Коралл
| Кораллы Временной диапазон: | |
|---|---|
 | |
| Обнажение кораллов на Большом Барьерном рифе , Австралия. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Книдарийцы |
| Подтип: | Антозоа Эренберг , 1834 г. |
| Подразделения | |
Кораллы колониальные морские класса Anthozoa — типа Cnidaria . беспозвоночные Обычно они образуют компактные колонии из множества идентичных отдельных полипов . К видам кораллов относятся важные строители рифов , населяющие тропические океаны и выделяющие карбонат кальция, образующий твердый скелет.
Коралловая «группа» представляет собой колонию очень многих генетически идентичных полипов. Каждый полип представляет собой мешковидное животное, обычно всего несколько миллиметров в диаметре и несколько сантиметров в высоту. набор щупалец Центральное ротовое отверстие окружает . Каждый полип выделяет экзоскелет возле основания. Таким образом, на протяжении многих поколений колония создает характерный для этого вида скелет, размер которого может достигать нескольких метров. Отдельные колонии растут путем бесполого размножения полипов. Кораллы также размножаются половым путем, нерестясь : полипы одного и того же вида выпускают гаметы одновременно в течение ночи, часто около полнолуния . Оплодотворенные яйца образуют планулы, подвижную раннюю форму кораллового полипа, которая, когда созревает, оседает, образуя новую колонию.
Хотя некоторые кораллы способны ловить планктон и мелкую рыбу , используя стрекательные клетки на своих щупальцах, большинство кораллов получают большую часть своей энергии и питательных веществ от фотосинтетических одноклеточных динофлагеллят рода Symbiodinium , которые живут в их тканях. Они широко известны как зооксантеллы и придают коралловый цвет. Такие кораллы требуют солнечного света и растут в прозрачной мелкой воде, обычно на глубине менее 60 метров (200 футов; 33 сажени), но кораллы рода Leptoseris были обнаружены на глубине до 172 метров (564 фута; 94 сажени). [1] Кораллы вносят основной вклад в физическую структуру коралловых рифов , которые развиваются в тропических и субтропических водах, таких как Большой Барьерный риф у побережья Австралии . Эти кораллы все чаще подвергаются риску обесцвечивания , когда полипы выделяют зооксантеллы в ответ на стресс, такой как высокая температура воды или токсины.
Другие кораллы не зависят от зооксантелл и могут жить по всему миру в гораздо более глубоких водах, например, холодноводный род Lophelia , который может выжить на глубине до 3300 метров (10 800 футов; 1800 саженей). [2] Некоторые из них были найдены на севере, вплоть до Дарвиновских курганов , к северо-западу от мыса Гнева в Шотландии, а другие — у побережья штата Вашингтон и Алеутских островов .
Таксономия
[ редактировать ]Классификация кораллов обсуждалась на протяжении тысячелетий из-за их сходства как с растениями, так и с животными. Теофраст Ученик Аристотеля описал красный коралл , кораллион , в своей книге о камнях, подразумевая, что это минерал, но он описал его как глубоководное растение в своих «Исследованиях о растениях» , где он также упоминает крупные каменистые растения, обнаруживающие яркие цветы под водой в Заливе Героев . [3] Плиний Старший смело заявил, что некоторые морские существа, включая морскую крапиву и губки, «не являются ни животными, ни растениями, а обладают третьей природой ( tertia natura )». [4] Петр Гиллий скопировал Плиния, введя термин Zoophyta для этой третьей группы в своей книге 1535 года «О французских и латинских названиях рыб региона Марселя» ; широко распространено, но ошибочно полагают, что этот термин создал Аристотель. [4] Гиллий далее, вслед за Аристотелем, отметил, как трудно было определить, что такое растение, а что животное. [4] В Вавилонском Талмуде коралл упоминается в списке типов деревьев, а французский комментатор XI века Раши описывает его как «тип дерева (מין עץ), растущего под водой и носящего (французское) название «коралл». [5]
Персидский эрудит Аль-Бируни (ум. 1048) отнес губки и кораллы к животным, утверждая, что они реагируют на прикосновение. [6] Тем не менее, люди считали кораллы растениями до восемнадцатого века, когда Уильям Гершель с помощью микроскопа установил, что кораллы имеют характерные тонкие клеточные мембраны животных . [7]
пределах подклассов Hexacorallia и Octocorallia класса В настоящее Anthozoa типа Cnidaria время кораллы классифицируются как виды животных в . [8] Hexacorallia включает каменные кораллы, и в этих группах есть полипы , которые обычно имеют 6-кратную симметрию. Octocorallia включает синий коралл и мягкие кораллы , а виды Octocorallia имеют полипы с восьмикратной симметрией, каждый полип имеет восемь щупалец и восемь брыжейок . Группа кораллов является парафилетической , поскольку морские анемоны также относятся к подклассу Hexacorallia.
Систематика
[ редактировать ] в этом разделе Использование внешних ссылок может не соответствовать политике и рекомендациям Википедии . ( Август 2023 г. ) |
Определение видов кораллов является сложной задачей, поскольку гипотезы, основанные на морфологических признаках, противоречат гипотезам, сформированным с помощью процессов на основе молекулярных деревьев. [9] По состоянию на 2020 год выявлено 2175 отдельных видов кораллов, 237 из которых в настоящее время находятся под угрозой исчезновения. [10] придание различению кораллов первостепенной важности в усилиях по сдерживанию вымирания. [9] Адаптация и разграничение продолжают происходить у видов кораллов. [11] в целях борьбы с опасностями, вызванными климатическим кризисом. Кораллы — это колониальные модульные организмы, образованные бесполым путем и генетически идентичные модули, называемые полипами. Полипы соединяются живой тканью, образуя полноценный организм. [12] Живая ткань обеспечивает межмодульную связь (взаимодействие между каждым полипом), [12] который проявляется в морфологии колоний , образуемых кораллами, и является одной из основных отличительных характеристик вида кораллов. [12]
Существует две основные классификации кораллов: твердые кораллы (склерактиниевые и каменистые кораллы). [13] которые образуют рифы на основе карбоната кальция с полипами, несущими шесть жестких щупалец, [14] и мягкие кораллы (Alcyonacea и агерматипический коралл) [13] которые являются гибкими и образованы колонией полипов с восемью перистыми щупальцами. [14] Эти две классификации возникли в результате дифференциации экспрессии генов на кончиках их ветвей. [12] и основания, возникшие через сигнальные пути развития, такие как Hox , Hedgehog , Wnt , BMP и т. д.
Ученые обычно выбирают Acropora в качестве исследовательской модели, поскольку это самый разнообразный род твердых кораллов, насчитывающий более 120 видов. [12] У большинства видов этого рода есть диморфные полипы: [15] осевые полипы быстро растут и имеют более светлую окраску, тогда как радиальные полипы маленькие и имеют более темную окраску. [12] [16] У Acropora рода синтез гамет и фотосинтез происходят в базальном отделе. [17] полипы, рост происходит преимущественно на радиальных полипах. Рост на месте радиальных полипов включает в себя два процесса: бесполое размножение посредством митотической пролиферации клеток , [12] и отложение скелета карбоната кальция посредством белков внеклеточного матрикса (EMC), действующих как дифференциально экспрессируемые (DE) сигнальные гены. [12] между кончиками и основаниями ветвей. Эти процессы приводят к дифференциации колоний , которая является наиболее точным способом различения видов кораллов. [9] У рода Acropora дифференциация колоний посредством повышения и понижения регуляции DE. [12]
Систематические исследования видов мягких кораллов столкнулись с трудностями из-за отсутствия таксономических знаний. [9] Исследователи не обнаружили достаточной изменчивости внутри рода, чтобы уверенно определить сходные виды из-за низкой скорости мутаций митохондриальной ДНК . [18]
Факторы окружающей среды, такие как повышение температуры и уровня кислотности в наших океанах, являются причиной некоторых видообразований кораллов в виде утраченных видов . [12] Различные виды кораллов имеют белки теплового шока (HSP), которые также относятся к категории DE у разных видов. [12] Эти HSP помогают кораллам бороться с повышенными температурами, с которыми они сталкиваются, что приводит к денатурации белков, замедлению роста и, в конечном итоге, к гибели кораллов. [12] Примерно 33% видов кораллов включены в список исчезающих видов Международного союза охраны природы и находятся под угрозой исчезновения. [19] Закисление океана (падение уровня pH в океанах) ставит под угрозу продолжающийся рост видов и дифференциацию кораллов. [12] Скорость мутации Vibrio shilonii , рифового патогена , ответственного за обесцвечивание кораллов , значительно превышает типичную скорость размножения колоний кораллов при падении уровня pH. [20] Таким образом, кораллы не могут мутировать свои HSP и другие гены, предотвращающие изменение климата, чтобы бороться с повышением температуры и снижением pH с такой скоростью, которая конкурирует с этими патогенами, ответственными за обесцвечивание кораллов. [20] что приводит к потере вида.
Анатомия
[ редактировать ]
Большую часть своей жизни кораллы являются сидячими животными колоний генетически идентичных полипов . Диаметр каждого полипа варьируется от миллиметров до сантиметров, а колонии могут образовываться из многих миллионов отдельных полипов. Каменистый коралл, также известный как твердый коралл, полипы образуют скелет, состоящий из карбоната кальция, который укрепляет и защищает организм. Он откладывается полипами и ценосарком , живой тканью, которая их соединяет. Полипы располагаются в чашеобразных углублениях скелета, известных как кораллиты . Колонии каменистых кораллов заметно различаются по внешнему виду; один вид может иметь корковую, пластинчатую, кустистую, столбчатую или массивную твердую структуру, причем различные формы часто связаны с разными типами среды обитания, при этом различия в уровне освещенности и движении воды являются значительными. [21]
По строению тело полипа можно грубо сравнить с мешком , стенка которого состоит из двух слоев клеток . Внешний слой технически известен как эктодерма , внутренний слой — как энтодерма . Между эктодермой и энтодермой находится поддерживающий слой студенистого вещества, называемого мезоглеей , секретируемого клеточными слоями стенки тела. [22] В мезоглее могут содержаться скелетные элементы, происходящие из клеток, мигрировавших из эктодермы.
Построенное таким образом мешкообразное тело прикрепляется к твердой поверхности, которая у твердых кораллов представляет собой чашеобразные углубления в скелете, известные как кораллиты . В центре верхнего конца мешка находится единственное отверстие, называемое ртом, окруженное кругом щупалец , напоминающих пальцы-перчатки. Щупальца — это органы , служащие как для осязания, так и для захвата пищи. [22] Полипы вытягивают свои щупальца, особенно ночью, часто содержащие скрученные стрекательные клетки ( книдоциты ), которые пронзают, отравляют и прочно удерживают живую добычу, парализуя или убивая ее. Добычей полипов является планктон, такой как копеподы и личинки рыб. Продольные мышечные волокна, образованные из клеток эктодермы, позволяют щупальцам сокращаться, чтобы доставить пищу ко рту. Точно так же циркулярно расположенные мышечные волокна, образующиеся из энтодермы, позволяют щупальцам вытягиваться или выдвигаться наружу после их сокращения. [22] Как у каменистых, так и у мягких кораллов полипы могут втягиваться за счет сокращения мышечных волокон, при этом каменистые кораллы полагаются на свой твердый скелет и книдоциты для защиты. Мягкие кораллы обычно выделяют терпеноидные токсины, чтобы отпугнуть хищников. [21]
У большинства кораллов щупальца днем втягиваются, а ночью расправляются для ловли планктона и других мелких организмов. Мелководные виды как каменистых, так и мягких кораллов могут быть зооксантеллятами , при этом кораллы дополняют свой планктонный рацион продуктами фотосинтеза, производимыми этими симбионтами . [21] Полипы соединяются между собой сложной и хорошо развитой системой гастроваскулярных каналов, обеспечивающей значительный обмен питательными веществами и симбионтами. [23]
Внешняя форма полипа сильно варьирует. Колонна может быть длинной и тонкой или может быть настолько короткой в осевом направлении, что тело становится дискообразным. Щупальца могут насчитывать многие сотни или быть очень мало, в редких случаях только одно или два. Они могут быть простыми и неразветвленными или иметь перистый рисунок. Рот может находиться на уровне поверхности перистома или может быть выступающим и иметь трубчатую форму. [22]
Мягкие кораллы
[ редактировать ]Мягкие кораллы не имеют твердого экзоскелета как такового. Однако их ткани часто укреплены небольшими поддерживающими элементами, известными как склериты, состоящие из карбоната кальция. Полипы мягких кораллов обладают восьмикратной симметрией, что отражено в окто у Octocorallia. [24]
Мягкие кораллы значительно различаются по форме, большинство из них являются колониальными. Некоторые мягкие кораллы столонатные , но полипы большинства из них соединены листками ткани, называемыми ценосарками, а у некоторых видов эти листы толстые, и полипы глубоко погружены в них. Некоторые мягкие кораллы покрывают другие морские объекты или образуют лепестки. Другие имеют древовидную или кнутовидную форму и имеют центральный осевой скелет, встроенный у основания в матрицу опорной ветви. [25] Эти ветви состоят из волокнистого белка, называемого горгонином , или из кальцинированного материала.
Каменистые кораллы
[ редактировать ]
Полипы каменистых кораллов обладают шестикратной симметрией. У каменных кораллов щупальца имеют цилиндрическую форму и сужаются к концу, а у мягких кораллов они перистые с боковыми ветвями, известными как перышки. У некоторых тропических видов они превратились в простые пеньки, а у некоторых они срослись, придавая вид весла. [26]
Скелеты кораллов представляют собой биокомпозиты (минерал + органика) карбоната кальция в форме кальцита или арагонита. У склерактиновых кораллов «центры обызвествления» и волокна представляют собой четко различимые структуры, различающиеся как по морфологии, так и по химическому составу кристаллических единиц. [27] [28] Органические матрицы, извлеченные из различных видов, являются кислыми и содержат белки, сульфатированные сахара и липиды; они видоспецифичны. [29] Растворимые органические матрицы скелетов позволяют дифференцировать зооксантеллы и незооксантеллы. [30]
Экология
[ редактировать ]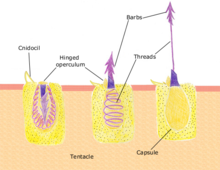
Кормление
[ редактировать ]Полипы питаются множеством мелких организмов, от микроскопического зоопланктона до мелких рыб. Щупальца полипа обездвиживают или убивают добычу с помощью стрекательных клеток, называемых нематоцистами . Эти клетки несут яд , который они быстро выделяют в ответ на контакт с другим организмом. Спящая нематоциста разряжается в ответ на прикосновение находящейся поблизости добычи к спусковому крючку ( Cnidocil ). Створка ( operculum ) открывается, и ее жалящий аппарат выпускает зазубрину в добычу. Яд вводится через полую нить, чтобы обездвижить добычу; затем щупальца направляют добычу в желудок. Как только добыча переварена, желудок вновь открывается, позволяя удалить отходы и начать следующий цикл охоты. [31] : 24
Внутриклеточные симбионты
[ редактировать ]Многие кораллы, а также другие группы книдарий, такие как морские анемоны , образуют симбиотические отношения с классом динофлагеллятных водорослей , зооксантеллами рода Symbiodinium , которые могут составлять до 30% ткани полипа. [31] : 23–24 Обычно каждый полип содержит один вид водорослей, а виды кораллов отдают предпочтение Symbiodinium . [32] Молодые кораллы не рождаются с зооксантеллами, а приобретают водоросли из окружающей среды, включая толщу воды и местные отложения. [33] Основным преимуществом зооксантелл является их способность к фотосинтезу, который снабжает кораллы продуктами фотосинтеза, включая глюкозу, глицерин, а также аминокислоты, которые кораллы могут использовать для получения энергии. [34] Зооксантеллы также приносят пользу кораллам, способствуя кальцификации кораллового скелета и удалению отходов. [35] [36] Помимо мягких тканей, микробиомы также обнаружены в слизи кораллов и (у каменистых кораллов) в скелете, причем последний демонстрирует наибольшее микробное богатство. [37]
полипа Зооксантеллы извлекают выгоду из безопасного места для жизни и потребляют углекислый газ , фосфаты и азотистые отходы . Кораллы, подвергшиеся стрессу, выбрасывают свои зооксантеллы, и этот процесс становится все более распространенным из-за нагрузки, оказываемой кораллам повышением температуры океана. Массовые выбросы известны как обесцвечивание кораллов , поскольку водоросли способствуют окраске кораллов; однако некоторые цвета обусловлены коралловыми пигментами-хозяевами, такими как зеленые флуоресцентные белки (GFP). Выброс увеличивает шансы полипа пережить кратковременный стресс, и если стресс утихнет, они смогут восстановить водоросли, возможно, другого вида, позже. Если стрессовые условия сохраняются, полип со временем погибает. [38] Зооксантеллы расположены внутри цитоплазмы коралла, и из-за фотосинтетической активности водорослей внутренний pH коралла может повышаться; такое поведение указывает на то, что зооксантеллы в некоторой степени ответственны за метаболизм кораллов-хозяев. [39] Болезнь потери ткани каменистых кораллов связана с нарушением физиологии зооксантелл-хозяев. [40] Кроме того, известно, что бактерии Vibrio обладают свойствами вирулентности, используемыми для повреждения тканей кораллов-хозяев и фотоингибирования водорослевых симбионтов. [41] Таким образом, и кораллы, и их симбиотические микроорганизмы могли эволюционировать и обрести черты, устойчивые к болезням и передаче инфекции.
Воспроизведение
[ редактировать ]Кораллы могут быть как гонохорическими (однополыми), так и гермафродитными , каждый из которых может размножаться половым и бесполым путем. Размножение также позволяет кораллам селиться на новых территориях. Размножение координируется химической связью. [ объяснить ]
Сексуальный
[ редактировать ]
Кораллы размножаются преимущественно половым путем . Около 25% герматипических кораллов (рифостроительных каменистых кораллов) образуют однополые ( гонохористические ) колонии, остальные — гермафродиты . [ нужна ссылка ] По оценкам, более 67% кораллов одновременно являются гермафродитами . [42]
Вещательные компании
[ редактировать ]| Внешние видео | |
|---|---|
 | |
Около 75% всех герматипических кораллов «транслируют икру». [ нужна ссылка ] выпуская гаметы — яйцеклетки и сперматозоиды — в воду, где они встречаются и оплодотворяются для распространения потомства. Кораллы часто синхронизируют время своего нереста. Эта репродуктивная синхронность необходима для того, чтобы мужские и женские гаметы могли встретиться. Нерест часто происходит вечером или ночью и может происходить нечасто, например, один раз в год и в течение 10–30 минут. [43] [44] Синхронный нерест очень типичен для коралловых рифов, и часто все кораллы нерестятся в одну и ту же ночь, даже если несколько видов . присутствуют [45] Синхронный нерест может образовывать гибриды и, возможно, участвует в видообразовании кораллов . [46]

Экологические сигналы, влияющие на выброс гамет в воду, варьируются от вида к виду. Сигналы включают изменение температуры, лунный цикл , продолжительность дня и, возможно, химические сигналы. [45] Другие факторы, влияющие на ритмичность организмов в морской среде обитания, включают соленость, механические силы и изменения давления или магнитного поля. [44]
Массовое нерест кораллов часто происходит ночью в дни после полнолуния. [43] [47] Полнолуние эквивалентно четырем-шести часам непрерывного воздействия тусклого света, который может вызвать светозависимые реакции в белке. [43] [44] Кораллы содержат светочувствительные криптохромы — белки, светопоглощающие флавиновые структуры которых чувствительны к различным типам света. Это позволяет кораллам, таким как Dipsastraea speciosa, обнаруживать изменения солнечного и лунного света и реагировать на них. [43] [44] [48]
Лунный свет сам по себе может фактически подавлять нерест кораллов. Самым непосредственным сигналом к нересту является темная часть ночи между закатом и восходом луны. В течение лунного цикла восход луны постепенно смещается позже, что происходит после захода солнца в день полнолуния. Возникающий в результате темный период между дневным и ночным светом устраняет подавляющий эффект лунного света и позволяет кораллам нереститься. [43] [47]
Событие нереста может быть визуально драматичным, поскольку обычно прозрачная вода замутняется гаметами. После выхода на поверхность гаметы оплодотворяются на поверхности воды и образуют микроскопическую личинку , называемую планулой , обычно розовую и эллиптическую по форме. Типичная колония кораллов должна выпускать несколько тысяч личинок в год, чтобы преодолеть препятствия к образованию новой колонии. [49] [50]
Исследования показывают, что световое загрязнение десинхронизирует нерест некоторых видов кораллов.В таких районах, как Красное море , до 10 из 50 видов могут демонстрировать асинхронность нереста по сравнению с тем, что было 30 лет назад. Образование новых кораллов в этом районе уменьшилось, а в некоторых случаях прекратилось. Ранее этот район считался убежищем для кораллов, поскольку массового обесцвечивания из-за изменения климата здесь не наблюдалось. [43] [51] Методы восстановления кораллов для управления коралловыми рифами разрабатываются для увеличения скорости оплодотворения, развития личинок и заселения новых кораллов. [52]
Брудеры
[ редактировать ]Высиживающие виды чаще всего агерматипичны (не строят рифы) в районах с сильным течением или волнением. Брудеры выделяют только сперму, которая имеет отрицательную плавучесть и опускается на ожидающих яйценосителей, которые неделями содержат неоплодотворенные яйца. Даже у этих видов иногда происходят синхронные нересты. [45] После оплодотворения кораллы выпускают планулу, готовую к оседанию. [35]

Планулы
[ редактировать ]Время от нереста до поселения личинок обычно составляет два-три дня, но может произойти сразу или до двух месяцев. [53] отложенные разбросом, Личинки планулы, развиваются на поверхности воды, прежде чем спуститься в поисках твердой поверхности бентоса, к которой они могут прикрепиться и начать новую колонию. [54] Личинкам часто нужен биологический сигнал, чтобы вызвать заселение, например, определенные виды корковых коралловых водорослей или микробные биопленки. [55] [56] На многих этапах этого процесса наблюдается высокий уровень неудач, и хотя каждая колония выпускает тысячи яиц, новых колоний образуется мало. Во время расселения личинки сдерживаются физическими барьерами, такими как осадки, [57] а также химические (аллелопатические) барьеры. [58] Личинки превращаются в один полип и в конечном итоге развиваются в ювенильную, а затем и во взрослую особь в результате бесполого почкования и роста.
Бесполый
[ редактировать ]
Внутри коралловой головки генетически идентичные полипы размножаются бесполым путем : либо путем почкования (почкования), либо путем деления, продольного или поперечного.
Почкование предполагает отделение полипа меньшего размера от взрослого. [49] По мере роста нового полипа из него формируются части тела . Расстояние между новыми и взрослыми полипами увеличивается, а вместе с ним и ценосарк (общее тело колонии). Почкование может быть внутрищупальцевым, из ротовых дисков, образуя полипы одинакового размера внутри кольца щупалец, или экстратентакулярным, из его основания, образуя полип меньшего размера.
При делении образуются два полипа, каждый из которых становится таким же большим, как и оригинал. Продольное деление начинается, когда полип расширяется, а затем разделяет свой кишечнополостной (тело), эффективно разделяясь по длине. Рот разделяется и образуются новые щупальца. Два созданных таким образом полипа затем создают недостающие части тела и экзоскелет. Поперечное деление происходит, когда полипы и экзоскелет разделяются поперечно на две части. Это означает, что у одного есть базальный диск (внизу), а у другого — оральный диск (вверху); новые полипы должны отдельно образовать недостающие части.
Бесполое размножение предлагает преимущества высокой репродуктивной способности, задержки старения и замены мертвых модулей, а также географического распространения. [ нужны разъяснения ] [59]
Подразделение колонии
[ редактировать ]Целые колонии могут размножаться бесполым путем, образуя две колонии с одинаковым генотипом. Возможные механизмы включают деление, спасение и фрагментацию. Деление происходит у некоторых кораллов, особенно среди семейства Fungiidae , где колония разделяется на две или более колоний на ранних стадиях развития. Спасение происходит, когда одиночный полип покидает колонию и селится на другом субстрате, чтобы создать новую колонию. Фрагментация включает в себя особей, вырвавшихся из колонии во время штормов или других потрясений. Отделившиеся особи могут основать новые колонии. [60]
Коралловые микробиомы
[ редактировать ]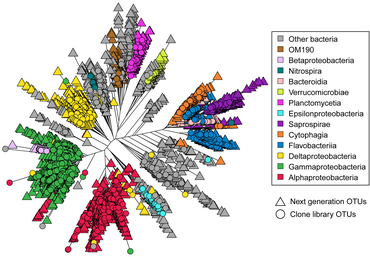
Кораллы являются одним из наиболее распространенных примеров животных-хозяев, чей симбиоз с микроводорослями может перерасти в дисбиоз , что заметно проявляется в обесцвечивании. Коралловые микробиомы изучались в различных исследованиях, которые демонстрируют, как изменения океанической окружающей среды, особенно температура, свет и неорганические питательные вещества, влияют на численность и продуктивность симбионтов микроводорослей, а также на кальцификацию и физиологию хозяина. [62] [63] [64]
Исследования также показали, что резидентные бактерии, археи и грибы дополнительно способствуют круговороту питательных веществ и органических веществ внутри коралла, а вирусы также, возможно, играют роль в структурировании состава этих членов, что дает один из первых проблесков многообразия. домен морских животных симбиоз. [65] Гаммапротеобактерия Endozoicomonas . становится центральным членом микробиома коралла и ведет гибкий образ жизни [66] [67] Учитывая недавнее массовое обесцвечивание рифов, [68] кораллы, вероятно, останутся полезной и популярной системой для исследования симбиоза и дисбиоза. [64]
Astrangia poculata , северный звездчатый коралл, представляет собой каменистый коралл умеренного пояса , широко встречающийся вдоль восточного побережья Соединенных Штатов. Коралл может жить как с зооксантеллами (водорослевыми симбионтами), так и без них, что делает его идеальным модельным организмом для изучения взаимодействий микробного сообщества, связанных с симбиотическим состоянием. Однако возможность разработки праймеров и зондов для более целенаправленного воздействия на ключевые микробные группы затруднена из-за отсутствия полноразмерных последовательностей 16S рРНК , поскольку последовательности, полученные с помощью платформы Illumina, имеют недостаточную длину (около 250 пар оснований) для дизайна. праймеров и зондов. [69] В 2019 году Голдсмит и др. продемонстрировало, что секвенирование по Сэнгеру способно воспроизводить биологически значимое разнообразие, обнаруженное с помощью более глубокого секвенирования следующего поколения , а также создавать более длинные последовательности, полезные исследовательскому сообществу для разработки зондов и праймеров (см. Диаграмму справа). [70]
Голобионты
[ редактировать ]Рифостроящие кораллы — это хорошо изученные холобионты , включающие в себя сам коралл вместе с его симбионтными зооксантеллами (фотосинтезирующими динофлагеллятами), а также ассоциированными с ним бактериями и вирусами. [71] Существуют модели коэволюции коралловых микробных сообществ и филогении кораллов. [72]

Известно, что микробиом и симбионт коралла влияют на здоровье хозяина, однако историческое влияние каждого члена на других недостаточно изучено. Склерактиновые кораллы диверсифицируются дольше, чем многие другие симбиотические системы, и известно, что их микробиомы частично видоспецифичны. [74] Было высказано предположение, что Endozoicomonas , широко распространенная бактерия в кораллах, продемонстрировала ко-диверсификацию со своим хозяином. [75] [76] Это намекает на сложный комплекс взаимоотношений между членами кораллового холобионта, которые развивались по мере эволюции этих членов.
Исследование, опубликованное в 2018 году [77] выявили доказательства филосимбиоза между кораллами и микробиомами их тканей и скелета. Скелет коралла, который представляет собой наиболее разнообразный из трех коралловых микробиомов, продемонстрировал наиболее убедительные доказательства филосимбиоза. состав и богатство Было обнаружено, что кораллового микробиома отражает филогению кораллов . Например, взаимодействие между бактериальной и эукариотической филогенией кораллов влияет на численность Endozoicomonas , очень распространенной бактерии в коралловом холобионте. Однако кофилогения микроорганизмов-хозяев , по-видимому, влияет только на подмножество бактерий, связанных с кораллами.

AMP = антимикробные пептиды, ROS = активные формы кислорода.

Рифы
[ редактировать ]
Многие кораллы отряда Scleractinia являются герматипическими , то есть участвуют в построении рифов. Большинство таких кораллов получают часть своей энергии от зооксантелл рода Symbiodinium . Это симбиотические фотосинтезирующие динофлагелляты , которым необходим солнечный свет; Поэтому кораллы, образующие рифы, встречаются в основном на мелководье. Они выделяют карбонат кальция, образуя твердые скелеты, которые становятся каркасом рифа. Однако не все кораллы, образующие рифы на мелководье, содержат зооксантеллы, а некоторые глубоководные виды, обитающие на глубинах, куда не проникает свет, образуют рифы, но не дают убежища симбионтам. [80]

Существуют различные типы мелководных коралловых рифов, в том числе окаймляющие рифы, барьерные рифы и атоллы; большинство из них встречается в тропических и субтропических морях. Они очень медленно растут, каждый год прибавляя примерно один сантиметр (0,4 дюйма) в высоту. Считается, что Большой Барьерный риф образовался около двух миллионов лет назад. Со временем кораллы фрагментируются и умирают, между кораллами скапливается песок и щебень, а раковины моллюсков и других моллюсков разлагаются, образуя постепенно развивающуюся структуру карбоната кальция. [81] Коралловые рифы представляют собой чрезвычайно разнообразные морские экосистемы, в которых обитает более 4000 видов рыб, огромное количество книдарий, моллюсков , ракообразных и многих других животных. [82]
Эволюция
[ редактировать ]
В определенные времена в геологическом прошлом кораллов было очень много. Подобно современным кораллам, их предки строили рифы, некоторые из которых заканчивались огромными сооружениями в осадочных породах . Наряду с окаменелостями кораллов появляются окаменелости других обитателей рифов — водорослей, губок, а также останки многих морских ежей , брахиопод , двустворчатых моллюсков , брюхоногих моллюсков и трилобитов . Это делает некоторые кораллы полезными индикаторными окаменелостями . [84] Окаменелости кораллов не ограничиваются остатками рифов, и многие одиночные окаменелости встречаются в других местах, например, Cyclocyathus в Англии , который встречается в глиняных формациях Голта .
Ранние кораллы
[ редактировать ]Кораллы впервые появились в кембрии около 535 миллионов лет назад . [85] Окаменелости встречаются крайне редко до ордовикского периода, 100 миллионов лет спустя, когда широкое распространение получили гелиолитиды, морщинистые и таблитчатые кораллы. Палеозойские кораллы часто содержали многочисленные эндобиотические симбионты. [86] [87]
Таблитчатые кораллы встречаются в известняках и известковых сланцах ордовика с пробелом в летописи окаменелостей из-за событий вымирания в конце ордовика. Кораллы вновь появились несколько миллионов лет спустя, в силурийский период, а таблитчатые кораллы часто образуют низкие подушки или ветвящиеся массы кальцита рядом с морщинистыми кораллами. Численность таблитчатых кораллов начала сокращаться в середине силурийского периода. [88]
Ругозные или роговые кораллы стали доминировать к середине силурийского периода, а в девонском периоде кораллы процветали, насчитывая более 200 родов. Морщинистые кораллы существовали в одиночных и колониальных формах и также состояли из кальцита. [89] И морщинистые, и таблитчатые кораллы вымерли во время пермско-триасового вымирания. [88] [90] 250 миллионов лет назад есть промежуток в десятки миллионов лет (вместе с 85% морских видов), и до появления новых форм кораллов в триасе .
 Таблитчатый коралл (сирингопорид); Известняк Буна (нижний карбон ) недалеко от Хивассе, Арканзас, масштабная линейка 2,0 см.
Таблитчатый коралл (сирингопорид); Известняк Буна (нижний карбон ) недалеко от Хивассе, Арканзас, масштабная линейка 2,0 см.
 Одиночный морщинистый коралл ( Grewingkia ) в трех видах; Ордовик, юго-восток Индианы
Одиночный морщинистый коралл ( Grewingkia ) в трех видах; Ордовик, юго-восток Индианы



Современные кораллы
[ редактировать ]Повсеместные в настоящее время каменистые кораллы Scleractinia появились в среднем триасе, чтобы заполнить нишу, освободившуюся вымершими складчатыми и таблитчатыми отрядами, и не имеют тесного родства с более ранними формами. В отличие от кораллов, преобладавших до пермского вымирания, которые образовывали скелеты из карбоната кальция, известного как кальцит , современные каменистые кораллы образуют скелеты, состоящие из арагонита . [91] Их окаменелости в небольшом количестве встречаются в породах триасового периода и становятся обычным явлением в юрском и более поздних периодах. [92] Хотя они геологически моложе таблитчатых и морщинистых кораллов, арагонит их скелетов сохраняется труднее, и их летопись окаменелостей, соответственно, менее полна.
 | |
Хронология основных летописей окаменелостей кораллов и событий от 650 млн лет назад до настоящего времени. [93] [94] | |
Статус
[ редактировать ]Угрозы
[ редактировать ]
Коралловые рифы по всему миру находятся под нагрузкой. [95] В частности, добыча кораллов, сельскохозяйственные и городские стоки , загрязнение (органическое и неорганическое), чрезмерный вылов рыбы , взрывной промысел , болезни, а также рытье каналов и доступ к островам и заливам представляют собой локальные угрозы коралловым экосистемам. Более широкими угрозами являются повышение температуры моря, повышение уровня моря и pH изменения в результате закисления океана , и все это связано с выбросами парниковых газов . [96] В 1998 году 16% рифов мира погибли в результате повышения температуры воды. [97]
Около 10% коралловых рифов мира мертвы. [98] [99] [100] Около 60% рифов мира находятся под угрозой из-за деятельности человека. [101] Угроза здоровью рифов особенно сильна в Юго-Восточной Азии , где 80% рифов находятся под угрозой исчезновения . [102] мира Более 50% коралловых рифов могут быть уничтожены к 2030 году; в результате большинство стран защищают их посредством законов об охране окружающей среды. [103]
В Карибском бассейне и тропической зоне Тихого океана прямой контакт примерно 40–70% обычных морских водорослей с кораллами вызывает обесцвечивание и гибель кораллов в результате жирорастворимых переноса метаболитов . [104] Морские водоросли и водоросли размножаются при наличии достаточного количества питательных веществ и ограниченном выпасе травоядных животных, таких как рыбы-попугаи .
Изменения температуры воды более чем на 1–2 ° C (1,8–3,6 ° F) или изменения солености могут привести к гибели некоторых видов кораллов. При таком экологическом стрессе кораллы вытесняют свой симбиодиниум ; без них ткани кораллов обнажают белизну своих скелетов — явление, известное как обесцвечивание кораллов . [105]
Подводные источники, обнаруженные вдоль побережья полуострова Юкатан в Мексике , производят воду с естественным низким pH (относительно высокой кислотностью), создавая условия, аналогичные тем, которые, как ожидается, получат широкое распространение по мере поглощения океанами углекислого газа. [106] Исследования обнаружили несколько видов живых кораллов, которые, по-видимому, хорошо переносят кислотность. Колонии были небольшими и неоднородно распределенными и не образовывали структурно сложных рифов, подобных тем, которые составляют близлежащую Мезоамериканскую систему барьерных рифов . [106]
Здоровье кораллов
[ редактировать ]Чтобы оценить уровень угрозы кораллам, ученые разработали коэффициент дисбаланса кораллов, Log (Средняя численность таксонов, связанных с болезнями / Средняя численность здоровых ассоциированных таксонов). Чем ниже это соотношение, тем здоровее микробное сообщество. Это соотношение было разработано после сбора и изучения микробной слизи кораллов. [107]
Последствия изменения климата
[ редактировать ]Повышение температуры поверхности моря в тропических регионах (~ 1 ° C (1,8 ° F)) в прошлом веке привело к значительному обесцвечиванию, гибели кораллов и, следовательно, к сокращению популяций кораллов. Хотя кораллы способны адаптироваться и акклиматизироваться, неясно, произойдет ли этот эволюционный процесс достаточно быстро, чтобы предотвратить значительное сокращение их численности. [108] Изменение климата приводит к более частым и сильным штормам, которые могут разрушить коралловые рифы . [109]
Годовые полосы роста некоторых кораллов, таких как глубоководные бамбуковые кораллы ( Isididae ), могут быть одними из первых признаков воздействия закисления океана на морскую жизнь. [110] Годовые кольца позволяют геологам строить год за годом хронологию — форму поэтапного датирования , которая лежит в основе записей с высоким разрешением прошлых климатических и экологических изменений с использованием геохимических методов. [111]
Некоторые виды образуют сообщества, называемые микроатоллами , которые представляют собой колонии, верхняя часть которых мертва и в основном находится над линией воды, но чей периметр в основном погружен под воду и жив. Средний уровень прилива ограничивает их высоту. Анализируя различные морфологии роста, микроатоллы позволяют получить данные об изменении уровня моря с низким разрешением. Окаменелые микроатоллы также можно датировать с помощью радиоуглеродного датирования . Такие методы могут помочь восстановить в голоцене уровень моря . [112]
Хотя кораллы имеют большие популяции, размножающиеся половым путем, их эволюция может быть замедлена обильным бесполым размножением . [113] Поток генов варьируется среди видов кораллов. [113] Согласно биогеографии видов кораллов, поток генов нельзя считать надежным источником адаптации, поскольку они являются очень стационарными организмами. Кроме того, долговечность кораллов может повлиять на их адаптивность. [113]
Однако во многих случаях была продемонстрирована адаптация к изменению климата , что обычно связано со сдвигом генотипов кораллов и зооксантелл . Эти сдвиги в частоте аллелей прогрессировали в сторону более толерантных типов зооксантелл. [114] Ученые обнаружили, что определенная склерактиновая зооксантелла становится все более распространенной там, где температура моря высока. [115] [116] Симбионты, способные переносить более теплую воду, похоже, фотосинтезируют медленнее, что подразумевает эволюционный компромисс. [116]
В Мексиканском заливе, где температура моря повышается, чувствительные к холоду оленьего рога и кораллы лосося сместились. [114] Было показано, что не только симбионты и отдельные виды меняются, но, по-видимому, существует определенная скорость роста, благоприятная для отбора. Медленно растущие, но более устойчивые к жаре кораллы стали более распространенными. [117] Изменения температуры и акклиматизации сложны. Некоторые рифы в нынешней тени представляют собой рефугиумы , которые помогут им приспособиться к неравенству окружающей среды, даже если в конечном итоге температура там может повышаться быстрее, чем в других местах. [118] Такое разделение населения климатическими барьерами приводит к значительному сокращению реализованной ниши по сравнению со старой фундаментальной нишей .
Геохимия
[ редактировать ]интегрируют кислород и микроэлементы в свои скелетные кристаллические структуры арагонита ( полиморфа кальцита Кораллы — это мелкие колониальные организмы , которые по мере роста ). Геохимические аномалии в кристаллических структурах кораллов представляют собой функцию температуры, солености и изотопного состава кислорода. Такой геохимический анализ может помочь в моделировании климата. [119] Отношение кислорода-18 к кислороду-16 (δ 18 O), например, является показателем температуры.
Аномалия соотношения стронций/кальций
[ редактировать ]Время можно отнести к аномалиям геохимии кораллов путем корреляции минимумов стронция / кальция с максимумами температуры поверхности моря (SST) с данными, собранными из NINO 3.4 SSTA . [120]
Аномалия изотопа кислорода
[ редактировать ]Сравнение коралловых минимумов стронция/кальция с максимумами температуры поверхности моря, данные, записанные с помощью NINO 3.4 SSTA , время может быть коррелировано с коралловым стронцией/кальцием и δ. 18 О вариации. Для подтверждения точности годовой зависимости между Sr/Ca и δ 18 Вариации , заметная связь с годовыми кольцами роста кораллов, подтверждают изменение возраста. Геохронология устанавливается путем объединения данных Sr/Ca, годовых колец и стабильных изотопов данных . Эль-Ниньо-Южное колебание (ЭНСО) напрямую связано с колебаниями климата, которые влияют на δ кораллов. 18 Коэффициент O из местных изменений солености связан с положением южно-тихоокеанской зоны конвергенции (SPCZ) и может использоваться для моделирования ЭНЮК . [120]
Температура поверхности моря и соленость поверхности моря
[ редактировать ]
На глобальный баланс влаги в первую очередь влияют тропические температуры поверхности моря из-за зоны внутритропической конвергенции (ITCZ). [121] Южное полушарие имеет уникальную метеорологическую особенность, расположенную в юго-западной части Тихоокеанского бассейна, называемую Южно-Тихоокеанской зоной конвергенции (SPCZ) , которая постоянно находится в Южном полушарии. В ЭНСО теплые периоды SPCZ меняет ориентацию, простираясь от экватора на юг через Соломоновы острова , Вануату , Фиджи и к Французским полинезийским островам ; и на восток в сторону Южной Америки, влияя на геохимию кораллов в тропических регионах. [122]
Геохимический анализ скелетных кораллов может быть связан с соленостью поверхности моря (SSS) и температурой поверхности моря (SST), от SSTA Эль-Ниньо 3.4 данных , от тропических океанов до морской воды δ. 18 Аномалии соотношения O в кораллах. Феномен ЭНЮК может быть связан с изменениями солености поверхности моря (SSS) и температуры поверхности моря (SST), которые могут помочь моделировать деятельность тропического климата. [123]
Ограниченные климатические исследования нынешних видов
[ редактировать ]
Климатические исследования живых видов кораллов ограничены несколькими изученными видами. Изучение кораллов Porites обеспечивает стабильную основу для геохимических интерпретаций, из которых гораздо проще физически извлекать данные по сравнению с видами Platygyra , где сложность скелетной структуры видов Platygyra создает трудности при физическом отборе проб, что является одной из немногих используемых записей о живых кораллах за несколько десятилетий. для моделирования кораллового палеоклимата . [123]
Защита
[ редактировать ]Морские охраняемые территории, биосферные заповедники , морские парки , национальные памятники всемирного наследия , статус , управление рыболовством и защита среды обитания могут защитить рифы от антропогенного ущерба. [124]
Многие правительства теперь запрещают удаление кораллов с рифов и информируют прибрежных жителей о защите рифов и экологии. Хотя местные действия, такие как восстановление среды обитания и защита травоядных животных, могут уменьшить локальный ущерб, долгосрочные угрозы закисления, изменения температуры и повышения уровня моря остаются проблемой. [96]
Защита сети разнообразных и здоровых рифов, а не только климатических рефугиумов , помогает обеспечить максимальную вероятность генетического разнообразия , которое имеет решающее значение для адаптации кораллов к новому климату. [125] Разнообразие методов сохранения, применяемых в морских и наземных экосистемах, находящихся под угрозой, делает адаптацию кораллов более вероятной и эффективной. [125]
Чтобы предотвратить уничтожение кораллов в их местных регионах, были начаты проекты по выращиванию кораллов в нетропических странах. [126] [127]
Отношение к людям
[ редактировать ]Местная экономика вблизи крупных коралловых рифов получает выгоду от обилия рыбы и других морских существ в качестве источника пищи. Рифы также обеспечивают рекреационное подводное плавание с аквалангом и подводное плавание. международные проекты, такие как Green Fins , которые поощряют центры дайвинга и снорклинга следовать Кодексу поведения, снижают эти риски. Такая деятельность может нанести ущерб кораллам, но доказано, что [128]
Ювелирные изделия
[ редактировать ]
Разнообразие цветов кораллов делает их привлекательными для ожерелий и других украшений . Ярко-красный коралл ценится как драгоценный камень. Иногда его называют огненным кораллом, но это не то же самое, что огненный коралл . Красный коралл очень редок из-за чрезмерного сбора урожая . [129] В целом нецелесообразно дарить кораллы, поскольку они приходят в упадок из-за таких факторов стресса, как изменение климата, загрязнение окружающей среды и неустойчивое рыболовство.
Всегда считавшийся драгоценным минералом, «китайцы издавна ассоциировали красный коралл с благоприятностью и долголетием из-за его цвета и сходства с оленьими рогами (то есть по ассоциации, добродетели, долгой жизни и высокому рангу»). [130] Пика своей популярности он достиг во времена маньчжурской династии или династии Цин (1644-1911), когда он почти исключительно предназначался для использования императором либо в виде коралловых бус (часто в сочетании с жемчугом) для придворных украшений, либо в качестве декоративных пенцзин (декоративных миниатюр). минеральные деревья). Коралл был известен как шаньху по-китайски. «Ранняя современная «коралловая сеть» [началась] в Средиземном море [и попала] в цинский Китай через английскую Ост-Индскую компанию ». [131] в 1759 году , существовали строгие правила его использования В кодексе, установленном императором Цяньлуном .
Лекарство
[ редактировать ]
В медицине химические соединения кораллов потенциально могут быть использованы для лечения рака, неврологических заболеваний, воспалений, включая артрит, боли, потери костной массы, высокого кровяного давления и для других терапевтических целей. [133] [134] Скелеты кораллов, например, Isididae, исследуются на предмет их потенциального использования в ближайшем будущем для костной пластики у людей. [135] как Праваль Бхасма Коралловый калькс, известный на санскрите , широко используется в традиционной системе индийской медицины в качестве добавки при лечении различных метаболических нарушений костей, связанных с дефицитом кальция. [136] принимать внутрь измельченный коралл, который состоит в основном из слабоосновного карбоната кальция рекомендовали для успокоения язв желудка В классические времена Гален и Диоскорид . [137]
Строительство
[ редактировать ]Коралловые рифы в таких местах, как побережье Восточной Африки, используются в качестве источника строительного материала . [138] Древний (ископаемый) коралловый известняк, в частности, формация Коралловая тряпка на холмах вокруг Оксфорда (Англия), когда-то использовался в качестве строительного камня, и его можно увидеть в некоторых из старейших зданий этого города, включая саксонскую башню Святого Михаила. у Нортгейта , башни Св. Георгия в Оксфордском замке и средневековых стен города. [139]
Защита береговой линии
[ редактировать ]Здоровые коралловые рифы поглощают 97 процентов энергии волн, защищая береговую линию от течений, волн и штормов, помогая предотвратить гибель людей и материальный ущерб. Береговые линии, защищенные коралловыми рифами, также более устойчивы к эрозии, чем те, которых нет. [140]
Местная экономика
[ редактировать ]Прибрежные сообщества вблизи коралловых рифов во многом зависят от них. Во всем мире более 500 миллионов человек зависят от коралловых рифов в плане пропитания, дохода, защиты прибрежных зон и многого другого. [141] Общая экономическая стоимость услуг коралловых рифов в Соединенных Штатах, включая рыболовство, туризм и защиту побережья, составляет более 3,4 миллиарда долларов в год.
Аквариум
[ редактировать ]
В последние годы хобби по разведению морской рыбы расширилось и теперь включает рифовые аквариумы , аквариумы, в которых содержится большое количество живых камней , на которых кораллы могут расти и распространяться. [142] Эти резервуары либо содержатся в естественном состоянии, с водорослями (иногда в форме очистителя из водорослей ) и глубоким песчаным слоем, обеспечивающим фильтрацию, [143] или как «танки для выступлений», где скала в основном лишена водорослей и микрофауны , которые обычно ее населяют, [144] чтобы выглядеть опрятно и чисто.
Самый популярный вид сохраняемых кораллов — мягкие кораллы , особенно зоантиды и грибообразные кораллы, которые особенно легко выращивать и размножать в самых разных условиях, поскольку они происходят из закрытых частей рифов, где водные условия меняются, а освещение может быть менее надежным. и прямой. [145] Более серьезные рыболовы могут держать небольшие каменистые кораллы- полипы , которые происходят из открытых, ярко освещенных рифов и, следовательно, гораздо более требовательны, в то время как каменные кораллы-полипы представляют собой своего рода компромисс между ними.
Аквакультура
[ редактировать ]Коралловая аквакультура , также известная как коралловое выращивание или коралловое садоводство , представляет собой выращивание кораллов в коммерческих целях или восстановление коралловых рифов. Аквакультура обещает стать потенциально эффективным инструментом восстановления коралловых рифов , численность которых во всем мире сокращается. [146] [147] [148] Этот процесс обходит ранние стадии роста кораллов, когда они наиболее подвержены риску гибели. Фрагменты кораллов, известные как «семена», выращиваются в питомниках, а затем пересаживаются на риф. [149] Кораллы выращиваются коралловыми фермерами, которые живут поблизости от рифов и занимаются сельским хозяйством для сохранения рифов или для получения дохода. Его также выращивают ученые для исследований, предприятия по поставке живых и декоративных кораллов, а также частные аквариумов любители .
Галерея
[ редактировать ]Дополнительные изображения: Commons:Категория:Коралловые рифы и Commons:Категория:Кораллы
 Грибки сп. скелет
Грибки сп. скелет Полипы Eusmilia fastigiata
Полипы Eusmilia fastigiata Столбчатый коралл Dendrogyra cylindricus
Столбчатый коралл Dendrogyra cylindricus Мозговой коралл Diploria labyrinthiformis.
Мозговой коралл Diploria labyrinthiformis. Мозговой коралл, откладывающий яйца
Мозговой коралл, откладывающий яйца










См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Русе, Элоиза; Галанд, Пьер Э.; Медина, Моника; Бонгертс, Пим; Смолл, Майкл; Перес-Росалес, Гонсало; Торда, Гергели; Мойя, Орели; Райна, Жан-Батист; Эдуэн, Летиция (9 мая 2021 г.). «Симбиотические ассоциации самых глубоких зарегистрированных фотосинтезирующих склерактиновых кораллов (глубина 172 м)» . Журнал ISME . 15 (5): 1564–1568. Бибкод : 2021ISMEJ..15.1564R . дои : 10.1038/s41396-020-00857-y . ПМК 8115523 . ПМИД 33452473 .
- ^ Сквайрс, Д.Ф. (1959). «Глубоководные кораллы, собранные Геологической обсерваторией Ламонта. 1. Атлантические кораллы» (PDF) . Американский музей Novitates (1965): 23.
- ^ Леруа, Арман Мари (2014). Лагуна: как Аристотель изобрел науку . Блумсбери. п. 271. ИСБН 978-1-4088-3622-4 .
- ^ Перейти обратно: а б с Боуэн, Джеймс (2015). Эра коралловых рифов: от открытия к упадку: история научных исследований с 1600 года до эпохи антропоцена . Спрингер. стр. 5–7. ISBN 978-3-319-07479-5 .
- ^ Вавилонский Талмуд, Рош ха-Шана 23а и комментарий Раши (24-я узкая строка)
- ^ Эгертон, Фрэнк Н. (2012). Корни экологии: от древности до Хакеля . Издательство Калифорнийского университета. п. 24. ISBN 978-0-520-95363-5 .
- ^ Светт, К. (5 марта 2020 г.). Кораллы: тайны их рифообразующих колоний . Capstone Global Library Ltd. ISBN 9781474771009 .
- ^ Хоксема, Берт (2015). «Антозоа» . ВОРМС . Всемирный регистр морских видов . Проверено 24 апреля 2015 г.
- ^ Перейти обратно: а б с д Рамирес-Портилья, Каталина; Бэрд, Эндрю Х; Коуман, Питер Ф; Кваттрини, Андреа М; Харии, Саки; Синнигер, Фредерик; Фло, Жан-Франсуа (01 марта 2022 г.). «Решение загадки разграничения видов кораллов» . Систематическая биология . 71 (2): 461–475. дои : 10.1093/sysbio/syab077 . ISSN 1063-5157 . ПМИД 34542634 .
- ^ Ричи, Ханна; Розер, Макс (15 апреля 2021 г.). «Биоразнообразие» . Наш мир в данных .
- ^ Хьюм, Бенджамин CC; Д'Анджело, Сесилия; Берт, Джон А.; Виденманн, Йорг (2018). «Мелкомасштабное определение биогеографических границ и разрешение субпопуляций в группе коралловых симбионтов Symbiodinium thermophilum из Персидского/Арабского залива и Оманского залива» . Границы морской науки . 5 . дои : 10.3389/fmars.2018.00138 . hdl : 10754/627647 . ISSN 2296-7745 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Хемонд, Элизабет М; Калузяк, Стефан Т; Воллмер, Стивен В. (17 декабря 2014 г.). «Генетика формы и функционирования колоний карибских кораллов Acropora» . БМК Геномика . 15 : 1133. дои : 10.1186/1471-2164-15-1133 . ISSN 1471-2164 . ПМЦ 4320547 . ПМИД 25519925 .
- ^ Перейти обратно: а б Министерство торговли США, Национальное управление океанических и атмосферных исследований. «Программа сохранения коралловых рифов NOAA (CRCP) — факты о кораллах» . coralreef.noaa.gov . Проверено 26 апреля 2022 г.
- ^ Перейти обратно: а б «Мягкие кораллы: они похожи на растения, но на самом деле являются животными» . МысльКо . Проверено 26 апреля 2022 г.
- ^ ДеВиктор, Сьюзен Т.; Мортон, Стив Л. (2010). «Морфология октокораллов» . Путеводитель по мелководным (0-200 м) восьмикоралам Южно-Атлантического залива . Окленд, Новая Зеландия: Магнолия Пресс. ISBN 978-1-86977-584-1 .
- ^ Гилберт, Скотт Ф. (2000). «Эмбриональное развитие» . Биология развития (6-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 0-87893-243-7 .
- ^ Эрик М., Энгстрем; Ижаки, Анат; Боуман, Джон Л. (июль 2004 г.). «Избиение промоутеров, микроРНК и гены Knox. Новые идеи, регуляторы и цели регулирования в установлении полярности латеральных органов у Arabidopsis» . Физиология растений . 135 (2): 685–694. дои : 10.1104/стр.104.040394 . ПМК 514105 . PMID 15208415 – через ResearchGate.
- ^ Стеммер, Кристина; Бургхардт, Инго; Майер, Кристоф; Райнике, Гетц Б.; Вегеле, Хайке; Толлриан, Ральф; Лиз, Флориан (июнь 2013 г.). «Морфологический и генетический анализ разнообразия мягких кораллов ксениид (Octocorallia; Alcyonacea)» . Разнообразие и эволюция организмов . 13 (2): 135–150. дои : 10.1007/s13127-012-0119-x . ISSN 1439-6092 . S2CID 17511020 .
- ^ «Коралловые рифы» . МСОП . Архивировано из оригинала 27 апреля 2022 г. Проверено 27 апреля 2022 г.
- ^ Перейти обратно: а б Штраус, Хлоя; Лонг, Хонган; Паттерсон, Кейтлин Э.; Те, Рональд; Линч, Майкл (6 сентября 2017 г.). Моран, Нэнси А. (ред.). «Реакция скорости полногеномных мутаций на изменение pH у возбудителя коралловых рифов Vibrio shilonii AK1» . мБио . 8 (4): e01021–17. дои : 10.1128/mBio.01021-17 . ISSN 2161-2129 . ПМЦ 5565966 . ПМИД 28830944 .
- ^ Перейти обратно: а б с Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание . Cengage Обучение. стр. 132–48. ISBN 978-81-315-0104-7 .
- ^ Перейти обратно: а б с д Одно или несколько предыдущих предложений включают текст из публикации, которая сейчас находится в свободном доступе : Минчин, Эдвард Альфред (1911). « Полип ». В Чисхолме, Хью (ред.). Британская энциклопедия . Том. 22 (11-е изд.). Издательство Кембриджского университета. п. 37.
- ^ Д. Гатено; А. Израиль; Ю. Барки; Б. Ринкевич (1998). «Гастроваскулярное кровообращение у восьмикораллов: свидетельства значительного транспорта кораллов и клеток-симбионтов» . Биологический вестник . 194 (2): 178–86. дои : 10.2307/1543048 . JSTOR 1543048 . ПМИД 28570841 . S2CID 19530967 .
- ^ Пил, Джон С. (2017). «Проблемный книдарий (Cambroctoconus; Octocorallia?) из кембрия (серии 2–3) Лаврентии» . Журнал палеонтологии . 91 (5): 871–882. Бибкод : 2017JPal...91..871P . дои : 10.1017/jpa.2017.49 . S2CID 134826884 .
- ^ Администрация Министерства торговли США, Национальная служба океанических и атмосферных исследований. «существующая и потенциальная ценность коралловых экосистем с точки зрения дохода и других экономических ценностей» . coralreef.noaa.gov . Архивировано из оригинала 05 февраля 2018 г. Проверено 4 февраля 2018 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Спрунг, Джулиан (1999). Кораллы: Краткое справочное руководство . Издательство Рикордеа. п. 145. ИСБН 978-1-883693-09-1 .
- ^ Куиф, Япония; Дофин, Ю. (1998). «Микроструктурная и физико-химическая характеристика« центров кальцификации »в перегородках некоторых современных склерактиновых кораллов». Палеонтологическая газета . 72 (3–4): 257–269. Бибкод : 1998PalZ...72..257C . дои : 10.1007/bf02988357 . ISSN 0031-0220 . S2CID 129021387 .
- ^ Куиф, Япония; Дофин, Ю.; Дусе, Дж.; Саломе, М.; Сусини, Дж. (2003). «XANES-картирование органических сульфатов в трех скелетах склерактиновых кораллов». Geochimica et Cosmochimica Acta . 67 (1): 75–83. Бибкод : 2003GeCoA..67...75C . дои : 10.1016/s0016-7037(02)01041-4 . ISSN 0016-7037 .
- ^ Дофин, Ю.; Куиф, Япония; Уильямс, Коннектикут (2008). «Растворимые органические матрицы арагонитовых скелетов мерулинид (Cnidaria, Anthozoa)». Сравнительная биохимия и физиология. Часть B: Биохимия и молекулярная биология . 150 (1): 10–22. дои : 10.1016/j.cbpb.2008.01.002 . ISSN 1096-4959 . ПМИД 18325807 .
- ^ Куиф, Япония; Дофин, Ю.; Фрейвальд, А.; Готре, П.; Зибровиус, Х. (1999). «Биохимические маркеры симбиоза зооксантелл в растворимых матрицах скелета 24 видов Scleractinia». Сравнительная биохимия и физиология. Часть A: Молекулярная и интегративная физиология . 123 (3): 269–278. дои : 10.1016/s1095-6433(99)00059-8 . ISSN 1095-6433 .
- ^ Перейти обратно: а б Мерфи, Ричард К. (2002). Коралловые рифы: города под морями . Дарвин Пресс. ISBN 978-0-87850-138-0 .
- ^ Юяма, Икуко (2014). «Сравнение влияния симбиотических водорослей (Symbiodinium) клад C1 и D на ранние стадии роста Acropora tenuis» . ПЛОС ОДИН . 9 (6): е98999. Бибкод : 2014PLoSO...998999Y . дои : 10.1371/journal.pone.0098999 . ПМК 4051649 . ПМИД 24914677 .
- ^ Ямасита, Хироши (2014). «Создание кораллово-водорослевого симбиоза требует привлечения и отбора» . ПЛОС ОДИН . 9 (5): e97003. Бибкод : 2014PLoSO...997003Y . дои : 10.1371/journal.pone.0097003 . ПМК 4019531 . ПМИД 24824794 .
- ^ «Зооксантеллы... Что это?» . Образование в области океанических услуг NOAA . Национальное управление океанических и атмосферных исследований . Проверено 1 декабря 2017 г.
- ^ Перейти обратно: а б Мадл, П.; Йип, М. (2000). «Полевая экскурсия в провинцию Милн-Бэй – Папуа-Новая Гвинея» . Архивировано из оригинала 11 мая 2020 г. Проверено 31 марта 2006 г.
- ^ ван де Плааше, Орсон (1986). Исследование уровня моря: руководство по сбору и оценке данных . Норидж, Великобритания: Geo Books. п. 196. ИСБН 978-94-010-8370-6 .
- ^ «Кораллы и их микробиомы развивались вместе | Университет штата Пенсильвания» . www.psu.edu .
- ^ В.В. Толлер; Р. Роуэн; Н. Ноултон (2001). «Репопуляция зооксантелл в карибских кораллах Montastraea annularis и M. faveolata после экспериментального и связанного с болезнями обесцвечивания» . Биологический вестник . 201 (3): 360–73. дои : 10.2307/1543614 . JSTOR 1543614 . ПМИД 11751248 . S2CID 7765487 . Архивировано из оригинала 25 февраля 2006 г. Проверено 30 марта 2006 г.
- ^ Браунли, Колин (2009). «Регуляция pH у симбиотических анемонов и кораллов: тонкий баланс» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (39): 16541–16542. Бибкод : 2009PNAS..10616541B . дои : 10.1073/pnas.0909140106 . ПМЦ 2757837 . ПМИД 19805333 .
- ^ Ландсберг и др., «Болезнь потери ткани каменистых кораллов во Флориде связана с нарушением физиологии хозяина-зооксантелл».
- ^ де О Сантос и др., «Геномный и протеомный анализ кораллового возбудителя Vibrio Coralliilyticus выявляет разнообразный репертуар вирулентности».
- ^ Авис, Джон К. (18 марта 2011 г.). Гермафродитизм: учебник по биологии, экологии и эволюции двойной сексуальности . Издательство Колумбийского университета. п. 83. ИСБН 978-0-231-52715-6 .
- ^ Перейти обратно: а б с д и ж Маркандейя, Вират (22 февраля 2023 г.). «Как лунные циклы управляют нерестом кораллов, червей и многого другого» . Знающий журнал . Ежегодные обзоры. doi : 10.1146/knowable-022223-2 . S2CID 257126558 . Проверено 6 марта 2023 г.
- ^ Перейти обратно: а б с д Хефкер, Н. Сёрен; Андреатта, Габриэле; Манцотти, Алессандро; Фальсиаторе, Анжела; Райбл, Флориан; Тессмар-Райбл, Кристин (16 января 2023 г.). «Ритмы и часы в морских организмах» . Ежегодный обзор морской науки . 15 (1): 509–538. Бибкод : 2023ARMS...15..509H . doi : 10.1146/annurev-marine-030422-113038 . ISSN 1941-1405 . ПМИД 36028229 . S2CID 251865474 .
- ^ Перейти обратно: а б с Верон, ДЖЕН (2000). Кораллы мира. Том 3 (3-е изд.). Австралия: Австралийский институт морских наук и CRR Qld. ISBN 978-0-642-32236-4 .
- ^ Хатта, М.; Фуками, Х.; Ван, В.; Омори, М.; Симойке, К.; Хаясибара, Т.; Ина, Ю.; Сугияма, Т. (1999). «Репродуктивные и генетические доказательства сетчатой эволюционной теории массового нереста кораллов» . Молекулярная биология и эволюция . 16 (11): 1607–13. doi : 10.1093/oxfordjournals.molbev.a026073 . ПМИД 10555292 .
- ^ Перейти обратно: а б Лин, Че-Хунг; Такахаши, Шуничи; Мулла, Азиз Дж.; Нодзава, Йоко (24 августа 2021 г.). «Время восхода луны является ключевым моментом для синхронного нереста кораллов Dipsastraea speciosa» . Труды Национальной академии наук . 118 (34): e2101985118. Бибкод : 2021PNAS..11801985L . дои : 10.1073/pnas.2101985118 . ISSN 0027-8424 . ПМЦ 8403928 . ПМИД 34373318 .
- ^ Джабр, Феррис (13 июня 2017 г.). «Лунное море» . Журнал Хакай . Проверено 6 марта 2023 г.
- ^ Перейти обратно: а б Барнс, Р. и; Хьюз, Р. (1999). Введение в морскую экологию (3-е изд.). Молден, Массачусетс: Блэквелл. стр. 117–41. ISBN 978-0-86542-834-8 .
- ^ Кэмерон, Керри А.; Харрисон, Питер Л. (26 марта 2020 г.). «Плотность личинок кораллов может влиять на заселение, численность колоний после поселения и коралловый покров при восстановлении личинок» . Научные отчеты . 10 (1): 5488. Бибкод : 2020NatSR..10.5488C . дои : 10.1038/s41598-020-62366-4 . ISSN 2045-2322 . ПМК 7099096 . ПМИД 32218470 .
- ^ Аялон, Инбал; Розенберг, Яэли; Бенишу, Дженнифер И.С.; Кампос, Селин Луиза Д.; Сайко, Шерри Лин Дж.; Нада, Майкл Анжелу Л.; Бакиран, Джейк Иван П.; Лигсон, Чарлон А.; Ависар, Дрор; Конако, Сесилия; Куечли, Хельга У.; Киба, Кристофер СМ; Кабейтан, Патрик С.; Леви, Орен (25 января 2021 г.). «Коллапс гаметогенеза кораллов при искусственном световом загрязнении» . Современная биология . 31 (2): 413–419.e3. дои : 10.1016/j.cub.2020.10.039 . ISSN 0960-9822 . ПМИД 33157030 . S2CID 226257589 .
- ^ Сузуки, Го; Окада, Ватару; Ясутаке, Йоко; Ямамото, Хидекадзу; Танита, Ивао; Ямасита, Хироши; Хаясибара, Такеши; Комацу, Тошиаки; Каньяма, Тору; Иноуэ, Масахито; Ямадзаки, Масаси (сентябрь 2020 г.). «Увеличение запасов личинок кораллов и производства саженцев с использованием специальной системы сбора связок «колыбель для личинок кораллов» для крупномасштабного восстановления кораллов» . Реставрационная экология . 28 (5): 1172–1182. Бибкод : 2020ResEc..28.1172S . дои : 10.1111/рек.13178 . ISSN 1061-2971 . S2CID 218796945 .
- ^ Джонс, ОА; Эндин, Р. (1973). Биология и геология коралловых рифов . Нью-Йорк, США: Харкорт Брейс Йованович. стр. 205–45. ISBN 978-0-12-389602-5 .
- ^ ХАРРИСОН, PL; УОЛЛЕС, CC (1990). «Размножение, расселение и пополнение склерактиновых кораллов» . Размножение, расселение и пополнение склерактиновых кораллов . 25 : 133–207. ISSN 0167-4579 .
- ^ Морс, Дэниел Э.; Хукер, Нил; Морс, Эйлин, Северная Каролина; Дженсен, Ребекка А. (24 мая 1988 г.). «Контроль метаморфоза личинок и пополнение в симпатрических агарицидных кораллах» . Журнал экспериментальной морской биологии и экологии . 116 (3): 193–217. дои : 10.1016/0022-0981(88)90027-5 . ISSN 0022-0981 .
- ^ Вебстер, Николь С.; Смит, Люк Д.; Хейворд, Эндрю Дж.; Уоттс, Джой Э.М.; Уэбб, Ричард И.; Блэколл, Линда Л .; Негри, Эндрю П. (1 февраля 2004 г.). «Метаморфоза склерактинового коралла в ответ на микробные биопленки» . Прикладная и экологическая микробиология . 70 (2): 1213–1221. Бибкод : 2004ApEnM..70.1213W . дои : 10.1128/АЕМ.70.2.1213-1221.2004 . ISSN 0099-2240 . ПМК 348907 . ПМИД 14766608 .
- ^ Рикардо, Джерард Ф.; Джонс, Росс Дж.; Нордборг, Микаэла; Негри, Эндрю П. (31 декабря 2017 г.). «Схемы расселения коралла Acropora millepora на поверхности, заполненной отложениями» . Наука об общей окружающей среде . 609 : 277–288. Бибкод : 2017ScTEn.609..277R . doi : 10.1016/j.scitotenv.2017.07.153 . ISSN 0048-9697 . ПМИД 28750231 .
- ^ Биррелл, CL; МакКук, LJ; Уиллис, БЛ; Харрингтон, Л. (30 июня 2008 г.). «Химическое воздействие макроводорослей на поселение личинок нерестящегося коралла Acropora millepora» . Серия «Прогресс в области морской экологии» . 362 : 129–137. Бибкод : 2008MEPS..362..129B . дои : 10.3354/meps07524 . ISSN 0171-8630 .
- ^ Гулько, Дэвид (1998). Экология гавайских коралловых рифов . Гонолулу, Гавайи: Взаимное издательство. п. 10. ISBN 978-1-56647-221-0 .
- ^ Шеппард, Чарльз Р.К.; Дэви, Саймон К.; Пиллинг, Грэм М. (25 июня 2009 г.). Биология коралловых рифов . ОУП Оксфорд. стр. 78–81. ISBN 978-0-19-105734-2 .
- ^ Шарп, Коти Х.; Пратте, Зои А.; Кервин, Эллисон Х.; Ротьян, Рэнди Д.; Стюарт, Фрэнк Дж. (2017). «Структурой микробиома коралла Astrangia poculata умеренного пояса управляет сезон, а не состояние симбионта» . Микробиом . 5 (1): 120. дои : 10.1186/s40168-017-0329-8 . ПМК 5603060 . ПМИД 28915923 .
- ^ Дубинский З. и Джокиел П.Л. (1994) «Соотношение потоков энергии и питательных веществ регулирует симбиоз между зооксантеллами и кораллами». Pacific Science , 48 (3): 313–324.
- ^ Энтони, К.Р., Клайн, Д.И., Диас-Пулидо, Г., Дав, С. и Хоег-Гульдберг, О. (2008) «Закисление океана вызывает обесцвечивание и потерю производительности строителей коралловых рифов». Труды Национальной академии наук , 105 (45): 17442–17446. дои : 10.1073/pnas.0804478105 .
- ^ Перейти обратно: а б Апприл, А. (2017) «Микробиомы морских животных: к пониманию взаимодействия хозяина и микробиома в меняющемся океане». Границы морской науки , 4 : 222. два : 10.3389/fmars.2017.00222 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Борн, Д.Г., Морроу, К.М. и Вебстер, Н.С. (2016) «Изучение кораллового микробиома: поддержка здоровья и устойчивости рифовых экосистем». Ежегодный обзор микробиологии , 70 : 317–340. doi : 10.1146/annurev-micro-102215-095440 .
- ^ Нив, М.Дж., Апприлль, А., Ферье-Пажес, К. и Вулстра, К.Р. (2016) «Разнообразие и функции распространенных симбиотических морских бактерий рода Endozoicomonas ». Прикладная микробиология и биотехнология , 100 (19): 8315–8324. два : 10.1007/s00253-016-7777-0 .
- ^ Нив, М.Дж., Мичелл, К.Т., Апприлл, А. и Вулстра, К.Р. (2017) «Геномы Endozoicomonas обнаруживают функциональную адаптацию и пластичность бактериальных штаммов, симбиотически связанных с различными морскими хозяевами». Научные отчеты , 7 : 40579. дои : 10.1038/srep40579 .
- ^ Хьюз, Т.П., Керри, Дж.Т., Альварес-Норьега, М., Альварес-Ромеро, Дж.Г., Андерсон, К.Д., Бэрд, А.Х., Бэбкок, Р.С., Бегер, М., Беллвуд, Д.Р., Беркельманс, Р. и Бридж, ТК (2017) «Глобальное потепление и периодическое массовое обесцвечивание кораллов». Природа , 543 (7645): 373–377. дои : 10.1038/nature21707 .
- ^ Ученые Геологической службы США публикуют давно читаемые последовательности микробиома кораллов умеренного пояса, предоставляя сообществу ресурсы для разработки зондов и праймеров , Геологическая служба США , 6 марта 2019 г. Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ б. Голдсмит, Дон; а. Пратте, Зоя; а. Келлог, Кристина; е. Снадер, Сара; час Шарп, Коти (2019). «Стабильность микробиома коралла Astrangia poculata умеренного пояса отражается в различных методологиях секвенирования» . АИМС Микробиология . 5 (1): 62–76. дои : 10.3934/микробиол.2019.1.62 . ПМК 6646935 . ПМИД 31384703 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Ноултон, Н. и Ровер, Ф. (2003) «Многовидовой микробный мутуализм на коралловых рифах: хозяин как среда обитания». Американский натуралист , 162 (S4): S51-S62. дои : 10.1086/378684 .
- ^ Поллок, Ф. Джозеф; МакМиндс, Райан; Смит, Стайлз; Борн, Дэвид Г.; Уиллис, Бетт Л.; Медина, Моника; Тербер, Ребекка Вега; Заневельд, Джесси Р. (22 ноября 2018 г.). «Бактерии, ассоциированные с кораллами, демонстрируют филосимбиоз и кофилогению» . Природные коммуникации . 9 (1): 4921. Бибкод : 2018NatCo...9.4921P . дои : 10.1038/s41467-018-07275-x . ISSN 2041-1723 . ПМК 6250698 . ПМИД 30467310 .
- ^ Пейшото, Р.С., Росадо, П.М., Лейте, DCDA, Росадо, А.С. и Борн, Д.Г. (2017) «Полезные микроорганизмы для кораллов (BMC): предлагаемые механизмы здоровья и устойчивости кораллов». Границы микробиологии , 8 : 341. два : 10.3389/fmicb.2017.00341 .
- ^ Апприлл, Эми; Вебер, Лаура Г.; Санторо, Элисон Э. (2016). «Различие среды обитания микробов раскрывает экологическую сложность коралловых микробиомов» . mSystems . 1 (5): e00143–16. дои : 10.1128/mSystems.00143-16 . ПМК 5080407 . ПМИД 27822559 .
- ^ Ла Ривьер, Мари; Гаррабоу, Хоаким; Балли, Марк (01 декабря 2015 г.). «Доказательства специфичности хозяина среди доминирующих бактериальных симбионтов горгониевых кораллов умеренного климата» . Коралловые рифы . 34 (4): 1087–1098. Бибкод : 2015CorRe..34.1087L . дои : 10.1007/s00338-015-1334-7 . ISSN 1432-0975 . S2CID 14309443 .
- ^ ван де Уотер, Йерун AJM; Мелконян, Реми; Вулстра, Кристиан Р.; Джунка, Ховард; Беро, Эрик; Аллеманд, Денис; Ферье-Пажес, Кристина (01 февраля 2017 г.). «Сравнительная оценка средиземноморских горгонарийных микробных сообществ выявила консервативность основных и локальных вариантов бактерий» . Микробная экология . 73 (2): 466–478. Бибкод : 2017MicEc..73..466V . дои : 10.1007/s00248-016-0858-x . ISSN 1432-184Х . ПМИД 27726033 . S2CID 22336906 .
- ^ Поллок, Ф. Джозеф; МакМиндс, Райан; Смит, Стайлз; Борн, Дэвид Г.; Уиллис, Бетт Л.; Медина, Моника; Тербер, Ребекка Вега; Заневельд, Джесси Р. (22 ноября 2018 г.). «Бактерии, ассоциированные с кораллами, демонстрируют филосимбиоз и кофилогению» . Природные коммуникации . 9 (1): 4921. Бибкод : 2018NatCo...9.4921P . дои : 10.1038/s41467-018-07275-x . ISSN 2041-1723 . ПМК 6250698 . ПМИД 30467310 .
- ^ Томпсон, Дж.Р., Ривера, Х.Э., Клосек, С.Дж. и Медина, М. (2015) «Микробы в коралловом холобионте: партнеры в эволюции, развитии и экологических взаимодействиях». Границы клеточной и инфекционной микробиологии , 4 : 176. два : 10.3389/fcimb.2014.00176 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Хомпсон, Дж.Р., Ривера, Х.Э., Клосек, С.Дж. и Медина, М. (2015) «Микробы в коралловом холобионте: партнеры в эволюции, развитии и экологических взаимодействиях». Границы клеточной и инфекционной микробиологии , 4 : 176. два : 10.3389/fcimb.2014.00176 .
- ^ Шумахер, Гельмут; Зибровиус, Хельмут (1985). «Что такое герматипический?». Коралловые рифы . 4 (1): 1–9. Бибкод : 1985КорРе...4....1С . дои : 10.1007/BF00302198 . S2CID 34909110 .
- ^ MSN Энкарта (2006). Большой Барьерный риф . Архивировано из оригинала 28 октября 2009 года . Проверено 25 апреля 2015 г.
- ^ Сполдинг, Марк; Равилиус, Коринна; Грин, Эдмунд (2001). Мировой атлас коралловых рифов . Беркли, Калифорния: Издательство Калифорнийского университета и ЮНЕП/WCMC. стр. 205–45 . ISBN 978-0-520-23255-6 .
- ^ Смитсоновский национальный музей flickr .
- ^ Олден, Эндрю. «Индексные окаменелости» . Об образовании. Архивировано из оригинала 1 декабря 2016 года . Проверено 25 апреля 2015 г.
- ^ Пратт, БР; Спинсер, БР; Вуд, РА; Журавлёв, А.Ю. (2001). «12: Экология и эволюция кембрийских рифов» (PDF) . Экология кембрийской радиации . Издательство Колумбийского университета. п. 259. ИСБН 978-0-231-10613-9 . Проверено 6 апреля 2007 г. [ постоянная мертвая ссылка ]
- ^ Винн, О.; Мытус, М.-А. (2008). «Самые ранние эндосимбиотические минерализованные трубчатые черви из силура Подолья, Украина» . Журнал палеонтологии . 82 (2): 409–14. Бибкод : 2008JPal...82..409V . дои : 10.1666/07-056.1 . S2CID 131651974 . Проверено 11 июня 2014 г.
- ^ Винн, О.; Мытус, М.-А. (2012). «Разнообразный комплекс ранних эндобиотических коралловых симбионтов из Катиана (позднего ордовика) Балтики». Палеогеография, Палеоклиматология, Палеоэкология . 321–322: 137–41. Бибкод : 2012PPP...321..137В . дои : 10.1016/j.palaeo.2012.01.028 .
- ^ Перейти обратно: а б «Введение в Табулату» . UCMP Беркли. Архивировано из оригинала 19 апреля 2015 года . Проверено 25 апреля 2015 г.
- ^ «Знакомство с Ругосой» . UCMP Беркли. Архивировано из оригинала 19 апреля 2015 года . Проверено 25 апреля 2015 г.
- ^ Сян-Донг Ван и Сяо-Хуан Ван (2007). «Схемы вымирания позднепермских (лопинских) кораллов в Китае» , Palaeoworld, 16, № 1–3, 31–38.
- ^ Райс Дж.Б., Стэнли С.М., Харди Л.А. (июль 2006 г.). «Склерактиновые кораллы производят кальцит и растут медленнее в искусственной морской воде мелового периода». Геология . 34 (7): 525–28. Бибкод : 2006Geo....34..525R . дои : 10.1130/G22600.1 .
- ^ «Эволюционная история» . ЦЕЛИ . Архивировано из оригинала 13 октября 2018 года . Проверено 25 апреля 2015 г.
- ^ Ваггонер, Бен М. (2000). Смит, Дэвид; Коллинз, Аллен (ред.). «Антозоа: Летопись окаменелостей» . Антозоа . УКМП . Проверено 9 марта 2020 г.
- ^ Оливер, Уильям А. младший (2003). «Кораллы: Таблица 1» . Ископаемые группы . Геологическая служба США . Архивировано из оригинала 9 января 2009 года . Проверено 9 марта 2020 г.
- ^ «Коралловые рифы мира» . Guardian.co.uk . 2 сентября 2009 г.
- ^ Перейти обратно: а б «Угрозы коралловым рифам» . Альянс коралловых рифов . 2010. Архивировано из оригинала 1 декабря 2011 года . Проверено 5 декабря 2011 г.
- ^ Потеря наших коралловых рифов – Экологические вопросы – Состояние планеты . Блоги.ei.columbia.edu. Проверено 1 ноября 2011 г.
- ^ Клейпас, Дж.А.; Фили, РА; Фабри, виджей; Лэнгдон, К.; Сабина, CL; Роббинс, LL (2006). «Воздействие закисления океана на коралловые рифы и другие морские кальцификаторы: руководство для будущих исследований» (PDF) . Национальный научный фонд , НОАА и Геологическая служба США . Архивировано из оригинала (PDF) 20 июля 2011 года . Проверено 7 апреля 2011 г.
- ^ Спасите наши моря, Летний информационный бюллетень 1997 г., Синди Хантер и Алан Фридлендер
- ^ Тун, К.; Чжоу, Л.М.; Кабанбан, А.; Туан, В.С.; Филрифс, С.; Йемин, Т.; Сухарсоно; Кислый, К.; Лейн, Д. (2004). «Состояние коралловых рифов, мониторинг и управление коралловыми рифами в Юго-Восточной Азии, 2004 г.» . В Уилкинсоне, К. (ред.). Состояние коралловых рифов мира: 2004 г. Таунсвилл, Квинсленд, Австралия: Австралийский институт морских наук. стр. 235–76 . Проверено 23 апреля 2019 г.
- ^ Берк, Лауретта; Рейтар, К.; Сполдинг, М.; Перри, А. (2011). Повторное посещение рифов, находящихся под угрозой . Вашингтон, округ Колумбия: Институт мировых ресурсов. п. 38 . ISBN 978-1-56973-762-0 .
- ^ Брайант, Дирк; Берк, Лауретта; Макманус, Джон; Сполдинг, Марк. «Рифы в опасности: картографический индикатор угроз коралловым рифам мира» (PDF) . НОАА. Архивировано из оригинала (PDF) 18 февраля 2013 г. Проверено 25 апреля 2015 г.
- ^ Норландер (8 декабря 2003 г.). «Коралловый кризис! Люди уничтожают эти шумные подводные города. Можно ли спасти коралловые рифы? (Наука о жизни: кораллы)» . Мир науки .
- ^ Рашер Д.Б., Хэй МЭ (май 2010 г.). «Химически богатые водоросли отравляют кораллы, если их не контролируют травоядные животные» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (21): 9683–88. Бибкод : 2010PNAS..107.9683R . дои : 10.1073/pnas.0912095107 . ПМЦ 2906836 . ПМИД 20457927 .
- ^ Хог-Гульдберг, О. (1999). «Изменение климата, обесцвечивание кораллов и будущее коралловых рифов мира». Морские и пресноводные исследования . 50 (8): 839–66. дои : 10.1071/MF99078 .
- ^ Перейти обратно: а б Стивенс, Тим (28 ноября 2011 г.). «Подводные источники позволяют увидеть влияние закисления океана на коралловые рифы» . Калифорнийский университет Санта-Крус . Проверено 25 апреля 2015 г.
- ^ «Признаки здоровья и болезней кораллового микробиома • iBiology» . iБиология . Проверено 14 мая 2020 г.
- ^ Хог-Гульдберг О. (1999). «Изменение климата, обесцвечивание кораллов и будущее коралловых рифов мира». Морские и пресноводные исследования . 50 (8): 839–99. дои : 10.1071/mf99078 .
- ^ Министерство торговли США, Национальное управление океанических и атмосферных исследований. «Как изменение климата влияет на коралловые рифы?» . Oceanservice.noaa.gov .
- ^ «Национальное управление океанических и атмосферных исследований – в ходе миссии, поддерживаемой НОАА, обнаружен новый глубоководный коралл» . www.noaanews.noaa.gov . Проверено 11 мая 2009 г.
- ^ Шраг, ДП; Линсли, БК (2002). «Кораллы, химия и климат». Наука . 296 (8): 277–78. дои : 10.1126/science.1071561 . ПМИД 11951026 . S2CID 82449130 .
- ^ Смитерс, Скотт Г.; Вудрофф, Колин Д. (2000). «Микроатоллы как индикаторы уровня моря на срединно-океаническом атолле». Морская геология . 168 (1–4): 61–78. Бибкод : 2000МГеол.168...61С . дои : 10.1016/S0025-3227(00)00043-8 .
- ^ Перейти обратно: а б с Хьюз, Т.; Бэрд, А.; Беллвуд, Д.; Кард, М.; Коннолли, С.; Фолке, К.; Гросберг, Р.; Хоэ-Гульдберг, О.; Джексон, Дж.; Клепас, Дж.; Лох, Дж.; Маршалл, П.; Нистром, М.; Палумби, С. ; Пандольфи, Дж.; Розен, Б.; и Рафгарден, Дж. (2003). «Изменение климата, воздействие человека и устойчивость коралловых рифов». Наука . 301 (5635): 929–33. Бибкод : 2003Sci...301..929H . дои : 10.1126/science.1085046 . ПМИД 12920289 . S2CID 1521635 .
- ^ Перейти обратно: а б Пармезан, К. (2006). «Экологические и эволюционные реакции на недавнее изменение климата». Ежегодный обзор экологии, эволюции и систематики . 37 : 637–69. doi : 10.1146/annurev.ecolsys.37.091305.110100 .
- ^ Бейкер, А. (2004). «Адаптивный ответ кораллов на изменение климата» . Природа . 430 (7001): 741. Бибкод : 2004Natur.430..741B . дои : 10.1038/430741a . ПМИД 15306799 . S2CID 32092741 .
- ^ Перейти обратно: а б Доннер, С.; Скирвинг, В.; Литтл, К.; Оппенгеймер, М.; Хог-Гульдберг, О. (2005). «Глобальная оценка обесцвечивания кораллов и необходимые темпы адаптации к изменению климата» (PDF) . Биология глобальных изменений . 11 (12): 2251–65. Бибкод : 2005GCBio..11.2251D . CiteSeerX 10.1.1.323.8134 . дои : 10.1111/j.1365-2486.2005.01073.x . ПМИД 34991281 . S2CID 84890014 . Архивировано из оригинала (PDF) 14 августа 2017 г. Проверено 25 октября 2017 г.
- ^ Баскетт, М.; Гейнс С. и Нисбет Р. (2009). «Разнообразие симбионтов может помочь коралловым рифам пережить умеренное изменение климата» (PDF) . Экологические приложения . 19 (1): 3–17. Бибкод : 2009EcoAp..19....3B . дои : 10.1890/08-0139.1 . ПМИД 19323170 . S2CID 1189125 .
- ^ МакКланахан, Т.; Атевеберхан, М.; Мухандо, К.; Майна Дж. и Мохаммед М. (2007). «Влияние климата и температуры морской воды на обесцвечивание кораллов и мораль». Экологические монографии . 77 (4): 503–25. Бибкод : 2007ЭкоМ...77..503М . CiteSeerX 10.1.1.538.970 . дои : 10.1890/06-1182.1 .
- ^ Килборн, К. Халимеда; Куинн, Терренс М.; Тейлор, Фредерик В.; Делькруа, Тьерри; Гуриу, Ив (2004). «Вариации солености, связанные с Эль-Ниньо и Южным колебанием, зафиксированные в геохимии скелета коралла Porites из Эспириту-Санто, Вануату» . Палеоокеанография . 19 (4): PA4002. Бибкод : 2004PalOc..19.4002K . дои : 10.1029/2004PA001033 .
- ^ Перейти обратно: а б Рен, Лей; Линсли, Брэддок К.; Веллингтон, Джерард М.; Шраг, Дэниел П.; Хоег-Гульдберг, Уве (2003). «Деконволюция δ 18 O компонент морской воды из субсезонных кораллов δ 18 O и Sr/Ca на Раротонге в юго-западной субтропической части Тихого океана за период с 1726 по 1997 год». Geochimica et Cosmochimica Acta . 67 (9): 1609–21. Бибкод : 2003GeCoA..67.1609R . doi : 10.1016/S0016-7037( 02)00917-1 .
- ^ Ву, Генри С.; Линсли, Брэддок К.; Дасси, Эмили П.; Ширальди, Бенедетто; деМенокал, Питер Б. (2013). «Океанографическая изменчивость в регионе зоны конвергенции южной части Тихого океана за последние 210 лет по данным коралловых записей Sr/Ca из нескольких мест» . Геохимия, геофизика, геосистемы . 14 (5): 1435–53. Бибкод : 2013GGG....14.1435W . дои : 10.1029/2012GC004293 .
- ^ Киладис, Джордж Н.; фон Шторх, Ганс; ван Лун, Гарри (1989). «Происхождение зоны конвергенции южной части Тихого океана» . Журнал климата . 2 (10): 1185–95. Бибкод : 1989JCli....2.1185K . doi : 10.1175/1520-0442(1989)002<1185:OOTSPC>2.0.CO;2 .
- ^ Перейти обратно: а б Лукас, Роджер; Линдстрем, Эрик (1991). «Смешанный слой западной экваториальной части Тихого океана». Журнал геофизических исследований . 96 (S1): 3343–58. Бибкод : 1991JGR....96.3343L . дои : 10.1029/90JC01951 .
- ^ «Восхождение Феникса» . Журнал «Нэшнл Географик» . Январь 2011. Архивировано из оригинала 18 декабря 2010 года . Проверено 30 апреля 2011 г.
- ^ Перейти обратно: а б Уолсворт, штат Теннесси; Шиндлер, Делавэр; Колтон, Массачусетс; Вебстер, штат Массачусетс; Палумби, СР; Мамби, Пи Джей; Эссингтон, штат Теннесси; Пинский, М.Л. (1 июля 2019 г.). «Управление разнообразием сетей ускоряет эволюционную адаптацию к изменению климата» . Исследования природы . 9 : 632–636.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ EcoDeco EcoologicalTechnology. Архивировано 7 марта 2011 г. в Wayback Machine . Ecodeco.nl. Проверено 29 ноября 2011 г.
- ^ Проект KoralenKAS. Архивировано 26 апреля 2012 г. в Wayback Machine . Кораалветеншап.нл. Проверено 29 ноября 2011 г.
- ^ Хант, Хлоя В.; Харви, Джеймс Дж.; Миллер, Энн; Джонсон, Вивьен; Фонгсуван, Нифон (2013). «Подход Green Fins к мониторингу и продвижению экологически устойчивых операций по подводному плаванию с аквалангом в Юго-Восточной Азии». Управление океаном и прибрежной зоной . 78 : 35–44. Бибкод : 2013OCM....78...35H . дои : 10.1016/j.ocecoaman.2013.03.004 .
- ^ Магсайсай, Мелисса (21 июня 2009 г.). «Коралл производит фурор» . Лос-Анджелес Таймс . Проверено 12 января 2013 г.
- ^ Уэлч, Патрисия Бьяаланд, Китайское искусство: Путеводитель по мотивам и визуальным образам . Токио, Ратленд и Сингапур: Таттл, 2008, с. 61
- ^ Лейси, Пиппа, «Коралловая сеть: торговля красными кораллами императорскому двору Цин в восемнадцатом веке» в «Глобальной жизни вещей » , изд. Энн Герритсен и Джорджио Айелло, Лондон: Ратледж, 2016, стр. 81
- ^ Фолио 391, Кодекс Юлианы Анисии
- ^ Медь, Эдвин; Хирабаяши, К.; Стричар, КБ; Саммарко, PW (2014). «Кораллы и их потенциальное применение в интегративной медицине» . Доказательная дополнительная и альтернативная медицина . 2014 : 9. дои : 10.1155/2014/184959 . ПМЦ 3976867 . ПМИД 24757491 .
- ^ Сентилкумар, Калимуту; Се-Квон, Ким (2013). «Натуральные продукты из морских беспозвоночных при противовоспалительных и хронических заболеваниях» . Доказательная дополнительная и альтернативная медицина . 2013 : 572859. doi : 10.1155/2013/572859 . ПМЦ 3893779 . ПМИД 24489586 .
- ^ Эрлих, Х.; Этнойер, П.; Литвинов, С.Д.; Оленникова, М.М.; Домашке, Х.; Ханке, Т.; Борн, Р.; Мейснер, Х.; Ворч, Х. (2006). «Структура биоматериала глубоководных бамбуковых кораллов (Anthozoa: Gorgonacea: Isididae): перспективы разработки костных имплантатов и шаблонов для тканевой инженерии». Materialwissenschaft und Werkstofftechnik . 37 (6): 552–57. дои : 10.1002/mawe.200600036 . S2CID 97972721 .
- ^ Редди П.Н., Лакшмана М., Удупа У.В. (декабрь 2003 г.). «Влияние Праваля бхасмы (кораллового камня), природного источника богатого кальция, на минерализацию костей у крыс». Фармакологические исследования . 48 (6): 593–99. дои : 10.1016/S1043-6618(03)00224-X . ПМИД 14527824 .
- ^ Педаниус Диоскорид - Венский Диоскорид, Codex medicus Graecus 1 Австрийской национальной библиотеки Грац: Akademische Druck- und Verlagsanstalt 1998 fol. 391 оборотная сторона (Том 2), комментарий, стр. 47 и 52. ISBN 3-201-01725-6
- ^ Пауэлс, Рэндалл Л. (6 июня 2002 г.). Рог и Полумесяц: культурные изменения и традиционный ислам на восточноафриканском побережье, 800–1900 гг . Издательство Кембриджского университета. п. 26. ISBN 978-0-521-52309-7 .
- ^ «Стратегическое исследование камня: Атлас строительных камней Оксфордшира» . Английское наследие. Март 2011 года . Проверено 23 апреля 2015 г.
- ^ Феррарио, Ф.; Бек, МВт; Сторлацци, CD; Микели, Ф.; Шепард, CC; Айролди, Л. (2014). «Эффективность коралловых рифов для снижения риска прибрежных опасностей и адаптации» . Природные коммуникации . 5 (3794): 3794. Бибкод : 2014NatCo...5.3794F . дои : 10.1038/ncomms4794 . ПМЦ 4354160 . ПМИД 24825660 .
- ^ «Состояние коралловых рифов мира: 2004 г., том 1» (PDF) . Глобальная сеть мониторинга коралловых рифов. Архивировано из оригинала (PDF) 17 июня 2019 г. Проверено 14 января 2019 г.
- ^ Аквариумные кораллы: коллекция и аквариумное содержание нефотосинтетических книдарий северо-восточной части Тихого океана. Архивировано 6 июня 2017 г. в Wayback Machine . Advancedaquarist.com (14 января 2011 г.). Проверено 13 июня 2016 г.
- ^ Reefkeeping 101 - Различные методы контроля питательных веществ . Reefkeeping.com. Проверено 13 июня 2016 г.
- ^ Советы по очистке аквариумного субстрата и живых камней. Архивировано 6 августа 2016 г. в Wayback Machine . Saltaquarium.about.com. Проверено 13 июня 2016 г.
- ^ Коралловые рифы. Архивировано 25 июня 2013 г. на archive.today . Marinebio.org. Проверено 13 июня 2016 г.
- ^ Хорошовский-Фридман Ю.Б., Ижаки И., Ринкевич Б. (2011). «Инженерия по обеспечению личинок коралловых рифов путем трансплантации беременных колоний, выращенных в питомниках». Журнал экспериментальной морской биологии и экологии . 399 (2): 162–66. дои : 10.1016/j.jembe.2011.01.005 .
- ^ Помрой, Роберт С.; Паркс, Джон Э.; Бальбоа, Кристина М. (2006). «Выращивание рифов: является ли аквакультура решением для снижения нагрузки на коралловые рифы в результате рыболовства?». Морская политика . 30 (2): 111–30. дои : 10.1016/j.marpol.2004.09.001 .
- ^ Ринкевич Б (2008). «Управление коралловыми рифами: мы ошиблись, пренебрегая активным восстановлением рифов» (PDF) . Бюллетень о загрязнении морской среды . 56 (11): 1821–24. Бибкод : 2008MarPB..56.1821R . дои : 10.1016/j.marpolbul.2008.08.014 . ПМИД 18829052 . Архивировано из оригинала (PDF) 23 мая 2013 г.
- ^ Ферзе, Себастьян, Калифорния (2010). «Плохая эффективность кораллов, пересаженных на недолговечные субстраты». Реставрационная экология . 18 (4): 399–407. Бибкод : 2010ResEc..18..399F . дои : 10.1111/j.1526-100X.2010.00682.x . S2CID 83723761 .
Источники
[ редактировать ]- Аллен, Греция; Р. Стин (1994). Полевое руководство по коралловым рифам Индо-Тихоокеанского региона . Исследование тропических рифов. ISBN 978-981-00-5687-2 .
- Кальфо, Энтони (2007). Книга о размножении кораллов . Чтение публикаций о деревьях. ISBN 978-0-9802365-0-7 .
- Колин, Польша; К. Арнесон (1995). Тропические тихоокеанские беспозвоночные . Пресса «Коралловый риф». ISBN 978-0-9645625-0-9 .
- Фагерстром, Дж. А. (1987). Эволюция рифовых сообществ . Уайли. ISBN 978-0-471-81528-0 .
- Гослинер, Т.; Д. Беренс; Г. Уильямс (1996). Животные коралловых рифов Индо-Тихоокеанского региона, жизнь животных от Африки до Гавайев (беспозвоночные) . Морские претенденты. ISBN 978-0-930118-21-1 .
- Нибаккен, JW (2004). Морская биология, экологический подход . Пирсон/Бенджамин Каммингс. ISBN 978-0-8053-4582-7 .
- Редхилл, Суррей. Кораллы мира: биология и полевой справочник .
- Сегалов, Нат; Пол Эриксон (1991). Риф оживает. Создание подводной выставки . Ф. Уоттс. ISBN 978-0-531-10994-6 .
- Шеппард, Чарльз Р.К.; Дэви, Саймон К.; Пиллинг, Грэм М. (25 июня 2009 г.). Биология коралловых рифов . ОУП Оксфорд. ISBN 978-0-19-105734-2 .
- Верон, ДЖЕН (1993). Кораллы Австралии и Индо-Тихоокеанского региона . Издательство Гавайского университета. ISBN 978-0-8248-1504-2 .
- Уэллс, Сьюзен (1988). Коралловые рифы мира . МСОП, ЮНЕП. ISBN 9782880329440 .
Внешние ссылки
[ редактировать ]- Коралловые рифы. Океанский портал Смитсоновского института.
- NOAA — Программа сохранения коралловых рифов
- NOAA CoRIS – Биология коралловых рифов
- Офис NOAA по управлению прибрежными зонами – краткие факты – Коралловые рифы
- NOAA Образование в океанических службах – кораллы
- «Что такое коралл?» . Стэнфордский проект микродокументов. Архивировано из оригинала 6 января 2014 г. Проверено 4 февраля 2017 г.
