Размножение млекопитающих

Большинство млекопитающих живородящие , рожают живых детенышей. [1] Однако пять видов однопроходных — утконосы и ехидны — откладывают яйца. У однопроходных система определения пола отличается от системы большинства других млекопитающих. [2] В частности, половые хромосомы утконоса больше похожи на хромосомы курицы, чем на хромосомы терийского млекопитающего. [3]
Молочные железы млекопитающих специализируются на выработке молока — жидкости, используемой новорожденными в качестве основного источника питания. Однопроходные рано ответвились от других млекопитающих и не имеют сосков, характерных для большинства млекопитающих, но у них есть молочные железы. Детеныши слизывают молоко с участка молочной железы на животе матери.
Живородящие млекопитающие относятся к подклассу Theria ; живущие сегодня относятся к инфраклассам Marsupialia и Placentalia . У сумчатого животного период беременности короткий , обычно короче его эстрального цикла , и он рождает недоразвитого ( альтрициального ) новорожденного, который затем подвергается дальнейшему развитию; у многих видов это происходит внутри мешочка, сумчатого матери , расположенного в передней части живота . Некоторые плацентарные особи, например морская свинка, рождают полностью развитых ( преждевременных ) детенышей, обычно после длительных периодов беременности, в то время как некоторые другие, например мыши, рождают недоразвитых детенышей.
Зрелость и репродуктивный возраст
[ редактировать ]Половая зрелость и, следовательно, самый ранний возраст, в котором млекопитающие могут размножаться, резко различаются у разных видов. Члены семейства грызунов Cricetidae могут достигать половой зрелости за 1–2 месяца, например норвежский лемминг ( Lemmus lemmus ) за 39 дней. Многим собакам ( семейство Canidae ) и быкам ( Bovidae ) требуется около года, чтобы достичь зрелости, тогда как приматам (включая человека ) и дельфинам ( Delphinidae ) требуется более 10 лет. Некоторым китам требуется еще больше времени, при этом наибольшая продолжительность зарегистрирована у гренландского кита ( Balaena mysticetus ), который достигает зрелости в возрасте всего около 23 лет. [4]
Репродуктивная система
[ редактировать ]Плацентарные млекопитающие
[ редактировать ]Самцы плацентарных млекопитающих
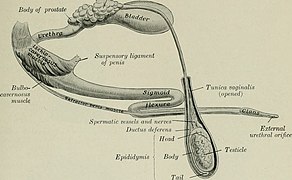
[ редактировать ]Мужская репродуктивная система млекопитающих состоит из двух основных отделов: полового члена и яичек , в последних из которых вырабатываются сперматозоиды, которые обычно хранятся в мошонке . У людей оба этих органа находятся за пределами брюшной полости , но у других животных они могут располагаться преимущественно внутри брюшной полости. Например, пенис собаки покрыт оболочкой полового члена, за исключением случаев спаривания. Расположение яичек вне брюшной полости лучше всего облегчает регулирование температуры сперматозоидов, которым для выживания необходимы определенные температуры. Внешнее расположение может также вызывать уменьшение вклада тепла в частоту спонтанных мутаций в мужской зародышевой ткани. [5] Сперматозоиды являются меньшими из двух гамет и, как правило, очень недолговечны, поэтому мужчинам приходится производить их непрерывно с момента половой зрелости до смерти. Произведенная сперма сохраняется в придатках яичка до эякуляции . Сперматозоиды подвижны и плавают, используя хвостообразные жгутики , чтобы продвинуться к яйцеклетке . Сперма следует температурным градиентам ( термотаксис ). [6] и химические градиенты ( хемотаксис ) для определения местоположения яйцеклетки.
Женские плаценты
[ редактировать ]Репродуктивная система самки млекопитающих состоит из трех основных отделов: влагалища и матки , которые действуют как вместилище для спермы, яичников самки , вырабатывающих яйцеклетки , и вульвы , состоящей из половых губ , преддверия и клитора . Влагалище, матка и яичники всегда находятся внутри, а вульва — снаружи. Влагалище прикрепляется к матке через шейку матки , а матка прикрепляется к яичникам через яйцеводы . Через определенные промежутки времени яичники выпускают яйцеклетку, которая через яйцевод попадает в матку.
Если во время этого транзита она встречается со спермой , яйцеклетка выбирает сперму, с которой она сливается; это называется оплодотворением . Оплодотворение обычно происходит в яйцеводах, но может произойти и в самой матке. Затем зигота имплантируется в стенку матки, где начинаются процессы эмбриогенеза и морфогенеза . Когда шейка матки развивается достаточно, чтобы выжить вне матки, шейка матки расширяется, и сокращения матки продвигают плод через родовые пути, то есть во влагалище.
Яйцеклетки, являющиеся женскими половыми клетками, намного крупнее сперматозоидов и обычно формируются в яичниках плода еще до его рождения. Они в основном фиксируются внутри яичника до момента перехода в матку и содержат питательные вещества для более поздней зиготы и эмбриона . Через регулярные промежутки времени в ответ на гормональные сигналы в процессе оогенеза созревает одна яйцеклетка, которая высвобождается и направляется по яйцеводам. Если яйцеклетка не оплодотворена, она выделяется во время менструации у людей и других человекообразных обезьян и реабсорбируется у других млекопитающих в цикле течки .
Беременность
[ редактировать ]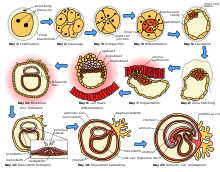
Беременность , называемая у человека беременностью , представляет собой период времени, в течение которого плод развивается, делясь посредством митоза внутри самки. В это время плод получает все необходимое питание и насыщенную кислородом кровь от самки, фильтруемую через плаценту плода , которая прикрепляется к брюшной полости через пуповину . Эта утечка питательных веществ может оказаться весьма обременительной для самки, которой приходится потреблять немного больше калорий . Кроме того, определенные витамины и другие питательные вещества требуются в больших количествах, чем обычно, что часто приводит к неправильным привычкам питания. Продолжительность беременности, называемая периодом беременности , сильно варьируется от вида к виду; у человека он составляет 40 недель, у жирафов 56–60 и у хомяков 16 дней .
Рождение
[ редактировать ]Как только плод достаточно развился, химические сигналы запускают процесс рождения. Это начинается с сокращения матки и расширения шейки матки. Затем плод опускается к шейке матки, где он выталкивается во влагалище и, в конечном итоге, из женщины. Новорожденный, которого у людей называют младенцем , обычно должен начать дышать самостоятельно вскоре после рождения. Вскоре после этого выходит и плацента.
Человеческие рождения
[ редактировать ]Человеческие младенцы уникальны в животном мире из-за большого размера головы по сравнению с телом. Это влияет на процесс родов у людей, поскольку из-за двуногой походки человека родовые пути становятся относительно узкими и искривленными посередине. В результате подавляющему большинству человеческих младенцев приходится вращаться внутри родовых путей, чтобы протиснуться через родовой канал и пройти через плоскости таза. Этот процесс известен как ротационное рождение, и хотя это не уникальный для человека процесс, люди уникальны тем, что почти все человеческие младенцы подвергаются этому процессу по необходимости. Основная гипотеза о том, почему происходит этот и другие процессы, из-за которых роды у людей становятся значительно более трудными, чем у других млекопитающих, известна как акушерская дилемма . [7]
однопроходные
[ редактировать ]Однопроходные , которых существует всего пять видов, все из Австралии и Новой Гвинеи , являются млекопитающими, откладывающими яйца . У них есть одно отверстие для выделения и размножения, называемое клоакой . Они удерживают яйца внутри в течение нескольких недель, обеспечивая их питательными веществами, а затем откладывают их и накрывают, как птицы . Как и сумчатые « джуи », однопроходные « мопсы » имеют личиночное и зародышевое состояние. [8] как и они, они не могут расширять свое туловище из-за наличия надлобковых костей, что вынуждает их производить неразвитое потомство.
Сумчатые
[ редактировать ]Репродуктивная система сумчатых заметно отличается от плацентарных млекопитающих. [9] [10] хотя, вероятно, это плезиоморфное состояние, обнаруженное у живородящих млекопитающих, включая неплацентарных плацентарных особей . [11] В процессе эмбрионального развития хориовителлиновая плацента у всех сумчатых формируется . У бандикутов образуется дополнительная хориоаллантоисная плацента , хотя в ней отсутствуют ворсинки хориона, встречающиеся в плацентах плаценты .

 мужского пола Домашний кот
мужского пола Домашний кот Домашняя кошка, самка
Домашняя кошка, самка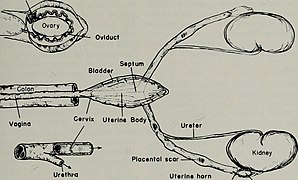 Самка енота
Самка енота Самец енота
Самец енота самец и самка Пятнистая гиена,
самец и самка Пятнистая гиена,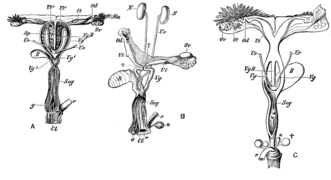 Самки сумчатых
Самки сумчатых -самец Кролик
-самец Кролик Самка кролика
Самка кролика










Гаметогенез
[ редактировать ]Животные, включая млекопитающих, производят гаметы (сперматозоиды и яйцеклетки) посредством мейоза в гонадах (яичках у самцов и яичниках у самок). Сперматозоиды производятся в процессе сперматогенеза , а яйцеклетки - в результате оогенеза . Эти процессы изложены в статье гаметогенез . Во время гаметогенеза у млекопитающих многие гены, кодирующие белки, участвующие в механизмах репарации ДНК, демонстрируют усиленную или специализированную экспрессию. [12] Эти механизмы включают мейотическую гомологичную рекомбинационную репарацию и репарацию ошибочного спаривания .
совокупление
[ редактировать ]











Сексуальное поведение можно разделить на поведенческие состояния, связанные с мотивацией вознаграждения (« желание »), получением вознаграждения, также известным как удовольствие («симпатия»), и насыщением («торможение»); [13] эти поведенческие состояния регулируются у млекопитающих посредством сексуального обучения, основанного на вознаграждении, колебаний различных нейрохимических веществ (например, дофамина – сексуального желания, также известного как «желание»; норадреналина – сексуального возбуждения ; окситоцина и меланокортинов – сексуального влечения ), а также половых гормональных циклов и на дальнейшее влияние оказывают половые феромоны и двигательные рефлексы (т. е. поведение лордоза ) у некоторых млекопитающих. [13] [14] Эти поведенческие состояния коррелируют с фазами цикла сексуальной реакции человека : мотивация – возбуждение; завершение — плато и оргазм; сытость − преломление. [13] Сексуальное обучение (форма ассоциативного обучения ) происходит, когда животное начинает ассоциировать особенности тела, личность, контекстуальные сигналы и другие стимулы с сексуальным удовольствием, вызванным половыми органами. [13] [14] Однажды сформировавшись, эти ассоциации, в свою очередь, влияют как на сексуальное желание, так и на сексуальное влечение.
У большинства самок млекопитающих акт совокупления контролируется несколькими врожденными нейробиологическими процессами, включая двигательный сексуальный рефлекс лордоза. [15] У самцов акт совокупления более сложен, поскольку необходимо определенное обучение, но врожденные процессы (ретроконтроль введения полового члена во влагалище , ритмичные движения таза, обнаружение женских феромонов) специфичны для совокупления. Эти врожденные процессы управляют гетеросексуальным совокуплением. [16] Поведение женского лордоза стало вторичным у гоминид и нефункциональным у людей. [17] Млекопитающие обычно совокупляются в дорсо-вентральной позе, хотя некоторые виды приматов совокупляются в вентро-вентральной позе. [18]
У большинства млекопитающих есть сошниково-носовой орган , который участвует в обнаружении феромонов, в том числе половых . [19] Несмотря на то, что у людей нет этого органа, взрослые люди, по-видимому, чувствительны к определенным феромонам млекопитающих, которые предполагаемые белки- рецепторы феромонов в обонятельном эпителии . способны обнаруживать [примечание 1] [19] Хотя половые феромоны явно играют роль в изменении сексуального поведения у некоторых млекопитающих, способность общего обнаружения феромонов и участие феромонов в сексуальном поведении человека еще не определена. [13]
Продолжительность совокупления значительно различается у разных видов млекопитающих. [23] и может коррелировать с массой тела, сохраняясь дольше у крупных млекопитающих, чем у мелких млекопитающих. [24] Продолжительность совокупления также может коррелировать с длиной бакулюма у млекопитающих. [25]
Самцы млекопитающих извергают сперму через половой член в репродуктивные пути самки во время совокупления. [26] Эякуляция обычно происходит только после одной интромиссии у людей, собак и копытных , но происходит после нескольких интромиссий у большинства видов млекопитающих. [27] [28]
Копуляция может вызвать овуляцию не происходит у видов млекопитающих, у которых овуляция спонтанно. [29]См. также
[ редактировать ]- Репродуктивная система млекопитающих
- Эволюция опустившихся семенников у млекопитающих
- Половое размножение § Млекопитающие
- Половой орган § Млекопитающие
- Репродукция человека
- Сексуальное поведение животных § Млекопитающие
- Беременность (млекопитающие)
- Репродуктивная система лошадей
- Хищники § Репродуктивная система
- Репродукция собак
- Дельфин § Размножение и сексуальность
- Лама § Размножение
- Воспроизводство домашних овец
Примечания
[ редактировать ]- ^ У людей и других животных следовые амино-ассоциированные рецепторы (TAAR), которые экспрессируются в обонятельном эпителии, действуют как обонятельные рецепторы , которые обнаруживают летучие аминные запахи , включая определенные феромоны ; [20] [21] эти TAAR предположительно функционируют как класс феромонных рецепторов, участвующих в обонятельном обнаружении социальных сигналов. [20] [21]
Обзор исследований с участием животных, не являющихся людьми, показал, что TAAR в обонятельном эпителии могут опосредовать привлекательные или отталкивающие поведенческие реакции на агонист . [20] В этом обзоре также отмечается, что поведенческая реакция, вызываемая TAAR, может различаться у разных видов. [20] Например, TAAR5 опосредует влечение к триметиламину у мышей и отвращение к триметиламину у крыс. [20] У людей hTAAR5 предположительно вызывает отвращение к триметиламину, который, как известно, действует как агонист hTAAR5 и обладает неприятным рыбным запахом, отталкивающим для человека; [20] [22] однако hTAAR5 — не единственный обонятельный рецептор, ответственный за обоняние триметиламина у человека. [20] [22] По состоянию на декабрь 2015 года [update] hTAAR5-опосредованная отвращение к триметиламину не изучалось в опубликованных исследованиях. [22]
Ссылки
[ редактировать ]- ^ Престон, Элизабет (13 февраля 2024 г.). «Любовь к себе важна, но мы, млекопитающие, застряли в сексе — некоторые самки птиц, рептилий и других животных могут выносить потомство самостоятельно. Но для таких млекопитающих, как мы, яйцеклетки и сперматозоиды нуждаются друг в друге» . Нью-Йорк Таймс . Архивировано из оригинала 13 февраля 2024 года . Проверено 16 февраля 2024 г.
- ^ Уоллис М.К., Уотерс П.Д., Делбридж М.Л., Кирби П.Дж., Паск А.Дж., Грюцнер Ф., Ренс В., Фергюсон-Смит М.А., Грейвс Дж.А. и др. (2007). «Определение пола у утконоса и ехидны: аутосомное расположение SOX3 подтверждает отсутствие SRY у однопроходных». Хромосомные исследования . 15 (8): 949–959. дои : 10.1007/s10577-007-1185-3 . ПМИД 18185981 . S2CID 812974 .
- ^ Маршалл Грейвс, Дженнифер А. (2008). «Странные геномы животных и эволюция пола и половых хромосом позвоночных» (PDF) . Ежегодный обзор генетики . 42 : 568–586. дои : 10.1146/annurev.genet.42.110807.091714 . ПМИД 18983263 . Архивировано из оригинала (PDF) 4 сентября 2012 г.
- ^ Пасифичи, Микела; Сантини, Лука; Марко, Морено Ди; Байсеро, Даниэле; Франкуччи, Лусилла; Маразини, Габриэле Гроттоло; Висконти, Пьеро; Рондинини, Карло (13 ноября 2013 г.). «Продолжительность поколений млекопитающих» . Охрана природы . 5 :89–94. doi : 10.3897/natureconservation.5.5734 . ISSN 1314-3301 . Архивировано из оригинала 26 января 2021 г. Проверено 03 марта 2021 г.
- ^ Бальц, Р.Х.; Бингэм, премьер-министр; Дрейк, JW (1976). «Тепловой мутагенез бактериофага Т4: путь перехода» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (4): 1269–73. Бибкод : 1976PNAS...73.1269B . дои : 10.1073/pnas.73.4.1269 . ПМК 430244 . ПМИД 4797 .
- ^ Бахат, Анат; Тур-Каспа, Илан; Гакамский, Анна; Джохалас, Лаура К.; Брейтбарт, Хаим; Айзенбах, Майкл (2003). «Термотаксис сперматозоидов млекопитающих: потенциальный механизм навигации в женских половых путях». Природная медицина . 9 (2): 149–50. дои : 10.1038/nm0203-149 . hdl : 11336/66658 . ПМИД 12563318 . S2CID 36538049 .
- ^ Треватан, Венда (5 марта 2015 г.). «Анатомия таза приматов и последствия для рождения» . Философские труды Королевского общества B: Биологические науки . 370 : 20140065.doi : (1663 ) 10.1098/rstb.2014.0065 . ISSN 0962-8436 . ПМК 4305166 . ПМИД 25602069 .
- ^ Мангер, Пол Р.; Холл, Лесли С.; Петтигрю, Джон Д. (1998). «Развитие внешних признаков утконоса (Ornithorhynchus anatinus)» . Философские труды Лондонского королевского общества. Серия Б: Биологические науки . 353 (1372): 1115–1125. дои : 10.1098/rstb.1998.0270 . ПМК 1692310 . ПМИД 9720109 .
- ^ Австралийское общество млекопитающих (декабрь 1978 г.). Австралийское общество млекопитающих . Австралийское общество млекопитающих.
- ^ Открытия кафедры биологии Университета штата Айова о размножении сумчатых, Анна Кинг, 2001. Веб-страница. Архивировано 5 сентября 2012 г. в Wayback Machine (примечание показывает код, расширение HTML опущено).
- ^ Джалломбардо, Андрес, 2009 г. Новые млекопитающие мелового периода из Монголии и ранняя диверсификация Eutheria Ph.D. диссертация, Колумбийский университет, 2009 г., 402 страницы; AAT 3373736 (аннотация) Происхождение плацентарных млекопитающих, Cimolestidae, Zalambdalestidae
- ^ Баарендс В.М., ван дер Лаан Р., Гротегоед Дж.А. (2001). «Механизмы репарации ДНК и гаметогенез» . Размножение . 121 (1): 31–9. дои : 10.1530/reprod/121.1.31 . hdl : 1765/9599 . ПМИД 11226027 .
- ^ Jump up to: а б с д и Георгиадис-младший, Крингельбах М.Л., Пфаус Дж.Г. (сентябрь 2012 г.). «Секс ради удовольствия: синтез нейробиологии человека и животных». Обзоры природы. Урология . 9 (9): 486–98. дои : 10.1038/nrurol.2012.151 . ПМИД 22926422 . S2CID 13813765 .
Цикл сексуального удовольствия соответствует базовой структуре циклов удовольствия, связанных с другими вознаграждениями (такими как еда), и поэтому может также быть выражен в терминах мотивация-удовлетворение-насыщение или желание-приязнь-торможение (рис. 1; вставка 2). 6,11,1 ... Подобно другим формам обучения, сексуальное поведение развивается с течением времени, когда люди учатся связывать такие стимулы, как особенности тела, личность и контекстуальные сигналы, с сексуальным удовольствием, вызываемым гениталиями.7 Подростковый возраст, возможно, является наиболее важным. фаза полового развития... Распространенное мнение гласит, что люди также реагируют на некоторые дистальные сексуальные стимулы (грудь, феромоны) безусловным образом, но это трудно оценить эмпирически (Вставка 1)... Сексуальное желание у обеих крыс а у людей взаимодействие между половыми гормонами и внешними стимулами становится сексуальным стимулом благодаря ассоциации с сексуальным вознаграждением, вызванным половыми органами; Таким образом, приятная генитальная стимуляция является основным фактором сексуального обучения...
• Безусловные сексуальные стимулы (то есть те, для получения удовольствия от которых не требуется обучения) включают тактильную стимуляцию проксимальных половых органов у людей и дистальные стимулы, такие как феромоны, запахи и некоторые слуховые вокализации у крыс.7,16
• Сексуальное торможение задействует схожие механизмы мозга у крыс и людей.
• Крысы демонстрируют аналогичную с людьми модель активации мозга в ответ на сигналы, связанные с сексуальным вознаграждением.
• Кортикальные, лимбические, гипоталамические и мозжечковые области активируются стимулами, связанными с полом, как у людей, так и у крыс. - ^ Jump up to: а б Шульц В. (2015). «Нейронное вознаграждение и сигналы решения: от теорий к данным» . Физиологические обзоры . 95 (3): 853–951. doi : 10.1152/physrev.00023.2014 . ПМЦ 4491543 . ПМИД 26109341 .
Сексуальное поведение обусловлено гормональным дисбалансом, по крайней мере у мужчин, но также во многом основано на удовольствии. Приобретение и следование этим основным пищевым и брачным вознаграждениям является основной причиной, по которой система вознаграждения мозга вообще развилась. Обратите внимание, что «основное» вознаграждение не относится к различию между безусловным и условным вознаграждением; действительно, большинство первичных наград усваиваются и, таким образом, обусловлены (еда — это основные награды, которые обычно усваиваются). ... Удовольствие — это не только одна из трех основных функций вознаграждения, но и определение вознаграждения. Поскольку гомеостаз объясняет функции лишь ограниченного числа вознаграждений, преобладающей причиной того, почему определенные стимулы, объекты, события, ситуации и действия приносят вознаграждение, может быть удовольствие. Это относится прежде всего к сексу (который занимался бы нелепой гимнастикой репродуктивной деятельности, если бы не удовольствие) и к первичным гомеостатическим вознаграждениям в виде еды и жидкости, а также к деньгам, вкусу, красоте, социальным контактам и нематериальным благам. , внутренне установленные и внутренние вознаграждения. ... Желание делает поведение целенаправленным и направляет его к опознаваемым целям. Таким образом, желание — это эмоция, которая помогает активно направлять поведение на известное вознаграждение, тогда как удовольствие — это пассивный опыт, возникающий в результате полученного или ожидаемого вознаграждения. Желание имеет множество отношений с удовольствием; оно может быть приятным само по себе (я чувствую приятное желание) и может приводить к удовольствию (я желаю получить приятный объект). Таким образом, удовольствие и желание имеют разные характеристики, но тесно переплетены. Они представляют собой самые важные положительные эмоции, вызываемые наградами. Они отдают приоритет нашей сознательной обработке информации и, таким образом, представляют собой важные компоненты поведенческого контроля. Эти эмоции также называются симпатией (для удовольствия) и желанием (для желания) в исследованиях зависимости (471) и решительно поддерживают функции вознаграждения, генерирующие обучение и подход. ... Некоторые стимулы и события, доставляющие удовольствие людям, могут даже не вызывать удовольствия у животных, а действовать вместо этого посредством врожденных механизмов. Мы просто не знаем. Тем не менее, изобретение удовольствия и желания в ходе эволюции имело огромное преимущество, поскольку позволило большому количеству стимулов, объектов, событий, ситуаций и занятий быть привлекательными. Этот механизм важно поддерживает основные функции вознаграждения при получении необходимых веществ и партнеров для спаривания.
- ^ Пфафф Дональд В., Шварц-Гиблин Сьюзен, Маккарти Маргарет М., Коу Ли-Минг: Клеточные и молекулярные механизмы женского репродуктивного поведения, в Кнобил Эрнест, Нил Джимми Д.: Физиология воспроизводства, Raven Press, 2-е издание, 1994 г.
- ^ Мейзель Роберт Л., Сакс Бенджамин Д.: Физиология мужского сексуального поведения. В Кнобил Эрнест, Нил Джимми Д. Физиология репродукции, Raven Press, 2-е издание, 1994 г.
- ^ Диксон А. Ф. Сексуальность приматов: сравнительные исследования полуобезьян, обезьян, человекообразных обезьян и людей . Издательство Оксфордского университета, 2-е издание, 2012 г.
- ^ Диксон, Алан Ф. (15 мая 2009 г.). Половой отбор и происхождение систем спаривания человека . ОУП Оксфорд. ISBN 978-0-19-156973-9 .
- ^ Jump up to: а б Ней М., Ниимура Ю., Нозава М. (декабрь 2008 г.). «Эволюция репертуара генов хемосенсорных рецепторов животных: роль случая и необходимости». Обзоры природы. Генетика . 9 (12): 951–63. дои : 10.1038/nrg2480 . ПМИД 19002141 . S2CID 11135361 .
Гены ОР преимущественно экспрессируются в сенсорных нейронах главного обонятельного эпителия (МОЭ) полости носа. Млекопитающие обнаруживают многие типы химических веществ в воздухе и некоторые в воде как запахи, тогда как рыбы распознают водорастворимые молекулы, такие как аминокислоты, желчные кислоты, половые стероиды и простагландины. Известно, что некоторые гены OR млекопитающих экспрессируются в других тканях, включая семенники, язык, мозг и плаценту17. Однако функциональное значение такой «эктопической экспрессии» OR-генов окончательно не известно. TAARs также выражены в MOE. Эти рецепторы были впервые идентифицированы как рецепторы головного мозга для следовых аминов, совокупности аминов, которые в низких концентрациях присутствуют в центральной нервной системе18. Первоначально предполагалось, что TAARs участвуют в психических расстройствах19, но теперь известно, что они функционируют как второй класс обонятельных рецепторов10. Некоторые мышиные TAAR распознают летучие амины, присутствующие в моче, и, по-видимому, функция TAAR заключается в обнаружении лигандов, связанных с социальными сигналами10. ... У большинства млекопитающих имеется дополнительный орган обоняния, называемый сошниково-носовым органом (VnO). ... Раньше считалось, что VnO является специализированным органом для обнаружения феромонов, но теперь известно, что VnO и MOE имеют некоторые перекрывающиеся функции22. ... Однако по крайней мере один из пяти генов V1R экспрессируется в обонятельной слизистой оболочке человека72. Более того, недавнее исследование предполагает, что эти пять генов могут активировать OR-подобный путь передачи сигнала в гетерологичной системе экспрессии73. Поэтому возможно, что продукты этих генов действуют как феромоны или обонятельные рецепторы. Взрослые люди не имеют VnO, но, по-видимому, чувствительны к феромонам74. Еще одно интересное наблюдение заключается в том, что у кур нет функциональных или нефункциональных генов V1R и V2R или VnO75, хотя птицы используют феромоны для выбора партнера и другого поведения76. Возможно, что некоторые OR-гены в MOE способны обнаруживать феромоны, как у людей74,77.
- ^ Jump up to: а б с д и ж г Либерлес СД (октябрь 2015 г.). «Отслеживание рецепторов, связанных с аминами: лиганды, нейронные цепи и поведение» . Курс. Мнение. Нейробиол . 34 : 1–7. дои : 10.1016/j.conb.2015.01.001 . ПМК 4508243 . ПМИД 25616211 .
Роль другого рецептора подтверждается TAAR5-независимыми триметиламиновыми аносмиями у людей [32]. ... Некоторые TAAR обнаруживают летучие и неприятные амины, но обонятельная система способна игнорировать лигандные или функциональные ограничения на эволюцию TAAR. Отдельные TAAR мутировали, чтобы распознавать новые лиганды, при этом почти вся клада костистых рыб потеряла канонический мотив распознавания аминов. Более того, хотя некоторые TAAR обнаруживают неприятные запахи, поведение, опосредованное TAAR, может различаться у разных видов. ... Способность определенных TAAR опосредовать поведение отвращения и влечения предоставляет прекрасную возможность для механистического раскрытия кодирования валентности запаха.
Рисунок 2: Таблица лигандов, моделей экспрессии и видоспецифичных поведенческих реакций для каждого TAAR. - ^ Jump up to: а б «Рецептор следовых аминов: Введение» . Международный союз фундаментальной и клинической фармакологии. Архивировано из оригинала 23 февраля 2014 года . Проверено 15 февраля 2014 г.
Важно отметить, что три лиганда, активирующие мышиные Taars, являются естественными компонентами мышиной мочи, основным источником социальных сигналов у грызунов. Мышиный Taar4 распознает β-фенилэтиламин, соединение, повышение которого в моче коррелирует с усилением стресса и стрессовых реакций как у грызунов, так и у людей. И Taar3, и Taar5 мыши обнаруживают соединения (изоамиламин и триметиламин соответственно), которые обогащены мочой самцов и самок мышей. Сообщается, что изоамиламин в моче самцов действует как феромон, ускоряя наступление полового созревания у самок мышей [34]. Авторы предполагают, что семейство Таар обладает хемосенсорной функцией, отличной от рецепторов запахов и связанной с обнаружением социальных сигналов. ... Эволюционная картина семейства генов TAAR характеризуется специфичной для линии филогенетической кластеризацией [26,30,35]. Эти характеристики очень похожи на те, которые наблюдаются в семействах генов обонятельных GPCR и вомероназальных (V1R, V2R) GPCR.
- ^ Jump up to: а б с Валлрабенштейн И., Сингер М., Пантен Дж., Хатт Х., Гиссельманн Г. (2015). «Тимберол® ингибирует TAAR5-опосредованные реакции на триметиламин и влияет на обонятельный порог у людей» . ПЛОС ОДИН . 10 (12): e0144704. Бибкод : 2015PLoSO..1044704W . дои : 10.1371/journal.pone.0144704 . ПМЦ 4684214 . ПМИД 26684881 .
В то время как мыши производят количество ТМА в моче в зависимости от пола и их привлекает ТМА, этот запах отталкивает крыс и отталкивает людей [19], что указывает на то, что должны существовать видоспецифичные функции. ... Более того, гомозиготный нокаут мышиного TAAR5 отменил поведение влечения к ТМА [19]. Таким образом, сделан вывод, что самого TAAR5 достаточно, чтобы опосредовать поведенческий ответ, по крайней мере, у мышей. ... Еще предстоит изучить, вызывает ли активация TAAR5 с помощью ТМА определенные поведенческие реакции, такие как поведение избегания у людей.
- ^ Нагиб, Марк (19 апреля 2020 г.). Достижения в изучении поведения . Академическая пресса. ISBN 978-0-12-820726-0 .
- ^ Столлманн, Роберт Р. и А.Х. Харкорт. « Размер имеет значение: (отрицательная) аллометрия продолжительности совокупления у млекопитающих. Архивировано 20 апреля 2022 г. в Wayback Machine ». Биологический журнал Линнеевского общества 87.2 (2006): 185–193. дои : 10.1111/j.1095-8312.2006.00566.x
- ^ DIXSON33, Алан, Н. ЙХОЛ Т. Дженна и Мэтт Андерсон. «Положительная связь между длиной бакулюма и продолжительной паттернами интромиссии у млекопитающих». 动物学报 50.4 (2004): 490-503.
- ^ Ломбарди, Джулиан (6 декабря 2012 г.). Сравнительное размножение позвоночных . Springer Science & Business Media. ISBN 978-1-4615-4937-6 .
- ^ Диксон, Алан Ф. (26 января 2012 г.). Сексуальность приматов: сравнительные исследования полуобезьян, обезьян, человекообразных обезьян и людей . ОУП Оксфорд. ISBN 978-0-19-954464-6 .
- ^ Энциклопедия поведенческой нейронауки . Эльзевир. 03.06.2010. ISBN 978-0-08-045396-5 .
- ^ Йохле, Вольфганг (1973). «Овуляция, вызванная коитусом» . Контрацепция . 7 (6): 523–564. дои : 10.1016/0010-7824(73)90023-1 . ISSN 0010-7824 .