Циклооксигеназа-2
| ПТГС2 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | PTGS2 , COX-2, COX2, GRIPGHS, PGG/HS, PGHS-2, PHS-2, hCox-2, простагландин-эндопероксидсинтаза 2 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 600262 ; МГИ : 97798 ; Гомологен : 31000 ; Генные карты : PTGS2 ; ОМА : PTGS2 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Номер ЕС | 1.14.99.1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





Циклооксигеназа-2 ( ЦОГ-2 ), также известная как простагландин-эндопероксидсинтаза 2 ( HUGO PTGS2 ), представляет собой фермент , который у человека кодируется PTGS2 геном . [ 5 ] У человека это одна из трех циклооксигеназ . Он участвует в превращении арахидоновой кислоты в простагландин Н 2 , важный предшественник простациклина , что проявляется при воспалении .
Функция
[ редактировать ]PTGS2 (COX-2) превращает арахидоновую кислоту (АК) в эндопероксид простагландина H2. PTGS являются мишенями для НПВП и специфических ингибиторов PTGS2 (ЦОГ-2), называемых коксибами. PTGS-2 представляет собой гомодимер последовательности. Каждый мономер фермента имеет пероксидазы и PTGS (ЦОГ) активный центр . Ферменты PTGS (COX) катализируют превращение арахидоновой кислоты в простагландины в два этапа. Сначала водород отрывается от углерода 13 арахидоновой кислоты, а затем с помощью PTGS2 (COX-2) добавляются две молекулы кислорода, образуя PGG2. Во-вторых, PGG2 восстанавливается до PGH2 в активном центре пероксидазы. Синтезированный PGH2 превращается в простагландины ( PGD2 , PGE2 , PGF2α . ), простациклин (PGI2) или тромбоксан А2 с помощью тканеспецифических изомераз (рис. 2) [ 6 ]
Метаболизируя арахидоновую кислоту преимущественно до PGG2, ЦОГ-2 также превращает эту жирную кислоту в небольшие количества рацемической смеси 15-гидроксиикозатетраеновых кислот (т.е. 15-НЕТЕ), состоящей из ~22% 15( R )-НЕТЕ и ~78% 15( S )-HETE Стереоизомеры , а также небольшое количество 11( R )-HETE. [ 7 ] Два стереоизомера 15-НЕТЕ обладают собственной биологической активностью, но, что, возможно, более важно, могут в дальнейшем метаболизироваться до основного класса агентов — липоксинов . Кроме того, ЦОГ-2, обработанный аспирином, метаболизирует арахидоновую кислоту почти исключительно до 15( R )-HETE, который может далее метаболизироваться до эпилипоксинов . [ 8 ] Липоксины и эпилипоксины являются мощными противовоспалительными средствами и могут способствовать общей активности обоих ЦОГ, а также аспирина. [ нужна ссылка ]
ЦОГ-2 естественным образом ингибируется кальцитриолом (активной формой витамина D). [ 9 ] [ 10 ]
Механизм
[ редактировать ]
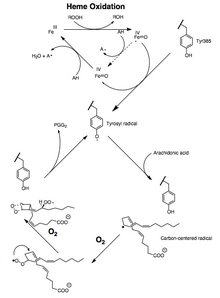
Активность как пероксидазы, так и PTGS инактивируется во время катализа с помощью механизмов первого порядка, что означает, что активность пероксидазы PGHS-2 или PTGS падает до нуля в течение 1–2 минут, даже в присутствии достаточного количества субстратов. [ 12 ] [ 13 ] [ 14 ]
Превращение арахидоновой кислоты в PGG2 можно представить как серию радикальных реакций, аналогичных полиненасыщенных жирных кислот автоокислению . [ 15 ] 13-про(S)-водород отрывается, а дикислород захватывает пентадиенильный радикал у углерода 11. 11-пероксильный радикал циклизуется по углероду 9, а углеродцентрированный радикал, образующийся при C-8, циклизуется по углероду 12, образуя эндопероксид . Образовавшийся аллильный ; радикал улавливается дикислородом у углерода 15 с образованием 15-(S)-пероксильного радикала этот радикал затем восстанавливается до PGG2 . Это подтверждается следующими данными: 1) значительный кинетический изотопный эффект при отщеплении 13-про(S)-водорода наблюдается ; 2) углеродцентрированные радикалы улавливаются в ходе катализа ; [ 16 ] 3) небольшие количества продуктов окисления образуются за счет захвата кислорода интермедиата аллильного радикала в положениях 13 и 15. [ 17 ] [ 18 ]
при котором 13-про(S)-водород депротонируется и карбанион окисляется до другой механизм , радикала . Теоретически возможен Однако оксигенация 10,10-дифторарахидоновой кислоты до 11-(S)-гидроксиэйкоза-5,8,12,14-тетраеновой кислоты не соответствует образованию промежуточного карбаниона, поскольку при этом происходит удаление фторида с образованием сопряженного диена. [ 19 ] Считалось, что отсутствие эндопероксидсодержащих продуктов, полученных из 10,10-дифторарахидоновой кислоты, указывает на важность карбокатиона C-10 в синтезе PGG2 . [ 20 ] Однако катионный механизм требует, чтобы образование эндопероксида происходило до удаления 13-про(S)-водорода. Это не согласуется с результатами изотопных экспериментов по оксигенации арахидоновой кислоты . [ 21 ]
Структура
[ редактировать ]
PTGS2 (COX-2) существует в виде гомодимера, каждый мономер имеет молекулярную массу около 70 кДа. Третичная и четвертичная структуры ферментов PTGS1 (ЦОГ-1) и PTGS2 (ЦОГ-2) практически идентичны. Каждая субъединица имеет три различных структурных домена: короткий домен N-концевого эпидермального фактора роста ( EGF ); α -спиральный мембраносвязывающий фрагмент; и С-концевой каталитический домен. Ферменты PTGS (ЦОГ, который можно спутать с « цитохромоксидазой ») представляют собой монотопные мембранные белки; Мембраносвязывающий домен состоит из серии амфипатических α-спиралей с несколькими гидрофобными аминокислотами , расположенными на монослое мембраны. PTGS1 (COX-1) и PTGS2 (COX-2) представляют собой бифункциональные ферменты, которые осуществляют две последовательные химические реакции в пространственно различных, но механически связанных активных центрах. Активные центры как циклооксигеназы , так и пероксидазы расположены в каталитическом домене, который составляет примерно 80% белка. Каталитический домен гомологичен пероксидазам млекопитающих, таким как миелопероксидаза . [ 23 ] [ 24 ]
Было обнаружено, что PTGS2 человека (COX-2) функционирует как конформационный гетеродимер, имеющий каталитический мономер (E-cat) и аллостерический мономер (E-алло). Гем связывается только с сайтом пероксидазы E-cat, тогда как субстраты, а также некоторые ингибиторы (например, целекоксиб ) связываются с сайтом ЦОГ E-cat. E-cat регулируется E-allo в зависимости от того, какой лиганд связан с E-allo. Субстратные и несубстратные жирные кислоты (ЖК) и некоторые ингибиторы PTGS (COX) (например, напроксен ) преимущественно связываются с сайтом PTGS (COX) E-allo. Арахидоновая кислота может связываться с E-cat и E-allo, но сродство АК к E-allo в 25 раз выше, чем к Ecat. Пальмитиновая кислота, эффективный стимулятор huPGHS-2 , связывает только E-allo в сокристаллах пальмитиновой кислоты/мышиного PGHS-2. Несубстратные жирные кислоты могут усиливать или ослаблять действие ингибиторов PTGS (ЦОГ) в зависимости от жирной кислоты и от того, связывает ли ингибитор E-cat или E-allo. Исследования показывают, что концентрация и состав пула свободных жирных кислот в среде, в которой функционирует PGHS-2 в клетках, также называемый тонусом FA, является ключевым фактором, регулирующим активность PGHS-2 и его ответ на PTGS ( ЦОГ) ингибиторы. [ 22 ]
Клиническое значение
[ редактировать ]
PTGS2 (COX-2) не экспрессируется в нормальных условиях в большинстве клеток, но повышенные уровни обнаруживаются во время воспаления . PTGS1 (COX-1) конститутивно экспрессируется во многих тканях и является преобладающей формой в слизистой оболочке желудка и почках. Ингибирование PTGS1 (COX-1) снижает базальную выработку цитопротекторных PGE2 и PGI2 в желудке , что может способствовать изъязвлению желудка . Поскольку PTGS2 (ЦОГ-2) обычно экспрессируется только в клетках, где простагландины активируются (например, во время воспаления), предполагалось, что препараты-кандидаты, избирательно ингибирующие PTGS2 (ЦОГ-2), проявляют меньше побочных эффектов. [ 24 ] но оказалось, что они существенно увеличивают риск сердечно-сосудистых событий, таких как сердечный приступ и инсульт. Два разных механизма могут объяснить противоречивые эффекты. Низкие дозы аспирина защищают от сердечных приступов и инсультов, блокируя образование простагландина PTGS1 (ЦОГ-1) под названием тромбоксан А2. Он склеивает тромбоциты и способствует свертыванию крови; подавление этого помогает предотвратить болезни сердца. С другой стороны, PTGS2 (COX-2) является более важным источником простагландинов, особенно простациклина, который содержится в слизистой оболочке кровеносных сосудов. Простациклин расслабляет или отклеивает тромбоциты, поэтому селективные ингибиторы ЦОГ-2 (коксибы) повышают риск сердечно-сосудистых событий из-за тромбообразования. [ 26 ]
Нестероидные противовоспалительные препараты (НПВП) ингибируют выработку простагландинов PTGS1 (ЦОГ-1) и PTGS2 (ЦОГ-2). НПВП, селективные для ингибирования PTGS2 (ЦОГ-2), с меньшей вероятностью, чем традиционные препараты, вызывают побочные эффекты со стороны желудочно-кишечного тракта , но могут вызывать сердечно-сосудистые события, такие как сердечная недостаточность , инфаркт миокарда и инсульт . Исследования фармакологии и генетики человека , генетически модифицированных грызунов и других животных моделей, а также рандомизированные исследования показывают, что это происходит из-за подавления PTGS2 (ЦОГ-2)-зависимых кардиозащитных простагландинов , простациклина . в частности [ 27 ]
Экспрессия PTGS2 (COX-2) повышается при многих видах рака. Сверхэкспрессия PTGS2 (COX-2) наряду с повышенным ангиогенезом и экспрессией SLC2A1 (GLUT-1) в значительной степени связана с карциномами желчного пузыря. [ 28 ] Кроме того, продукт PTGS2 (COX-2), PGH 2, преобразуется синтазой простагландина E2 в PGE 2 , что, в свою очередь, может стимулировать прогрессирование рака. Следовательно, ингибирование PTGS2 (ЦОГ-2) может принести пользу в профилактике и лечении этих типов рака. [ 29 ] [ 30 ]
Экспрессия ЦОГ-2 была обнаружена в идиопатических эпиретинальных мембранах человека. [ 31 ] Блокирование циклооксигеназ лорноксикамом в острой стадии воспаления снижало частоту мембранообразования на 43% в диспазной модели ПВР и на 31% в конканавалиновой . Лорноксикам не только нормализовал экспрессию циклооксигеназ в обеих моделях ПВР, но и нейтрализовал изменения сетчатки и толщины сосудистой оболочки , вызванные введением провоспалительных агентов. Эти факты подчеркивают важность циклооксигеназ и простагландинов в развитии ПВР. [ 32 ]
Активация гена PTGS2 также связана с несколькими стадиями репродукции человека. Присутствие гена обнаружено в хорионической пластинке , в эпителии амниона , синцитиотрофобластах , ворсинчатых фибробластах, хорионических трофобластах , амниотических трофобластах , а также в базальной пластинке плаценты , в децидуальных клетках и вневорсинчатых цитотрофобластах . Во время процесса хориоамнионита /децидуита повышение регуляции PTGS2 в амнионе и хориодецидуе является одним из трех ограниченных эффектов воспаления в матке . Повышенная экспрессия гена PTGS2 в плодных оболочках связана с наличием воспаления, вызывающего экспрессию гена простагландина матки и иммунолокализацию белков пути простагландина в клетках хорионического трофобласта и прилегающей децидуальной оболочки, или хориодецидуальной оболочки. PTGS2 связан с воспалительной системой и наблюдается в воспалительных лейкоцитах . Было отмечено, что существует положительная корреляция с экспрессией PTGS2 в амнионе во время самопроизвольных родов, и было обнаружено, что экспрессия увеличивается с гестационным возрастом после наличия родов, при этом никаких изменений не наблюдается в амнионе и хориодецидуальной оболочке ни во время преждевременных, ни в доношенных родах. Кроме того, окситоцин стимулирует экспрессию PTGS2 в клетках миометрия . [ 33 ]
Было показано, что носители мутантного аллеля PTGS2 5939C среди населения ханьцев имеют более высокий риск развития рака желудка . Кроме того, была обнаружена связь между заражением Helicobacter pylori и наличием аллеля 5939С. [ 34 ]
Взаимодействия
[ редактировать ]Было показано, что PTGS2 взаимодействует с кавеолином 1 . [ 35 ]
История
[ редактировать ]PTGS2 (COX-2) был открыт в 1991 году лабораторией Симмонса . Дэниела [ 36 ] [ нужен лучший источник ] в Университете Бригама Янга.
См. также
[ редактировать ]- Арахидоновая кислота
- Циклооксигеназа
- Циклооксигеназа 1
- НПВП
- Открытие и разработка селективных ингибиторов ЦОГ-2.
- Селективный ингибитор ЦОГ-2
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000073756 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000032487 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Хла Т, Нилсон К. (август 1992 г.). «КДНК циклооксигеназы-2 человека» . Учеб. Натл. акад. наук. США . 89 (16): 7384–8. Бибкод : 1992PNAS...89.7384H . дои : 10.1073/pnas.89.16.7384 . ПМК 49714 . ПМИД 1380156 .
- ^ О'Бэнион МК (1999). «Циклооксигеназа-2: молекулярная биология, фармакология и нейробиология». Крит преподобный Нейробиол . 13 (1): 45–82. дои : 10.1615/critrevneurobiol.v13.i1.30 . ПМИД 10223523 .
- ^ Мулугета С., Сузуки Т., Эрнандес Н.Т., Гриссер М., Боглин В.Е., Шнайдер С. (2010). «Идентификация и абсолютная конфигурация дигидроксиарахидоновых кислот, образующихся в результате оксигенации 5S-HETE нативным и ацетилированным аспирином ЦОГ-2» . Дж. Липид Рес . 51 (3): 575–85. дои : 10.1194/jlr.M001719 . ПМЦ 2817587 . ПМИД 19752399 .
- ^ Серхан CN (2005). «Липоксины и 15-эпилипоксины, запускаемые аспирином, являются первыми липидными медиаторами эндогенного противовоспалительного процесса и разрешения». Простагландины Лейкот. Сущность. Жирные кислоты . 73 (3–4): 141–62. дои : 10.1016/j.plefa.2005.05.002 . ПМИД 16005201 .
- ^ Ван Ц, Хэ Ю, Шэнь Ю, Чжан Ц, Чэнь Д, Цзо С, Цинь Дж, Ван Х, Ван Дж, Ю Ю (2014). «Витамин D ингибирует экспрессию ЦОГ-2 и воспалительную реакцию, воздействуя на члена 4 суперсемейства тиоэстеразы» . J Биол Хим . 289 (17): 11681–11694. дои : 10.1074/jbc.M113.517581 . ПМК 4002078 . ПМИД 24619416 .
- ^ Касси Э., Адамопулос С., Басдра Е.К., Папавассилиу А.Г. (2013). «Роль витамина D при атеросклерозе» . Тираж . 128 (23): 2517–2531. doi : 10.1161/CIRCULATIONAHA.113.002654 . ПМИД 24297817 .
- ^ ПДБ : 3ОЛТ
- ^ Смит В.Л., Гаравито Р.М., ДеВитт Д.Л. (декабрь 1996 г.). «Простагландин-эндопероксид Н-синтазы (циклооксигеназы)-1 и -2» . Ж. Биол. Хим . 271 (52): 33157–60. дои : 10.1074/jbc.271.52.33157 . ПМИД 8969167 .
- ^ Ву Г, Вэй С., Кулмач Р.Дж., Осава Ю., Цай А.Л. (апрель 1999 г.). «Механистическое исследование самоинактивации пероксидазной активности простагландин H-синтазы-1» . Ж. Биол. Хим . 274 (14): 9231–7. дои : 10.1074/jbc.274.14.9231 . ПМИД 10092596 .
- ^ Каллан, Огайо, Со Ой, Суинни, округ Колумбия (февраль 1996 г.). «Кинетические факторы, которые определяют сродство и селективность медленного ингибирования связывания человеческой простагландин H-синтазы 1 и 2 индометацином и флурбипрофеном» . Ж. Биол. Хим . 271 (7): 3548–54. дои : 10.1074/jbc.271.7.3548 . ПМИД 8631960 .
- ^ Портер Н.А. (1986). «Механизмы автоокисления полиненасыщенных липидов». Отчеты о химических исследованиях . 19 (9): 262–8. дои : 10.1021/ar00129a001 .
- ^ Мейсон Р.П., Кальянараман Б., Тайнер Б.Е., Элинг Т.Е. (июнь 1980 г.). «Углеродоцентрированный свободный радикальный промежуточный продукт при окислении арахидоновой кислоты простагландинсинтетазой. Исследования спинового захвата и поглощения кислорода» . Ж. Биол. Хим . 255 (11): 5019–22. дои : 10.1016/S0021-9258(19)70741-8 . ПМИД 6246094 .
- ^ Хекер М., Ульрих В., Фишер К., Миз Ко (ноябрь 1987 г.). «Идентификация новых метаболитов арахидоновой кислоты, образуемых синтетазой простагландина H» . Евро. Дж. Биохим . 169 (1): 113–23. дои : 10.1111/j.1432-1033.1987.tb13587.x . ПМИД 3119336 .
- ^ Сяо Г., Цай А.Л., Палмер Г., Бояр В.К., Маршалл П.Дж., Кулмач Р.Дж. (февраль 1997 г.). «Анализ тирозильных радикалов, индуцированных гидропероксидом, и активности липоксигеназы в обработанной аспирином человеческой простагландин H-синтазе-2». Биохимия . 36 (7): 1836–45. дои : 10.1021/bi962476u . ПМИД 9048568 .
- ^ Квок П.Ю., Мюлльнер Ф.В., Фрид Дж. (июнь 1987 г.). «Ферментативные превращения 10,10-дифторарахидоновой кислоты с помощью PGH-синтазы и липоксигеназы сои». Журнал Американского химического общества . 109 (12): 3692–3698. дои : 10.1021/ja00246a028 .
- ^ Дин AM, Дин FM (май 1999 г.). «Карбокатионы в синтезе простагландинов циклооксигеназой PGH-синтазы? Радикальный сдвиг!» . Белковая наука . 8 (5): 1087–98. дои : 10.1110/ps.8.5.1087 . ПМК 2144324 . ПМИД 10338019 .
- ^ Хамберг М., Самуэльссон Б. (ноябрь 1967 г.). «О механизме биосинтеза простагландинов Е-1 и F-1-альфа» . Ж. Биол. Хим . 242 (22): 5336–43. дои : 10.1016/S0021-9258(18)99433-0 . ПМИД 6070851 .
- ^ Jump up to: а б Донг Л., Веккио А.Дж., Шарма Н.П., Джурбан Б.Дж., Малковски М.Г., Смит В.Л. (май 2011 г.). «Человеческая циклооксигеназа-2 представляет собой гомодимер последовательности, который функционирует как конформационный гетеродимер» . Ж. Биол. Хим . 286 (21): 19035–46. дои : 10.1074/jbc.M111.231969 . ПМК 3099718 . ПМИД 21467029 .
- ^ Пико Д., Лолл П.Дж., Гаравито Р.М. (январь 1994 г.). «Рентгеновская кристаллическая структура мембранного белка простагландин H2-синтазы-1». Природа . 367 (6460): 243–9. Бибкод : 1994Natur.367..243P . дои : 10.1038/367243a0 . ПМИД 8121489 . S2CID 4340064 .
- ^ Jump up to: а б Курумбайл Р.Г., Кифер-младший, Марнетт Л.Дж. (декабрь 2001 г.). «Ферменты циклооксигеназы: катализ и ингибирование». Курс. Мнение. Структура. Биол . 11 (6): 752–60. дои : 10.1016/S0959-440X(01)00277-9 . ПМИД 11751058 .
- ^ ПДБ : 3PGH
- ^ Руан Ч., Со СП, Руан К.Х. (2011). «Индуцибельный ЦОГ-2 доминирует над ЦОГ-1 в биосинтезе простациклина: механизмы риска развития сердечно-сосудистых заболеваний ингибитором ЦОГ-2» . Науки о жизни . 88 (1–2): 24–30. дои : 10.1016/j.lfs.2010.10.017 . ПМК 3046773 . ПМИД 21035466 .
- ^ Ван Д., Патель В.В., Риччиотти Э., Чжоу Р., Левин М.Д., Гао Е., Ю З, Феррари В.А., Лу ММ, Сюй Дж., Чжан Х., Хуэй Ю., Ченг Ю., Петренко Н., Ю Ю., Фитцджеральд Г.А. (май 2009 г.) ). «Кардиомиоцитарная циклооксигеназа-2 влияет на сердечный ритм и функцию» . Учеб. Натл. акад. наук. ОЛЕНЬ . 106 (18): 7548–52. Бибкод : 2009PNAS..106.7548W . дои : 10.1073/pnas.0805806106 . ПМК 2670242 . ПМИД 19376970 .
- ^ Леган М. (август 2010 г.). «Циклооксигеназа-2, р53 и транспортер глюкозы-1 как предикторы злокачественности при развитии карцином желчного пузыря» . Босн Дж. Базовые медицинские науки . 10 (3): 192–6. дои : 10.17305/bjbms.2010.2684 . ПМК 5504494 . ПМИД 20846124 .
- ^ EnterGene 5743
- ^ Ментер Д.Г., Шильский Р.Л., Дюбуа Р.Н. (март 2010 г.). «Циклооксигеназа-2 и лечение рака: понимание риска должно стоить награды» . Клин. Рак Рез . 16 (5): 1384–90. дои : 10.1158/1078-0432.CCR-09-0788 . ПМЦ 4307592 . ПМИД 20179228 .
- ^ КАСЕ С., САИТО В., ОНО С., ИШИДА С. (2010). «Экспрессия циклооксигеназы-2 в идиопатической эпиретинальной мембране человека». Ретина . 30 (5): 719–723. дои : 10.1097/iae.0b013e3181c59698 . ПМИД 19996819 . S2CID 205650971 .
- ^ Тихонович М.В., Эрдиаков А.К., Гаврилова С.А. (21.06.2017). «Нестероидная противовоспалительная терапия подавляет развитие пролиферативной витреоретинопатии более эффективно, чем стероидная». Международная офтальмология . 38 (4): 1365–1378. дои : 10.1007/s10792-017-0594-3 . ISSN 0165-5701 . ПМИД 28639085 . S2CID 4017540 .
- ^ Филлипс, Роберт Дж. и др. «На экспрессию генов пути простагландинов в плаценте человека, амнионе и хориодецидуе по-разному влияют преждевременные и доношенные роды, а также воспаление матки». Беременность и роды BMC, том. 14 241. 22 июля 2014 г., номер номера: 10.1186/1471-2393-14-241.
- ^ Ли Ю, Хэ В, Лю Т, Чжан Ц (декабрь 2010 г.). «Новый вариант гена циклооксигеназы-2 у ханьской популяции связан с повышенным риском рака желудка». Мол Диагн Тер . 14 (6): 351–5. дои : 10.1007/bf03256392 . ПМИД 21275453 . S2CID 1229751 .
- ^ Лиу Дж.Ю., Дэн В.Г., Гилрой Д.В., Шюэ С.К., Ву К.К. (сентябрь 2001 г.). «Колокализация и взаимодействие циклооксигеназы-2 с кавеолином-1 в фибробластах человека» . Ж. Биол. Хим . 276 (37): 34975–82. дои : 10.1074/jbc.M105946200 . ПМИД 11432874 .
- ^ Се В.Л., Чипман Дж.Г., Робертсон Д.Л., Эриксон Р.Л., Симмонс Д.Л. (апрель 1991 г.). «Экспрессия митоген-чувствительного гена, кодирующего простагландинсинтазу, регулируется сплайсингом мРНК» . Учеб. Натл. акад. наук. США . 88 (7): 2692–6. Бибкод : 1991PNAS...88.2692X . дои : 10.1073/pnas.88.7.2692 . ПМК 51304 . ПМИД 1849272 .
Дальнейшее чтение
[ редактировать ]- Ричардс Дж.А., Буревестник Т.А., Брюггемайер Р.В. (февраль 2002 г.). «Сигнальные пути, регулирующие ароматазу и циклооксигеназы в нормальных и злокачественных клетках молочной железы». J. Стероидная биохимия. Мол. Биол . 80 (2): 203–12. дои : 10.1016/S0960-0760(01)00187-X . ПМИД 11897504 . S2CID 12728545 .
- Ву Т, Ву Х, Ван Дж, Ван Дж (2011). «Экспрессия и клеточная локализация циклооксигеназ и синтаз простагландина Е в геморрагическом мозге» . J Нейровоспаление . 8:22 . дои : 10.1186/1742-2094-8-22 . ПМК 3062590 . ПМИД 21385433 .
- Коки А.Т., Хан Н.К., Вернер Б.М., Зайберт К., Хармон Дж.Л., Данненберг А.Дж., Сослоу Р.А., Масферрер Дж.Л. (январь 2002 г.). «Характеристика циклооксигеназы-2 (ЦОГ-2) во время онкогенеза при эпителиальном раке человека: доказательства потенциальной клинической пользы ингибиторов ЦОГ-2 при эпителиальном раке». Простагландины Лейкот. Сущность. Жирные кислоты . 66 (1): 13–8. дои : 10.1054/plef.2001.0335 . ПМИД 12051953 .
- Саукконен К., Ринтахака Дж., Сивула А., Бускенс С.Дж., Ван Рис Б.П., Рио М.К., Хаглунд С., Ван Ланшот Дж.Дж., Офферхаус Г.Дж., Ристимаки А. (октябрь 2003 г.). «Циклооксигеназа-2 и канцерогенез желудка». АПМИС . 111 (10): 915–25. дои : 10.1034/j.1600-0463.2003.1111001.x . ПМИД 14616542 . S2CID 23257867 .
- Синикроп Ф.А., Гилл С. (2004). «Роль циклооксигеназы-2 при колоректальном раке». Раковые метастазы Rev. 23 (1–2): 63–75. дои : 10.1023/А:1025863029529 . ПМИД 15000150 . S2CID 21521040 .
- Джайн С., Хури Ф.Р., Шин Д.М. (2004). «Профилактика рака головы и шеи: современное состояние и перспективы». Карр Проблема рака . 28 (5): 265–86. doi : 10.1016/j.currproblcancer.2004.05.003 . ПМИД 15375804 .
- Саба Н., Джайн С., Хури Ф. (2004). «Химиопрофилактика рака легких». Карр Проблема рака . 28 (5): 287–306. doi : 10.1016/j.currproblcancer.2004.05.005 . ПМИД 15375805 .
- Кардилло И, Спугнини Э.П., Вердина А., Галати Р., Ситро Дж., Бальди А. (октябрь 2005 г.). «Кокс и мезотелиома: обзор» Гистол. Гистопатол . 20 (4): 1267–74. ПМИД 16136507 .
- Брюггемайер Р.В., Диас-Крус Э.С. (март 2006 г.). «Связь между ароматазой и циклооксигеназой при раке молочной железы: потенциал новых терапевтических подходов». Минерва Эндокринол . 31 (1): 13–26. ПМИД 16498361 .
- Фудзимура Т., Охта Т., Ояма К., Мияшита Т., Мива К. (март 2006 г.). «Роль циклооксигеназы-2 в канцерогенезе рака желудочно-кишечного тракта: обзор и отчет из личного опыта» . Мир Дж. Гастроэнтерол . 12 (9): 1336–45. дои : 10.3748/wjg.v12.i9.1336 . ПМК 4124307 . ПМИД 16552798 .
- Бингхэм С., Бесвик П.Дж., Блюм Д.Э., Грей Н.М., Чесселл И.П. (октябрь 2006 г.). «Роль пути циклооксигеназы в ноцицепции и боли». Семин. Сотовое развитие. Биол . 17 (5): 544–54. дои : 10.1016/j.semcdb.2006.09.001 . ПМИД 17071117 .
- Мингетти Л., Поккьяри М. (2007). Циклооксигеназа-2, простагландин Е2 и активация микроглии при прионных заболеваниях . Международное обозрение нейробиологии. Том. 82. С. 265–75. дои : 10.1016/S0074-7742(07)82014-9 . ISBN 978-0-12-373989-6 . ПМИД 17678966 .
Внешние ссылки
[ редактировать ]- Nextbio. Архивировано 18 октября 2019 г. в Wayback Machine.
- НПВП и сердечно-сосудистый риск объяснены согласно исследованиям Медицинской школы Перельмана
- Вулф М.М. (декабрь 2004 г.). «Рофекоксиб, Merck и FDA». Н. англ. Дж. Мед . 351 (27): 2875–8, ответ автора 2875–8. дои : 10.1056/NEJM200412303512719 . ПМИД 15625749 .