Размножение рыб

рыб К репродуктивным органам относятся семенники и яичники . У большинства видов гонады представляют собой парные органы одинакового размера, которые могут частично или полностью слиться. [1] Также может существовать ряд вторичных органов, повышающих репродуктивную способность. Генитальный сосочек представляет собой небольшую мясистую трубку позади ануса у некоторых рыб, из которой сперматозоиды или яйца выделяются ; пол рыбы часто можно определить по форме ее сосочка.
Анатомия
[ редактировать ]Яички
[ редактировать ]У большинства самцов рыб есть два семенника одинакового размера. У акул семенники с правой стороны обычно крупнее. [ нужна ссылка ] . У примитивных бесчелюстных рыб имеется только один семенник, расположенный по средней линии тела, хотя и он образуется в результате слияния парных структур в зародыше. [2]
Белая оболочка, рыб, под прочной перепончатой оболочкой семенник некоторых костистых содержит очень тонкие спиральные трубки, называемые семенными канальцами . Канальцы выстланы слоем клеток ( зародышевых клеток ), которые от полового созревания до старости развиваются в сперматозоиды или (также известные как сперматозоиды мужские гаметы ). Развивающиеся сперматозоиды проходят через семенные канальцы к сети семенника , расположенной в средостении яичка , к выносящим протокам , а затем к придатку яичка , где созревают вновь созданные сперматозоиды (см. Сперматогенез ). Сперматозоиды попадают в семявыносящий проток и в конечном итоге выбрасываются через уретру и из отверстия уретры посредством мышечных сокращений.
Однако у большинства рыб семенные канальцы отсутствуют. Вместо этого сперматозоиды производятся в сферических структурах, называемых ампулами сперматозоидов . Это сезонные структуры, выделяющие свое содержимое в период размножения, а затем реабсорбируемые организмом. Перед следующим сезоном размножения начинают формироваться и созревать новые ампулы сперматозоидов. В остальном ампулы по существу идентичны семенным канальцам высших позвоночных , включая тот же диапазон типов клеток. [2]
По распределению сперматогоний строение семенников костистых рыб имеет два типа: у наиболее распространенных сперматогонии располагаются по всему ходу семенных канальцев , а у атериноморфных рыб они приурочены к дистальной части этих структур. У рыб может наблюдаться кистозный или полукистозный сперматогенез в зависимости от фазы выхода зародышевых клеток из кист в просвет семенных канальцев . [1]
Яичники
[ редактировать ]
Многие особенности яичников являются общими для всех позвоночных, включая наличие фолликулярных клеток и белочной оболочки. В яичнике рыбы в любой момент времени могут присутствовать сотни или даже миллионы оплодотворенных яиц. Свежие яйца могут развиваться из зародышевого эпителия на протяжении всей жизни. Желтые тела встречаются только у млекопитающих и у некоторых пластиножаберных рыб; у других видов остатки фолликула быстро рассасываются яичником. [2] Яичник костистых рыб часто содержит полое, заполненное лимфой пространство, которое открывается в яйцевод и куда откладываются яйца. [2] У большинства нормальных самок рыб есть два яичника. У некоторых пластиножаберных полностью развивается только правый яичник. У примитивных бесчелюстных рыб и некоторых костистых рыб имеется только один яичник, образовавшийся в результате слияния парных органов в зародыше. [2]
Яичники рыб могут быть трех типов: гимнновариальные, вторично-гимновариальные и цистовариальные. При первом типе ооциты выходят непосредственно в целомическую полость, затем поступают в устье , затем через яйцевод и элиминируются. Вторичные гимноварианские яичники выделяют яйцеклетки в целом , откуда они попадают непосредственно в яйцевод. При третьем типе ооциты выводятся наружу через яйцевод . [3] Гимноварии — примитивное состояние, встречающееся у двоякодышащих рыб , осетровых и боуфинов . Кистовариумы характерны для большинства костистых рыб, у которых просвет яичника неразрывно связан с яйцеводом. [1] Вторичные гимноварии встречаются у лососевых и некоторых других костистых рыб.
Яйца
[ редактировать ]

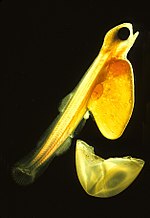
Икра рыб и земноводных желеобразная. Яйца хрящевых рыб (акулы, скаты, скаты, химеры) оплодотворяются внутренне и демонстрируют большое разнообразие как внутреннего, так и внешнего эмбрионального развития. Большинство видов рыб выметывают икру, которая оплодотворяется извне, обычно самец оплодотворяет икру после того, как самка ее откладывает. Эти яйца не имеют скорлупы и высыхают на воздухе. Даже дышащие воздухом земноводные откладывают яйца в воду или в защитную пену, как, например, прибрежная древесная лягушка Chiromantis xerampelina , дышащая воздухом .
Органы-нарушители
[ редактировать ]

Самцы хрящевых рыб (акулы и скаты), а также самцы некоторых живородящих лучепёрых рыб имеют плавники, которые были модифицированы и функционируют как интромиттивные органы , репродуктивные придатки, которые обеспечивают внутреннее оплодотворение . У лучепёрых рыб они называются гоноподиями или андроподиями , а у хрящевых рыб — класперами .
Гоноподии встречаются у самцов некоторых видов семейств Anablepidae и Poeciliidae . Это анальные плавники, которые были модифицированы и функционируют как подвижные внутренние органы и используются для оплодотворения самок молоком во время спаривания. Третий, четвертый и пятый лучи анального плавника самца сформированы в трубчатую структуру, в которую выбрасывается сперма рыбы. [6] Когда гонопий готов к спариванию, он становится прямым и направлен вперед в сторону самки. Самец вскоре вставляет орган в половое отверстие самки с помощью крючкообразных приспособлений, которые позволяют рыбе захватывать самку, чтобы обеспечить оплодотворение. Если самка остается неподвижной, а ее партнер касается ее отверстия своим гоноподием, она оплодотворяется. Сперма сохраняется в яйцеводе самки. Это позволяет самкам оплодотворить себя в любое время без дополнительной помощи самцов. У некоторых видов гоноподий может составлять половину общей длины тела. Иногда плавник слишком длинный, чтобы его можно было использовать, как, например, у лирохвостых пород Xiphophorus helleri . У женщин, получающих гормональные препараты, могут развиться гоноподии. Они бесполезны для разведения.
Подобные органы со схожими характеристиками встречаются и у других рыб, например, андроподия у Hemirhamphodon или Goodeidae . [7]
Класперы встречаются у самцов хрящевых рыб . самки Они представляют собой заднюю часть брюшных плавников, которые также были модифицированы для выполнения функций интромиттирующих органов и используются для направления спермы в клоаку во время совокупления. Акт спаривания у акул обычно включает в себя поднятие одной из застежек, чтобы вода попала в сифон через определенное отверстие . Затем застежку вставляют в клоаку, где она раскрывается как зонтик, фиксируя свое положение. Затем сифон начинает сокращаться, вытесняя воду и сперму. [8] [9]
Физиология
[ редактировать ]Развитие оогоний у костистых рыб варьируется в зависимости от группы, а определение динамики оогенеза позволяет понять процессы созревания и оплодотворения. Изменения в ядре , ооплазме и окружающих слоях характеризуют процесс созревания ооцита. [1]
Постовуляторные фолликулы представляют собой структуры, образующиеся после выхода ооцита; они не выполняют эндокринной функции, имеют широкий неровный просвет и быстро реабсорбируются в процессе, включающем апоптоз фолликулярных клеток. Дегенеративный процесс, называемый фолликулярной атрезией, реабсорбирует непророжденные вителлогенные ооциты. Этот процесс также может происходить, но реже, в ооцитах на других стадиях развития. [1]
Некоторые рыбы являются гермафродитами , имеющими семенники и яичники либо на разных стадиях жизненного цикла, либо, как в деревнях , имеют их одновременно.
Репродуктивные стратегии
[ редактировать ]У рыб оплодотворение икры может быть как наружным, так и внутренним. У многих видов рыб плавники модифицированы для обеспечения внутреннего оплодотворения . Точно так же развитие эмбриона может быть внешним или внутренним, хотя у некоторых видов наблюдаются различия между ними на разных стадиях развития эмбриона. Тьерри Лоде описал репродуктивные стратегии с точки зрения развития зиготы и взаимоотношений с родителями; Существует пять классификаций - яйцерождение, яйцерождение, яйцеживорождение, гистотрофное живорождение и гемотрофное живорождение. [10]
Яйцерождение
[ редактировать ]Яйцерождение означает, что самка откладывает неоплодотворенные яйца (яйцеклетки), которые затем необходимо оплодотворить извне. [10] Примеры яйцекладущих рыб включают лосося , золотую рыбку , цихлид , тунца и угрей . У большинства этих видов оплодотворение происходит вне тела матери, при этом самцы и самки сбрасывают свои гаметы в окружающую воду.
Яйцерождение
[ редактировать ]Яйцерождение — это когда оплодотворение происходит внутри организма, и тогда самка сбрасывает зиготы (или вновь развивающиеся эмбрионы) в воду. [10] часто с добавлением важных внешних тканей. Более 97% всех известных рыб являются яйцекладущими (требуется подтверждение, поскольку яйцекладание - это новый термин, который можно спутать с яйцерождением. Если используется яйцерождение, большинство рыб имеют стратегию размножения яйцекладущую). [11] У яйцекладущих рыб внутреннее оплодотворение требует, чтобы самец использовал какой-то интромиттирующий орган для доставки спермы в половое отверстие самки. Примеры включают яйцекладущих акул, таких как рогатая акула , и яйцекладущих скатов, таких как скаты . В этих случаях самец оснащен парой модифицированных брюшных плавников, известных как класперы .
Морская рыба может производить большое количество икры, которая часто выбрасывается в толщу открытой воды. Яйца имеют средний диаметр 1 миллиметр (0,039 дюйма). Яйца обычно окружены внезародышевыми оболочками , но вокруг этих оболочек не образуется твердая или мягкая оболочка. У некоторых рыб толстая кожистая шерсть, особенно если им приходится выдерживать физическую силу или высыхание. Яйца такого типа также могут быть очень маленькими и хрупкими.
 Яйцо миноги
Яйцо миноги Яйцо кошачьей акулы ( кошелек русалки )
Яйцо кошачьей акулы ( кошелек русалки ) Яйцо бычьей акулы
Яйцо бычьей акулы Яйцо химеры
Яйцо химеры

Только что вылупившуюся молодь яйцекладущих рыб называют личинками . Они обычно плохо сформированы, несут большой желточный мешок (для питания) и по внешнему виду сильно отличаются от молодых и взрослых особей. Личиночный период у яйцекладущих рыб относительно короткий (обычно всего несколько недель), личинки быстро растут и меняют внешний вид и структуру (процесс, называемый метаморфозом ), превращаясь в молодь. Во время этого перехода личинки должны переключиться с желточного мешка на питание добычей зоопланктона , этот процесс обычно зависит от недостаточной плотности зоопланктона, приводящей к голоданию многих личинок.
Яйцеживорождение
[ редактировать ]У яйцеживородящих рыб икра развивается внутри тела матери после внутреннего оплодотворения, но получает мало или вообще не получает питания непосредственно от матери, вместо этого в зависимости от запаса пищи внутри яйца, желтка . [10] Каждый эмбрион развивается в своей яйцеклетке. Известные примеры яйцеживородящих рыб включают гуппи , акулы-ангелы и целаканты .
Живорождение
[ редактировать ]Существует два типа живорождения , различающиеся тем, как потомство получает питательные вещества.
- Гистотрофное (поедание тканей) живорождение означает, что эмбрионы развиваются в яйцеводах самки, но получают питательные вещества, потребляя другие ткани, такие как яйцеклетки ( оофагия ) или зиготы. [10] Это наблюдалось в первую очередь среди акул, таких как короткоплавник мако и porbeagle , но также известно и для некоторых костистых рыб, таких как полуклюв Nomorhamphus ebrardtii . [12] Необычным способом вивипирования является адельфофагия или внутриутробный каннибализм , при котором самые крупные эмбрионы поедают более слабых и меньших неродившихся братьев и сестер. Это чаще всего встречается среди акул, таких как серая акула-нянька , но также сообщалось и о Nomorhamphus ebrardtii . [12]
- Гемотрофное (поедание крови) живорождение означает, что эмбрионы развиваются в яйцеводе самки (или самца), а питательные вещества предоставляются непосредственно родителем, обычно через структуру, подобную или аналогичную плаценте, наблюдаемой у млекопитающих . [10] Примеры гемотрофных рыб включают серфингистов , расщепленных плавников , лимонную акулу , морских коньков и игл .
Аквариумисты обычно называют яйцеживородящих и живородящих рыб живородящими .
Гермафродитизм
[ редактировать ]


Гермафродитизм возникает, когда данная особь вида обладает как мужскими, так и женскими репродуктивными органами или может попеременно обладать то одним, то другим. Гермафродитизм часто встречается у беспозвоночных, но редко у позвоночных. Его можно противопоставить гонохоризму , при котором каждая особь вида является либо самцом, либо самкой и остается таковой на протяжении всей своей жизни. Большинство рыб — гонохористы, но известно, что гермафродитизм встречается у 14 семейств костистых рыб. [13]
Обычно гермафродиты являются последовательными , то есть могут менять пол , обычно с женского на мужской ( протогиния ). Это может произойти, если из группы самок удалить доминирующего самца. Самая крупная самка в гареме может за несколько дней сменить пол и заменить доминирующего самца. [13] Он встречается среди рыб коралловых рифов, таких как морские окуни , рыбы-попугаи и губаны . Реже самец переходит в самку ( протандрия ). [14] : 162 Например, большинство губанов являются протогинными гермафродитами в рамках гаремной системы спаривания. [15] [16] Гермафродитизм допускает сложные системы спаривания. Губаны демонстрируют три различные системы спаривания: полигинную, токообразную и беспорядочную. [17] Групповой нерест и парный нерест происходят в системах спаривания. Тип нереста зависит от размера тела самца. [16] Лаброиды обычно нерестятся широко, выделяя большое количество планктонных яиц, которые разносятся приливными течениями; взрослые губаны не взаимодействуют с потомством. [18] Губаны особой подгруппы семейства Labridae, Labrini, не имеют массового нереста.
Реже гермафродиты могут быть синхронными , то есть они одновременно обладают и яичниками, и яичками и могут функционировать как представители любого пола в любой момент времени. Черные деревушки «по очереди выпускают сперму и икру во время нереста. Поскольку такая торговля икрой выгодна обеим особям, деревни обычно моногамны в течение коротких периодов времени - необычная ситуация для рыб». [19] Пол многих рыб не фиксирован, но может меняться в зависимости от физических и социальных изменений среды, в которой обитают рыбы. [20]
Особенно среди рыб гермафродитизм может окупиться в ситуациях, когда у одного пола больше шансов выжить и размножиться, возможно, потому, что он крупнее. [21] Рыбы-анемоны — последовательные гермафродиты, которые рождаются самцами и становятся самками только в зрелом возрасте. Рыбы-анемоны моногамно живут вместе в анемоне , защищенном укусами анемона. Самцам не приходится конкурировать с другими самцами, а самки анемонов обычно крупнее. Когда самка умирает, молодь (самец) рыбы-анемоны приближается, и «резидентный самец затем превращается в самку, и репродуктивные преимущества комбинации крупной самки и маленького самца сохраняются». [22] У других рыб смена пола обратима. Например, если некоторых бычков сгруппировать по полу (самцы или самки), некоторые поменяют пол. [14] : 164 [21]
Мангровый ручей Kryptolebias marmoratus производит как яйца, так и сперму путем мейоза и обычно размножается путем самооплодотворения. Каждый отдельный гермафродит обычно оплодотворяет себя, когда яйцеклетка и сперма, произведенная внутренним органом, соединяются внутри тела рыбы. [23] В природе этот способ размножения может давать высоко гомозиготные линии, состоящие из особей, настолько генетически однородных, что фактически идентичны друг другу. [24] [25] Способность к самоопылению у этих рыб, по-видимому, сохраняется по крайней мере несколько сотен тысяч лет. [26]
Хотя инбридинг , особенно в крайней форме самооплодотворения, обычно считается вредным, поскольку он приводит к проявлению вредных рецессивных аллелей, самооплодотворение действительно обеспечивает преимущество гарантии оплодотворения (гарантии воспроизводства) в каждом поколении. [24]
Половой паразитизм
[ редактировать ]
Половой паразитизм — это способ полового размножения , уникальный для удильщиков , при котором самцы вида намного меньше самок и полагаются на самок в качестве пищи и защиты от хищников. Самцы ничего не отдают взамен, кроме спермы , которая необходима самкам для производства следующего поколения.
Некоторые удильщики, например, представители глубоководной группы цератиоидов , используют этот необычный метод спаривания. Поскольку особи распределены очень редко, встречи также очень редки. Поэтому найти пару проблематично. Когда ученые впервые начали ловить цератиоидных удильщиков, они заметили, что все экземпляры были самками. Эти особи были размером в несколько сантиметров, и почти у всех из них было прикреплено что-то, похожее на паразитов . Оказалось, что этими «паразитами» были сильно редуцированные самцы цератиоидных удильщиков. Это указывает на то, что удильщики используют полиандрическую систему спаривания.
Методы, с помощью которых удильщики находят себе пару, различаются. У некоторых видов крошечные глаза, непригодные для идентификации самок, в то время как у других недоразвиты ноздри, что делает маловероятным их эффективное обнаружение самок по обонянию. [27] Когда самец находит самку, он кусает ее кожу и выделяет фермент , который переваривает кожу его рта и ее тела, сплавляя пару до уровня кровеносных сосудов. [28] Самец становится зависимым от самки-хозяина в плане выживания, получая питательные вещества через свою теперь общую систему кровообращения и взамен предоставляя сперму самке. После слияния самцы увеличиваются в объёме и становятся значительно крупнее свободноживущих самцов вида. Они живут и остаются репродуктивно функциональными, пока жива самка, и могут принимать участие в нескольких нерестах. [27] Этот крайний половой диморфизм гарантирует, что, когда самка будет готова к нересту, у нее сразу же появится партнер. [29] Несколько самцов могут быть объединены в одну особь самки, причем у некоторых видов может быть до восьми самцов, хотя для некоторых таксонов, по-видимому, существует правило: один самец на самку. [27] В дополнение к физиологическим адаптациям иммунная система изменяется, чтобы обеспечить соединение. [30]
Другой формой полового паразитизма является гибридогенез , при котором используются репродуктивные усилия другого вида. Одним из таких примеров является вид рода Poeciliopsis . Эта рыба состоит только из самок, которые спариваются с самцами близкородственного вида. Все отцовские хромосомы отбрасываются во время производства яиц. Самки передают свой материнский геном следующему поколению, в отличие от самцов других видов. [31]
Одним из объяснений эволюции полового паразитизма является то, что относительная низкая плотность самок в глубоководной среде оставляет мало возможностей для выбора партнера среди удильщиков. Самки остаются крупными, чтобы обеспечить плодовитость , о чем свидетельствуют их большие яичники и яйца. Ожидается, что самцы уменьшатся в размерах, чтобы снизить метаболические затраты в условиях скудных ресурсов, и у них разовьются узкоспециализированные способности по поиску самок. Если самцу удается найти паразитическую привязанность самки, то в конечном итоге у него больше шансов улучшить приспособленность на всю жизнь по сравнению со свободной жизнью, особенно когда перспектива найти будущих партнеров невелика. Дополнительным преимуществом паразитизма является то, что сперму самца можно использовать при многократном оплодотворении, поскольку она всегда остается доступной самке для спаривания. Более высокая плотность встреч самцов и самок может коррелировать с видами, которые демонстрируют факультативный паразитизм или просто используют более традиционное временное контактное спаривание. [32]
Партеногенез
[ редактировать ]Партеногенез — форма бесполого размножения , при которой рост и развитие зародышей происходят без оплодотворения . У животных партеногенез означает развитие зародыша из неоплодотворенной яйцеклетки . Первое женское (однополое) размножение позвоночных было описано у амазонской моллинезии в 1932 году. [33] С тех пор было описано не менее 50 видов однополых позвоночных, в том числе не менее 20 рыб, 25 ящериц, один вид змей, лягушек и саламандр. [34] Как и в случае со всеми типами бесполого размножения, с партеногенезом связаны как издержки (низкое генетическое разнообразие и, следовательно, восприимчивость к неблагоприятным мутациям), так и преимущества (воспроизведение без необходимости в самце).
У акул подтвержден партеногенез у чепчиковых голов. [35] и акула-зебра . [36] Другие, обычно половые виды, могут иногда размножаться партеногенетически, а акулы-молот и черноперые акулы [37] являются недавними дополнениями к известному списку факультативно партеногенетических позвоночных.
Частным случаем партеногенеза является гиногенез . стимулируется просто наличием сперматозоида — При этом типе размножения потомство производится по тому же механизму, что и при партеногенезе, однако развитие яйцеклетки сперматозоиды не передают потомству никакого генетического материала. Поскольку все гиногенетические виды являются самками, активация их яиц требует спаривания с самцами близкородственного вида для получения необходимого стимула. Амазонская молли (на фото) размножается гиногенезом.
Другие
[ редактировать ]Лосорог-бычок ( Alcichthys elongatus ) — морское костистое животное с уникальным репродуктивным способом, называемым «внутренней гаметической ассоциацией». Сперматозоиды вводятся в яичник путем копуляции и затем попадают в микропилярный канал овулировавшей яйцеклетки в полость яичника. Однако фактического слияния сперматозоида и яйцеклетки не происходит до тех пор, пока яйцеклетки не попадут в морскую воду. [38]
Инбридинг
[ редактировать ]Инбредная депрессия
[ редактировать ]Влияние инбридинга на репродуктивное поведение изучали на пецилидных рыбках Heterandria formosa . [39] Было обнаружено, что одно поколение спаривания полнородных братьев снижает репродуктивные способности и, вероятно, репродуктивный успех потомства мужского пола. Другими признаками, демонстрирующими инбредную депрессию, были жизнеспособность потомства и время созревания как самцов, так и самок.
Воздействие на рыбу-зебру химического агента окружающей среды, аналогичного тому, которое вызвано антропогенным загрязнением, усилило влияние инбридинга на ключевые репродуктивные признаки. [40] Жизнеспособность эмбрионов была значительно снижена у инбредных рыб, подвергшихся воздействию, и наблюдалась тенденция у инбредных самцов давать меньше потомства.
Поведение молоди кижуча с низким и средним инбридингом сравнивали в парных состязаниях. [41] Рыбы с низким инбридингом проявляли почти в два раза более агрессивное преследование при защите территории, чем рыбы со средним инбридингом, а также имели более высокий удельный темп роста. Также было обнаружено значительное влияние инбридинговой депрессии на выживаемость молоди, но только в конкурентной среде с высокой плотностью населения, что позволяет предположить, что внутривидовая конкуренция может усилить пагубные последствия инбридинга.
Избегание инбридинга
[ редактировать ]Инбридинг обычно имеет негативные последствия для приспособленности (инбредная депрессия), и в результате виды выработали механизмы, позволяющие избегать инбридинга. многочисленные механизмы предотвращения инбридинга, Описаны действующие до спаривания. Однако механизмы предотвращения инбридинга, действующие после совокупления, менее известны. У гуппи возникает посткопулятивный механизм избежания инбридинга, основанный на конкуренции между сперматозоидами конкурирующих самцов за достижение оплодотворения . [42] В соревнованиях между сперматозоидами неродственного самца и полнокровного самца наблюдалось значительное смещение отцовства в сторону неродственного самца. [42]
Считается, что инбредная депрессия во многом обусловлена проявлением гомозиготных вредных рецессивных мутаций. [43] Ауткроссинг между неродственными особями приводит к полезной маскировке вредных рецессивных мутаций в потомстве. [44]
Сексуальные стратегии
[ редактировать ]Стратегии нереста
[ редактировать ]Нерестилища
[ редактировать ]Примеры
[ редактировать ]

- Золотая рыбка
Золотые рыбки , как и все карповые , являются яйцекладущими. Обычно они приступают к размножению после значительного изменения температуры, часто весной. Самцы преследуют самок, побуждая их выпустить яйца, ударяя и подталкивая их. Пока самка золотой рыбки выметывает икру, самец остается рядом и оплодотворяет ее. Их яйца клейкие и прикрепляются к водной растительности. Яйца вылупляются в течение 48–72 часов. Примерно через неделю мальки начинают принимать окончательную форму, хотя может пройти год, прежде чем они приобретут зрелый цвет золотой рыбки; до тех пор они имеют металлически-коричневый цвет, как и их дикие предки. В первые недели жизни мальки растут быстро — адаптация, вызванная высоким риском быть съеденными взрослой золотой рыбкой.
- Карп
Карп, представитель семейства карповых, нерестится в период с апреля по август, что во многом зависит от климата и условий, в которых они живут. Уровень кислорода в воде, наличие пищи, размер каждой рыбы, возраст, количество раз, когда рыба нерестилась. до и температура воды - все это известные факторы, влияющие на то, когда и сколько икринок каждый карп будет нерестить одновременно. [45]
- Сиамская бойцовая рыбка
Перед нерестом самцы сиамских бойцовских рыб строят пузырьковые гнезда на поверхности воды. Когда самец начинает интересоваться самкой, он раздувает жабры, крутит тело и расправляет плавники. Самка темнеет и изгибает тело вперед и назад. Акт нереста происходит в «брачных объятиях», когда самец обвивает свое тело вокруг самки, каждое объятие приводит к высвобождению 10-40 икринок до тех пор, пока у самки не закончатся икринки. Самец со своей стороны выпускает молоки в воду и оплодотворение происходит снаружи. Во время и после нереста самец использует рот, чтобы подобрать тонущую икру и отложить ее в пузырьковое гнездо. Как только самка выпустит все яйца, ее прогоняют с территории самца, так как вполне вероятно, что она съест яйца из-за голода. [46] Яйца остаются на попечении самца. Он держит их в пузырьковом гнезде, следя за тем, чтобы ни один из них не упал на дно, и при необходимости ремонтирует гнездо. Инкубация длится 24–36 часов, а только что вылупившиеся личинки остаются в гнезде следующие 2–3 дня, пока их желточные мешки полностью не рассасываются. После этого мальки покидают гнездо и наступает стадия свободного плавания. [47]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Гимарайнш-Крус, Родриго Х., Родриго Х.; Сантос, Хосе Э. дос; Сантос, Гилмар Б. (2005). «Структура гонад и гаметогенез Loricaria lentiginosa Isbrücker (Pisces, Teleostei, Siluriformes)» . Преподобный Брас. Зоол . 22 (3): 556–564. дои : 10.1590/S0101-81752005000300005 . ISSN 0101-8175 .
- ^ Jump up to: а б с д и Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 385–386. ISBN 978-0-03-910284-5 .
- ^ Брито, МФГ; Базцоли, Н. (2003). «Размножение сома-сурубима (Pisces, Pimelodidae) в реке Сан-Франциско, регион Пирапора, Минас-Жерайс, Бразилия» . Бразильский архив ветеринарной медицины и зоотехники . 55 (5): 624–633. дои : 10.1590/S0102-09352003000500018 . ISSN 0102-0935 .
- ^ Мастерсон, Дж. « Gambusia affinis » . Смитсоновский институт . Проверено 21 октября 2011 г.
- ^ Кунц, Альберт (1913). «Заметки о повадках, морфологии репродуктивных органов и эмбриологии живородящих рыб Gambusia affinis» . Бюллетень Бюро рыболовства США . 33 : 181–190.
- ^ Капур Б.Г. и Ханна Б. (2004) Справочник по ихтиологии, стр. 497–498, Springer Science & Business Media. ISBN 9783540428541 .
- ^ Хелфман Г., Коллетт Б.Б., Фейси Д.Х. и Боуэн Б.В. (2009) Разнообразие рыб: биология, эволюция и экология, с. 35, Уайли-Блэквелл. ISBN 978-1-4051-2494-2 .
- ^ «Системный словарь» . ФишБаза . Проверено 15 февраля 2013 г.
- ^ Хейнике, Мэтью П.; Нэйлор, Гэвин Дж. П.; Хеджес, С. Блэр (2009). Древо жизни: хрящевые рыбы (Chondrichthyes) . Издательство Оксфордского университета. п. 320. ИСБН 978-0191560156 .
- ^ Jump up to: а б с д и ж Лоде, Т. (2012). «Яйцерождение или живорождение? Вот в чем вопрос…». Репродуктивная биология . 12 (3): 259–264. дои : 10.1016/j.repbio.2012.09.001 . ПМИД 23153695 .
- ^ Питер Скотт: Живородящие рыбы , с. 13. Тетра Пресс, 1997. ISBN 1-56465-193-2
- ^ Jump up to: а б Мейснер, А; Бернс, Дж (1997). «Живорождение полуклювов родов Dermogenys и Nomorhamphus (Teleostei: Hemiramphidae)». Журнал морфологии . 234 (3): 295–317. doi : 10.1002/(SICI)1097-4687(199712)234:3<295::AID-JMOR7>3.0.CO;2-8 . ПМИД 29852651 . S2CID 46922423 .
- ^ Jump up to: а б Шапиро Д.Ю. (1984) «Реверсия пола и социально-демографические процессы у коралловых рифовых рыб», страницы 103–116 в Г.В. Поттсе и Р.К. Вутуне, ред., Размножение рыб: стратегии и тактика , Academic Press.
- ^ Jump up to: а б Мойл П.Б. и Чех Дж.Дж. (2004) Рыбы, Введение в ихтиологию. 5-е изд, Бенджамин Каммингс. ISBN 978-0-13-100847-2
- ^ Робертсон, доктор медицинских наук; Р. Р. Уорнер (1978). «Половые особенности лаброидов Западного Карибского бассейна II: рыбы-попугаи (Scaridae)». Смитсоновский вклад в зоологию . 255 (255): 1–26. дои : 10.5479/si.00810282.255 .
- ^ Jump up to: а б Казанчиоглу, Э.; С.Х. Алонзо (август 2010 г.). «Сравнительный анализ смены пола у Labridae подтверждает гипотезу преимущества в размерах» . Эволюция . 64 (8): 2254–2264. дои : 10.1111/j.1558-5646.2010.01016.x . ПМИД 20394662 .
- ^ Колин, Польша; Эл Джей Белл (1992). «Аспекты нереста лабридных и скаридовых рыб (Pisces, Labroidei) на атолле Эниветак, Маршалловы острова с примечаниями о других семействах (исправленная перепечатка)» . Экологическая биология рыб . 33 (3): 330–345. дои : 10.1007/BF00005881 .
- ^ Ханель, Р.; МВт Вестнит; К. Штурмбауэр (декабрь 2002 г.). «Филогенетические отношения, эволюция поведения по уходу за выводком и географическое видообразование в племени губанов Лабрини». Журнал молекулярной эволюции . 55 (6): 776–789. Бибкод : 2002JMolE..55..776H . дои : 10.1007/s00239-002-2373-6 . ПМИД 12486536 . S2CID 3002410 .
- ^ Фишер Э.А., Петерсон К.В. (1987). «Эволюция сексуальности морских окуней». Бионаука . 37 (7): 482–489. дои : 10.2307/1310420 . JSTOR 1310420 .
- ^ Чан СТХ и Юнг ВСБ (1983) «Контроль над сексом и смена пола у рыб в естественных условиях». Страницы 171–222 в WS Hoar, DJ Randall и EM Donaldson, ред., Физиология рыб 9B: Размножение, поведение и контроль фертильности . Академическая пресса.
- ^ Jump up to: а б Кувамура, Тецуо; Накашима, Ясухиро (1998). «Новые аспекты смены пола среди рифовых рыб: недавние исследования в Японии». Экологическая биология рыб . 52 (1–3): 125–135. Бибкод : 1998EnvBF..52..125K . дои : 10.1023/А:1007389519702 . S2CID 30244987 .
- ^ Фрике, Ганс; Фрике, Симона (1977). «Моногамия и смена пола в результате агрессивного доминирования у коралловых рифовых рыб». Природа . 266 (5605): 830–832. Бибкод : 1977Natur.266..830F . дои : 10.1038/266830a0 . ПМИД 865603 . S2CID 4177265 .
- ^ Сакакура, Ю.; Сояно, К.; Ноукс, DLG; Хагивара, А. (2006). «Морфология гонад у самооплодотворяющейся мангровой рыбы-киллифиса Kryptolebias marmoratus». Ихтиологические исследования . 53 (4): 427–430. Бибкод : 2006IchtR..53..427S . дои : 10.1007/s10228-006-0362-2 . hdl : 10069/35713 . S2CID 9474211 .
- ^ Jump up to: а б Авизе Ю.С., Татаренков А (2012). «Аргумент Алларда против утверждения Бейкера об адаптивном значении самоопыления у гермафродитных рыб» . Учеб. Натл. акад. наук. США . 109 (46): 18862–7. Бибкод : 2012PNAS..10918862A . дои : 10.1073/pnas.1217202109 . ПМЦ 3503157 . ПМИД 23112206 .
- ^ Эрли Р.Л., Ханнинен А.Ф., Фуллер А., Гарсия М.Дж., Ли Э.А. (2012). «Фенотипическая пластичность и интеграция мангрового ручья (Kryptolebias marmoratus): проспект» . Интегр. Комп. Биол . 52 (6): 814–27. дои : 10.1093/icb/ics118 . ПМК 3501102 . ПМИД 22990587 .
- ^ Татаренков А, Лима С.М., Тейлор Д.С., Avise JC (2009). «Длительное сохранение самоопыления в кладе рыб» . Учеб. Натл. акад. наук. США . 106 (34): 14456–9. Бибкод : 2009PNAS..10614456T . дои : 10.1073/pnas.0907852106 . ПМЦ 2732792 . ПМИД 19706532 .
- ^ Jump up to: а б с Питч, Теодор В. (25 августа 2005 г.). «Возвращение к диморфизму, паразитизму и полу: способы размножения глубоководных цератиоидных удильщиков (Teleostei: Lophiiformes)». Ихтиологические исследования . 52 (3): 207–236. Бибкод : 2005IchtR..52..207P . дои : 10.1007/s10228-005-0286-2 . S2CID 24768783 .
- ^ Гулд, Стивен Джей (1983). Куриные зубы и лошадиные пальцы . Нью-Йорк: WW Norton & Company. ISBN 978-0-393-01716-8 .
- ^ Теодор В. Питч (июль 1975 г.). «Преждевременный половой паразитизм у глубоководных цератиоидных удильщиков Cryptopsaras couesi Gill». Природа . 256 (5512): 38–40. Бибкод : 1975Natur.256...38P . дои : 10.1038/256038a0 . S2CID 4226567 .
- ^ Суонн, Джереми Б. (30 июля 2020 г.). «Иммуногенетика полового паразитизма» . Наука . 369 (6511): 1608–1615. Бибкод : 2020Sci...369.1608S . дои : 10.1126/science.aaz9445 . hdl : 21.11116/0000-0006-CE67-F . ПМИД 32732279 . S2CID 220893482 .
- ^ Лаванши, Гийом; Швандер, Таня (2019). «Гибридогенез» . Современная биология . 29 (1): С9–Р11. Бибкод : 2019CBio...29...R9L . дои : 10.1016/j.cub.2018.11.046 . ПМИД 30620918 .
- ^ Мия, Масаки; Питч, Теодор В.; Орр, Джеймс В.; Арнольд, Рэйчел Дж; Сато, Такаши П; Шедлок, Эндрю М; Хо, Сюань-Цзин; Симадзаки, Мицуоми; Ябэ, Мамору; Нисида, Муцуми (1 января 2010 г.). «Эволюционная история удильщиков (Teleostei: Lophiiformes): митогеномная перспектива» . Эволюционная биология BMC . 10 (1): 58. Бибкод : 2010BMCEE..10...58M . дои : 10.1186/1471-2148-10-58 . ПМЦ 2836326 . ПМИД 20178642 .
- ^ Хаббс, CL; Хаббс, LC (1932). «Видимый партеногенез в природе у рыб гибридного происхождения». Наука . 76 (1983): 628–630. Бибкод : 1932Sci....76..628H . дои : 10.1126/science.76.1983.628 . ПМИД 17730035 .
- ^ Вриенгук, Р.К., Р.М. Доули, Си.Дж. Коул и Дж.П. Богарт. 1989. Список известных однополых позвоночных, стр. 19–23 в : «Эволюция и экология однополых позвоночных». Р. М. Доули и Дж. П. Богарт (ред.), Бюллетень 466, Музей штата Нью-Йорк, Олбани, Нью-Йорк.
- ^ Чепмен, Демиан Д.; Шивджи, Махмуд С.; Луи, Эд; Соммер, Джули; Флетчер, Хью; Продёль, Пауло А. (2007). «Непорочное зачатие у акулы-молота» . Письма по биологии . 3 (4): 425–427. дои : 10.1098/rsbl.2007.0189 . ПМК 2390672 . ПМИД 17519185 .
- ^ Робинсон, ДП; Баверсток, В.; Аль-Джару, А.; Хайленд, К.; Хазанедари, К.А. (2011). «Ежегодно повторяющийся партеногенез у акулы-зебры Stegostoma fasciatum ». Журнал биологии рыб . 79 (5): 1376–1382. Бибкод : 2011JFBio..79.1376R . дои : 10.1111/j.1095-8649.2011.03110.x . ПМИД 22026614 .
- ^ Чепмен, Д.Д.; Фирчау, Б.; Шивджи, М.С. (2008). «Партеногенез у крупнотелой акулы-реквиема, черноперой». Журнал биологии рыб . 73 (6): 1473–1477. дои : 10.1111/j.1095-8649.2008.02018.x .
- ^ Коя Ю., Мунехара Х. и Такано К. (2002). «Хранение спермы и подвижность в яичнике морского бычка Alcichthys alcicornis (Teleostei: Scorpaeniformes) с внутренней гаметической ассоциацией» . Журнал экспериментальной зоологии . 292 (2): 145–155. дои : 10.1002/jez.1150 . ПМИД 11754030 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ала-Хонкола О, Уддстрём А, Паули Б.Д., Линдстрём К (2009). «Сильная инбредная депрессия в брачном поведении самцов пецилидных рыб». Дж. Эвол. Биол . 22 (7): 1396–406. дои : 10.1111/j.1420-9101.2009.01765.x . ПМИД 19486236 .
- ^ Бикли Л.К., Браун А.Р., Хоскен Д.Д., Гамильтон П.Б., Ле Пейдж Дж., Полл Г.К., Оуэн С.Ф., Тайлер Ч.Р. (2013). «Интерактивное влияние инбридинга и эндокринных нарушений на воспроизводство модельной лабораторной рыбы» . Эволюционные приложения . 6 (2): 279–89. Бибкод : 2013EvApp...6..279B . дои : 10.1111/j.1752-4571.2012.00288.x . ПМЦ 3689353 . ПМИД 23798977 .
- ^ Галлардо Х.А., Нейра Р. (2005). «Экологическая зависимость инбредной депрессии у культивируемого кижуча (Oncorhynchus kisutch): агрессивность, доминирование и внутривидовая конкуренция» . Наследственность (Эдинб) . 95 (6): 449–56. дои : 10.1038/sj.hdy.6800741 . ПМИД 16189545 .
- ^ Jump up to: а б Фитцпатрик Дж.Л., Эванс Дж.П. (2014). «Избежание посткопулятивного инбридинга у гуппи» (PDF) . Дж. Эвол. Биол . 27 (12): 2585–94. дои : 10.1111/jeb.12545 . ПМИД 25387854 .
- ^ Чарльзворт Д., Уиллис Дж. Х. (2009). «Генетика инбредной депрессии». Нат. Преподобный Жене . 10 (11): 783–96. дои : 10.1038/nrg2664 . ПМИД 19834483 . S2CID 771357 .
- ^ Бернштейн Х., Хопф Ф.А., Мишод Р.Э. (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–70. дои : 10.1016/s0065-2660(08)60012-7 . ISBN 9780120176243 . ПМИД 3324702 .
- ^ http://www.carp.me.uk. Архивировано 21 марта 2011 г. на сайте Wayback Machine. Информация о нересте карпа.
- ^ Леонг, Пол (2004). «Советы по нересту петушков» . Архивировано из оригинала 2 марта 2009 года . Проверено 13 марта 2009 г. . Проверено 13 марта 2009 г.
- ^ Rainwate FL и Miller EJ (1967) «Ухаживание и репродуктивное поведение сиамской бойцовой рыбы Betta splendens Regan » Труды Академии наук Оклахомы , Университет штата Оклахома.
Дальнейшие ссылки
[ редактировать ]- Агарвал Н.К. (2008) Издательство APH по воспроизводству рыб . ISBN 9788131303573 .
- Бабин П.Дж., Серда Дж. и Лубценс Э. (редакторы) (2007) Ооцит рыбы: от фундаментальных исследований к биотехнологическим применениям Springer. ISBN 9781402062339 .
- Боун Кью и Мур Р. (2008) Биология рыб, глава 8: Размножение и истории жизни, стр. 217–255. Тейлор и Фрэнсис. ISBN 9781134186310 .
- Кабрита Э., Роблес В. и Пас Херраес П. (редакторы) (2008 г.) Методы репродуктивной аквакультуры: морские и пресноводные виды CRC Press. ISBN 9780849380549 .
- Коул, Кэтлин Сабина (редактор) (2010) Размножение и сексуальность морских рыб: закономерности и процессы University of California Press. ISBN 9780520264335 .
- Хоар В.С., Рэндалл Д.Д. и Дональдсон Э.М. (редакторы) (1983) Физиология рыб: Том 9: Размножение, Часть A: Эндокринные ткани и гормоны. Академическая пресса. ISBN 9780080585291 .
- Хоар В.С., Рэндалл Дж.Д. и Дональдсон Э.М. (редакторы) (1983) Физиология рыб: Том 9: Размножение, Часть B: Поведение и контроль фертильности. Академическая пресса. ISBN 9780080585307 .
- Якобсен Т., Фогарти М.Дж., Мегрей Б.А. и Мокснесс Э. (редакторы) (2009 г.) Репродуктивная биология рыб: значение для оценки и управления John Wiley & Sons. ISBN 9781444312126 .
- Меламед П. и Шервуд Н. (редакторы) (2005) Гормоны и их рецепторы в воспроизводстве рыб World Scientific. ISBN 9789812569189 .
- Поттс Г.В., Вуттон Р.Дж. и Вуттон Р.Дж. (редакторы) (1984) Размножение рыб: стратегии и тактика Academic Press. ISBN 9780125636605 .
- Роча М.Дж., Арукве А. и Капур Б.Г. (редакторы) (2008) Fish Reproduction CRC Press. ISBN 9781578083312 .
