Хемотаксис
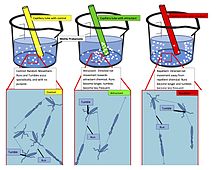
Хемотаксис (от хемо- + таксис ) — это движение организма или сущности в ответ на химический раздражитель. [1] Соматические клетки , бактерии и другие одноклеточные или многоклеточные организмы направляют свои движения в соответствии с определенными химическими веществами в их среде обитания. Бактериям важно находить пищу (например, глюкозу ), плавая к самой высокой концентрации пищевых молекул, или спасаться от ядов (например, фенола ). У многоклеточных организмов хемотаксис имеет решающее значение для раннего развития (например, движения сперматозоидов к яйцеклетке во время оплодотворения ) и развития (например, миграции нейронов или лимфоцитов ), а также для нормального функционирования и здоровья (например, миграция лейкоцитов во время травмы или инфекция). [2] Кроме того, было признано, что механизмы, обеспечивающие хемотаксис у животных, могут быть нарушены во время рака метастазирования . [3] Аберрантный хемотаксис лейкоцитов и лимфоцитов также способствует воспалительным заболеваниям, таким как атеросклероз, астма и артрит. [4] [5] [6] [7] Субклеточные компоненты, такие как участок полярности, генерируемый спаривающимися дрожжами, также могут проявлять хемотаксическое поведение. [8]
Положительный хемотаксис возникает, если движение направлено в сторону более высокой концентрации рассматриваемого химического вещества; отрицательный хемотаксис, если движение происходит в противоположном направлении. Химически вызванный кинезис (произвольно направленный или ненаправленный) можно назвать хемокинезом .
История хемотаксиса исследований
еще на заре развития микроскопии Хотя миграция клеток была обнаружена Левенгуком , в лекции по хемотаксису в Калифорнийском технологическом институте утверждается, что «эрудитическое описание хемотаксиса было впервые сделано Т. В. Энгельманном (1881) и В. Ф. Пфеффером (1884) у бактерий, и HS Jennings (1906) у инфузорий . [9] Лауреат Нобелевской премии И. Мечников также внес вклад в изучение этой области в 1882—1886 гг., исследуя процесс как начальный этап фагоцитоза . [10] Значение хемотаксиса в биологии и клинической патологии было широко признано в 1930-х годах, и к этому времени были разработаны наиболее фундаментальные определения, лежащие в основе этого явления. [ кем? ] Важнейшие аспекты контроля качества исследований хемотаксиса были описаны Х. Харрисом в 1950-х годах. [11] В 1960-х и 1970-х годах революция в современной клеточной биологии и биохимии предоставила ряд новых методов, которые стали доступны для исследования мигрирующих клеток-ответчиков и субклеточных фракций, ответственных за хемотаксическую активность. [12] Доступность этой технологии привела к открытию C5a, основного хемотаксического фактора, участвующего в остром воспалении. Новаторские работы Дж. Адлера модернизировали капиллярный анализ Пфеффера и явились важным поворотным моментом в понимании всего процесса внутриклеточной передачи сигналов бактерий. [13] [14]
Бактериальный хемотаксис характеристика общая -

Некоторые бактерии , такие как E. coli , имеют несколько жгутиков на клетку (обычно 4–10). Они могут вращаться двумя способами:
- Вращение против часовой стрелки объединяет жгутики в один вращающийся пучок, заставляя бактерию плавать по прямой линии; и
- Вращение по часовой стрелке разбивает пучок жгутиков на части, так что каждый жгутик направлен в разном направлении, в результате чего бактерия падает на месте. [15]
Направления вращения указаны для наблюдателя вне клетки, смотрящего вниз по жгутикам в сторону клетки. [16]
Поведение [ править ]
Общее движение бактерии является результатом чередования фаз падения и плавания, называемых бегущим движением . [17] В результате траектория движения бактерии, плавающей в однородной среде, будет представлять собой случайное блуждание с относительно прямым плаванием, прерываемым случайными падениями, которые переориентируют бактерию. [18] Бактерии, такие как E. coli, не могут выбирать направление своего плавания и не могут плавать по прямой более нескольких секунд из-за вращательной диффузии ; другими словами, бактерии «забывают» направление, в котором они движутся. Постоянно оценивая свой курс и корректируя свое движение в неправильном направлении, бактерии могут направить свое случайное движение в сторону благоприятных мест. [19]
При наличии химического градиента бактерии будут хемотаксировать или направлять свое общее движение в зависимости от градиента. Если бактерия почувствует, что движется в правильном направлении (к аттрактанту/от репеллента), она будет продолжать плыть по прямой в течение более длительного времени, прежде чем упасть; однако, если он движется в неправильном направлении, он упадет раньше. Бактерии, такие как E. coli, используют временное зондирование, чтобы решить, улучшается ли их ситуация или нет, и таким образом находят место с самой высокой концентрацией аттрактанта, обнаруживая даже небольшие различия в концентрации. [20]
Это предвзятое случайное блуждание является результатом простого выбора между двумя методами случайного движения; а именно акробатика и прямое плавание. [21] Спиральная природа отдельной жгутиковой нити имеет решающее значение для возникновения этого движения. Белковая структура, составляющая жгутиковую нить, флагеллин , консервативна среди всех жгутиковых бактерий. [22] Позвоночные животные, похоже, воспользовались этим фактом, обладая иммунным рецептором ( TLR5 ), предназначенным для распознавания этого консервативного белка. [23]
Как и во многих случаях в биологии, существуют бактерии, которые не следуют этому правилу. Многие бактерии, такие как вибрионы , моножгутиковые и имеют один жгутик на одном полюсе клетки. Их метод хемотаксиса различен. У других есть единственный жгутик, который находится внутри клеточной стенки. Эти бактерии передвигаются, вращая всю клетку, имеющую форму штопора. [24] [ нужна страница ]
Преобразование сигнала [ править ]

Химические градиенты воспринимаются множеством трансмембранных рецепторов , называемых метил-принимающими белками хемотаксиса (MCP), которые различаются по молекулам, которые они обнаруживают. [25] Известно, что во всем бактериальном царстве кодируются тысячи рецепторов MCP. [26] Эти рецепторы могут связывать аттрактанты или репелленты прямо или косвенно посредством взаимодействия с белками периплазматического пространства . [27] Сигналы от этих рецепторов передаются через плазматическую мембрану в цитозоль , где белки Che . активируются [28] Белки Che изменяют частоту переворота и изменяют рецепторы. [28]
Регуляция жгутика [ править ]
Белки CheW и CheA связываются с рецептором. Отсутствие активации рецептора приводит к CheA по аутофосфорилированию гистидинкиназы одному высококонсервативному остатку гистидина. [29] [ нужен лучший источник ] CheA, в свою очередь, переносит фосфорильные группы на консервативные остатки аспартата в регуляторах ответа CheB и CheY; CheA представляет собой гистидинкиназу и не переносит активно фосфорильную группу, скорее, регулятор ответа CheB забирает фосфорильную группу у CheA. [ нужна ссылка ] Этот механизм передачи сигнала называется двухкомпонентной системой и является распространенной формой передачи сигнала у бактерий. [ нужна ссылка ] CheY вызывает переворачивание путем взаимодействия с белком-переключателем жгутика FliM, вызывая изменение вращения жгутика с против часовой стрелки на вращение по часовой стрелке. Изменение состояния вращения одного жгутика может разрушить весь пучок жгутиков и вызвать его переворачивание. [ нужна ссылка ]
рецепторов Регуляция
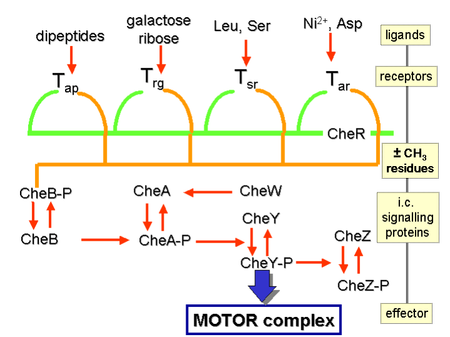
CheB при активации CheA действует как метилэстераза , удаляя метильные группы из остатков глутамата на цитозольной стороне рецептора; он действует антагонистически с CheR, метилтрансферазой , которая добавляет метильные остатки к тем же остаткам глутамата. [30] Если уровень аттрактанта останется высоким, то уровень фосфорилирования CheA (а, следовательно, CheY и CheB) останется низким, клетка будет плавно плавать, а уровень метилирования MCP увеличится (поскольку CheB-P отсутствует для деметилирования). [30] MCP больше не реагируют на аттрактант, когда они полностью метилированы; следовательно, даже если уровень аттрактанта может оставаться высоким, уровень CheA-P (и CheB-P) увеличивается, и клетка начинает разрушаться. [30] MCP могут быть деметилированы CheB-P, и когда это произойдет, рецепторы снова смогут реагировать на аттрактанты. [30] В отношении репеллентов ситуация противоположная: полностью метилированные МЦП лучше всего реагируют на репелленты, а наименее метилированные МЦП хуже всего реагируют на репелленты. [ нужна ссылка ] Эта регуляция позволяет бактерии «запоминать» химические концентрации из недавнего прошлого, на несколько секунд, и сравнивать их с теми, которые она испытывает в настоящее время, таким образом «знать», движется ли она вверх или вниз по градиенту. [31] Что бактерии имеют химические градиенты, то в повышении абсолютной величины чувствительности на том или ином фоне участвуют другие механизмы. Хорошо известными примерами являются сверхчувствительная реакция мотора на сигнал CheY-P и кластеризация хеморецепторов. [32] [33]
Хемоаттрактанты и хемопелленты [ править ]
Хемоаттрактанты и хеморепелленты — неорганические или органические вещества, обладающие индуктором хемотаксиса в подвижных клетках. Эти хемотаксические лиганды создают градиенты химических концентраций, к которым организмы, прокариотические и эукариотические, движутся навстречу или от них соответственно. [34]
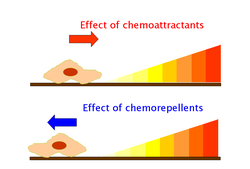
Эффекты хемоаттрактантов вызываются через хеморецепторы, такие как метил-принимающие белки хемотаксиса (MCP). [35] MCP в E.coli включают Tar, Tsr, Trg и Tap. [36] Хемоаттрактанты Trg включают рибозу и галактозу с фенолом в качестве хемопеллента. Tap и Tsr распознают дипептиды и серин как хемоаттрактанты соответственно. [36]
Хемоаттрактанты или хемопелленты связывают MCP во внеклеточном домене; внутриклеточный сигнальный домен передает изменения концентрации этих хемотаксических лигандов нижестоящим белкам, таким как CheA, который затем передает этот сигнал жгутиковым моторам через фосфорилированный CheY (CheY-P). [35] Затем CheY-P может контролировать вращение жгутиков, влияя на направление подвижности клеток. [35]
Для E.coli , S. meliloti и R. spheroides связывание хемоаттрактантов с MCP ингибирует активность CheA и, следовательно, CheY-P, что приводит к плавному прохождению, но для B. substilis активность CheA увеличивается. [35] События метилирования в E.coli приводят к тому, что MCP имеют более низкое сродство к хемоаттрактантам, что вызывает повышение активности CheA и CheY-P, что приводит к падению. [35] Таким образом, клетки способны адаптироваться к немедленной концентрации хемоаттрактанта и обнаруживать дальнейшие изменения для модуляции подвижности клеток. [35]
Хемоаттрактанты эукариот хорошо характеризуют иммунные клетки. Формильные пептиды , такие как fMLF , привлекают лейкоциты , такие как нейтрофилы и макрофаги , вызывая движение к местам инфекции. [37] Неацилированные метионинильные пептиды не действуют как хемоаттрактанты на нейтрофилы и макрофаги. [37] Лейкоциты также движутся к хемоаттрактантам C5a, компоненту комплемента и патоген -специфическим лигандам бактерий. [37]
Механизмы действия хемопеллентов менее известны, чем механизмы хемоаттрактантов. Хотя хемопелленты вызывают у организмов реакцию избегания, Tetrahymena thermophila адаптируется к хеморепелленту, пептиду Нетрин-1 , в течение 10 минут после воздействия; однако воздействие хемопеллентов, таких как GTP , PACAP-38 и ноцицептин , не приводит к такой адаптации. [38] ГТФ и АТФ являются хеморепеллентами в микромолярных концентрациях как для Tetrahymena , так и для Paramecium . Эти организмы избегают этих молекул, вызывая реакции избегания, чтобы переориентироваться от градиента. [39]
Эукариотический хемотаксис [ править ]
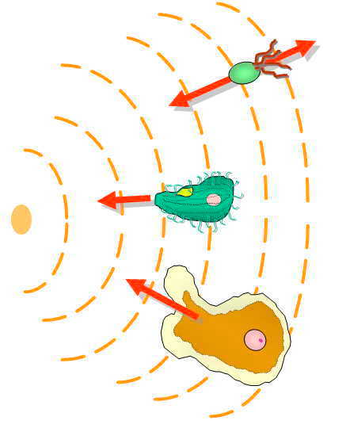
Механизм хемотаксиса эукариотических клеток совершенно отличается от механизма хемотаксиса бактерий E. coli ; однако обнаружение химических градиентов по-прежнему остается важным шагом в этом процессе. [40] [ нужен лучший источник ] Из-за своего небольшого размера и других биофизических ограничений E. coli не может напрямую обнаруживать градиент концентрации. [41] Вместо этого они используют определение временного градиента, при котором они перемещаются на большие расстояния, в несколько раз превышающие их собственную ширину, и измеряют скорость изменения воспринимаемой химической концентрации. [42] [43]
Эукариотические клетки намного крупнее прокариот и имеют рецепторы, равномерно расположенные по всей клеточной мембране . [42] Хемотаксис эукариот включает пространственное обнаружение градиента концентрации путем сравнения асимметричной активации этих рецепторов на разных концах клетки. [42] Активация этих рецепторов приводит к миграции к хемоаттрактантам или от хеморепеллентов. [42] У спаривающихся дрожжей, которые неподвижны, участки полярных белков на клеточной коре могут перемещаться хемотаксическим образом вверх по градиентам феромонов. [44] [8]
Также было показано, что как прокариотические, так и эукариотические клетки способны к хемотаксической памяти. [43] [45] У прокариот этот механизм включает метилирование рецепторов, называемых метил-принимающими белками хемотаксиса (MCP). [43] Это приводит к их десенсибилизации и позволяет прокариотам «помнить» и адаптироваться к химическому градиенту. [43] Напротив, хемотаксическая память у эукариот может быть объяснена моделью локального возбуждения и глобального торможения (LEGI). [45] [46] LEGI включает баланс между быстрым возбуждением и отсроченным торможением, который контролирует нижестоящую передачу сигналов, такую как активация Ras и выработка PIP3 . [47]
Уровни рецепторов, внутриклеточные сигнальные пути и эффекторные механизмы представляют собой разнообразные компоненты эукариотического типа. В эукариотических одноклеточных клетках основными эффекторами являются амебоидное движение и ресничка или эукариотический жгутик (например, Amoeba или Tetrahymena ). [48] [49] Некоторые эукариотические клетки высшего происхождения позвоночных , например иммунные клетки, также перемещаются туда, где им необходимо быть. Помимо иммунокомпетентных клеток ( гранулоцитов , моноцитов , лимфоцитов ), большая группа клеток, ранее считавшихся фиксированными в тканях, также подвижна в особых физиологических (например, тучные клетки , фибробласты , эндотелиальные клетки ) или патологических состояниях (например, метастазы ). . [50] Хемотаксис имеет большое значение на ранних стадиях эмбриогенеза , поскольку развитие зародышевых листков направляется градиентами сигнальных молекул. [51] [52]
Подвижность [ править ]
В отличие от подвижности при бактериальном хемотаксисе, механизм физического перемещения эукариотических клеток неясен. По-видимому, существуют механизмы, с помощью которых внешний хемотаксический градиент воспринимается и превращается во внутриклеточный градиент PIP3 к градиенту и активации сигнального пути, кульминацией которого является полимеризация актиновых филаментов , что приводит . Растущий дистальный конец актиновых филаментов развивает связи с внутренней поверхностью плазматической мембраны через различные наборы пептидов и приводит к образованию передних псевдоподий и задних уроподий . Реснички эукариотических клеток также могут вызывать хемотаксис; в данном случае это в основном Ca 2+ -зависимая индукция системы микротрубочек базального тельца и ритма 9+2 микротрубочек внутри ресничек. Организованное биение сотен ресничек синхронизируется подмембранной системой, построенной между базальными тельцами.Детали сигнальных путей до сих пор не совсем ясны.
хемотаксисом связанные с

Хемотаксис относится к направленной миграции клеток в ответ на химические градиенты; Существует несколько вариантов химической миграции, перечисленных ниже.
- Хемокинез означает увеличение подвижности клеток в ответ на химические вещества в окружающей среде. В отличие от хемотаксиса, миграция, стимулируемая хемокинезом, лишена направленности и вместо этого усиливает поведение сканирования окружающей среды. [53]
- При гаптотаксисе градиент . хемоаттрактанта выражен или связан на поверхности, в отличие от классической модели хемотаксиса, в которой градиент развивается в растворимой жидкости [54] Наиболее распространенной биологически активной гаптотактической поверхностью является внеклеточный матрикс (ECM); наличие связанных лигандов отвечает за индукцию трансэндотелиальной миграции и ангиогенеза .
- Некротаксис представляет собой особый тип хемотаксиса, когда молекулы хемоаттрактанта высвобождаются из некротических или апоптотических клеток. В зависимости от химического характера выделяемых веществ некротаксис может накапливать или отталкивать клетки, что подчеркивает патофизиологическую значимость этого явления.
Рецепторы [ править ]
В общем, эукариотические клетки ощущают присутствие хемотаксических стимулов посредством использования 7-трансмембранных (или серпентиновых) гетеротримерных рецепторов, связанных с G-белком , - класса, представляющего значительную часть генома . [55] Некоторые члены этого суперсемейства генов используются в зрении ( родопсины ), а также в обонянии (обонянии). [56] [57] Основные классы рецепторов хемотаксиса запускаются:
- Формильные пептиды – формилпептидные рецепторы (FPR),
- Хемокины - хемокиновые рецепторы (CCR или CXCR) и
- Лейкотриены – лейкотриеновые рецепторы (БЛТ). [58]
Однако индукция широкого набора мембранных рецепторов (например, циклических нуклеотидов , аминокислот , инсулина , вазоактивных пептидов) также вызывает миграцию клетки. [59]
Хемотаксический отбор [ править ]

В то время как некоторые рецепторы хемотаксиса экспрессируются на поверхности мембраны с долговременными характеристиками, поскольку они детерминированы генетически, другие имеют краткосрочную динамику, поскольку они собираются ad hoc в присутствии лиганда. [60] Разнообразные особенности рецепторов и лигандов хемотаксиса позволяют отбирать клетки-ответчики на хемотаксис с помощью простого анализа хемотаксиса. С помощью хемотаксического отбора мы можем определить, действует ли еще не охарактеризованная молекула через долгосрочный или кратковременный рецепторный путь. [61] Термин «хемотаксический отбор» также используется для обозначения метода, который разделяет эукариотические или прокариотические клетки в соответствии с их хемотаксической чувствительностью к селекторным лигандам. [62] [ нужен неосновной источник ] [ нужен неосновной источник ]
Хемотаксические лиганды [ править ]


Число молекул, способных вызывать хемотаксические ответы, относительно велико, и можно выделить первичные и вторичные хемотаксические молекулы. [ нужна ссылка ] Основные группы первичных лигандов следующие:
- Формильные пептиды представляют собой ди-, три-, тетрапептиды бактериального происхождения, формилированные на N-конце пептида. [ нужна ссылка ] [63] Они выделяются из бактерий in vivo или после разложения клетки. Типичным представителем этой группы является N-формилметиониллейцилфенилаланин (сокращенно fMLF или fMLP). [ нужна ссылка ] Бактериальный fMLF является ключевым компонентом воспаления и оказывает характерное хемоаттрактантное действие на нейтрофильные гранулоциты и моноциты. [ нужна ссылка ] Лиганды и рецепторы хемотаксических факторов, связанные с формильными пептидами, обобщены в соответствующей статье « Рецепторы формилпептидов» .
- Комплемент 3а ( С3а ) и комплемент 5а ( С5а ) являются промежуточными продуктами каскада комплемента. [ нужна ссылка ] Их синтез сопряжен с тремя альтернативными путями (классическим, лектинзависимым и альтернативным) активации комплемента ферментом конвертазой. [ нужна ссылка ] Основными клетками-мишенями этих производных являются также нейтрофильные гранулоциты и моноциты. [ нужна ссылка ]
- Хемокины принадлежат к особому классу цитокинов ; их группы (хемокины C, CC, CXC, CX 3 C) не только представляют собой структурно родственные молекулы с особым расположением дисульфидных мостиков, но и их специфичность к клеткам-мишеням разнообразна. [ нужна ссылка ] Хемокины CC действуют на моноциты (например, RANTES ), а хемокины CXC специфичны для нейтрофильных гранулоцитов (например, IL-8 ). [ нужна ссылка ] Исследования трехмерной структуры хемокинов показали, что характерный состав бета-листов и альфа-спирали обеспечивает экспрессию последовательностей, необходимых для взаимодействия с хемокиновыми рецепторами. [ нужна ссылка ] Образование димеров и их повышенная биологическая активность были продемонстрированы кристаллографией нескольких хемокинов, например IL-8. [ нужна ссылка ]
- Метаболиты полиненасыщенных жирных кислот
- Лейкотриены представляют собой эйкозаноидные липидные медиаторы, образующиеся в результате метаболизма арахидоновой кислоты с помощью ALOX5 (также называемого 5-липоксигеназой). Их наиболее известным членом с активностью хемотаксического фактора является лейкотриен B4 , который вызывает адгезию, хемотаксис и агрегацию лейкоцитов. Хемоаттрактантное действие LTB4 индуцируется посредством любого из двух рецепторов, связанных с G-белком , BLT1 и BLT2 , которые высоко экспрессируются в клетках, участвующих в воспалении и аллергии . [64]
- Семейство эйкозаноидов 5-гидроксикозатетраеновой кислоты представляет собой метаболиты арахидоновой кислоты, также образуемые ALOX5. Три члена семейства образуются естественным путем и обладают заметной хемотаксической активностью. Они перечислены в порядке убывания эффективности: 5-оксо-эйкозатетраеновая кислота , 5-оксо-15-гидрокси-эйкозатетраеновая кислота и 5-гидроксиэйкозатетраеновая кислота . Это семейство агонистов стимулирует хемотаксические реакции в эозинофилах , нейтрофилах и моноцитах человека путем связывания с оксоэйкозаноидным рецептором 1 , который, как и рецепторы лейкотриена B4, является рецептором, связанным с G-белком. [64] Помимо кожи, нейтрофилы являются первой линией защиты организма от бактериальных инфекций. Покинув близлежащие кровеносные сосуды, эти клетки распознают химические вещества, вырабатываемые бактериями в порезе или царапине, и мигрируют «в сторону запаха».
- 5-гидроксиэйкозатриеновая кислота и 5-оксоэйкозатриеновая кислота являются метаболитами Мидовой кислоты (5 Z ,8 Z ,11 Z -эйкозатриреноидная кислота); они стимулируют хемотаксис лейкоцитов через оксоэйкозаноидный рецептор 1 [65] при этом 5-оксоэйкозатриеновая кислота столь же эффективна, как и ее аналог, производный арахидоновой кислоты, 5-оксо-эйкозатетраеновая кислота, в стимуляции хемотаксиса эозинофилов и нейтрофилов крови человека . [64]
- 12-Гидроксикозатетраеновая кислота представляет собой эйкозаноидный метаболит арахидоновой кислоты, образуемый ALOX12 , который стимулирует хемотаксис лейкоцитов через лейкотриеновый рецептор B4, BLT2. [64]
- Простагландин D2 представляет собой эйкозаноидный метаболит арахидононовой кислоты, вырабатываемый циклооксигеназой 1 или циклооксигеназой 2 , который стимулирует хемотаксис через рецептор простагландина DP2 . Он вызывает хемотаксические реакции в эозинофилах, базофилах и Т-хелперных клетках подтипа Th2. [66] [ нужен неосновной источник ] [ нужен неосновной источник ]
- 12-Гидроксигептадекатриеновая кислота представляет собой неэйкозаноидный метаболит арахидоновой кислоты, образуемый циклооксигеназой 1 или циклооксигеназой 2, который стимулирует хемотаксис лейкоцитов через лейкотриеновый B4-рецептор, BLT2. [67] [ нужен неосновной источник ] [ нужен неосновной источник ]
- 15-оксо-эйкозатетраеновая кислота представляет собой эйкозаноидный метаболит арахидоновой кислоты, полученный моим ALOX15 ; он обладает слабой хемотаксической активностью в отношении моноцитов человека (см. 15-Гидроксикозатетраеновая кислота#15-оксо-ЭТЕ ). [68] [ нужен неосновной источник ] [ нужен неосновной источник ] Рецептор или другой механизм, с помощью которого этот метаболит стимулирует хемотаксис, не выяснен.
диапазона хемотаксического Подбор

Хемотаксические реакции, вызываемые взаимодействиями лиганд - рецептор, варьируются в зависимости от концентрации лиганда. Исследования семейств лигандов (например, аминокислот или олигопептидов ) показывают, что хемоаттрактантная активность проявляется в широком диапазоне, тогда как хемопеллентная активность имеет узкие диапазоны. [69]
Клиническое значение
Измененный миграционный потенциал клеток имеет сравнительно большое значение в развитии ряда клинических симптомов и синдромов.Изменение хемотаксической активности внеклеточных (например, Escherichia coli ) или внутриклеточных (например, Listeria monocytogenes ) патогенов само по себе представляет собой важную клиническую мишень. Модификация эндогенной хемотаксической способности этих микроорганизмов фармацевтическими агентами может снизить или ингибировать частоту инфекций или распространение инфекционных заболеваний.Помимо инфекций, существуют и другие заболевания, при которых нарушение хемотаксиса является основным этиологическим фактором, например синдром Чедиака-Хигаси , при котором гигантские внутриклеточные везикулы ингибируют нормальную миграцию клеток.
| Тип заболевания | Хемотаксис увеличен | Хемотаксис снизился |
|---|---|---|
| Инфекции | Воспаления | СПИД , Бруцеллез |
| Хемотаксис приводит к заболеванию | — | Синдром Чедиака-Хигаси , синдром Картагенера |
| Нарушается хемотаксис | Атеросклероз , артрит , пародонтит , псориаз , реперфузионное повреждение , метастатические опухоли | Рассеянный склероз , болезнь Ходжкина , мужское бесплодие |
| Отравление | Асбест , бензпирен | Hg и Cr Соли , озон |
Математические модели [ править ]
Разработано несколько математических моделей хемотаксиса в зависимости от типа
- Миграция (например, основные различия бактериального плавания, перемещение одноклеточных эукариот с ресничками / жгутиками и амебоидная миграция)
- Физико-химические характеристики химических веществ (например, диффузия ), действующих как лиганды.
- Биологические характеристики лигандов (аттрактантные, нейтральные и репеллентные молекулы)
- Системы анализа, применяемые для оценки хемотаксиса (см. время инкубации, развитие и стабильность градиентов концентрации)
- Другие воздействия окружающей среды, оказывающие прямое или косвенное влияние на миграцию (освещение, температура, магнитные поля и т. д.)
Хотя взаимодействие перечисленных выше факторов делает поведение решений математических моделей хемотаксиса достаточно сложным, основной феномен движения, обусловленного хемотаксисом, можно описать прямолинейно. Действительно, обозначим через пространственно неоднородная концентрация хемоаттрактанта и как его градиент. Затем хемотаксический клеточный поток (также называемый током) генерируемый хемотаксисом, связан с вышеуказанным градиентом по закону: [70]




где - пространственная плотность клеток и это так называемый «коэффициент хемотаксиса» - часто является не постоянной, а убывающей функцией хемоаттрактанта. Для некоторого количества который подвержен полному потоку и срок генерации/уничтожения , можно сформулировать уравнение неразрывности :





где это расхождение . Это общее уравнение применимо как к плотности клеток, так и к хемоаттрактанту. Следовательно, если включить диффузионный поток в общий поток, взаимодействия между этими величинами будут определяться набором связанных производных реакция-диффузия, дифференциальных уравнений в частных описывающих изменение и : [70]

![{\displaystyle {\begin{aligned}{\partial C \over {\partial t}}&=f(C)+\nabla \cdot \left[D_{C}\nabla CC\chi (\varphi)\nabla \varphi \right]\\{\partial \varphi \over {\partial t}}&=g(\varphi ,C)+\nabla \cdot (D_ {\varphi }\nabla \varphi )\end{aligned} }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/924b3d46d7dc2a27228a7d98cf43d035a4622c41)
где описывает рост плотности клеток, представляет собой термин кинетики/источника для хемоаттрактанта, а коэффициенты диффузии для плотности клеток и хемоаттрактанта соответственно равны и .




Пространственная экология почвенных микроорганизмов является функцией их хемотаксической чувствительности к субстрату и другим организмам. [71] [ нужен неосновной источник ] [ нужен неосновной источник ] Было доказано, что хемотаксическое поведение бактерий приводит к нетривиальным структурам популяций даже в отсутствие гетерогенности окружающей среды. Наличие структурных неоднородностей порового масштаба оказывает дополнительное влияние на возникающие бактериальные структуры.
хемотаксиса Измерение
Доступен широкий спектр методов для оценки хемотаксической активности клеток или хемоаттрактантного и хеморепеллентного характера лигандов.Основные требования к измерениям следующие:
- Градиенты концентрации могут развиваться относительно быстро и длительно сохраняться в системе.
- Различают хемотаксическую и хемокинетическую активность.
- Миграция клеток свободна вперед и назад по оси градиента концентрации.
- Обнаруженные ответы являются результатом активной миграции клеток.
Несмотря на то, что идеальный анализ хемотаксиса до сих пор не доступен, существует несколько протоколов и оборудования, которые хорошо соответствуют условиям, описанным выше. Наиболее часто используемые приведены в таблице ниже:
| Тип анализа | Анализы на агаровых пластинках | Двухкамерные анализы | Другие |
|---|---|---|---|
| Примеры |
|
|
|
Искусственные хемотаксические системы
химические роботы , которые используют искусственный хемотаксис для автономной навигации. Были разработаны [72] [73] Приложения включают адресную доставку лекарств в организм. [74] Совсем недавно молекулы ферментов также продемонстрировали положительное хемотаксическое поведение в градиенте своих субстратов. [75] Термодинамически выгодное связывание ферментов со специфическими субстратами считается причиной ферментативного хемотаксиса. [76] Кроме того, ферменты в каскадах также демонстрируют хемотаксическую агрегацию, управляемую субстратом. [77]
Помимо активных ферментов, нереагирующие молекулы также проявляют хемотаксическое поведение. Это было продемонстрировано с помощью молекул красителя, которые движутся направленно в градиентах раствора полимера за счет благоприятных гидрофобных взаимодействий. [78]
См. также [ править ]
Ссылки [ править ]
- ^ Чисхолм, Хью , изд. (1911). . Британская энциклопедия . Том. 6 (11-е изд.). Издательство Кембриджского университета. п. 77.
- ^ де Оливейра С., Розовский Э.Э., Хуттенлохер А. (май 2016 г.). «Миграция нейтрофилов при инфекции и заживлении ран: движение вперед в обратном направлении» . Обзоры природы. Иммунология . 16 (6): 378–91. дои : 10.1038/nri.2016.49 . ПМК 5367630 . ПМИД 27231052 .
- ^ Стелтен CH, Parent CA, Montell DJ (май 2018 г.). «Подвижность клеток при инвазии и метастазировании рака: данные простых модельных организмов» . Обзоры природы. Рак . 18 (5): 296–312. дои : 10.1038/nrc.2018.15 . ПМК 6790333 . ПМИД 29546880 .
- ^ Ли Дж., Лей К. (январь 2015 г.). «Миграция лимфоцитов в атеросклеротическую бляшку» . Атеросклероз, тромбоз и сосудистая биология . 35 (1): 40–9. дои : 10.1161/ATVBAHA.114.303227 . ПМЦ 4429868 . ПМИД 25301842 .
- ^ Гельфанд РЭБ (октябрь 2017). «Важность путей лейкотриенов B4-BLT1 и LTB4-BLT2 при астме» . Семинары по иммунологии . 33 : 44–51. дои : 10.1016/j.smim.2017.08.005 . ПМЦ 5679233 . ПМИД 29042028 .
- ^ Планагума А, Доменек Т, Пон М, Калама Е, Гарсиа-Гонсалес В, Лопес Р, Аули М, Лопес М, Фонкерна С, Рамос И, де Альба Х, Нуэда А, Пратс Н, Сегарра В, Миральпейкс М, Ленер МД (октябрь 2015 г.). «Комбинированная терапия анти-CXC-рецепторами 1 и 2 является многообещающим противовоспалительным лечением респираторных заболеваний за счет уменьшения миграции и активации нейтрофилов». Легочная фармакология и терапия . 34 : 37–45. дои : 10.1016/j.pupt.2015.08.002 . ПМИД 26271598 .
- ^ Рана АК, Ли Й, Данг К, Ян Ф (декабрь 2018 г.). «Моноциты при ревматоидном артрите: циркулирующие предшественники макрофагов и остеокластов и их роль гетерогенности и пластичности в патогенезе РА». Международная иммунофармакология . 65 : 348–359. дои : 10.1016/j.intimp.2018.10.016 . ПМИД 30366278 . S2CID 53116963 .
- ↑ Перейти обратно: Перейти обратно: а б Гхош, Дебрадж; Джейкобс, Кэтрин; Рамирес, Самуэль; Элстон, Тимоти; Лью, Дэниел (1 июня 2021 г.). «Хемотаксическое перемещение участка полярности позволяет дрожжевым клеткам находить себе пару» . Труды Национальной академии наук . 118 (22): e2025445118. Бибкод : 2021PNAS..11825445G . дои : 10.1073/pnas.2025445118 . ISSN 0027-8424 . ПМК 8179161 . ПМИД 34050026 .
- ^ Лекция по хемотаксису. Загружено в 2007 г. Доступно по адресу: http://www.rpgroup.caltech.edu/courses/aph161/2007/lectures/ChemotaxisLecture.pdf. Архивировано 19 июня 2010 г. в Wayback Machine (последняя проверка: 15.04.17).
- ^ Эли Мечников». Британская энциклопедия. Британская энциклопедия, Inc.
- ^ Модели Келлера-Сегеля для хемотаксиса. 2012. доступно по адресу: http://www.isn.ucsd.edu/courses/Beng221/problems/2012/BENG221_Project%20-%20Roberts%20Chung%20Yu%20Li.pdf . Архивировано 29 августа 2017 г. в Wayback Machine (последняя проверка). к апрелю 2017 г.)
- ^ Снайдерман Р., Гевурц Х., Мергенхаген С.Е. (август 1968 г.). «Взаимодействие системы комплемента с эндотоксичными липополисахаридами. Генерация фактора хемотаксиса полиморфно-ядерных лейкоцитов» . Журнал экспериментальной медицины . 128 (2): 259–75. дои : 10.1084/jem.128.2.259 . ПМК 2138524 . ПМИД 4873021 .
- ^ Адлер Дж., Цо В.В. (июнь 1974 г.). « Принятие решений у бактерий: хемотаксический ответ Escherichia coli на противоречивые стимулы». Наука . 184 (4143): 1292–4. Бибкод : 1974Sci...184.1292A . дои : 10.1126/science.184.4143.1292 . ПМИД 4598187 . S2CID 7221477 .
- ^ Берг, Ховард (2004). Берг, Ховард С. (ред.). Кишечная палочка в движении . Биологическая и медицинская физика, Биомедицинская инженерия. Спрингер. п. 15 , 19–29. дои : 10.1007/b97370 . ISBN 0-387-00888-8 . S2CID 35733036 .
- ^ Юань Дж., Фарнер К.А., Тернер Л., Берг Х.К. (июль 2010 г.). «Асимметрия вращения бактериального жгутикового мотора по и против часовой стрелки» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (29): 12846–9. Бибкод : 2010PNAS..10712846Y . дои : 10.1073/pnas.1007333107 . ПМЦ 2919929 . ПМИД 20615986 .
- ^ «Бактериальный хемотаксис» (PDF) . Архивировано (PDF) из оригинала 6 мая 2017 года.
- ^ Берг Х.К., Браун Д.А. (октябрь 1972 г.). «Хемотаксис Escherichia coli, анализируемый с помощью трехмерного отслеживания». Природа . 239 (5374): 500–504. Бибкод : 1972Natur.239..500B . дои : 10.1038/239500a0 . ПМИД 4563019 . S2CID 1909173 .
- ^ Сурджик В., Вингрин Н.С. (апрель 2012 г.). «Реакция на химические градиенты: бактериальный хемотаксис» . Современное мнение в области клеточной биологии . 24 (2): 262–268. дои : 10.1016/j.ceb.2011.11.008 . ПМК 3320702 . ПМИД 22169400 .
- ^ Берг, Ховард К. (1993). Случайные блуждания в биологии (Расширенная, переработанная ред.). Принстон, Нью-Джерси: Princeton Univ. Нажимать. стр. 83–94. ISBN 978-0-691-00064-0 .
- ^ Сурджик В., Вингрин Н. (апрель 2012 г.). «Реакция на химические градиенты: бактериальный хемотаксис» . Современное мнение в области клеточной биологии . 24 (2): 262–8. дои : 10.1016/j.ceb.2011.11.008 . ПМК 3320702 . ПМИД 22169400 .
- ^ Макнаб Р.М., Кошланд, Д.Э. (сентябрь 1972 г.). «Механизм восприятия градиента в бактериальном хемотаксисе» . Труды Национальной академии наук Соединенных Штатов Америки . 69 (9): 2509–12. Бибкод : 1972PNAS...69.2509M . дои : 10.1073/pnas.69.9.2509 . ПМК 426976 . ПМИД 4560688 .
- ^ Неделькович, Марко; Састре, Диего; Сундберг, Эрик (14 июля 2021 г.). «Бактериальная жгутиковая нить: супрамолекулярная многофункциональная наноструктура» . Международный журнал молекулярных наук . 22 (14): 7521. doi : 10.3390/ijms22147521 . ПМК 8306008 . ПМИД 34299141 .
- ^ Чжун, Маохуа; Ян, Хуэйминь; Ли, Яомин (октябрь 2017 г.). «Флагеллин: уникальная молекулярная структура, связанная с микробами, и многогранный иммуномодулятор» . Клеточная и молекулярная иммунология . 14 (10): 862–864. дои : 10.1038/cmi.2017.78 . ISSN 2042-0226 . ПМЦ 5649114 . ПМИД 28845044 .
- ^ Берг ХК (2003). кишечной палочки Перемещение . Нью-Йорк, штат Нью-Йорк: Спрингер. ISBN 978-0-387-00888-2 . [ нужна страница ]
- ^ Уодхамс, Джордж Х.; Армитидж, Джудит П. (декабрь 2004 г.). «Понимание всего этого: бактериальный хемотаксис». Nature Reviews Молекулярно-клеточная биология . 5 (12): 1024–1037. дои : 10.1038/nrm1524 . ПМИД 15573139 . S2CID 205493118 .
- ^ Гальперин, Михаил (июнь 2005 г.). «Перепись мембраносвязанных и внутриклеточных белков, передающих сигнал, у бактерий: бактериальный IQ, экстраверты и интроверты» . БМК Микробиология . 5:35 . дои : 10.1186/1471-2180-5-35 . ПМЦ 1183210 . ПМИД 15955239 .
- ^ Дженнаро Аулетта (2011). Когнитивная биология: работа с информацией от бактерий до разума . США: Издательство Оксфордского университета. п. 266. ИСБН 978-0-19-960848-5 .
- ↑ Перейти обратно: Перейти обратно: а б Фальке, Джозеф Дж.; Басс, Рэндал Б.; Батлер, Скотт Л.; Червиц, Стивен А.; Дэниэлсон, Марк А. (1997). «ДВУХКОМПОНЕНТНЫЙ СИГНАЛЬНЫЙ ПУТЬ БАКТЕРИАЛЬНОГО ХЕМОТАКСИСА: Молекулярный взгляд на передачу сигнала рецепторами, киназами и адаптационными ферментами» . Ежегодный обзор клеточной биологии и биологии развития . 13 : 457–512. дои : 10.1146/annurev.cellbio.13.1.457 . ISSN 1081-0706 . ПМК 2899694 . ПМИД 9442881 .
- ^ ToxCafe (2 июня 2011 г.). «Хемотаксис» . Архивировано из оригинала 11 июля 2015 года . Проверено 23 марта 2017 г. - через YouTube.
- ↑ Перейти обратно: Перейти обратно: а б с д Уодхамс, Джордж Х.; Армитидж, Джудит П. (декабрь 2004 г.). «Понимание всего этого: бактериальный хемотаксис». Nature Reviews Молекулярно-клеточная биология . 5 (12): 1024–1037. дои : 10.1038/nrm1524 . ISSN 1471-0080 . ПМИД 15573139 . S2CID 205493118 .
- ^ Шу Чиен; Питер С.И. Чен; Ю. К. Фунг (2008). Вводный текст в биоинженерию (Продвинутая серия по биомеханике - Том 4) . Сингапур: World Scientific Publishing Co. Pte. ООО с. 418. ИСБН 9789812707932 .
- ^ Клюзель П., Сюретт М., Лейблер С. (март 2000 г.). «Сверхчувствительный бактериальный мотор, выявленный путем мониторинга сигнальных белков в отдельных клетках». Наука . 287 (5458): 1652–5. Бибкод : 2000Sci...287.1652C . дои : 10.1126/science.287.5458.1652 . ПМИД 10698740 . S2CID 5334523 .
- ^ Сурджик В. (декабрь 2004 г.). «Кластеризация рецепторов и обработка сигналов в хемотаксисе E. coli». Тенденции в микробиологии . 12 (12): 569–76. CiteSeerX 10.1.1.318.4824 . дои : 10.1016/j.tim.2004.10.003 . ПМИД 15539117 .
- ^ Сюй Ф, Бирман Р, Хили Ф, Нгуен Х (2016). «Многомасштабная модель хемотаксиса Escherichia coli от внутриклеточного сигнального пути до подвижности и поглощения питательных веществ в градиенте питательных веществ и изотропной жидкой среде» . Компьютеры и математика с приложениями . 71 (11): 2466–2478. дои : 10.1016/j.camwa.2015.12.019 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Сзурмант Х., Ордал Г.В. (июнь 2004 г.). «Разнообразие механизмов хемотаксиса бактерий и архей» . Обзоры микробиологии и молекулярной биологии . 68 (2): 301–19. дои : 10.1128/MMBR.68.2.301-319.2004 . ПМК 419924 . ПМИД 15187186 .
- ↑ Перейти обратно: Перейти обратно: а б Ямамото К., Макнаб Р.М., Имаэ Ю. (январь 1990 г.). «Функции репеллентного ответа хеморецепторов Trg и Tap Escherichia coli» . Журнал бактериологии . 172 (1): 383–8. дои : 10.1128/jb.172.1.383-388.1990 . ПМК 208443 . ПМИД 2403544 .
- ↑ Перейти обратно: Перейти обратно: а б с Шиффманн Э., Коркоран Б.А., Валь С.М. (март 1975 г.). «N-формилметиониловые пептиды как хемоаттрактанты лейкоцитов» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (3): 1059–62. Бибкод : 1975PNAS...72.1059S . дои : 10.1073/pnas.72.3.1059 . ПМК 432465 . ПМИД 1093163 .
- ^ Курувилла Х, Шмидт Б, Сонг С, Бхаджан М, Мерикал М, Элли С, Гриффин С, Йодер Д, Хейн Дж, Коль Д, Паффенбергер С, Петрофф Д, Новичок Е, Гуд К, Хестон Дж, Хуртубис А (2016) . «Пептид нетрин-1 является хеморепеллентом у Tetrahymena thermophila» . Международный журнал пептидов . 2016 : 7142868. doi : 10.1155/2016/7142868 . ПМЦ 4830718 . ПМИД 27123011 .
- ^ Хеннесси ТМ (июнь 2005 г.). «Реакция инфузорий Tetrahymena и Paramecium на внешние АТФ и ГТФ» . Пуринергическая сигнализация . 1 (2): 101–10. дои : 10.1007/s11302-005-6213-1 . ПМК 2096533 . ПМИД 18404496 .
- ^ Кохидай Л. (2016), «Хемотаксис как выражение коммуникации тетрахимен», в Вицани Г., Новацки М. (ред.), Биокоммуникация инфузорий , стр. 65–82, doi : 10.1007/978-3-319-32211- 7_5 , ISBN 978-3-319-32211-7
- ^ Берг, ХК; Перселл, Э.М. (ноябрь 1977 г.). «Физика хеморецепции» . Биофизический журнал . 20 (2): 193–219. Бибкод : 1977BpJ....20..193B . дои : 10.1016/s0006-3495(77)85544-6 . ISSN 0006-3495 . ПМЦ 1473391 . ПМИД 911982 .
- ↑ Перейти обратно: Перейти обратно: а б с д Левин, Герберт; Раппель, Воутер-Ян (1 февраля 2013 г.). «Физика хемотаксиса эукариот» . Физика сегодня . 66 (2): 24–30. Бибкод : 2013ФТ....66б..24Л . дои : 10.1063/PT.3.1884 . ПМЦ 3867297 . ПМИД 24363460 .
- ↑ Перейти обратно: Перейти обратно: а б с д Владимиров Н., Сурджик В. (ноябрь 2009 г.). «Хемотаксис: как бактерии используют память». Биологическая химия . 390 (11): 1097–104. дои : 10.1515/BC.2009.130 . ПМИД 19747082 . S2CID 207440927 .
- ^ Гхош, Дебрадж; Лью, Дэниел (1 мая 2020 г.). «Механистическое понимание движения участков полярности, управляемого актином, у дрожжей» . Молекулярная биология клетки . 31 (10): 1085–1102. дои : 10.1091/mbc.e20-01-0040 . ISSN 1059-1524 . ПМЦ 7346724 . ПМИД 32186970 .
- ↑ Перейти обратно: Перейти обратно: а б Скоге М., Юэ Х., Эрикстад М., Бэ А., Левин Х., Гройсман А., Лумис В.Ф., Раппель В.Дж. (октябрь 2014 г.). «Клеточная память в хемотаксисе эукариот» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (40): 14448–53. Бибкод : 2014PNAS..11114448S . дои : 10.1073/pnas.1412197111 . ПМК 4210025 . ПМИД 25249632 .
- ^ Кучер Б., Девреотес П., Иглесиас П.А. (февраль 2004 г.). «Локальное возбуждение, глобальный механизм торможения для определения градиента: интерактивный апплет». СТКЭ науки . 2004 (219): табл.3. дои : 10.1126/stke.2192004pl3 . ПМИД 14872096 . S2CID 4660870 .
- ^ Сюн Ю, Хуан Ч., Иглесиас П.А., Девреотес П.Н. (октябрь 2010 г.). «Клетки перемещаются с помощью возбудимой сети с локальным возбуждением и глобальным торможением» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (40): 17079–86. Бибкод : 2010PNAS..10717079X . дои : 10.1073/pnas.1011271107 . ПМЦ 2951443 . ПМИД 20864631 .
- ^ Багорда А., родитель, Калифорния (август 2008 г.). «Краткий обзор хемотаксиса эукариот» . Журнал клеточной науки . 121 (Часть 16): 2621–4. CiteSeerX 10.1.1.515.32 . дои : 10.1242/jcs.018077 . ПМЦ 7213762 . ПМИД 18685153 .
- ^ Кохидай Л (1999). «Хемотаксис: правильный физиологический ответ для оценки филогении сигнальных молекул». Акта Биологика Хунгарика . 50 (4): 375–94. дои : 10.1007/BF03543060 . ПМИД 10735174 . S2CID 248703226 .
- ^ Кедрин Д., ван Ринен Дж., Эрнандес Л., Конделис Дж., Сегал Дж.Э. (сентябрь 2007 г.). «Подвижность клеток и регуляция цитоскелета при инвазии и метастазировании». Журнал биологии молочной железы и неоплазии . 12 (2–3): 143–52. дои : 10.1007/s10911-007-9046-4 . ПМИД 17557195 . S2CID 31704677 .
- ^ Солница-Крезель Л., Сепич Д.С. (2012). «Гаструляция: образование и формирование зародышевых листков». Ежегодный обзор клеточной биологии и биологии развития . 28 : 687–717. doi : 10.1146/annurev-cellbio-092910-154043 . ПМИД 22804578 . S2CID 11331182 .
- ^ Шеллард А., мэр Р. (июль 2016 г.). «Хемотаксис во время миграции нервного гребня». Семинары по клеточной биологии и биологии развития . 55 : 111–8. дои : 10.1016/j.semcdb.2016.01.031 . ПМИД 26820523 .
- ^ Беккер Э.Л. (октябрь 1977 г.). «Стимулированное передвижение нейтрофилов: хемокинез и хемотаксис». Архивы патологии и лабораторной медицины . 101 (10): 509–13. ПМИД 199132 .
- ^ Картер С.Б. (январь 1967 г.). «Гаптотаксис и механизм подвижности клеток». Природа . 213 (5073): 256–60. Бибкод : 1967Natur.213..256C . дои : 10.1038/213256a0 . ПМИД 6030602 . S2CID 4212997 .
- ^ Ким Дж. Я., Хаастерт П. В., Девреотес П. Н. (апрель 1996 г.). «Социальные чувства: сигнальные пути рецепторов, связанных с G-белком, у Dictyostelium discoideum» . Химия и биология . 3 (4): 239–43. дои : 10.1016/s1074-5521(96)90103-9 . ПМИД 8807851 .
- ^ Монтелл, Крейг (ноябрь 1999 г.). «Визуальная трансдукция у дрозофилы». Ежегодный обзор клеточной биологии и биологии развития . 15 (1): 231–268. дои : 10.1146/annurev.cellbio.15.1.231 . ПМИД 10611962 . S2CID 14193715 .
- ^ Антунес Дж., Симоэш де Соуза FM (2016). «Передача сигналов обонятельных рецепторов». G-белковые рецепторы – передача сигналов, транспортировка и регуляция . Методы клеточной биологии. Том. 132. стр. 127–45. дои : 10.1016/bs.mcb.2015.11.003 . ISBN 9780128035955 . ПМИД 26928542 .
- ^ Томас, Моника А.; Клейст, Эндрю Б.; Волкман, Брайан Ф. (2018). «Расшифровка хемотаксического сигнала» . Журнал биологии лейкоцитов . 104 (2): 359–374. дои : 10.1002/JLB.1MR0218-044 . ПМК 6099250 . ПМИД 29873835 .
- ^ ван Хаастерт П.Дж., Де Вит Р.Дж., Конейн Т.М. (август 1982 г.). «Антагонисты хемоаттрактантов выявляют отдельные рецепторы цАМФ, фолиевой кислоты и птерина у Dictyostelium» (PDF) . Экспериментальные исследования клеток . 140 (2): 453–6. дои : 10.1016/0014-4827(82)90139-2 . ПМИД 7117406 . S2CID 27784085 .
- ^ Вицани, Гюнтер; Новацкий, Мариуш (2016). Биокоммуникация инфузорий . Спрингер. ISBN 978-3-319-32211-7 . [ нужна страница ]
- ^ Кохидай, Ласло (2016). «Хемотаксис как выражение коммуникации тетрахимен» . В Вицани, Гюнтер; Новацкий, Мариуш (ред.). Биокоммуникация инфузорий . Спрингер. стр. 65–82. дои : 10.1007/978-3-319-32211-7_5 . ISBN 978-3-319-32211-7 .
- ^ Кохидай Л., Чаба Г. (июль 1998 г.). «Хемотаксис и хемотаксический отбор, индуцированный цитокинами (IL-8, RANTES и TNF-альфа) у одноклеточных Tetrahymenapyriformis». Цитокин . 10 (7): 481–6. дои : 10.1006/cyto.1997.0328 . ПМИД 9702410 . S2CID 33755476 .
- ^ Зигмонд С.Х. (ноябрь 1977 г.). «Способность полиморфно-ядерных лейкоцитов ориентироваться в градиентах хемотаксических факторов» . Журнал клеточной биологии . 75 (2, часть 1): 606–16. дои : 10.1083/jcb.75.2.606 . ПМК 2109936 . ПМИД 264125 .
- ↑ Перейти обратно: Перейти обратно: а б с д Пауэлл В.С., Рокач Дж. (апрель 2015 г.). «Биосинтез, биологические эффекты и рецепторы гидроксиэйкозатетраеновых кислот (HETE) и оксоэйкозатетраеновых кислот (оксо-ETE), полученных из арахидоновой кислоты» . Biochimica et Biophysical Acta (BBA) - Молекулярная и клеточная биология липидов . 1851 (4): 340–55. дои : 10.1016/j.bbalip.2014.10.008 . ПМК 5710736 . ПМИД 25449650 .
- ^ Пауэлл В.С., Рокач Дж. (октябрь 2013 г.). «Хемоаттрактант эозинофилов 5-оксо-ETE и рецептор OXE» . Прогресс в исследованиях липидов . 52 (4): 651–65. дои : 10.1016/j.plipres.2013.09.001 . ПМК 5710732 . ПМИД 24056189 .
- ^ Мацуока Т., Нарумия С. (сентябрь 2007 г.). «Передача сигналов рецептором простагландина при заболевании» . Научный мировой журнал . 7 : 1329–47. дои : 10.1100/tsw.2007.182 . ПМК 5901339 . ПМИД 17767353 .
- ^ Ёкомизо Т. (февраль 2015 г.). «Два различных рецептора лейкотриена B4, BLT1 и BLT2». Журнал биохимии . 157 (2): 65–71. дои : 10.1093/jb/mvu078 . ПМИД 25480980 .
- ^ Соццани С., Чжоу Д., Локати М., Бернаскони С., Луини В., Мантовани А., О'Флаэрти Дж.Т. (ноябрь 1996 г.). «Стимулирующие свойства 5-оксо-эйкозаноидов для моноцитов человека: синергизм с моноцитарными хемотаксическими белками-1 и -3» . Журнал иммунологии . 157 (10): 4664–71. дои : 10.4049/jimmunol.157.10.4664 . ПМИД 8906847 . S2CID 23499393 .
- ^ Кохидай Л., Ланг О и Чаба Г. (2003). «Соответствие хемотаксического диапазона аминокислот и его корреляция с физико-химическими параметрами у Tetrahymenapyriformis - Эволюционные последствия». Клеточная и молекулярная биология . 49 : ОЛ487–95. ПМИД 14995080 .
- ↑ Перейти обратно: Перейти обратно: а б Мюррей, Джеймс Д. (2002). Математическая биология I: Введение (PDF) . Междисциплинарная прикладная математика. Том. 17 (3-е изд.). Нью-Йорк: Спрингер. стр. 395–417. дои : 10.1007/b98868 . ISBN 978-0-387-95223-9 . Архивировано (PDF) из оригинала 6 мая 2022 года.
- ^ Гарасу М., Центлер Ф., Фетцер И., Таллнер М. (2014). «Как хемотаксические характеристики бактерий могут определять структуру их популяций». Биология и биохимия почвы . 69 : 346–358. doi : 10.1016/j.soilbio.2013.11.019 .
- ^ Маккензи, Дана (6 марта 2023 г.). «Как животные следуют за своим носом» . Знающий журнал . Ежегодные обзоры. doi : 10.1146/knowable-030623-4 . S2CID 257388244 . Проверено 13 марта 2023 г.
- ^ Редди, Гаутам; Мурти, Венкатеш Н.; Вергассола, Массимо (10 марта 2022 г.). «Обонятельное зондирование и навигация в турбулентной среде» . Ежегодный обзор физики конденсированного состояния . 13 (1): 191–213. Бибкод : 2022ARCMP..13..191R . doi : 10.1146/annurev-conmatphys-031720-032754 . ISSN 1947-5454 . S2CID 243966350 .
- ^ Лагзи, Иштван (2013). «Химическая робототехника — хемотаксические носители лекарств» . Центральноевропейский медицинский журнал . 8 (4): 377–382. дои : 10.2478/s11536-012-0130-9 . S2CID 84150518 .
- ^ Сенгупта С., Дей К.К., Муддана Х.С., Табуйо Т., Ибеле М.Е., Батлер П.Дж., Сен А. (январь 2013 г.). «Молекулы ферментов как наномоторы». Журнал Американского химического общества . 135 (4): 1406–14. дои : 10.1021/ja3091615 . ПМИД 23308365 .
- ^ Мохаджерани, Ф; Чжао, X; Сомасундар, А; Велегол, Д; Сен, А (2018). «Теория ферментативного хемотаксиса: от экспериментов к моделированию». Биохимия . 57 (43): 6256–6263. arXiv : 1809.02530 . doi : 10.1021/acs.biochem.8b00801 . ПМИД 30251529 . S2CID 52816076 .
- ^ Чжао X, Палаччи Х., Ядав В., Спиринг М.М., Гилсон М.К., Батлер П.Дж., Хесс Х., Бенкович С.Дж., Сен А. (март 2018 г.). «Хемотаксическая сборка, управляемая субстратом, в ферментном каскаде». Природная химия . 10 (3): 311–317. Бибкод : 2018НатЧ..10..311З . дои : 10.1038/nchem.2905 . ПМИД 29461522 .
- ^ Гуха Р., Мохаджерани Ф., Коллинз М., Гош С., Сен А., Велегол Д. (ноябрь 2017 г.). «Хемотаксис молекулярных красителей в полимерных градиентах в растворе». Журнал Американского химического общества . 139 (44): 15588–15591. дои : 10.1021/jacs.7b08783 . ПМИД 29064685 .
Дальнейшее чтение [ править ]
- Альбертс Б., Джонсон А., Льюис Дж., Уолтер П., Рафф MC (2002). «Бактериальный хемотаксис зависит от двухкомпонентного сигнального пути, активируемого гистидин-киназными рецепторами» . Молекулярная биология клетки . Группа Тейлор и Фрэнсис. ISBN 978-0-8153-4069-0 . Проверено 18 сентября 2017 г.
- Багорда А., родитель, Калифорния (август 2008 г.). «Краткий обзор хемотаксиса эукариот» . Журнал клеточной науки . 121 (Часть 16): 2621–4. CiteSeerX 10.1.1.515.32 . дои : 10.1242/jcs.018077 . ПМЦ 7213762 . ПМИД 18685153 .
- Берг ХК (1993). Случайные блуждания в биологии (Расширенная, переработанная ред.). Принстон, Нью-Джерси: Princeton Univ. Нажимать. ISBN 978-0-691-00064-0 .
- Берг ХК (2003). Кишечная палочка в движении . Том. 58. Нью-Йорк: Спрингер. стр. 64–65. Бибкод : 2005ФТ....58б..64Б . дои : 10.1063/1.1897527 . ISBN 978-0-387-00888-2 .
{{cite book}}:|journal=игнорируется ( помогите ) - Дюзенбери Д.Б. (2009). Жизнь на микроуровне: неожиданная физика маленького размера . Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-03116-6 .
- Айзенбах М. (2004). Ленгелер Дж.В. (ред.). Хемотаксис . Лондон: Издательство Имперского колледжа. ISBN 978-1-86094-413-0 .
- Айзенбах, Майкл (декабрь 2011 г.). «Бактериальный хемотаксис». Энциклопедия наук о жизни . дои : 10.1002/9780470015902.a0001251.pub3 . ISBN 978-0470016176 .
{{cite book}}:|journal=игнорируется ( помогите ) - Hazelbauer GL (13 октября 2012 г.). «Бактериальный хемотаксис: первые годы молекулярных исследований» . Ежегодный обзор микробиологии . 66 (1): 285–303. doi : 10.1146/annurev-micro-092611-150120 . ПМЦ 3989901 . ПМИД 22994495 .
- Джин Т., Херельд Д. (2016). Хемотаксис: методы и протоколы . Хумана Пресс. ISBN 978-1-4939-3480-5 .
- Миллер Л.Д., Рассел М.Х., Александр Г. (2009). Разнообразие бактериальных хемотаксических реакций и адаптация к нише . Том. 66. стр. 53–75. дои : 10.1016/S0065-2164(08)00803-4 . ISBN 9780123747884 . ПМИД 19203648 .
{{cite book}}:|journal=игнорируется ( помогите ) - Рао К.В., Кирби-младший, Аркин А.П. (февраль 2004 г.). «Дизайн и разнообразие бактериального хемотаксиса: сравнительное исследование Escherichia coli и Bacillus subtilis» . ПЛОС Биология . 2 (2): Е49. дои : 10.1371/journal.pbio.0020049 . ПМК 340952 . ПМИД 14966542 .
- Уильямс AH (20 декабря 2010 г.). «Хемотаксис в движении – средство активного обучения» . Журнал микробиологии и биологического образования . 11 (2): 177–8. дои : 10.1128/jmbe.v11i2.216 . ПМК 3577161 . ПМИД 23653726 .