . Проверено 7 июня 2021 г.
- ^ Флинн, Дж.; Свишер, CC III (1995). «Кайнозойский возраст наземных млекопитающих Южной Америки: корреляция с глобальной геохронологией». У Уильяма А. Берггрена; Деннис В. Кент; Мари-Пьер Обри; Ян Харденбол (ред.). Геохронологические временные шкалы и глобальная стратиграфическая корреляция . Общество осадочной геологии. стр. 317–333. дои : 10.2110/pec.95.04.0317 .
- ^ Кульманн, Г.; К.Г. Ланжерайс; Д. Мюнстерман; Р.-Ж. ван Леувен; Р. Веррейссель; Дж. Э. Меленкамп; Т.Э. Вонг (2006). «Комплексная хроностратиграфия плиоцен-плейстоценового интервала и ее связь с региональными стратиграфическими этапами в южной части Северного моря» (PDF) . Нидерландский журнал геонаук . 85 (1): 19–35. Бибкод : 2006NJGeo..85...19K . дои : 10.1017/S0016774600021405 . S2CID 62803118 .
- ^ Фокетт, Северин; Бертини, Адель (28 июня 2008 г.). «Количественная оценка климата плиоцена северной Италии по данным пыльцы: свидетельства очень своеобразного климата» . Борей . 32 (2): 361–369. дои : 10.1111/j.1502-3885.2003.tb01090.x .
- ^ Цинь, Цзе; Чжан, Руй; Кравчинский Вадим А.; Вале, Жан-Пьер; Саньотти, Леонардо; Ли, Цзяньсин; Сюй, Юн; Анвар, Таслима; Юэ, Лепинг (2 апреля 2022 г.). «1.2 Мировой диапазон модуляции наклона Земли и Марса в эволюции холодного позднего миоцена к теплому раннему плиоцену» . Твердая Земля . 127 (4). Бибкод : 2022JGRB..12724131Q . дои : 10.1029/2022JB024131 . S2CID 247933545 . Проверено 24 ноября 2022 г.
- ^ Робинсон, М.; Доусетт, HJ; Чендлер, Массачусетс (2008). «Роль плиоцена в оценке будущих воздействий на климат». Эос, Транзакции, Американский геофизический союз . 89 (49): 501–502. Бибкод : 2008EOSTr..89..501R . дои : 10.1029/2008eo490001 .
- ^ «Решения: реагирование на изменение климата» . Climate.Nasa.gov . Проверено 1 сентября 2016 г.
- ^ Дуайер, Г.С.; Чендлер, Массачусетс (2009). «Уровень моря в середине плиоцена и объем континентального льда на основе связанных палеотемператур бентоса Mg / Ca и изотопов кислорода». Фил. Пер. Р. Сок. А. 367 (1886): 157–168. Бибкод : 2009RSPTA.367..157D . дои : 10.1098/rsta.2008.0222 . hdl : 10161/6586 . ПМИД 18854304 . S2CID 3199617 .
- ^ Бартоли, Г.; и др. (2005). «Окончательное закрытие Панамы и начало оледенения северного полушария» . Планета Земля. наук. Летт . 237 (1–2): 3344. Бибкод : 2005E&PSL.237...33B . дои : 10.1016/j.epsl.2005.06.020 .
- ^ Ван Андел (1994), с. 226.
- ^ «Эпоха плиоцена» . Музей палеонтологии Калифорнийского университета . Проверено 25 марта 2008 г.
- ^ Доусетт, HJ; Чендлер, Массачусетс; Кронин, ТМ; Дуайер, Г.С. (2005). «Изменчивость температуры поверхности моря в среднем плиоцене» (PDF) . Палеоокеанография . 20 (2): ПА2014. Бибкод : 2005PalOc..20.2014D . CiteSeerX 10.1.1.856.1776 . дои : 10.1029/2005PA001133 . Архивировано из оригинала (PDF) 22 октября 2011 года.
- ^ Мейсон, Джон. «В последний раз концентрация углекислого газа составляла около 400 частей на миллион: снимок из арктической Сибири» . Скептическая наука . Проверено 30 января 2014 г.
- ^ Шен, Синъянь; Ван, Шиминг; Колин, Кристоф; Тада, Рюдзи; Ши, Сюэфа; Пей, Вэньцян; Тан, Ян; Цзян, Сюэцзюнь; Ли, Аньчунь (15 ноября 2018 г.). «Повышенная сезонность и засушливость привели к расширению производства C4 в Центральной Азии после границы миоцена и плиоцена» . Письма о Земле и планетологии . 502 : 74–83. Бибкод : 2018E&PSL.502...74S . дои : 10.1016/j.epsl.2018.08.056 . S2CID 134183141 . Проверено 1 января 2023 г.
- ^ Галлахер, Тимоти М.; Серах, Лили; Сехон, Наташа; Чжан, Ханжи; Ван, Ханлинь; Цзи, Шуньчуань; Чанг, Си; Лу, Хуаюй; Брикер, Дэниел О. (25 ноября 2021 г.). «Региональные закономерности миоцен-плиоценовой засушливости на китайском лессовом плато, выявленные с помощью записей палеопочвенного карбоната и окклюдированного органического вещества с высоким разрешением» . Палеоокеанография и палеоклиматология . 32 (12). Бибкод : 2021PaPa...36.4344G . дои : 10.1029/2021PA004344 . S2CID 244702210 . Проверено 1 января 2023 г.
- ^ Сан, Юбин; Ань, Чжишэн (1 декабря 2005 г.). «Позднеплиоцен-плейстоценовые изменения темпов массового накопления эоловых отложений на Центрально-Китайском Лёссовом плато» . Журнал геофизических исследований . 110 (Д23): 1–8. Бибкод : 2005JGRD..11023101S . дои : 10.1029/2005JD006064 .
- ^ Сюфке, финн; Кабот-Барр, Стефани; Вэй, Го-Йен; Чуанг, Чи-Кай; Гутжар, Маркус; Просс, Йорг; Фридрих, Оливер (15 сентября 2022 г.). «Усиление азиатских пыльных бурь во время теплого периода среднего плиоцена (3,25–2,96 млн лет назад), зафиксированное в керне отложений Южно-Китайского моря» . Четвертичные научные обзоры . 292 . Бибкод : 2022QSRv..29207669S . doi : 10.1016/j.quascirev.2022.107669 . S2CID 251426879 . Проверено 25 июня 2023 г.
- ^ Амидон, Уильям Х.; Фишер, Г. Берч; Бербанк, Дуглас В.; Чиччоли, Патрисия Л.; Алонсо, Рикардо Н.; Горин, Андрей Л.; Сильверхарт, Перри Х.; Киландер-Кларк, Эндрю Р.К.; Кристофферсен, Майкл С. (12 июня 2017 г.). «Мио-плиоценовая засушливость в южно-центральных Андах, связанная с холодными периодами в южном полушарии» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (25): 6474–6479. Бибкод : 2017PNAS..114.6474A . дои : 10.1073/pnas.1700327114 . ПМЦ 5488932 . ПМИД 28607045 .
- ^ Лерой, Сюзанна; Дюпон, Лиди (июнь 1994 г.). «Развитие растительности и континентальной засушливости в северо-западной Африке в позднем плиоцене: запись пыльцы на участке ODP 658» . Палеогеография, Палеоклиматология, Палеоэкология . 109 (2–4): 295–316. Бибкод : 1994PPP...109..295L . дои : 10.1016/0031-0182(94)90181-3 . Проверено 31 декабря 2022 г.
- ^ Блюменталь, Скотт А.; Левин, Наоми Э.; Браун, Фрэнсис Х.; Бругаль, Жан-Филип; Хритц, Кендра Л.; Харрис, Джон М.; Йеле, Глинис Э.; Серлинг, Туре Э. (26 июня 2017 г.). «Засушливость и среда обитания человека» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (28): 7331–7336. Бибкод : 2017PNAS..114.7331B . дои : 10.1073/pnas.1700597114 . ПМК 5514716 . ПМИД 28652366 .
- ^ Додсон-младший; Макфейл, МК (июль 2004 г.). «Палинологические свидетельства засушливости и изменения растительности во время среднего плиоцена, теплого периода на юго-западе Австралии» . Глобальные и планетарные изменения . 41 (3–4): 285–307. Бибкод : 2004GPC....41..285D . дои : 10.1016/j.gloplacha.2004.01.013 . Проверено 31 декабря 2022 г.
- ^ Федоров А.В.; и др. (2006). «Парадокс плиоцена (механизмы постоянного Эль-Ниньо)». Наука . 312 (5779): 1485–1489. Бибкод : 2006Sci...312.1485F . CiteSeerX 10.1.1.143.5772 . дои : 10.1126/science.1122666 . ПМИД 16763140 . S2CID 36446661 .
- ^ Федоров Алексей Владимирович; Бриерли, Кристофер М.; Эмануэль, Керри (февраль 2010 г.). «Тропические циклоны и постоянное Эль-Ниньо в эпоху раннего плиоцена». Природа . 463 (7284): 1066–1070. Бибкод : 2010Natur.463.1066F . дои : 10.1038/nature08831 . hdl : 1721.1/63099 . ISSN 0028-0836 . ПМИД 20182509 . S2CID 4330367 .
- ^ Нэйш, Т.; и др. (2009). «Наклонные плиоценовые колебания ледникового покрова Западной Антарктики» . Природа . 458 (7236): 322–328. Бибкод : 2009Natur.458..322N . дои : 10.1038/nature07867 . ПМИД 19295607 . S2CID 15213187 .
- ^ Поллард, Д.; ДеКонто, РМ (2009). «Моделирование роста и разрушения ледникового покрова Западной Антарктики за последние пять миллионов лет». Природа . 458 (7236): 329–332. Бибкод : 2009Natur.458..329P . дои : 10.1038/nature07809 . ПМИД 19295608 . S2CID 4427715 .
- ^ Уэбб, С. Дэвид (1991). «Экогеография и великий американский обмен». Палеобиология . 17 (3): 266–280. Бибкод : 1991Pbio...17..266W . дои : 10.1017/S0094837300010605 . JSTOR 2400869 . S2CID 88305955 .
- ^ Бартоли, Г.; Сарнтейн, М.; Вайнельт, М.; Эрленкейзер, Х.; Гарбе-Шенберг, Д.; Леа, Д.В. (август 2005 г.). «Окончательное закрытие Панамы и начало оледенения северного полушария» . Письма о Земле и планетологии . 237 (1–2): 33–44. Бибкод : 2005E&PSL.237...33B . дои : 10.1016/j.epsl.2005.06.020 .
- ^ Готье Ф., Клаузон Г., Сюк Дж. П., Краватт Дж., Виоланти Д., 1994. Возраст и продолжительность Мессинского кризиса солености. ЧР акад. Sci., Париж (IIA) 318, 1103–1109.
- ^ Крийгсман, В. (август 1996 г.). «Новая хронология континентальных рекордов среднего и позднего миоцена в Испании» (PDF) . Письма о Земле и планетологии . 142 (3–4): 367–380. Бибкод : 1996E&PSL.142..367K . дои : 10.1016/0012-821X(96)00109-4 .
- ^ Чанг, Зихан; Чжоу, Липин (декабрь 2019 г.). «Доказательства изменения происхождения глубоководных отложений Бенгальского конуса: запись IODP U1444A за 7 миллионов лет» . Журнал азиатских наук о Земле . 186 . Бибкод : 2019JAESc.18604008C . дои : 10.1016/j.jseaes.2019.104008 . S2CID 202902163 . Проверено 6 июля 2023 г.
- ^ Гладенков Андрей Ю; Олейник, Антон Е; Маринкович, Луи; Баринов, Константин Б (июль 2002 г.). «Утонченный возраст самого раннего открытия Берингова пролива» . Палеогеография, Палеоклиматология, Палеоэкология . 183 (3–4): 321–328. Бибкод : 2002PPP...183..321G . дои : 10.1016/S0031-0182(02)00249-3 . Проверено 6 июля 2023 г.
- ^ Жиберт, Хорди Мария де; Мартинелл, Хорди (январь 1995 г.). «Осадочный субстрат и следы ископаемых комплексов в морских плиоценовых отложениях на северо-востоке Испании». Геобиос . 28 : 197–206. Бибкод : 1995Geobi..28R.197G . дои : 10.1016/S0016-6995(95)80166-9 .
- ^ Демере, Томас А. (1983). «Неогеновый бассейн Сан-Диего: обзор морской плиоценовой формации Сан-Диего». Кайнозойская морская седиментация, Тихоокеанская окраина . Тихоокеанское отделение Общества осадочной геологии. стр. 187–195 . Проверено 7 июня 2021 г.
- ^ Саул, Г.; Наиш, ТР; Эбботт, Северная Каролина; Картер, РМ (1 апреля 1999 г.). «Осадочная цикличность в морском плиоцене-плейстоцене бассейна Вангануи (Новая Зеландия): последовательность стратиграфических мотивов, характерных для последних 2,5 млн лет назад» Бюллетень Геологического общества Америки . 111 (4): 524–537. Бибкод : 1999GSAB..111..524S . doi : 10.1130/0016-7606(1999)111<0524:SCITMP>2.3.CO;2 . Проверено 6 июля 2023 г.
- ^ Селли, Раймондо (сентябрь 1965 г.). «Граница плиоцена и плейстоцена в морских разрезах Италии и ее связь с континентальной стратиграфией». Прогресс в океанографии . 4 : 67–86. Бибкод : 1965Proce...4...67S . дои : 10.1016/0079-6611(65)90041-8 .
- ^ Япсен, Питер; Грин, Пол Ф.; Чалмерс, Джеймс А.; Бонов, Йохан М. (17 мая 2018 г.). «Горы самой южной Норвегии: поднятые миоценовые пенеплены и вновь обнаженные мезозойские поверхности» . Журнал Геологического общества . 175 (5): 721–741. Бибкод : 2018JGSoc.175..721J . дои : 10.1144/jgs2017-157 . S2CID 134575021 .
- ^ Лидмар-Бергстрем, Карна ; Ольвмо, Матс; Боноу, Йохан М. (2017). «Южно-Шведский купол: ключевая структура для идентификации пенеплена и выводов о фанерозойской тектонике древнего щита» . ГФФ . 139 (4): 244–259. Бибкод : 2017GFF...139..244L . дои : 10.1080/11035897.2017.1364293 . S2CID 134300755 .
- ^ Розе, Симон (28 августа 2019 г.). «Самый древний ископаемый череп австралопитека обнаружен в Эфиопии» . RFI (на французском языке) . Проверено 18 февраля 2023 г.
- ^ Хайле-Селассие, Ю (27 октября 2010 г.). «Филогения ранних австралопитеков: новые ископаемые свидетельства из Ворансо-Милле (центральный Афар, Эфиопия)» . Философские труды Королевского общества B: Биологические науки . 365 (1556): 3323–3331. дои : 10.1098/rstb.2010.0064 . ПМК 2981958 . ПМИД 20855306 .
- ^ Льюис, Барри; и др. (2013). Понимание людей: введение в физическую антропологию и археологию (11-е изд.). Бельмонт, Калифорния: Издательство Wadsworth.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Стрингер CB (1994). «Эволюция первых людей». В Джонс С., Мартин Р., Пилбим Д. (ред.). Кембриджская энциклопедия эволюции человека . Кембридж: Издательство Кембриджского университета. п. 242.
- ^ Перейти обратно: а б Поттс, Р. (2007), Бобе, Рене; Алемсегед, Зересенай; Беренсмейер, Анна К. (ред.), «Экологические гипотезы эволюции человека в плиоцене», Среда обитания гомининов в восточноафриканском плиоцене: оценка фаунистических данных , серия палеобиологии и палеоантропологии позвоночных, Дордрехт: Springer Нидерланды, стр. 25–49 , doi : 10.1007/978-1-4020-3098-7_2 , ISBN 978-1-4020-3098-7
- ^ Перейти обратно: а б с Поттс, Ричард (3 января 1999 г.). «Экологические гипотезы эволюции человекообразных». Американский журнал биологической антропологии . 107 (С27): 93–136. doi : 10.1002/(SICI)1096-8644(1998)107:27+<93::AID-AJPA5>3.0.CO;2-X . ПМИД 9881524 .
- ^ деМенокаль, Питер Б. (3 марта 2004 г.). «Изменение климата Африки и эволюция фауны в плиоцене – плейстоцене» . Письма о Земле и планетологии . 220 (1): 3–24. Бибкод : 2004E&PSL.220....3D . дои : 10.1016/S0012-821X(04)00003-2 . ISSN 0012-821X .
- ^ Дарт, Р. (1953). «Хищный переход от обезьяны к человеку». Межд. Антроп. Линг. Преподобный . 1 : 201–218.
- ^ Уошберн, СЛ (1960). «Инструменты и эволюция человека». наук. Являюсь . 203 (3): 63–75. Бибкод : 1960SciAm.203c..62W . doi : 10.1038/scientificamerican0960-62 . JSTOR 24940615 . ПМИД 13843002 .
- ^ Поттс 1999 , стр. 106–108.
- ^ Марес, Майкл А., изд. (1999). «Миоцен». Энциклопедия пустынь . Университет Оклахомы Пресс. ISBN 0-8061-3146-2 .
- ^ Чарльз А. Репеннинг, Ричард Х. Тедфорд (2013). Окаменелости Карпатского региона . Издательство Университета Индианы. п. 373. ИСБН 9780253009876 . Проверено 23 августа 2022 г.
- ^ Строц, Люк К.; Саупе, Эрин Э.; Киммиг, Жюльен; Либерман, Брюс С. (29 августа 2018 г.). «Скорость метаболизма, климат и макроэволюция: тематическое исследование с использованием моллюсков неогена» . Труды Королевского общества B: Биологические науки . 285 (1885): 20181292. doi : 10.1098/rspb.2018.1292 . ISSN 0962-8452 . ПМК 6125889 . ПМИД 30135165 . Проверено 1 мая 2024 г.
- ^ Бадд, AF (апрель 2000 г.). «Разнообразие и вымирание в кайнозойской истории карибских рифов» . Коралловые рифы . 19 :25–35. дои : 10.1007/s003380050222 . S2CID 1062406 . Проверено 10 мая 2023 г.
- ^ Кейгвин, Ллойд Д. (1 октября 1978 г.). «Плиоценовое закрытие Панамского перешейка на основе биостратиграфических данных из близлежащих кернов Тихого океана и Карибского моря» . Геология . 6 (10): 630–634. Бибкод : 1978Geo.....6..630K . doi : 10.1130/0091-7613(1978)6<630:PCOTIO>2.0.CO;2 . ISSN 0091-7613 .
Дальнейшее чтение [ править ]
- Коминс, Нил Ф.; Уильям Дж. Кауфманн III (2005). Открытие Вселенной (7-е изд.). Нью-Йорк, штат Нью-Йорк: Сьюзен Финнемор Бреннан. ISBN 978-0-7167-7584-3 .
- Градштейн, FM; Огг, Дж.Г. и Смит, АГ ; 2004 : Геологическая шкала времени 2004 , Издательство Кембриджского университета .
- Огг, Джим (июнь 2004 г.). «Обзор разрезов и точек стратотипов глобальной границы (GSSP)» . Архивировано из оригинала 23 апреля 2006 года . Проверено 30 апреля 2006 г.
- Ван Андел, Тьерд Х. (1994). Новые взгляды на старую планету: история глобальных изменений (2-е изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-44243-5 .
Внешние ссылки [ править ]
- Глобальное потепление в середине плиоцена: моделирование климата НАСА/GISS
- Палеос-плиоцен
- Изменение PBS: Глубокое время: плиоцен
- Возможная сверхновая плиоцена
- «Сверхновая звезда привела к гибели людей на Земле? Звездные взрывы, возможно, убили древнюю морскую жизнь» Science News Online, получено 2 февраля 2002 г.
- Страница UCMP Беркли, эпоха плиоцена
- Плиоценовые микрофоссилии: более 100 изображений плиоценовых фораминифер
- Хронология человечества (интерактивная) – Смитсоновский институт , Национальный музей естественной истории (август 2016 г.).