Рыба
| Рыба Временной диапазон: Средний кембрий - современный | |
|---|---|
 | |
| Бала акула , костистая рыба | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Ольфакторы |
| Подтип: | Позвоночные животные |
| Группы включены | |
| Кладистически включенные, но традиционно исключенные таксоны | |
Рыба или ( мн.р.: рыба плавниками рыбы ) — , амниотическое , жаберное позвоночное водное животное с плавательными и твердым черепом , но лишенное конечностей с пальцами . Рыб можно разделить на более базальных бесчелюстных рыб и более обычных челюстных рыб , причем последние включают всех ныне живущих хрящевых и костных рыб , а также вымерших плакодерм и акантодий . Большинство рыб хладнокровны , температура их тела варьируется в зависимости от окружающей воды, хотя некоторые крупные активные пловцы, такие как белая акула и тунец, могут поддерживать более высокую внутреннюю температуру . Многие рыбы могут акустически общаться друг с другом, например, во время ухаживания .
Самые ранние рыбы появились в кембрийском периоде как маленькие фильтраторы ; они продолжали развиваться на протяжении палеозоя , приобретая множество форм. Самые ранние рыбы со специальными дыхательными жабрами и парными плавниками , остракодермы , имели тяжелые костные пластины , которые служили защитным экзоскелетом от беспозвоночных хищников . Первые рыбы с челюстями , плакодермы, появились в силурийском периоде и значительно разнообразились в девонском периоде , «эпохе рыб».
Костные рыбы, отличающиеся наличием плавательных пузырей , а затем и окостеневшего эндоскелета , стали доминирующей группой рыб после того, как вымирание в конце девона уничтожило верхушечные плакодермы. Костные рыбы подразделяются на лопастеперые и лучепёрые . Около 96% всех ныне живущих видов рыб являются костистыми , коронной группой лучепёрых рыб, которые могут выдвигать вперед челюсти . Четвероногие дышащие , в основном наземная клада позвоночных, которые доминировали на высших трофических уровнях в как в водных , так и наземных экосистемах, начиная с позднего палеозоя , произошли от лопастных рыб в каменноугольном воздухом, периоде, у них развились легкие, гомологичные плавательным пузырям. Несмотря на кладистическое происхождение, четвероногих обычно не считают рыбами, что делает «рыб» парафилетической группой.
Рыба была важным природным ресурсом для человека с доисторических времен, особенно в качестве пищи . Коммерческие и натуральные рыболовы ловят рыбу в диких промыслах или выращивают ее в прудах или в садках для разведения в океане. Рыбу ловят для развлечения или выращивают рыбоводы в качестве украшения для частных и общественных выставок в аквариумах и садовых прудах . Рыбы на протяжении веков играли роль в человеческой культуре , служа божествами, религиозными символами, а также объектами искусства, книг и фильмов.
Этимология
Слово рыба унаследовано от протогерманского языка и связано с немецким Fisch , латинским piscis и древнеирландским īasc , хотя точный корень неизвестен; некоторые авторитеты реконструируют протоиндоевропейский корень * peysk- , встречающийся только в италийском , кельтском и германском языках . [1] [2] [3] [4]
Эволюция
Ископаемая история

Около 530 миллионов лет назад во время кембрийского взрыва появились рыбоподобные животные с хордой и глазами в передней части тела, такие как Haikouichthys в летописи окаменелостей . [5] В позднем кембрии появляются другие бесчелюстные формы, такие как конодонты . [6] [7]
Челюстные позвоночные появляются в силурийском периоде с гигантскими панцирными плакодермами , такими как Dunkleosteus . [8] Челюстные рыбы тоже появились в силурийском периоде: [9] хрящевой хондрихтиес [10] [11] и костлявый Остейхтис . [12]
В течение девона разнообразие рыб значительно увеличилось, в том числе среди плакодерм, лопастеперых рыб и ранних акул, за что девон получил прозвище «эпоха рыб». [13] [14]
Филогения
Рыбы представляют собой парафилетическую группу, поскольку любая клада , содержащая всех рыб, например Gnathostomata или (для костистых рыб) Osteichthyes , также содержит кладу четвероногих (четырехногих позвоночных, в основном наземных), которые обычно не считаются рыбами. [15] [16] Некоторые четвероногие, такие как китообразные и ихтиозавры , вторично приобрели рыбоподобную форму тела в результате конвергентной эволюции . [17] «Рыбы мира» комментируют, что «все более широко признается, что четвероногие, включая нас самих, являются просто модифицированными костными рыбами, и поэтому нам удобно использовать таксон Osteichthyes в качестве клады, которая теперь включает всех четвероногих». [16] Биоразнообразие ; современных рыб распределено по различным группам неравномерно костистые рыбы, способные выдвигать челюсти , составляют 96% видов рыб. [18] [16] Кладограмма [19] показаны эволюционные взаимоотношения всех групп живых рыб (с их соответствующим разнообразием). [16] ) и четвероногих. [20] Вымершие группы отмечены кинжалом ( †); группы неопределенного размещения [19] обозначаются знаком вопроса (?) и пунктирными линиями (- - - - -).
| Позвоночные животные |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




Таксономия
Рыбы (без четвероногих) представляют собой парафилетическую группу, и по этой причине класс Рыб, встречавшийся в старых справочниках, больше не используется в формальных классификациях. Традиционная классификация делит рыб на три существующих класса (Agnatha, Chondrichthyes и Osteichthyes), а вымершие формы иногда классифицируются внутри этих групп, а иногда как отдельные классы. [21]
Рыбы составляют более половины видов позвоночных. По состоянию на 2016 год описано более 32 000 видов костистых рыб, более 1100 видов хрящевых рыб, более 100 миксин и миног. Треть из них приходится на девять крупнейших семей; от самых крупных к самым мелким — это Cyprinidae , Gobiidae , Cichlidae , Characidae , Loricariidae , Balitoridae , Serranidae , Labridae и Scorpaenidae . Около 64 семейств являются монотипными , содержащими только один вид. [16]
Разнообразие
Размеры рыб варьируются от огромной 16-метровой китовой акулы . [22] до некоторых крошечных костистых рыб длиной всего 8 миллиметров (0,3 дюйма), таких как карповые Paedocypris progenetica. [23] и толстая рыбка . [24]
 Самая большая: китовая акула
Самая большая: китовая акула Самый маленький: например, P. progenetica.
Самый маленький: например, P. progenetica.


Способность плавать варьируется от таких рыб, как тунец, лосось и окунь , которые могут преодолевать 10–20 длин тела в секунду, до таких видов, как угри и скаты , которые плавают со скоростью не более 0,5 длины тела в секунду. [25]
 Самый быстрый: например, лосось , 10-20 длин тела в секунду.
Самый быстрый: например, лосось , 10-20 длин тела в секунду. Самый медленный: например , угорь , 0,5 длины тела в секунду.
Самый медленный: например , угорь , 0,5 длины тела в секунду.


Типичная рыба хладнокровна , имеет обтекаемое тело для быстрого плавания, добывает кислород из воды с помощью жабр, имеет два набора парных плавников, один или два спинных плавника, анальный плавник и хвостовой плавник, челюсти, кожу, покрытую чешуей. , и откладывает яйца. Каждый критерий имеет исключения, создающие большое разнообразие форм тела и образа жизни. Например, некоторые быстроплавающие рыбы являются теплокровными, а некоторые медленноплавающие рыбы отказались от обтекаемости в пользу других форм тела. [26]






Экология
Места обитания

Виды рыб примерно поровну разделены между пресноводными и морскими (океаническими) экосистемами; существует около 15 200 пресноводных видов и около 14 800 морских видов. [27] Коралловые рифы в Индо-Тихоокеанском регионе представляют собой центр разнообразия морских рыб. [28] тогда как континентальные пресноводные рыбы наиболее разнообразны в крупных речных бассейнах тропических лесов , особенно в бассейнах Амазонки , Конго и Меконга . [29] пресных водах обитает более 5600 видов рыб Только в неотропических , так что неотропические рыбы составляют около 10% всех видов позвоночных на Земле. [30]
Рыба многочисленна в большинстве водоемов. Их можно найти почти во всех водных средах, от высоких горных ручьев (например, голец и пескарь ) до абиссальных и даже хадальных глубин самых глубоких океанов (например, олени и улитки ), хотя ни один из них не был обнаружен в самых глубоких местах. 25% океана. [31] Самая глубокая живая рыба в океане, обнаруженная на данный момент, — это обыкновенный угорь Abyssobrotula galatheae , обнаруженный на дне впадины Пуэрто-Рико на высоте 8370 м (27 460 футов). [32]
Что касается температуры, ледяная рыба Ионы живет в холоде. [а] воды Южного океана, в том числе под шельфовым ледником Фильшнера-Ронне на 79° ю.ш., [34] в то время как пустынные куколки обитают в пустынных источниках, ручьях и болотах, иногда сильно соленых, с температурой воды до 36 C. [35] [36]
Некоторые рыбы живут в основном на суше или откладывают икру на суше возле воды. [37] Илистые прыгуны питаются и взаимодействуют друг с другом на илистых отмелях, а также уходят под воду, чтобы спрятаться в своих норах. [38] Единственный неописанный вид Phreatobius был назван настоящей «сухопутной рыбой» , поскольку этот червеобразный сом обитает исключительно среди заболоченных листьев . [39] [40] Пещерные рыбы нескольких семейств обитают в подземных озерах , подземных реках или водоносных горизонтах . [41]
Паразиты и хищники
Как и другие животные, рыбы страдают паразитизмом . Некоторые виды используют рыбу-чистильщика для удаления внешних паразитов. Самыми известными из них являются чистильщики коралловых рифов Индийского Тихого и губаны - океанов. Эти мелкие рыбы содержат станции очистки, где собираются другие рыбы, и совершают определенные движения, чтобы привлечь внимание чистильщиков. [42] Чистящее поведение наблюдалось у ряда групп рыб, включая интересный случай между двумя цихлидами одного и того же рода, Etroplus maculatus , чистильщицей, и гораздо более крупной E. suratensis . [43]
Рыбы занимают множество трофических уровней в пресноводных и морских пищевых цепях . Рыбы на более высоких уровнях являются хищными , и значительная часть их добычи состоит из других рыб. [44] Кроме того, млекопитающие, такие как дельфины и тюлени рыбой питаются , а также птицы, такие как олуши и бакланы . [45]


 Барракуда рыбу охотится на более мелкую
Барракуда рыбу охотится на более мелкую Морской лев , хищное млекопитающее, поедающее крупного лосося.
Морской лев , хищное млекопитающее, поедающее крупного лосося. Баклан с добычей рыбы
Баклан с добычей рыбы





Анатомия и физиология
Анатомия и передвижение
Тело типичной рыбы приспособлено к эффективному плаванию за счет попеременного сокращения парных наборов мышц по обе стороны позвоночника. Эти сокращения образуют S-образные кривые, которые движутся вниз по телу. Когда каждая кривая достигает хвостового плавника, к воде прикладывается сила, перемещающая рыбу вперед. Остальные плавники действуют как управляющие поверхности, подобно закрылкам самолета, позволяя рыбе поворачивать в любом направлении. [46]
 Анатомия типичной рыбы ( показан фонарь ):
Анатомия типичной рыбы ( показан фонарь ):
1) жаберная крышка 2) боковая линия 3) спинной плавник 4) жировой плавник
5) хвостовой стебель 6) хвостовой плавник 7) анальный плавник 8) фотофоры 9) брюшные плавники 10) грудные плавники

Поскольку ткани тела плотнее воды, рыба должна компенсировать разницу, иначе она утонет. У многих костных рыб есть внутренний орган, называемый плавательным пузырем , который позволяет им регулировать плавучесть , увеличивая или уменьшая количество содержащегося в нем газа. [47]
Чешуя рыбы обеспечивает защиту от хищников за счет увеличения жесткости и веса. [48] Рыбья чешуя часто обладает высокой отражающей способностью; это серебрение обеспечивает маскировку в открытом океане. Поскольку вода вокруг одинакового цвета, отражение изображения воды обеспечивает почти невидимость. [49]


Тираж
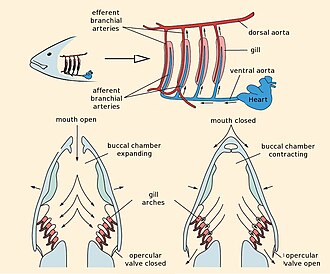
У рыб замкнутая система кровообращения . Сердце ; перекачивает кровь по одному контуру по всему телу для сравнения, сердце млекопитающего имеет две петли: одну для того, чтобы легкие поглощали кислород, а другую для доставки кислорода организму. У рыб сердце перекачивает кровь через жабры. Богатая кислородом кровь затем течет без дальнейшего перекачивания, в отличие от млекопитающих, к тканям тела. Наконец, обедненная кислородом кровь возвращается к сердцу. [50]
Дыхание
Жабры
Для обмена газов рыбы используют жабры по обе стороны глотки . Жабры состоят из гребневидных структур, называемых нитями. Каждая нить содержит капиллярную сеть, обеспечивающую большую площадь поверхности для обмена кислорода и углекислого газа . Рыбы обменивают газы, втягивая через рот богатую кислородом воду и перекачивая ее через жабры. Капиллярная кровь в жабрах течет в направлении, противоположном воде, что приводит к эффективному противоточному обмену . Жабры выталкивают бедную кислородом воду через отверстия по бокам глотки. Хрящевые рыбы имеют множество жаберных отверстий: у акул их обычно пять, иногда шесть или семь пар; им часто приходится плавать, чтобы насытить жабры кислородом. У костных рыб по одному жаберному отверстию с каждой стороны, скрытому под защитным костным покровом или жаберной крышкой . Они способны насыщать кислородом свои жабры с помощью мышц головы. [51]
Дыхание воздухом
Около 400 видов рыб из 50 семейств могут дышать воздухом, что позволяет им жить в бедной кислородом воде или выходить на сушу. [52] Способность рыб делать это потенциально ограничена их одноконтурным кровообращением, поскольку насыщенная кислородом кровь из органа воздушного дыхания смешивается с дезоксигенированной кровью, возвращающейся к сердцу из остального тела. Двоякодышащие рыбы, бичиры, веревки, боуфины, рыбы-змеи и африканские рыбы-ножи эволюционировали так, чтобы уменьшить такое смешивание и уменьшить потерю кислорода из жабр в воду с низким содержанием кислорода. У бичиров и двоякодышащих рыб легкие парные, как у четвероногих, поэтому им приходится всплывать на поверхность, чтобы глотать воздух, и это делает их обязательными для дыхания воздухом. Многие другие рыбы, в том числе обитатели каменных заводей и приливной зоны , являются факультативными дышащими воздухом, способными дышать воздухом, находясь вне воды, что может происходить ежедневно во время отлива , и использовать свои жабры, когда они находятся в воде. Некоторые прибрежные рыбы, такие как скальные прыгуны и илистые прыгуны, предпочитают покидать воду, чтобы питаться в местах обитания, временно открытых для воздуха. [52] Некоторые сомы поглощают воздух через пищеварительный тракт. [53]
Пищеварение
Пищеварительная система состоит из трубки-кишки, ведущей ото рта к заднему проходу. Рот большинства рыб содержит зубы, позволяющие захватывать добычу, откусывать или царапать растительный материал, а также измельчать пищу. Пищевод доставляет пищу в желудок , где она может храниться и частично перевариваться. Сфинктер, привратник, через определенные промежутки времени выпускает пищу в кишечник. У многих рыб вокруг привратника имеются пальцеобразные мешочки, привратниковые придатки , функции которых сомнительны. Поджелудочная железа выделяет в кишечник ферменты для переваривания пищи; другие ферменты секретируются непосредственно самим кишечником. Печень желчь вырабатывает , которая помогает расщеплять жир в эмульсию, которая может всасываться в кишечнике. [54]
Экскреция
Большинство рыб выделяют азотистые отходы в виде аммиака . может выводиться через жабры или фильтроваться почками Он . Соль выводится ректальной железой. [55] Морские рыбы имеют тенденцию терять воду в результате осмоса ; их почки возвращают воду в организм и производят концентрированную мочу. происходит обратное У пресноводных рыб : они имеют тенденцию набирать воду осмотически и производить разбавленную мочу. У некоторых рыб есть почки, способные работать как в пресной, так и в соленой воде. [56]
Мозг

У рыб мозг маленький по сравнению с размером тела по сравнению с другими позвоночными, обычно он составляет одну пятнадцатую массы мозга птицы или млекопитающего такого же размера. [57] Однако у некоторых рыб мозг относительно большой, особенно у мормирид и акул , у которых мозг примерно такого же размера по сравнению с массой тела, как у птиц и сумчатых . [58] В передней части мозга расположены обонятельные доли — пара структур, которые получают и обрабатывают сигналы от ноздрей через два обонятельных нерва . Рыбы, которые охотятся преимущественно по запаху, например миксины и акулы, имеют очень большие обонятельные доли. За ними находится конечный мозг , который у рыб в основном отвечает за обоняние. Вместе эти структуры образуют передний мозг. Передний мозг со средним мозгом соединяет промежуточный мозг ; он работает с гормонами и гомеостазом . находится Шишковидное тело чуть выше промежуточного мозга; он обнаруживает свет, поддерживает циркадные ритмы и контролирует изменения цвета. содержит Средний мозг две оптические доли . Они очень крупные у видов, которые охотятся визуально, таких как радужная форель и цихлиды . контролирует Задний мозг плавание и равновесие. Однодольный мозжечок — самая большая часть мозга; он мал у миксин и миног , но очень велик у мормирид , обрабатывая их электрическое чувство . Ствол головного мозга, или продолговатый мозг, контролирует некоторые мышцы и органы тела, а также управляет дыханием и дыханием. осморегуляция . [57]
Сенсорные системы
Система боковой линии представляет собой сеть датчиков в коже, которая обнаруживает слабые течения и вибрации, а также улавливает движение близлежащей рыбы, будь то хищники или добыча. [59] Это можно считать как осязанием, так и слухом . Слепые пещерные рыбы ориентируются почти полностью за счет ощущений системы боковой линии. [60] У некоторых рыб, таких как сом и акулы, есть ампулы Лоренцини , электрорецепторы , которые улавливают слабые электрические токи порядка милливольт. [61]
Зрение – важная сенсорная система рыб. [62] Глаза рыб похожи на глаза наземных позвоночных, таких как птицы и млекопитающие, но имеют более сферическую линзу . [62] Их сетчатка обычно имеет как палочки , так и колбочки (для скотопического и фотопического зрения ); многие виды обладают цветовым зрением , часто с тремя типами колбочек. [62] Кистистые кости могут видеть поляризованный свет ; [63] у некоторых, таких как карповые, есть четвертый тип колбочек, который улавливает ультрафиолет . [62] Среди бесчелюстных рыб хорошо у миноги развиты глаза. [64] а у миксины только примитивные глазные пятна. [65]
Слух также является важной сенсорной системой рыб. Рыбы чувствуют звук с помощью боковых линий и отолитов в ушах, внутри головы. Некоторые могут улавливать звук через плавательный пузырь. [66]
Некоторые рыбы, в том числе лосось, способны к магниторецепции ; когда ось магнитного поля меняется вокруг круглого резервуара с молодыми рыбами, они переориентируются в соответствии с полем. [67] [68] Механизм магниторецепции рыб остается неизвестным; [69] эксперименты на птицах предполагают механизм квантово-радикальной пары . [70]
Познание
Когнитивные способности рыб включают самосознание , как видно из зеркальных тестов . Манты и губаны , поставленные перед зеркалом, постоянно проверяют, повторяет ли поведение их отражения движения их тела. [71] [72] Губан -черодон , рыба-стрелок и атлантическая треска могут решать проблемы и изобретать инструменты. [73] Моногамная Amatitlania цихлида siquia демонстрирует пессимистическое поведение, когда ей не позволяют быть со своим партнером. [74] Рыбы ориентируются по ориентирам; они могут использовать мысленные карты, основанные на нескольких ориентирах. Рыбы способны научиться преодолевать лабиринты, показывая, что они обладают пространственной памятью и зрительным распознаванием. [75]
Электрогенез

У электрических рыб , таких как рыбы-слоны , африканские рыбы-ножи и электрические угри , некоторые мышцы приспособлены для генерации электрических полей . Они используют поле для обнаружения и идентификации объектов, таких как добыча, в воде вокруг них, которая может быть мутной или темной. [61] Рыбы с сильным электрическим током, такие как электрический угорь, кроме того, могут использовать свои электрические органы для создания ударов, достаточно сильных, чтобы оглушить добычу. [77]
Эндотермия
Большинство рыб исключительно холоднокровные или экзотермные . Однако Scombroidei являются теплокровными (эндотермными), включая марлиновых и тунцов. [78] Опа . , ламприформ , использует эндотермию всего тела, генерируя тепло с помощью плавательных мышц, чтобы согреть свое тело, в то время как противотоковый обмен минимизирует потерю тепла [79] акулы семейств Lamnidae (например, большая белая акула) и Alopiidae Среди хрящевых рыб эндотермичны (молотильные акулы). Степень эндотермии варьируется от марлиновых, которые согревают только глаза и мозг, до голубого тунца и белой акулы , которые поддерживают температуру тела более чем на 20 ° C (68 ° F) выше окружающей воды. [78] [80] [81]
Размножение и жизненный цикл
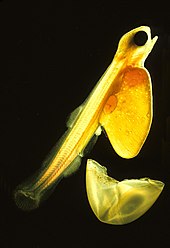
Первичными репродуктивными органами являются парные семенники и яичники . [82] Яйца выходят из яичника в яйцеводы . [83] Более 97% рыб, включая лосося и золотых рыбок, яйцекладущие , то есть икра попадает в воду и развивается вне тела матери. [84] Икра обычно оплодотворяется вне тела матери, при этом самцы и самки сбрасывают свои гаметы в окружающую воду. У некоторых яйцекладущих рыб, таких как скаты , оплодотворение внутреннее: самец использует интромиттирующий орган для доставки спермы в половое отверстие самки. [85] Морские рыбы выпускают большое количество мелкой икры в толщу открытой воды. Только что вылупившаяся молодь яйцекладущих рыб представляет собой планктонные личинки . Они имеют большой желточный мешок и не похожи на молодь или взрослых рыб. Личиночный период у яйцекладущих рыб обычно длится всего несколько недель, личинки быстро растут и меняют структуру , превращаясь в молодь. Во время этого перехода личинки должны переключиться с желточного мешка на питание добычей зоопланктона . [85] Некоторые рыбы, такие как окуни-серфингисты , плавники и лимонные акулы , являются живородящими или живородящими, а это означает, что мать сохраняет икру и питает эмбрионы через структуру, аналогичную плаценте, чтобы соединить кровоснабжение матери с кровоснабжением эмбриона. [85]
восстановление ДНК
Эмбрионы видов рыб, оплодотворенных извне, во время своего развития подвергаются непосредственному воздействию условий окружающей среды, которые могут повредить их ДНК , таких как загрязняющие вещества, ультрафиолетовый свет и активные формы кислорода . [86] множество различных путей восстановления ДНК . Чтобы справиться с такими повреждениями ДНК, эмбрионы рыб во время своего развития используют [86] В последние годы рыбки данио стали полезной моделью для оценки загрязнителей окружающей среды, которые могут быть генотоксичными, то есть вызывать повреждение ДНК. [87]
Защита от болезней
Рыбы обладают как неспецифической, так и иммунной защитой от болезней. Неспецифическая защита включает кожу и чешуйки, а также слой слизи, выделяемый эпидермисом , который улавливает и подавляет рост микроорганизмов . Если патогены нарушают эту защиту, врожденная иммунная система может вызвать воспалительную реакцию , которая увеличивает приток крови к инфицированной области и доставляет лейкоциты , которые пытаются неспецифически уничтожить патогены. Специфическая защита реагирует на определенные антигены, такие как белки на поверхности патогенных бактерий , распознаваемые адаптивной иммунной системой . [88] Иммунная система развилась у вторичноротых , как показано на кладограмме. [89]
Иммунные органы различаются в зависимости от вида рыб. У бесчелюстных рыб имеется лимфоидная ткань в передней части почки и гранулоциты в кишечнике. У них есть свой собственный тип адаптивной иммунной системы ; он использует вариабельные рецепторы лимфоцитов (VLR) для создания иммунитета к широкому спектру антигенов. Результат во многом похож на результат челюстных рыб и четвероногих, но, возможно, он развивался отдельно . [89] Все челюстные рыбы имеют адаптивную иммунную систему с В- и Т- лимфоцитами, несущими иммуноглобулины и Т-клеточные рецепторы соответственно. При этом используется перегруппировка «переменная-разнообразие-соединение» (V(D)J) для создания иммунитета к широкому спектру антигенов. Эта система возникла однажды и является базальной для клада челюстных позвоночных. [89] У хрящевых рыб есть три специализированных органа, содержащих клетки иммунной системы: эпигональные органы вокруг половых желез, орган Лейдига в пищеводе и спиральный клапан в кишечнике, а их тимус и селезенка выполняют функции, аналогичные функциям тех же органов иммунной системы. системы четвероногих. [90] У костистых рыб лимфоциты находятся в тимусе, а другие иммунные клетки — в селезенке и других органах. [91] [92]
Поведение
Мелководье и школьное обучение

Косяк — это слабо организованная группа , в которой каждая рыба плавает и добывает пищу независимо, но ее привлекают другие члены группы, и она корректирует свое поведение, например скорость плавания, так, чтобы оставаться рядом с другими членами группы. Стая — это гораздо более сплоченная группа, синхронизирующая свое плавание так, что все рыбы движутся с одинаковой скоростью и в одном направлении. [94] Обучение иногда является адаптацией против хищников , обеспечивающей повышенную бдительность против хищников. Зачастую более эффективно собирать пищу, работая в группе, и отдельные рыбы оптимизируют свои стратегии, решая присоединиться или покинуть косяк. Когда хищник замечен, рыба-жертва реагирует оборонительно, что приводит к коллективному поведению косяка, например, к синхронным движениям. Ответные меры заключаются не только в попытках спрятаться или убежать; Тактика борьбы с хищниками включает, например, рассеяние и повторную сборку. Рыба также собирается в косяки для нереста. [93] Мойва ежегодно мигрирует большими стаями между местами нагула и нерестилищами. [95]
Коммуникация
Рыбы общаются, передавая друг другу акустические сигналы (звуки). Чаще всего это происходит в контексте кормления, агрессии или ухаживания. [96] Издаваемые звуки различаются в зависимости от вида и стимула. Рыбы могут издавать либо стридуляционные звуки, перемещая компоненты скелетной системы, либо могут издавать нестридуляционные звуки, манипулируя специальными органами, такими как плавательный пузырь. [97]

Некоторые рыбы издают звуки, потирая или перетирая кости. Эти звуки являются стридуляционными. Haemulon flavolineatum , французская хрюкающая рыба, поскольку она издает хрюкающий звук, скрежетая зубами, особенно когда терпит бедствие. Ворчание происходит с частотой около 700 Гц и длится примерно 47 миллисекунд. [97] Длиннорылый морской конёк, Hippocampus reidi, издаёт две категории звуков: «щелчки» и «рычание», потирая венечную кость о рифленую часть нейрокраниума. [98] Щелчки издаются во время ухаживания и кормления, частота щелчков находится в диапазоне 50–800 Гц. Частоты находятся на верхнем конце диапазона во время нереста, когда самка и самец находились на расстоянии менее пятнадцати сантиметров друг от друга. Рычание издается, когда H. reidi находится в состоянии стресса. Звуки «рычания» состоят из серии звуковых импульсов и издаются одновременно с вибрациями тела. [99]
Некоторые виды рыб создают шум, задействуя специальные мышцы, которые сокращаются и вызывают вибрации плавательного пузыря. Рыба-устрица издает громкое ворчание, сокращая звуковые мышцы по бокам плавательного пузыря. [100] Самки и самцы рыб-жаб издают короткое ворчание, часто в ответ на испуг. [101] Помимо кратковременного хрюканья, самцы рыб-жаб издают «лодочные свистки». [102] Эти звонки более продолжительны, менее часты и в основном используются для привлечения партнеров. [102] Различные звуки имеют частотный диапазон от 140 Гц до 260 Гц. [102] Частота звуков зависит от скорости сокращения звуковых мышц. [103] [100]
Красный барабан Sciaenops ocellatus издает барабанные звуки за счет вибрации плавательного пузыря. Вибрации вызываются быстрым сокращением звуковых мышц, окружающих дорсальную часть плавательного пузыря. Эти вибрации приводят к повторяющимся звукам с частотами от 100 до >200 Гц. S. ocellatus издает разные звуки в зависимости от задействованных стимулов, таких как ухаживание или нападение хищника. Самки не издают звуков и у них отсутствуют звукопроизводящие (звуковые) мышцы. [104]
Сохранение
2024 года МСОП В Красный список включено 2168 видов рыб, находящихся под угрозой исчезновения или находящихся под угрозой исчезновения. [105] Включены такие виды, как атлантическая треска , [106] Куколка Дьявольской Дыры , [107] целаканты , [108] и большие белые акулы . [109] Поскольку рыбы живут под водой, их труднее изучать, чем наземных животных и растения, а информация о популяциях рыб часто отсутствует. Однако пресноводные рыбы кажутся особенно уязвимыми, поскольку они часто обитают в относительно небольших водоемах. Например, детеныш Дьявольской дыры занимает только один бассейн размером 3 на 6 метров (10 на 20 футов). [110]
Перелов
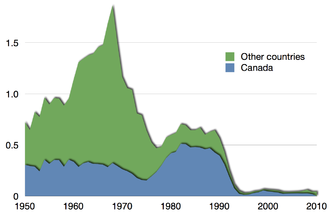
Продовольственная и сельскохозяйственная организация сообщает, что «в 2017 году 34 процента рыбных запасов мирового морского рыболовства были классифицированы как перелавливаемые». [112] Чрезмерный вылов рыбы представляет собой серьезную угрозу для съедобных рыб, таких как треска и тунец . [113] [114] Чрезмерный вылов рыбы в конечном итоге приводит к исчезновению рыбных запасов , поскольку выжившие не могут произвести достаточно молоди, чтобы заменить выловленную. Такое коммерческое исчезновение не означает, что вид вымер, просто он больше не может поддерживать промысел. В случае с тихоокеанским промыслом сардин у побережья Калифорнии вылов неуклонно снижался с пика 1937 года в 800 000 тонн до экономически нерентабельных 24 000 тонн в 1968 году. [115] В случае промысла трески на северо-западе Атлантического океана из-за чрезмерного вылова популяция рыбы к 1992 году сократилась до 1% от исторического уровня. [111] Ученые-рыболовы и представители рыбной промышленности имеют совершенно разные взгляды на устойчивость рыболовства к интенсивному вылову рыбы. Во многих прибрежных регионах рыбная промышленность является крупным работодателем, поэтому правительства готовы ее поддерживать. [116] [117] С другой стороны, ученые и защитники природы настаивают на строгой защите, предупреждая, что многие запасы могут быть уничтожены в течение пятидесяти лет. [118] [119]
Другие угрозы
Ключевым стрессом как для пресноводных, так и для морских экосистем является деградация среды обитания , включая загрязнение воды , строительство плотин, изъятие воды для использования человеком и интродукция экзотических видов, включая хищников. [120] Пресноводная рыба, особенно если она является эндемичной для определенного региона (и больше нигде не встречается), может оказаться под угрозой исчезновения по всем этим причинам, как в случае с тремя из десяти эндемичных пресноводных рыб Испании. [121] Речные плотины, особенно такие крупные сооружения, как плотина Кариба (река Замбези) и Асуанская плотина ( река Нил ) на реках с экономически важным рыболовством, привели к значительному сокращению улова рыбы. [122] Промышленное донное траление может нанести вред обитателям морского дна , как это произошло на банке Жорж в Северной Атлантике. [123] Интродукция водных инвазивных видов широко распространена. Это изменяет экосистемы, вызывая потерю биоразнообразия и может нанести вред рыболовству. Вредные виды включают рыбу, но не ограничиваются ею; [124] приход гребневика в Черное море нанес ущерб тамошнему промыслу хамсы . [125] [124] Открытие Суэцкого канала в 1869 году вызвало миграцию лессепсисов , в ходе которой сотни видов рыб, водорослей и беспозвоночных проникли в Средиземное море , изменив его экологию. [126] Хищный нильский окунь был намеренно завезен в озеро Виктория в 1960-х годах как промысловая и спортивная рыба. Озеро отличалось высоким биоразнообразием: около 500 эндемичных видов цихлид . Это радикально изменило экологию озера и упростило промысел с нескольких видов до трех: нильского окуня, серебристого карпового и еще одной завезенной рыбы, нильской тилапии . Популяции гаплохроминовых цихлид сократились. [127] [128]
Важность для человека
Экономический

На протяжении всей истории люди использовали рыбу в качестве источника пищевого белка . Как исторически, так и сегодня большая часть рыбы, добываемой для потребления человеком, добывается путем ловли дикой рыбы. Однако рыбоводство, которое практикуется примерно с 3500 г. до н. э. в древнем Китае, [129] становится все более важным во многих странах. В целом, по оценкам, около одной шестой мирового белка обеспечивается рыбой. [130] Соответственно, рыболовство является крупным глобальным бизнесом, который обеспечивает доход миллионам людей. [130] У Фонда защиты окружающей среды есть руководство, согласно которому рыба безопасна для употребления в пищу, учитывая уровень загрязнения в современном мире, и какая рыба добывается экологически чистым способом. [131] По состоянию на 2020 год было выловлено более 65 миллионов тонн морской рыбы и 10 миллионов тонн пресноводной рыбы, а также выращено около 50 миллионов тонн рыбы, в основном пресноводной. Из морских видов, выловленных в 2020 году, анчоус составлял 4,9 млн тонн, минтай — 3,5 млн тонн, тунец — 2,8 млн тонн, а атлантическая сельдь и желтоперый тунец — по 1,6 млн тонн каждый; еще восемь видов имели уловы более 1 млн тонн. [132]
Отдых
Рыба была признана источником красоты почти с тех пор, как она использовалась в пищу, появлялась в наскальных рисунках , выращивалась как декоративная рыба в прудах и выставлялась в аквариумах в домах, офисах или общественных местах. Любительская рыбалка – это рыбалка в первую очередь для удовольствия или соревнований; его можно противопоставить коммерческому рыболовству, то есть лову рыбы с целью получения прибыли, или кустарному рыболовству , которое занимается ловлей рыбы в первую очередь ради еды. В наиболее распространенной форме любительской рыбалки используются удочка , катушка , леска , крючки и широкий выбор наживок . Любительское рыболовство особенно популярно в Северной Америке и Европе; правительственные учреждения часто активно управляют целевыми видами рыб. [133] [134]
Культура
Рыбная тематика имеет символическое значение во многих религиях. В древней Месопотамии рыбные подношения богам приносились с самых давних времен. [135] Рыба также была главным символом Энки , бога воды. [135] Рыбы часто появляются в качестве мотивов заполнения цилиндрических печатей старовавилонского 1830 г. до н . ( ок. э. – ок. 1531 г. до н. э.) и неоассирийского (911–609 до н. э.) периодов. [135] Начиная с касситского периода ( ок. 1600 г. до н.э. – ок. 1155 г. до н.э.) и продолжаясь до раннего персидского периода (550–30 гг. до н.э.), целители и экзорцисты одевались в ритуальные одежды, напоминающие тела рыб. [135] Говорят, что в период Селевкидов (312–63 гг. До н. э.) легендарный герой вавилонской культуры Оаннес носил рыбью шкуру. [135] Рыба была священной сирийской богиней Атаргатис. [136] и во время ее праздников только ее священникам разрешалось есть их. [136] В « Книге Ионы » центральная фигура, пророк по имени Иона , проглатывается гигантской рыбой после того, как команда корабля, на котором он путешествует, выбросила его за борт. [137] Ранние христиане использовали ихтис , символ рыбы, для обозначения Иисуса. [136] [138] Среди божеств, которые, как говорят, принимают форму рыбы, Икатере полинезийцев есть . [139] бог-акула Камохоали с Гавайев , [140] и Матсья индусов. [141] Созвездие Рыб («Рыбы») связано с легендой Древнего Рима о том, что Венеру и ее сына Купидона спасли две рыбы. [142]
Рыбы занимают видное место в искусстве. [143] в таких фильмах, как « В поисках Немо» [144] и такие книги, как «Старик и море» . [145] Крупные рыбы, особенно акулы, часто становились объектами фильмов ужасов и триллеров , в частности, романа «Челюсти» , по которому был снят фильм, который, в свою очередь, неоднократно пародировался и имитировался. [146] Пираньи показаны в таком же свете, как и акулы в таких фильмах, как «Пираньи» . [147]

 Рыбная лавка , Бартоломео Пассеротти , 1580-е годы.
Рыбная лавка , Бартоломео Пассеротти , 1580-е годы. Золотая рыбка , Анри Матисс , 1912 год.
Золотая рыбка , Анри Матисс , 1912 год.



См. также
- Глубоководная рыба
- Синдром острой токсичности рыбы
- Развитие рыбы
- Кормовая рыба
- Ихтиология
- Список распространенных названий рыб
- Список семейств рыб
- Ртуть в рыбе
- Отолит (кость, используемая для определения возраста рыбы)
- Беременность (рыбы)
- Ходячая рыба
Примечания
- ^ Температура часто составляет около 0 C. Температура замерзания морской воды на поверхности составляет -1,85 C, а на глубине 1000 метров падает до -2,62 C. Однако воду можно переохладить несколько ниже этих температур. [33]
Ссылки
- ^ «DWDS – Цифровой словарь немецкого языка» . DWDS (на немецком языке). Архивировано из оригинала 31 июля 2020 года . Проверено 21 января 2023 г.
- ^ Уинфред Филипп Леманн, Хелен-Джо Дж. Хьюитт, Зигмунд Файст, Готский этимологический словарь , 1986, sv fisks p. 118
- ^ «рыба, №1» . Издательство Оксфордского университета . Архивировано из оригинала 17 марта 2023 года . Проверено 21 января 2023 г.
- ^ Бак, Карл Дарлинг (1949). «раздел 3.65». Словарь избранных синонимов основных индоевропейских языков . п. 184.
- ^ Шу, Д.Г.; Конвей Моррис, С.; Хан, Дж.; Чжан, ZF; Ясуи, К.; Жанвье, П.; и др. (2003). «Голова и позвоночник раннекембрийского позвоночного Haikouichthys » . Природа . 421 (6922): 526–529. Бибкод : 2003Natur.421..526S . дои : 10.1038/nature01264 . ПМИД 12556891 . S2CID 4401274 .
- ^ Донохью, Филип CJ; Пурнелл, Марк А. (2009). «Эволюционное возникновение позвоночных животных из числа их бесхребетных сородичей» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 204–212. дои : 10.1007/s12052-009-0134-3 . ISSN 1936-6426 .
- ^ Миллер, Джеймс Ф.; Кларк, Д.Л. (1984). «Эволюция конодонтов кембрийского и раннего ордовика, биофации и провинциализм» . Специальный доклад Геологического общества Америки . Специальные статьи Геологического общества Америки. 196 (196): 43–68. дои : 10.1130/SPE196-p43 . ISBN 978-0-8137-2196-5 .
- ^ «Рыба-монстр сокрушила противника сильнейшим укусом» . Smh.com.au. 30 ноября 2006 г. Архивировано из оригинала 2 апреля 2013 г. Проверено 26 февраля 2013 г.
- ^ Чу, Брайан; Чжу, Мин; Чжао, Вэньцзинь; Цзя, Ляотао; Чжу, Юань (2014). «Крупнейшее силурийское позвоночное животное и его палеоэкологические последствия» . Научные отчеты . 4 : 5242. Бибкод : 2014NatSR...4E5242C . дои : 10.1038/srep05242 . ПМК 4054400 . ПМИД 24921626 .
- ^ Андреев, Пламен С.; Сансом, Иван Дж.; Ли, Цян; Чжао, Вэньцзинь; Ван, Цзяньхуа; Ван, Чун-Чье; и др. (сентябрь 2022 г.). «Колючий хондрихт из нижнего силура Южного Китая» . Природа . 609 (7929): 969–974. Бибкод : 2022Natur.609..969A . дои : 10.1038/s41586-022-05233-8 . ПМИД 36171377 . S2CID 252570103 .
- ^ Андреев, Пламен С.; Сансом, Иван Дж.; Ли, Цян; Чжао, Вэньцзинь; Ван, Цзяньхуа; Ван, Чун-Чье; и др. (сентябрь 2022 г.). «Самые древние зубы гнатостома» . Природа . 609 (7929): 964–968. Бибкод : 2022Natur.609..964A . дои : 10.1038/s41586-022-05166-2 . ISSN 0028-0836 . ПМИД 36171375 . S2CID 252569771 .
- ^ Берг, Линда Р.; Соломон, Эльдра Перл; Мартин, Диана В. (2004). Биология . Cengage Обучение . п. 599. ИСБН 978-0-534-49276-2 .
- ^ Бентон 2005 , с. 35: Рис. 2.10, с. 73: Рис. 3.25.
- ^ Далтон, Рекс (январь 2006 г.). «Подсел на окаменелости» . Природа . 439 (7074): 262–263. дои : 10.1038/439262а . ПМИД 16421540 . S2CID 4357313 .
- ^ Грин, Гарри В. (1 января 1998 г.). «Мы приматы и мы рыбы: преподавание монофилетической биологии организма». Интегративная биология . 1 (3): 108–111. doi : 10.1002/(sici)1520-6602(1998)1:3<108::aid-inbi5>3.0.co;2-t . ISSN 1520-6602 .
- ^ Jump up to: Перейти обратно: а б с д и Нельсон 2016 , с. 3
- ^ Дэвис, RW (2019). «Возвращение в море: эволюция морских млекопитающих». В Дэвисе, RW (ред.). Морские млекопитающие: приспособления к водной жизни . Нью-Йорк: Международное издательство Springer . стр. 7–27. ISBN 978-3-3199-8278-6 .
- ^ Бентон 2005 , стр. 175–184.
- ^ Jump up to: Перейти обратно: а б Фридман, Мэтт; Саллан, Лорен Коул (июнь 2012 г.). «Пятьсот миллионов лет вымирания и восстановления: фанерозойское исследование крупномасштабного разнообразия рыб» . Палеонтология . 55 (4): 707–742. Бибкод : 2012Palgy..55..707F . дои : 10.1111/j.1475-4983.2012.01165.x . S2CID 59423401 .
- ^ «Сводная статистика» . Красный список исчезающих видов МСОП . 2023.1 . Проверено 5 февраля 2024 г. Таблица 1а: Количество видов, оцененное по отношению к общему числу описанных видов, и количество видов, находящихся под угрозой исчезновения, по основным группам организмов
- ^ Бентон, MJ (1998). «Качество летописи окаменелостей позвоночных». В Доноване, СК; Пол, CRC (ред.). Адекватность летописи окаменелостей . Уайли . С. 269–303, рис. 2.
- ^ Макклейн, Крейг Р.; Балк, Меган А.; Бенфилд, Марк К.; Бранч, Тревор А.; Чен, Кэтрин; Косгроув, Джеймс; и др. (13 января 2015 г.). «Определение размеров океанских гигантов: закономерности внутривидовых изменений размеров морской мегафауны» . ПерДж . 3 : е715. дои : 10.7717/peerj.715 . ISSN 2167-8359 . ПМЦ 4304853 . ПМИД 25649000 .
- ^ Коттелат, Морис; Бритц, Ральф; Хок Хуэй, Тан; Витте, Кай-Эрик (2005). « Paedocypris , новый род карповых рыб Юго-Восточной Азии с замечательным половым диморфизмом, включает в себя самых маленьких позвоночных в мире» (PDF) . Труды Королевского общества Б. 273 (1589): 895–899. дои : 10.1098/rspb.2005.3419 . ПМЦ 1560243 . ПМИД 16627273 . Архивировано из оригинала (PDF) 12 июля 2009 года . Проверено 26 октября 2012 г.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2017). « Schindleria brevipinguis » в FishBase . Версия за сентябрь 2017 года.
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 103.
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 3, 33–36.
- ^ Манель, Стефани; Герен, Пьер-Эдуард; Муйо, Давид; Бланше, Саймон; Велес, Лора; Альбуи, Камилла; Пеллиссье, Лоик (10 февраля 2020 г.). «Глобальные детерминанты генетического разнообразия пресноводных и морских рыб» . Природные коммуникации . 11 (1): 692. Бибкод : 2020NatCo..11..692M . дои : 10.1038/s41467-020-14409-7 . ISSN 2041-1723 . ПМК 7010757 . ПМИД 32041961 .
- ^ Юбер, Николя; Мейер, Кристофер П.; Брюггеманн, Генрих Дж.; Герен, Фабьен; Комено, Роберто Дж.Л.; Эспио, Бенуа; Косс, Ромен; Уильямс, Джеффри Т.; Самолеты, Серж (15 марта 2012 г.). «Загадочное разнообразие коралловых рифовых рыб Индо-Тихоокеанского региона, выявленное с помощью штрих-кодирования ДНК, обеспечивает новую поддержку гипотезы о центре перекрытия» . ПЛОС ОДИН . 7 (3): e28987. Бибкод : 2012PLoSO...728987H . дои : 10.1371/journal.pone.0028987 . ISSN 1932-6203 . ПМЦ 3305298 . ПМИД 22438862 .
- ^ ван дер Слин, Питер; Альберт, Джеймс С. (2022). «Закономерности разнообразия пресноводных рыб». Энциклопедия внутренних вод . Эльзевир. стр. 243–255. дои : 10.1016/b978-0-12-819166-8.00056-6 . ISBN 978-0-12-822041-2 .
- ^ Альберт, Джеймс С.; Карвалью, Тьяго П.; Петри, Пауло (июнь 2011 г.). «Водное биоразнообразие в бассейне Амазонки: специализация среды обитания и географическая изоляция способствуют богатству видов» . Животные . 1 (2): 205–241. дои : 10.3390/ani1020205 . ПМЦ 4513461 . ПМИД 26486313 .
- ^ Янси, PH; Герринджер, Мэн; Дражен, Дж. К.; Роуден, А.А.; Джеймисон, А. (2014). «Морские рыбы могут быть биохимически ограничены в заселении самых глубоких океанских глубин» . Proc Natl Acad Sci США . 111 (12): 4461–4465. Бибкод : 2014PNAS..111.4461Y . дои : 10.1073/pnas.1322003111 . ПМЦ 3970477 . ПМИД 24591588 .
- ^ «Какая самая глубоководная рыба?» . Австралийский музей . 23 декабря 2014 года . Проверено 18 сентября 2015 г.
- ^ Хауманн, Ф. Александр; Мурман, Рут; Райзер, Стивен С.; Смедсруд, Ларс Х.; Максим, Тед; Вонг, Энни PS; Уилсон, Эрл А.; Друкер, Роберт; Тэлли, Линн Д.; Джонсон, Кеннет С.; Ки, Роберт М.; Сармьенто, Хорхе Л. (28 октября 2020 г.). «Переохлажденные воды Южного океана». Письма о геофизических исследованиях . 47 (20). Бибкод : 2020GeoRL..4790242H . дои : 10.1029/2020GL090242 . hdl : 1912/26495 .
- ^ Персер, Отен; Хехеманн, Лаура; Берингер, Лилиан; Типпенхауэр, Сандра; Веге, Миа; Борнеманн, Хорст; и др. (2022). «В Антарктике обнаружена огромная колония ледяной рыбы» . Современная биология . 32 (4): 842–850.e4. Бибкод : 2022CBio...32E.842P . дои : 10.1016/j.cub.2021.12.022 . hdl : 2263/90796 . ПМИД 35030328 . S2CID 245936769 .
- ^ Марш, Пол С.; Сада, Дональд В. (1993). «План восстановления пустынной рыбки (Cyprinodon macularius)» (PDF) . Служба охраны рыбы и дикой природы США . Архивировано (PDF) из оригинала 17 октября 2011 года.
- ^ Шроде, Джой Б.; Геркинг, Шелби Д. (1977). «Влияние постоянных и переменных температур на репродуктивную функцию пустынной куколки Cyprinodon n. nevadensis» . Физиологическая зоология . 50 (1): 1–10. дои : 10.1086/physzool.50.1.30155710 . ISSN 0031-935X . S2CID 82166135 .
- ^ Мартин, KLM (2014). Рыбы, нерестящиеся на пляже: воспроизводство в исчезающей экосистеме . ЦРК Пресс . ISBN 978-1-4822-0797-2 .
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2006). « Periophthalmus barbarus » в FishBase . Версия ноября 2006 г.
- ^ «Cat-eLog: Heptapteridae: Phreatobius : Phreatobius sp. (1)» . Планета Сом . Архивировано из оригинала 23 октября 2006 года . Проверено 26 ноября 2006 г.
- ^ Хендерсон, Пенсильвания; Уокер, И. (1990). «Пространственная организация и плотность населения рыбного сообщества подстилки в центральном черноморском потоке Амазонки». Журнал биологии рыб . 37 (3): 401–411. Бибкод : 1990JFBio..37..401H . дои : 10.1111/j.1095-8649.1990.tb05871.x .
- ^ Хельфман, Г.С. (2007). Сохранение рыбы: Руководство по пониманию и восстановлению глобального водного биоразнообразия и рыбных ресурсов . Айленд Пресс . стр. 41–42. ISBN 978-1-55963-595-0 .
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 380.
- ^ Вайман, Ричард Л.; Уорд, Джек А. (1972). «Очистительный симбиоз между цихлидами Etroplus maculatus и Etroplus suratensis. I. Описание и возможная эволюция». Копейя . 1972 (4): 834–838. дои : 10.2307/1442742 . JSTOR 1442742 .
- ^ Майерс, Рэнсом А.; Червь, Борис (2003). «Быстрое истощение хищных рыбных сообществ во всем мире». Природа . 423 (6937). Springer Science and Business Media : 280–283. Бибкод : 2003Natur.423..280M . дои : 10.1038/nature01610 . ISSN 0028-0836 . ПМИД 12748640 .
- ^ «Хищничество» . Северо-Западный совет по энергетике и охране окружающей среды . Проверено 10 февраля 2024 г.
- ^ Сфакиотакис, М.; Лейн, DM; Дэвис, JBC (1999). «Обзор способов плавания рыб для передвижения в воде» (PDF) . Журнал IEEE океанической инженерии . 24 (2): 237–252. Бибкод : 1999IJOE...24..237S . дои : 10.1109/48.757275 . S2CID 17226211 . Архивировано из оригинала (PDF) 24 декабря 2013 года.
- ^ «Актиноптеригии: Еще о морфологии» . Музей палеонтологии Калифорнийского университета . Проверено 10 февраля 2024 г.
- ^ Цюань, Хаочэн; Ян, Вэнь; Лаперьер, морской пехотинец; Шайбле, Эрик; Ричи, Роберт О.; Мейерс, Марк А. (2020). «Структура и механическая адаптируемость современной эласмоидной чешуи обыкновенного карпа». Иметь значение . 3 (3): 842–863. дои : 10.1016/j.matt.2020.05.011 .
- ^ Сельдь, Питер (2002). Биология глубокого океана . Издательство Оксфордского университета . стр. 192–195. ISBN 978-0-19-854956-7 .
- ^ «Системы кровообращения животных» . Технологический институт Джорджии . Проверено 10 февраля 2024 г.
- ^ Ромер, Альфред Шервуд ; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия: Холт-Сондерс Интернэшнл. стр. 316–327. ISBN 0-03-910284-Х .
- ^ Jump up to: Перейти обратно: а б Грэм, Джеффри Б.; Вегнер, Северная Каролина (2010). «6. Дыхание воздухом в воде и на воздухе: рыбы, дышащие воздухом». Нильссон, Йоран Э. (ред.). Респираторная физиология позвоночных . Нью-Йорк: Издательство Кембриджского университета . стр. 174–221. дои : 10.1017/CBO9780511845178.007 . ISBN 978-0-521-87854-8 .
- ^ Армбрустер, Джонатан В. (1998). «Модификации пищеварительного тракта для удержания воздуха у лорикариевых и сколоплацидных сомов» (PDF) . Копейя . 1998 (3): 663–675. дои : 10.2307/1447796 . JSTOR 1447796 . Архивировано из оригинала (PDF) 26 марта 2009 года . Проверено 25 июня 2009 г.
- ^ «Пищеварительная система» . Университет Теннесси . Проверено 10 февраля 2024 г.
- ^ Бертон, Дерек; Бертон, Маргарет (21 декабря 2017 г.). «Экскреция». Оксфордская стипендия онлайн . Издательство Оксфордского университета . дои : 10.1093/oso/9780198785552.003.0008 .
- ^ Маец, Дж. (20 августа 1971 г.). «Рыбьи жабры: механизмы переноса солей в пресной и морской воде». Философские труды Лондонского королевского общества Б. 262 (842): 209–249. Бибкод : 1971РСПТБ.262..209М . дои : 10.1098/rstb.1971.0090 . ISSN 0080-4622 .
- ^ Jump up to: Перейти обратно: а б Хелфман, Коллетт и Фейси, 1997 , стр. 48–49.
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 191.
- ^ Блекманн, Хорст; Зелик, Рэнди (1 марта 2009 г.). «Система боковой линии рыб» . Интегративная зоология . 4 (1): 13–25. дои : 10.1111/j.1749-4877.2008.00131.x . ПМИД 21392273 .
- ^ Годфри-Смит, Питер (2020). «Королевская рыба». Метазоа . Нью-Йорк: Фаррар, Штраус и Жиру . ISBN 9780374207946 .
- ^ Jump up to: Перейти обратно: а б Альберт, Дж.С.; Крэмптон, WG (2006). «Электрорецепция и электрогенез». В Лутце, PL (ред.). Физиология рыб . Бока-Ратон, Флорида: CRC Press. стр. 429–470. ISBN 978-0-8493-2022-4 .
- ^ Jump up to: Перейти обратно: а б с д Гатри, DM (1986). «Роль зрения в поведении рыб». Поведение костистых рыб . Бостон, Массачусетс: Спрингер. стр. 75–113. дои : 10.1007/978-1-4684-8261-4_4 . ISBN 978-1-4684-8263-8 .
- ^ Гавришин, Крейг В. (2010). «Ультрафиолетовое поляризационное зрение и визуально управляемое поведение рыб». Мозг, поведение и эволюция . 75 (3): 186–194. дои : 10.1159/000314275 . ISSN 0006-8977 . ПМИД 20733294 .
- ^ Мейер-Рохов, В. Бенно; Стюарт, Дункан (1996). «Обзор личиночной и постличиночной ультраструктуры глаз миноги (круглоротых) с особым акцентом на Geotria australis (серый)». Микроскопические исследования и техника . 35 (6): 431–444. doi : 10.1002/(SICI)1097-0029(19961215)35:6<431::AID-JEMT3>3.0.CO;2-L . ПМИД 9016447 . S2CID 22940203 .
- ^ Лэмб, Тревор Д.; Коллин, Шон П.; Пью, Эдвард Н. (2007). «Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и глазная чашечка» . Обзоры природы Неврология . 8 (12): 960–976. дои : 10.1038/nrn2283 . ISSN 1471-003X . ПМК 3143066 . ПМИД 18026166 . См. также статью Ламба и др. «Происхождение глаза позвоночных», 2008 г.
- ^ Хокинс, AD (1981). «6. Слух рыб». В Таволге, Уильям Н.; Поппер, Артур Н.; Фэй, Ричард Р. (ред.). Слух и звуковая коммуникация рыб . Спрингер . стр. 109–138. ISBN 978-1-4615-7188-9 .
- ^ Куинн, Томас П. (1980). «Доказательства ориентации по небесному и магнитному компасу у молоди мигрирующей по озеру нерки». Журнал сравнительной физиологии А. 137 (3): 243–248. дои : 10.1007/bf00657119 . S2CID 44036559 .
- ^ Тейлор, П.Б. (май 1986 г.). «Экспериментальные доказательства геомагнитной ориентации молоди лосося Oncorhynchus tschawytscha Walbaum». Журнал биологии рыб . 28 (5): 607–623. Бибкод : 1986JFBio..28..607T . дои : 10.1111/j.1095-8649.1986.tb05196.x .
- ^ Формицкий, Кшиштоф; Корзелецка-Оркиш, Агата; Таньский, Адам (2019). «Магниторецепция у рыб». Журнал биологии рыб . 95 (1): 73–91. Бибкод : 2019JFBio..95...73F . дои : 10.1111/jfb.13998 . ISSN 0022-1112 . ПМИД 31054161 .
- ^ Хор, Питер Дж .; Муритсен, Хенрик (апрель 2022 г.). «Квантовая природа миграции птиц» . Научный Американ : 24–29.
- ^ Ари, Чилла; Д'Агостино, Доминик П. (1 мая 2016 г.). «Проверка на случай непредвиденных обстоятельств и самостоятельное поведение гигантских скатов манты: есть ли у пластиножаберных самосознание?» . Журнал этологии . 34 (2): 167–174. дои : 10.1007/s10164-016-0462-z . ISSN 1439-5444 . S2CID 254134775 . Архивировано из оригинала 17 марта 2023 года . Проверено 21 января 2023 г.
- ^ Кода, Масанори; Хотта, Такаши; Такеяма, Томохиро; Авата, Сатоши; Танака, Хирокадзу; Асаи, Джун-я; Джордан, Л. Алекс (21 августа 2018 г.). «Губаны-чистильщики проходят тест на отметку. Каковы последствия для тестирования сознания и самосознания у животных?» . ПЛОС Биология . 17 (2): 397067. bioRxiv 10.1101/397067 . дои : 10.1371/journal.pbio.3000021 . ПМК 6366756 . ПМИД 30730878 . S2CID 91375693 .
- ^ Балкомб, Джонатан (1 мая 2017 г.). «Рыбы используют инструменты для решения проблем и изобретают» . Научный американец . Архивировано из оригинала 17 марта 2023 года.
- ^ Лаубу, Хлоя; Луапре, Филипп; Дешом-Моншармон, Франсуа-Ксавье (2019). «Связывание пар влияет на эмоциональное состояние моногамных видов рыб» . Учеб. Р. Сок. Б. 286 (1904). 20190760. дои : 10.1098/rspb.2019.0760 . ПМК 6571461 . ПМИД 31185864 .
- ^ наук, Журнал студенческой жизни. «Подходящая методология лабиринта для изучения обучения рыб» (PDF) . Архивировано из оригинала (PDF) 6 июля 2011 года . Проверено 28 мая 2009 г.
- ^ фон дер Эмде, Г. (15 мая 1999 г.). «Активная электролокация предметов у слабоэлектрических рыб». Журнал экспериментальной биологии . 202 (10): 1205–1215. дои : 10.1242/jeb.202.10.1205 . ПМИД 10210662 .
- ^ Катания, Кеннет К. (20 октября 2015 г.). «Электрические угри используют высокое напряжение, чтобы отслеживать быстро движущуюся добычу» . Природные коммуникации . 6 : 8638. Бибкод : 2015NatCo...6.8638C . дои : 10.1038/ncomms9638 . ПМЦ 4667699 . ПМИД 26485580 .
- ^ Jump up to: Перейти обратно: а б Блок, бакалавр; Финнерти, младший (1993). «Эндотермия у рыб: филогенетический анализ ограничений, предрасположенностей и давления отбора» (PDF) . Экологическая биология рыб . 40 (3): 283–302. дои : 10.1007/BF00002518 . S2CID 28644501 . Архивировано из оригинала 6 ноября 2020 года . Проверено 1 октября 2018 г.
- ^ Вегнер, Николас К.; Снодграсс, Оуин Э.; Дьюар, Хайди; Хайд, Джон Р. (15 мая 2015 г.). «Энотермия всего тела у мезопелагической рыбы опы, Lampris Guttatus». Наука . 348 (6236): 786–789. Бибкод : 2015Sci...348..786W . дои : 10.1126/science.aaa8902 . ISSN 0036-8075 . ПМИД 25977549 . S2CID 17412022 .
- ^ Голдман, К.Дж. (1997). «Регуляция температуры тела белой акулы Carcharodon carcharias » . Журнал сравнительной физиологии . B Биохимическая системная и экологическая физиология. 167 (6): 423–429. дои : 10.1007/s003600050092 . S2CID 28082417 . Архивировано из оригинала 6 апреля 2012 года . Проверено 12 октября 2011 г.
- ^ Кэри, ФГ; Лоусон, К.Д. (февраль 1973 г.). «Регулирование температуры у свободноплавающего голубого тунца». Сравнительная биохимия и физиология А. 44 (2): 375–392. дои : 10.1016/0300-9629(73)90490-8 . ПМИД 4145757 .
- ^ Гимарайнш-Крус, Родриго Х.; дос Сантос, Хосе Э.; Сантос, Гилмар Б. (июль – сентябрь 2005 г.). «Структура гонад и гаметогенез Loricaria lentiginosa Isbrücker (Pisces, Teleostei, Siluriformes)» . Преподобный Брас. Зоол . 22 (3): 556–564. дои : 10.1590/S0101-81752005000300005 . ISSN 0101-8175 .
- ^ Брито, МФГ; Базцоли, Н. (2003). «Размножение сома-сурубима (Pisces, Pimelodidae) в реке Сан-Франциско, регион Пирапора, Минас-Жерайс, Бразилия» . Бразильский архив ветеринарной медицины и зоотехники . 55 (5): 624–633. дои : 10.1590/S0102-09352003000500018 . ISSN 0102-0935 .
- ^ Скотт, Питер (1997). Живородящие рыбы . Тетра Пресс. п. 13. ISBN 1-56465-193-2 .
- ^ Jump up to: Перейти обратно: а б с Миллер, Брюс; Кендалл, Артур В. (2009). «1. Размножение рыб» (PDF) . История ранней жизни морских рыб . Издательство Калифорнийского университета. стр. 11–37. ISBN 9780520249721 . Архивировано (PDF) из оригинала 7 марта 2016 г.
- ^ Jump up to: Перейти обратно: а б Дей А., Флайшанс М., Пшеничка М., Газо И. Гены репарации ДНК играют разнообразную роль в развитии эмбрионов рыб. Front Cell Dev Biol. 1 марта 2023 г.; 11: 1119229. doi: 10.3389/fcell.2023.1119229. ПМИД 36936683; PMCID: PMC10014602
- ^ Канедо А, Роча TL. Рыбка данио (Danio rerio) в качестве модели для оценки генотоксичности и восстановления ДНК: исторический обзор, современное состояние и тенденции. Научная Тотальная Окружающая среда. 25 марта 2021 г.;762:144084. doi: 10.1016/j.scitotenv.2020.144084. Epub, 14 декабря 2020 г. PMID 33383303
- ^ Хелфман, Коллетт и Фейси, 1997 , стр. 95–96.
- ^ Jump up to: Перейти обратно: а б с Флайник, МФ; Касахара, М. (2010). «Происхождение и эволюция адаптивной иммунной системы: генетические события и селективное давление» . Обзоры природы Генетика . 11 (1): 47–59. дои : 10.1038/nrg2703 . ПМК 3805090 . ПМИД 19997068 .
- ^ Сапата, АГ; Чиба, А.; Вара, А. (1996). «Клетки и ткани иммунной системы рыб». В Иваме, Г. Ивама; Наканиши, Т. (ред.). Иммунная система рыб: организм, возбудитель и окружающая среда . Иммунология рыб. Нью-Йорк: Академическая пресса. стр. 1–55.
- ^ Чильмончик, С. (1992). « Тимус рыб: развитие и возможная функция в иммунном ответе ». Ежегодный обзор болезней рыб . 2 : 181–200. дои : 10.1016/0959-8030(92)90063-4 .
- ^ Хансен, доктор юридических наук; Сапата, АГ (1998). « Развитие лимфоцитов у рыб и амфибий ». Иммунологические обзоры . 166 : 199–220. дои : 10.1111/j.1600-065x.1998.tb01264.x . ПМИД 9914914 . S2CID 7965762 .
- ^ Jump up to: Перейти обратно: а б Питчер, Тони Дж. (1986). «12. Функции мелководного поведения костистых рыб». Поведение костистых рыб . Спрингер. стр. 294–337. дои : 10.1007/978-1-4684-8261-4_12 . ISBN 978-1-4684-8263-8 .
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 375.
- ^ Гьёсетер, Х. (1998). «Популяционная биология и промысел мойвы ( Mallotus villosus ) в Баренцевом море» . Сарсия . 83 (6): 453–496. дои : 10.1080/00364827.1998.10420445 .
- ^ Вайнманн, СР; Блэк, АН; Рихтер, МЛ; Ицковиц, М.; Бургер, РМ (февраль 2017 г.). «Территориальная вокализация у симпатрических стрекоз: акустические характеристики и распознавание злоумышленников». Биоакустика . 27 (1): 87–102. дои : 10.1080/09524622.2017.1286263 . S2CID 89625932 .
- ^ Jump up to: Перейти обратно: а б Бертуччи, Ф.; Руппе, Л.; Вассенберг, СВ; Компер, П.; Пармантье, Э. (29 октября 2014 г.). «Новые сведения о роли глоточно-челюстного аппарата в механизме производства звука Haemulon Flavolineatum (Haemulidae)» . Журнал экспериментальной биологии . 217 (21): 3862–3869. дои : 10.1242/jeb.109025 . hdl : 10067/1197840151162165141 . ПМИД 25355850 .
- ^ Колсон, диджей; Патек, С.Н.; Брейнерд, Эл.; Льюис, С.М. (февраль 1998 г.). «Извлечение звука во время кормления морских коньков гиппокампа (Syngnathidae)». Экологическая биология рыб . 51 (2): 221–229. Бибкод : 1998EnvBF..51..221C . дои : 10.1023/А:1007434714122 . S2CID 207648816 .
- ^ Оливейра, TPR; Ладич, Ф.; Абед-Наванди, Д.; Соуто, А.С.; Роза, Иллинойс (26 июня 2014 г.). «Звуки, издаваемые длиннорылым морским коньком: исследование их строения и функций». Журнал зоологии . 294 (2): 114–121. дои : 10.1111/jzo.12160 .
- ^ Jump up to: Перейти обратно: а б Хорошо, Л.Ф.; Кинг, CB; Кэмерон, ТМ (16 октября 2009 г.). «Акустические свойства плавательного пузыря устрицы-жабы Opsanus tau » . Журнал экспериментальной биологии . 212 (21): 3542–3552. дои : 10.1242/jeb.033423 . ПМЦ 2762879 . ПМИД 19837896 .
- ^ Хорошо, МЛ; Уэйбрайт, Т.Д. (15 октября 2015 г.). «Хрюкающая вариация устрицы-жабы Opsanus tau: влияние размера и пола» . ПерДж . 3 (1330): е1330. дои : 10.7717/peerj.1330 . ПМЦ 4662586 . ПМИД 26623178 .
- ^ Jump up to: Перейти обратно: а б с Риччи, Юго-Запад; Боненштиль, доктор медицинских наук; Эгглстон, Д.Б.; Келлог, ML; Лион, РП (8 августа 2017 г.). «Обнаружение свистков лодок-устриц (Opsanus tau) и закономерности на крупномасштабном участке восстановления устриц» . ПЛОС ОДИН . 12 (8): e0182757. Бибкод : 2017PLoSO..1282757R . дои : 10.1371/journal.pone.0182757 . ПМЦ 5549733 . ПМИД 28792543 .
- ^ Скоглунд, ЧР (1 августа 1961 г.). «Функциональный анализ мышц плавательного пузыря, обеспечивающих здоровую продуктивность рыбы-жабы» . Журнал клеточной биологии . 10 (4): 187–200. дои : 10.1083/jcb.10.4.187 . ПМК 2225107 . ПМИД 19866593 .
- ^ Парментье, Э.; Ток, Дж.; Фальгьер, JC; Бошо, М. (22 мая 2014 г.). «Звуковое производство у Sciaenops ocellatus: предварительное исследование развития акустических сигналов в аквакультуре» (PDF) . Аквакультура . 432 : 204–211. Бибкод : 2014Aquac.432..204P . doi : 10.1016/j.aquacultural.2014.05.017 . Архивировано (PDF) из оригинала 3 июня 2021 года . Проверено 21 января 2019 г.
- ^ «Поиск «Рыб» (глобальный, CR-находящийся под угрозой исчезновения, находящийся под угрозой исчезновения, виды)» . Проверено 27 февраля 2024 г.
- ^ Собел, Дж. (1996). « Гадус морхуа » . Красный список исчезающих видов МСОП . 1996 : e.T8784A12931575. doi : 10.2305/IUCN.UK.1996.RLTS.T8784A12931575.en . Проверено 11 ноября 2021 г.
- ^ ПриродаСерв (2014). « Ципринодон диаболис » . Красный список исчезающих видов МСОП . 2014 : e.T6149A15362335. doi : 10.2305/IUCN.UK.2014-3.RLTS.T6149A15362335.en . Проверено 11 ноября 2021 г.
- ^ Мьюзик, Дж. А. (2000). « Латимерия халюмная » . Красный список исчезающих видов МСОП . 2000 : e.T11375A3274618. doi : 10.2305/IUCN.UK.2000.RLTS.T11375A3274618.en . Проверено 11 ноября 2021 г.
- ^ Ригби, CL; Баррето, Р.; Карлсон, Дж.; Фернандо, Д.; Фордхэм, С.; Фрэнсис, член парламента; и др. (2019). « Кархародон кархариас » . Красный список исчезающих видов МСОП . 2019 : e.T3855A2878674 . Проверено 19 декабря 2019 г.
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 449–450.
- ^ Jump up to: Перейти обратно: а б Гамильтон, Лоуренс К.; Батлер, MJ (январь 2001 г.). «Адаптация к внешнему виду: социальные показатели в условиях кризиса трески в Ньюфаундленде» . Обзор экологии человека . 8 (2): 1–11.
- ^ Состояние мирового рыболовства и аквакультуры в 2020 году . Продовольственная и сельскохозяйственная организация . 2020. с. 54. дои : 10.4060/ca9229en . hdl : 10535/3776 . ISBN 978-92-5-132692-3 . S2CID 242949831 .
- ^ «Призыв остановить чрезмерный вылов трески » . Новости Би-би-си . 5 января 2007 года. Архивировано из оригинала 17 января 2007 года . Проверено 18 января 2006 г.
- ^ «Группы тунца борются с чрезмерным выловом рыбы» . Новости Би-би-си . 26 января 2007 года. Архивировано из оригинала 21 января 2009 года . Проверено 18 января 2006 г.
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 462.
- ^ «Великобритания «должна защищать рыбную промышленность» » . Новости Би-би-си . 3 ноября 2006 г. Архивировано из оригинала 30 ноября 2006 г. Проверено 18 января 2006 г.
- ^ «Соглашение ЕС по квотам на рыбу заключено» . Новости Би-би-си . 21 декабря 2006 г. Архивировано из оригинала 26 декабря 2006 г. Проверено 18 января 2006 г.
- ^ «Исследование океана предсказывает крах всего рыболовства морепродуктов к 2050 году» . физ.орг . Архивировано из оригинала 15 марта 2007 года . Проверено 13 января 2006 г.
- ^ «Атлантический голубой тунец вскоре может исчезнуть с коммерческой точки зрения» . WWF. Архивировано из оригинала 30 апреля 2007 года . Проверено 18 января 2006 г.
- ^ Хелфман, Коллетт и Фейси 1997 , стр. 463.
- ^ Эльвира, Бениньо (1995). «Статус сохранения эндемической пресноводной рыбы в Испании». Биологическая консервация . 72 (2). Эльзевир: 129–136. Бибкод : 1995BCons..72..129E . дои : 10.1016/0006-3207(94)00076-3 . ISSN 0006-3207 .
- ^ Джексон, Дональд С.; Мармулла, Герд (2001). Влияние плотин на речное рыболовство (PDF) . Том. Технический документ 419. Рыболовство ФАО. стр. 1–44.
- ^ Дуплиси, Дэниел Э.; Фриск, Майкл Г.; Тренкель, Верена М. (28 ноября 2016 г.). «Долг вымирания и кредит колонизаторов в рыболовном банке, нарушенном средой обитания» . ПЛОС ОДИН . 11 (11). Публичная научная библиотека (PLoS): e0166409. Бибкод : 2016PLoSO..1166409D . дои : 10.1371/journal.pone.0166409 . ISSN 1932-6203 . ПМК 5125594 . ПМИД 27893775 .
- ^ Jump up to: Перейти обратно: а б Ловелл, Сабрина Дж.; Стоун, Сьюзен Ф.; Фернандес, Линда (2006). «Экономическое воздействие водных инвазивных видов: обзор литературы». Обзор экономики сельского хозяйства и ресурсов . 35 (1): 195–208. дои : 10.1017/S1068280500010157 .
- ^ Ноулер, Д.; Барбье, Э.Б. (2000). «Экономика вторгающихся видов: теоретическая модель и практический пример». В Перрингсе, К.; Уильямсон, М.; Далмаццоне, С. (ред.). Экономика биологических инвазий . Челтнем: Эдвард Элгар. стр. 70–93.
- ^ Голани, Дэниел (1998). «Влияние рыб-мигрантов из Красного моря через Суэцкий канал на водную среду Восточного Средиземноморья». Серия бюллетеней Йельской школы лесного хозяйства и экологических исследований (103): 375–387.
- ^ Коултер, Джордж В.; Аллансон, Брайан Р.; Брутон, Майкл Н.; Гринвуд, П. Хамфри; Харт, Роберт С.; Джексон, Питер Б.Н.; Риббинк, Энтони Дж. (1986). «Уникальные качества и особые проблемы Великих африканских озер». Экологическая биология рыб . 17 (3). Springer Science and Business Media: 161–183. Бибкод : 1986EnvBF..17..161C . дои : 10.1007/bf00698196 . ISSN 0378-1909 .
- ^ Ачиенг, AP (1990). «Влияние интродукции нильского окуня Lates niloticus (L.) на рыболовство в озере Виктория». Журнал биологии рыб . 37, Доп. А: 17–23. Бибкод : 1990JFBio..37S..17A . дои : 10.1111/j.1095-8649.1990.tb05016.x .
- ^ Сполдинг, Марк (11 июля 2013 г.). «Устойчивая древняя аквакультура» . Нэшнл Географик . Архивировано из оригинала 18 мая 2015 года . Проверено 13 августа 2015 г.
- ^ Jump up to: Перейти обратно: а б Хелфман, Джин С. (2007). Сохранение рыбы: Руководство по пониманию и восстановлению глобального водного биоразнообразия и рыбных ресурсов . Остров Пресс. п. 11. ISBN 978-1-59726-760-1 .
- ^ «Селектор морепродуктов EDF: выбор рыбы, полезной для вас и океанов» . Фонд защиты окружающей среды . Проверено 21 января 2024 г.
- ^ ФАО (2022 г.). Состояние мирового рыболовства и аквакультуры в 2022 году. На пути к синей трансформации . Рим: ФАО. дои : 10.4060/cc0461en . hdl : 10535/3776 . ISBN 978-92-5-136364-5 .
- ^ Берд, Т. Дуглас, изд. (2011). Рыболов в окружающей среде: социальные, экономические, биологические и этические аспекты . Бетесда, Мэриленд: Американское общество рыболовства. п. 365. ИСБН 978-1-934874-24-0 .
- ^ Хикли, Фил; Томпкинс, Хелена, ред. (1998). Любительское рыболовство: социальные, экономические и управленческие аспекты . Уайли-Блэквелл. п. 328. ИСБН 978-0-852-38248-6 .
- ^ Jump up to: Перейти обратно: а б с д и Блэк, Джереми; Грин, Энтони (1992). Боги, демоны и символы Древней Месопотамии: Иллюстрированный словарь . Издательство Британского музея. стр. 82–83. ISBN 978-0-7141-1705-8 . Архивировано из оригинала 20 февраля 2018 года.
- ^ Jump up to: Перейти обратно: а б с Хайд, Уолтер Вудберн (2008) [1946]. От язычества к христианству в Римской империи . Юджин, Орегон: Wipf и Stock Publishers. стр. 57–58. ISBN 978-1-60608-349-9 . Архивировано из оригинала 17 марта 2023 года . Проверено 12 декабря 2020 г. .
- ^ Шервуд, Ивонн (2000), Библейский текст и его загробная жизнь: выживание Ионы в западной культуре , Cambridge University Press, стр. 1–8, ISBN 978-0-521-79561-6 , заархивировано из оригинала 17 марта 2023 года , получено 12 декабря 2020 года.
- ^ Коффман, Элеша (8 августа 2008 г.). «Каково происхождение христианского символа рыбы?» . Христианство сегодня . Архивировано из оригинала 30 января 2016 года . Проверено 13 августа 2015 г.
- ^ « 'Нгарара – рептилии, Страница 2. От моря до суши', Те Ара - Энциклопедия Новой Зеландии» . Брэдфорд Хаами . Проверено 4 мая 2018 г.
- ^ Трум, Томас (1907). Гавайские народные сказки . AC МакКлерг . п. 86.
- ^ Бандйопадхьяя, Джаянтануджа (2007). Класс и религия в Древней Индии . Гимн Пресс. п. 136. ИСБН 978-1-84331-332-8 . Архивировано из оригинала 8 октября 2022 года . Проверено 10 июля 2022 г.
- ^ Овидий Фасти 2.457ff
- ^ Мойл, Питер Б .; Мойл, Мэрилин А. (май 1991 г.). «Введение в образы рыб в искусстве». Экологическая биология рыб . 31 (1): 5–23. Бибкод : 1991EnvBF..31....5M . дои : 10.1007/bf00002153 . S2CID 33458630 .
- ^ Тидвелл, Кристи (2009). « Рыбы такие же, как люди, только более хрупкие»: экологическая практика и теория в поисках Немо » . Американа: Журнал американской популярной культуры (8).
- ^ Дурга, П.; Сай, Канака (2017). «Природа экзистенциальной борьбы в «Старик и море ». Журнал английского языка и литературы JOELL . 4 (4): 19–21.
- ^ Алебастр, Джей (2023). «Гуфи-белая: челюсти и наша любовь к высшему хищнику». В Джексоне — Кэти Мерлок; Симпсон, Филип Л. (ред.). Эта акула, проглотит тебя целиком»: Очерки культурного влияния челюстей . МакФарланд. Стр. 124–. ISBN 978-1-4766-7745-3 .
- ^ Золлингер, Сью Энн (3 июля 2009 г.). «Пиранья – свирепый боец или мягкотелый мусорщик?» . Момент науки . Общественные СМИ Индианы. Архивировано из оригинала 17 октября 2015 года . Проверено 1 ноября 2015 г.
Источники
- Бентон, MJ (2005). Палеонтология позвоночных (3-е изд.). Джон Уайли. ISBN 9781405144490 .
- Хельфман, Г.; Коллетт, Б.; Фейси, Д. (1997). Разнообразие рыб (1-е изд.). Уайли-Блэквелл. ISBN 978-0-86542-256-8 .
- Нельсон, Джозеф С. (2016). «Таксономическое разнообразие». Рыбы мира . Джон Уайли и сыновья . ISBN 978-1-118-34233-6 .
Дальнейшее чтение
- Эшмейер, Уильям Н.; Фонг, Джон Дэвид (2013). «Каталог рыб» . Калифорнийская академия наук. Архивировано из оригинала 21 ноября 2018 года . Проверено 28 февраля 2013 г.
- Хельфман, Г.; Коллетт, Б.; Фейси, Д.; Боуэн, Б. (2009). Разнообразие рыб: биология, эволюция и экология (2-е изд.). Уайли-Блэквелл. ISBN 978-1-4051-2494-2 . Архивировано из оригинала 26 августа 2021 года . Проверено 26 января 2010 г.
- Мойл, Питер Б. (1993) Рыба: Путеводитель для энтузиастов. Архивировано 17 марта 2023 года в издательстве Wayback Machine University of California Press. ISBN 978-0-520-91665-4 – хороший нестандартный текст.
- Мойл, Питер Б.; Чех, Джозеф Дж. (2003). Рыбы, Введение в ихтиологию (5-е изд.). Бенджамин Каммингс. ISBN 978-0-13-100847-2 .
- Весы, Хелен (2018). Глаз мелководья: Путеводитель для рыболовов по жизни, океану и всему остальному . Блумсбери Сигма. ISBN 978-1-4729-3684-4 .
- Шубин, Нил (2009). Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела . Винтажные книги. ISBN 978-0-307-27745-9 . Архивировано из оригинала 17 марта 2023 года . Проверено 15 декабря 2015 г. UCTV. Интервью Архивировано 14 января 2021 г. в Wayback Machine.
Внешние ссылки
- ANGFA – Иллюстрированная база данных пресноводных рыб Австралии и Новой Гвинеи.
- FishBase онлайн – обширная база данных с информацией о более чем 29 000 видах рыб.
- Центр рыболовства и аквакультуры Иллинойса - источник данных для исследовательского центра рыболовства и аквакультуры в центральной части США на archive.today (архивировано 15 декабря 2012 г.)
- The Native Fish Conservancy - Сохранение и изучение пресноводных рыб Северной Америки на Wayback Machine (архивировано 12 марта 2008 г.)
- Организация Объединенных Наций – Департамент рыболовства и аквакультуры: Использование рыбы и морепродуктов
| О рыбе |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| сенсорный системы | |||||
| Воспроизведение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| По среде обитания | |||||
| Другие типы | |||||
| Коммерция |
| ||||
| Основные группы | |||||
| Списки | |||||


морской водной экосистемы Компоненты |
|---|
