сетчатка
| сетчатка | |
|---|---|
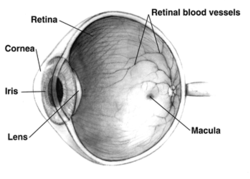 правого человеческого глаза Поперечное сечение ; Глаза значительно различаются у животных. | |
| Подробности | |
| Произношение | Великобритания : / ˈ r ɛ t ɪ n ə / , США : / ˈ r ɛ t ən ə / , пл. сетчатка /- н и / |
| Часть | Глаз |
| Система | Визуальная система |
| Артерия | Центральная артерия сетчатки |
| Идентификаторы | |
| латинский | rēte, внутренние луковицы туники |
| МеШ | D012160 |
| ТА98 | А15.2.04.002 |
| ТА2 | 6776 |
| ФМА | 58301 |
| Анатомическая терминология | |
Сетчатка латинского от ( rete «сеть»; мн. retinae или retinas — самый внутренний светочувствительный слой ткани глаза большинства и позвоночных ) некоторых моллюсков . Оптика глаза создает на сетчатке сфокусированное двухмерное изображение зрительного мира, которое затем обрабатывает это изображение внутри сетчатки и отправляет нервные импульсы по зрительному нерву в зрительную кору для создания зрительного восприятия . Сетчатка выполняет функцию, во многом аналогичную функции пленки или датчика изображения в фотоаппарате .
Нервная сетчатка состоит из нескольких слоев нейронов, соединенных между собой синапсами и поддерживается наружным слоем пигментированных эпителиальных клеток. Основными светочувствительными клетками сетчатки являются фоторецепторные клетки , которые бывают двух типов: палочки и колбочки . Палочки функционируют преимущественно при тусклом свете и обеспечивают монохроматическое зрение. Колбочки функционируют в хорошо освещенных условиях и отвечают за восприятие цвета посредством использования ряда опсинов , а также за остроту зрения, используемую для таких задач, как чтение. Третий тип светочувствительных клеток, светочувствительные ганглиозные клетки , важен для управления циркадными ритмами и рефлекторными реакциями, такими как зрачковый световой рефлекс .
Свет, попадающий на сетчатку, инициирует каскад химических и электрических событий, которые в конечном итоге запускают нервные импульсы, которые посылаются в различные зрительные центры мозга через волокна зрительного нерва . Нейронные сигналы от палочек и колбочек подвергаются обработке другими нейронами, выходные данные которых принимают форму потенциалов действия в ганглиозных клетках сетчатки которых , аксоны образуют зрительный нерв. [1]
При эмбриональном развитии позвоночных сетчатка и зрительный нерв возникают как выросты развивающегося мозга, в частности, эмбрионального промежуточного мозга ; таким образом, сетчатка считается частью центральной нервной системы (ЦНС) и фактически представляет собой ткань мозга. [2] [3] Это единственная часть ЦНС, которую можно визуализировать неинвазивно . Как и большая часть головного мозга, сетчатка изолирована от сосудистой системы гематоэнцефалическим барьером . Сетчатка является частью тела с наибольшим постоянным потреблением энергии. [4]
Структура [ править ]
неинвертированная и сетчатка Инвертированная
Сетчатка позвоночных перевернута в том смысле, что светочувствительные клетки расположены в задней части сетчатки, поэтому свету приходится проходить через слои нейронов и капилляров, прежде чем он достигнет светочувствительных участков палочек и колбочек. [5] Ганглиозные клетки, аксоны которых образуют зрительный нерв, находятся в передней части сетчатки; следовательно, зрительный нерв должен пересечь сетчатку на пути к мозгу. В этой области нет фоторецепторов, что приводит к возникновению слепого пятна . [6] Напротив, в сетчатке головоногих фоторецепторы расположены спереди, а обрабатывающие нейроны и капилляры позади них. Благодаря этому у головоногих моллюсков нет слепого пятна.
Хотя вышележащая нервная ткань частично прозрачна, а сопутствующие глиальные клетки действуют как оптоволоконные каналы для транспортировки фотонов непосредственно к фоторецепторам, [7] [8] рассеяние света . происходит [9] У некоторых позвоночных, в том числе и у человека, имеется участок центральной сетчатки, приспособленный для остроты зрения. Эта область, называемая центральной ямкой , бессосудистая (не имеет кровеносных сосудов) и имеет минимальное количество нервной ткани перед фоторецепторами, что сводит к минимуму рассеяние света. [9]
Головоногие моллюски имеют неперевернутую сетчатку, сравнимую по разрешающей способности с глазами многих позвоночных. Глаза кальмара не имеют аналога пигментного эпителия сетчатки (ПЭС) позвоночных. Хотя их фоторецепторы содержат белок ретинохром, который перерабатывает сетчатку и повторяет одну из функций РПЭ позвоночных, фоторецепторы головоногих, вероятно, не поддерживаются так хорошо, как у позвоночных, и в результате полезный срок службы фоторецепторов у беспозвоночных значительно увеличивается. короче, чем у позвоночных. [10] Легко заменяя стебельчатые глаза (некоторые омары) или сетчатку (некоторые пауки, например дейнопис [11] ) встречается редко.
Сетчатка головоногих не возникает как отросток головного мозга, как у позвоночных. Это различие предполагает, что глаза позвоночных и головоногих не гомологичны , а развивались отдельно. С эволюционной точки зрения, более сложная структура, такая как инвертированная сетчатка, обычно может возникнуть как следствие двух альтернативных процессов — выгодного «хорошего» компромисса между конкурирующими функциональными ограничениями или как исторический неадаптивный реликт запутанного пути эволюции органов. и трансформация. Зрение является важной адаптацией у высших позвоночных.
Третья точка зрения на «перевернутый» глаз позвоночных заключается в том, что он сочетает в себе два преимущества: поддержание упомянутых выше фоторецепторов и снижение интенсивности света, необходимое для того, чтобы избежать ослепления фоторецепторов, которые основаны на чрезвычайно чувствительных глазах предков позвоночных. современная миксина (рыба, обитающая в очень глубокой темной воде). [12]
Недавнее исследование эволюционной цели перевернутой структуры сетчатки, проведенное APS (Американским физическим обществом) [13] говорит, что «Направление глиальных клеток помогает повысить четкость человеческого зрения. Но мы также заметили кое-что довольно любопытное: цвета, которые лучше всего проходят через глиальные клетки, были от зеленого до красного, которые глазу больше всего необходимы для дневного зрения. Глаз обычно получает слишком много синего цвета и, следовательно, имеет меньше колбочек, чувствительных к синему цвету.
Дальнейшее компьютерное моделирование показало, что зеленый и красный свет концентрируются в глиальных клетках и в соответствующих колбочках в пять-десять раз больше, чем синий свет. Вместо этого избыток синего света рассеивается на окружающие стержни. Эта оптимизация такова, что цветовое зрение в течение дня улучшается, а ночное зрение страдает очень незначительно».
Слои сетчатки [ править ]




Сетчатка позвоночных состоит из 10 отдельных слоев. [15] От ближайшего к наиболее дальнему от стекловидного тела:
- Внутренняя пограничная мембрана – базальная мембрана, образованная клетками Мюллера.
- Слой нервных волокон - аксоны тел ганглиозных клеток (между этим слоем и внутренней ограничивающей мембраной существует тонкий слой опорных пластинок клеток Мюллера)
- Слой ганглиозных клеток - содержит ядра ганглиозных клеток, аксоны которых становятся волокнами зрительного нерва, и некоторые смещенные амакриновые клетки. [2]
- Внутренний плексиформный слой - содержит синапс между аксонами биполярных клеток и дендритами ганглиозных и амакриновых клеток. [2]
- Внутренний ядерный слой – содержит ядра и окружающие их клеточные тела (перикарии) амакриновых клеток , биполярных клеток и горизонтальных клеток. [2]
- Наружный плексиформный слой – выступы палочек и колбочек, оканчивающиеся сферой палочки и ножкой колбочки соответственно, образуют синапсы с дендритами биполярных клеток и горизонтальных клеток. [2] В макулярной области это известно как слой волокон Генле .
- Наружный ядерный слой – тела клеток палочек и колбочек.
- Внешняя пограничная мембрана - слой, который отделяет части внутренних сегментов фоторецепторов от ядер их клеток.
- Внутренний сегмент/наружный сегментный слой – внутренние сегменты и внешние сегменты палочек и колбочек, внешние сегменты содержат узкоспециализированный светочувствительный аппарат. [16] [17]
- Пигментный эпителий сетчатки – однослойные кубовидные эпителиальные клетки (выступы на схеме не показаны). Этот слой наиболее близок к сосудистой оболочке и обеспечивает питание и поддержку нейронов сетчатки. Черный пигмент меланин в пигментном слое предотвращает отражение света по всему глазному яблоку; это чрезвычайно важно для ясного зрения. [18] [19] [20]
Эти слои можно сгруппировать в четыре основных этапа обработки: фоторецепция; передача в биполярные клетки ; передача на ганглиозные клетки , которые также содержат фоторецепторы, светочувствительные ганглиозные клетки ; и передача по зрительному нерву. На каждой синаптической стадии горизонтальные и амакриновые клетки также соединяются латерально.
Зрительный нерв — это центральный тракт многих аксонов ганглиозных клеток, соединяющийся в первую очередь с латеральным коленчатым телом , зрительной ретрансляционной станцией в промежуточном мозге (задняя часть переднего мозга). Он также проецируется на верхний холмик , супрахиазматическое ядро и ядро зрительного тракта . Он проходит через другие слои, образуя диск зрительного нерва у приматов. [21]
Дополнительные структуры, не связанные непосредственно со зрением, у некоторых групп позвоночных встречаются в виде выростов сетчатки. У птиц гребешок — сосудистая структура сложной формы , выступающая из сетчатки в стекловидное тело ; он снабжает глаза кислородом и питательными веществами, а также может способствовать улучшению зрения. Рептилии имеют похожее, но гораздо более простое строение. [22]
У взрослого человека вся сетчатка составляет около 72% сферы диаметром около 22 мм. Вся сетчатка содержит около 7 миллионов колбочек и от 75 до 150 миллионов палочек. Диск зрительного нерва, часть сетчатки, которую иногда называют «слепым пятном» из-за отсутствия фоторецепторов, расположен в сосочке зрительного нерва , где волокна зрительного нерва выходят из глаза. Он выглядит как овальная белая область размером 3 мм. 2 . Височно (в направлении висков) от этого диска находится макула , в центре которой находится ямка , ямка, которая отвечает за острое центральное зрение, но на самом деле менее чувствительна к свету из-за отсутствия палочек. , не относящихся к человеку У человека и приматов , имеется одна ямка, в отличие от некоторых видов птиц, таких как ястребы, которые являются двулистными, а также собак и кошек, у которых нет ямки, но имеется центральная полоса, известная как зрительная полоса. [ нужна ссылка ] Вокруг ямки простирается центральная сетчатка примерно на 6 мм, а затем периферическая сетчатка. Самый дальний край сетчатки определяется зубчатым краем . Расстояние от одного ора до другого (или макулы), наиболее чувствительной зоны по горизонтальному меридиану , составляет около 32 мм. [ нужны разъяснения ]
На разрезе сетчатка имеет толщину не более 0,5 мм. Он имеет три слоя нервных клеток и два синапса , включая уникальный ленточный синапс . Зрительный нерв несет ганглиозных клеток аксоны в мозг и кровеносные сосуды, снабжающие сетчатку. Ганглиозные клетки лежат внутри глаза, а фоторецепторные клетки — за его пределами. Из-за такого нелогичного устройства свет должен сначала пройти через ганглиозные клетки и вокруг них, а также через толщу сетчатки (включая ее капиллярные сосуды, не показаны), прежде чем достичь палочек и колбочек. Свет поглощается пигментным эпителием сетчатки или хориоидеей (оба непрозрачны).
Лейкоциты . в капиллярах перед фоторецепторами можно воспринимать как крошечные яркие движущиеся точки, если смотреть на синий свет Это известно как энтоптический феномен синего поля (или феномен Ширера).
Между слоем ганглиозных клеток и палочками и колбочками расположены два слоя нейропилей , где и осуществляются синаптические контакты. Слои нейропиля представляют собой внешний плексиформный слой и внутренний плексиформный слой . Во внешнем слое нейропиля палочки и колбочки соединяются с вертикально идущими биполярными клетками , а горизонтально ориентированные горизонтальные клетки соединяются с ганглиозными клетками.
В центральной части сетчатки преобладают колбочки, а в периферической — палочки. Всего сетчатка насчитывает около семи миллионов колбочек и ста миллионов палочек. В центре макулы находится фовеальная ямка, где колбочки узкие и длинные, расположены в виде шестиугольной мозаики , наиболее плотной, в отличие от гораздо более толстых колбочек, расположенных более периферически в сетчатке. [23] В фовеальной ямке другие слои сетчатки смещаются, а затем наращиваются вдоль фовеального наклона до тех пор, пока не будет достигнут край фовеа, или парафовеа , который является самой толстой частью сетчатки. Макула имеет желтую пигментацию из-за экранирующих пигментов и известна как желтое пятно. Область, непосредственно окружающая ямку, имеет наибольшую плотность палочек, сходящихся в одиночные биполярные клетки. Поскольку ее конусы имеют гораздо меньшую конвергенцию сигналов, ямка обеспечивает максимально четкое зрение, которого только может достичь глаз. [2]
Хотя палочка и колбочки представляют собой своего рода мозаику , передача от рецепторов к биполярным и ганглиозным клеткам не является прямой. Поскольку существует около 150 миллионов рецепторов и только 1 миллион волокон зрительного нерва, должна произойти конвергенция и, следовательно, смешивание сигналов. Более того, горизонтальное действие горизонтальных и амакриновых клеток может позволить одной области сетчатки контролировать другую (например, один стимул подавляет другой). Это торможение является ключом к уменьшению количества сообщений, отправляемых в высшие отделы мозга. У некоторых низших позвоночных (например, голубя ) контроль сообщений является «центробежным» — то есть один слой может контролировать другой, или более высокие области мозга могут управлять нервными клетками сетчатки, но у приматов этого не происходит. [2]
когерентной которые можно визуализировать с помощью оптической Слои , томографии
С помощью оптической когерентной томографии (ОКТ) в сетчатке можно идентифицировать 18 слоев. Слои и анатомическая корреляция: [24] [25] [26]

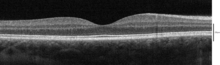

От самого внутреннего до самого внешнего слои, идентифицируемые OCT, следующие:
| # | Слой OCT/обычная этикетка | Анатомический коррелят | Отражательная способность | Специфический анатомический границы? | Дополнительный ссылки |
|---|---|---|---|---|---|
| 1 | Заднее корковое стекловидное тело | Заднее корковое стекловидное тело | Гиперотражающий | Да | [25] |
| 2 | Преретинальное пространство | В глазах, где стекловидное тело полностью или частично отслоилось от сетчатки, это пространство, образующееся между задней корковой поверхностью стекловидного тела и внутренней пограничной мембраной сетчатки. | Гипоотражающий | [25] | |
| 3 | Внутренняя ограничительная мембрана (ILM) | Сформировано ячеек Мюллера. конечными ножками (неясно, можно ли это наблюдать на ОКТ) | Гиперотражающий | Нет | [25] |
| Слой нервных волокон (НФЛ) | ганглиозных клеток Аксоны направляются к зрительному нерву. | ||||
| 4 | Слой ганглиозных клеток (GCL) | Тела ганглиозных клеток (и некоторые смещенные амакриновые клетки ) | Гипоотражающий | [25] | |
| 5 | Внутренний плексиформный слой (IPL) | Синапсы между биполярными , амакриновыми и ганглиозными клетками. | Гиперотражающий | [25] | |
| 6 | Внутренний ядерный слой (ВЯС) | а) Горизонтальные , биполярные и амакриновые клеточные тела б) клеток Мюллера ядра | Гипоотражающий | [25] | |
| 7 | Внешний плексиформный слой (OPL) | Синапсы между фоторецепторами , биполярными и горизонтальными клетками | Гиперотражающий | [25] | |
| 8 | (Внутренняя половина) Слой нервных волокон Генле (HL) | фоторецепторов Аксоны (наклонно ориентированные волокна; отсутствуют в средней или периферической части сетчатки) | Гипоотражающий | Нет | [25] |
| (Внешняя половина) Внешний ядерный слой (ОНЛ) | фоторецепторных Тела клеток | ||||
| 9 | Внешняя ограничительная мембрана (ELM) | Состоит из зонул, прикрепляющихся между клетками Мюллера и внутренними сегментами фоторецепторов. | Гиперотражающий | [25] | |
| 10 | Миоидная зона (МЗ) | Самая внутренняя часть внутреннего сегмента фоторецептора (IS), содержащая:
| Гипоотражающий | Нет | [27] [28] |
| 11 | Эллипсоидная зона (EZ) | Самая внешняя часть внутреннего сегмента фоторецептора (IS), заполненная митохондриями. | Очень гиперрефлексивный | Нет | [24] [29] [27] [25] [30] [31] |
| Соединение IS/OS или линия целостности фоторецепторов (PIL) | Фоторецептор, соединяющий реснички , соединяющие внутренний и внешний сегменты фоторецепторных клеток. | ||||
| 12 | Внешние сегменты фоторецепторов (ОС) | Внешние сегменты фоторецепторов (ОС) содержат диски, заполненные опсином — молекулой, поглощающей фотоны. | Гипоотражающий | [32] [25] | |
| 13 | Interdigitation zone (IZ) | Вершины клеток РПЭ , в которых заключена часть ОС колбочек. Плохо отличим от РПЭ. Ранее: «линия кончиков внешнего сегмента конуса» (COST). | Гиперотражающий | Нет | |
| 14 | РПЭ / Бруха комплекс | РПЭ Фагосомная зона | Очень гиперрефлексивный | Нет | [24] [25] |
| РПЭ Меланосомная зона | Гипоотражающий | ||||
| Зона митохондрий РПЭ + Соединение между РПЭ и мембраной Бруха | Очень гиперрефлексивный | ||||
| 15 | Хориокапилляры | Тонкий слой умеренной отражательной способности во внутренней сосудистой оболочке. | Нет | [25] | |
| 16 | Слой Саттлера | Толстый слой гиперрефлексивных профилей круглой или овальной формы с гипорефлексивными ядрами в средней части хориоидеи. | [25] | ||
| 17 | слой Галлера | Толстый слой гиперотражающих профилей овальной формы с гипорефлексивными ядрами во внешней сосудистой оболочке. | [25] | ||
| 18 | Хориоидально-склеральный переход | Зона внешней сосудистой оболочки с выраженным изменением текстуры, в которой крупные круглые или овоидные профили примыкают к однородная область переменной отражательной способности | [25] | ||
Развитие [ править ]
Развитие сетчатки начинается с формирования глазных полей, опосредованных белками SHH и SIX3 , с последующим развитием глазных пузырьков, регулируемых белками PAX6 и LHX2 . [33] Роль Pax6 в развитии глаз была элегантно продемонстрирована Walter Gehring и его коллегами, которые показали, что эктопическая экспрессия Pax6 может привести к образованию глаз на дрозофилы . усиках, крыльях и ногах [34] Зрительный пузырек дает начало трем структурам: нейральной сетчатке, пигментному эпителию сетчатки и зрительной ножке. Нейральная сетчатка содержит клетки-предшественники сетчатки (RPC), которые дают начало семи типам клеток сетчатки. Дифференцировка начинается с ганглиозных клеток сетчатки и заканчивается образованием глии Мюллера. [35] Хотя каждый тип клеток дифференцируется от RPC в последовательном порядке, существует значительное перекрытие во времени дифференцировки отдельных типов клеток. [33] Сигналы, определяющие судьбу дочерних клеток RPC, кодируются множеством семейств транскрипционных факторов, включая bHLH и гомеодоменные факторы. [36] [37]
Помимо определения судьбы клеток, в сетчатке существуют сигналы для определения дорсально-вентральной (DV) и носово-височной (NT) осей. Ось DV устанавливается вентрально-дорсальным градиентом VAX2 , тогда как ось NT координируется экспрессией факторов транскрипции forkhead FOXD1 и FOXG1 . Внутри сетчатки формируются дополнительные градиенты. [37] Такое пространственное распределение может помочь в правильном нацеливании на аксоны RGC, которые функционируют для создания ретинотопической карты. [33]
Кровоснабжение [ править ]
В этой статье есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|
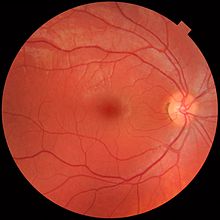
Сетчатка разделена на отдельные слои, каждый из которых содержит определенные типы клеток или клеточные компартменты. [38] которые имеют метаболизм с различными потребностями в питании. [39] Чтобы удовлетворить этим требованиям, глазная артерия раздваивается и снабжает сетчатку через две отдельные сосудистые сети: хориоидальную сеть, снабжающую сосудистую оболочку и внешнюю сетчатку, и сеть сетчатки, снабжающую внутренний слой сетчатки. [40]
Хотя перевернутая сетчатка у позвоночных кажется нелогичной, она необходима для правильного функционирования сетчатки. Слой фоторецепторов должен быть встроен в пигментный эпителий сетчатки (ПЭС), который выполняет как минимум семь жизненно важных функций: [41] Одним из наиболее очевидных способов является снабжение фоторецепторов кислородом и другими питательными веществами, необходимыми для их функционирования.
Энергетические требования
Энергетические потребности сетчатки даже больше, чем у мозга. [4] Это связано с дополнительной энергией, необходимой для постоянного обновления внешних сегментов фоторецепторов, 10% которых теряется ежедневно. [4] Потребности в энергии наиболее велики во время темновой адаптации, когда ее чувствительность наиболее повышена. [42] Сосудистая оболочка поставляет в сетчатку около 75% этих питательных веществ, а сосудистая сеть сетчатки — только 25%. [5]
Когда свет попадает на 11-цис-ретиналь (в дисках палочек и колбочек), 11-цис-ретиналь превращается в полностью транс-ретиналь, что затем вызывает изменения в опсинах. Однако внешние сегменты не регенерируют сетчатку обратно в цис-форму, как только она была изменена светом. Вместо этого сетчатка откачивается в окружающий РПЭ, где регенерируется и транспортируется обратно во внешние сегменты фоторецепторов. Эта функция рециркуляции RPE защищает фоторецепторы от фотоокислительного повреждения. [43] [44] и позволяет фоторецепторным клеткам иметь срок службы в течение десятилетий.
У птиц [ править ]
Сетчатка птиц лишена кровеносных сосудов, возможно, для того, чтобы обеспечить беспрепятственное прохождение света для формирования изображений, что обеспечивает лучшее разрешение. Таким образом, считается, что сетчатка птиц обеспечивает питание и снабжение кислородом специализированным органом, называемым «гребешком» или гребешком глаза , расположенным на слепом пятне или диске зрительного нерва. Этот орган чрезвычайно богат кровеносными сосудами и, как полагают, снабжает сетчатку птиц питанием и кислородом путем диффузии через стекловидное тело. Пектен очень богат активностью щелочной фосфатазы и поляризованными клетками в его мостиковой части – и то, и другое соответствует его секреторной роли. [45] Клетки гребешка заполнены темными гранулами меланина, которые, как предполагается, сохраняют этот орган в тепле за счет поглощения рассеянного света, падающего на гребень. Считается, что это увеличивает скорость метаболизма пектенов, тем самым экспортируя больше питательных молекул для удовлетворения строгих энергетических потребностей сетчатки в течение длительных периодов воздействия света. [46]
заболеваний Биометрическая и диагностика идентификация
Известно, что бифуркации и другие физические характеристики внутренней сосудистой сети сетчатки различаются у разных людей. [47] и эти индивидуальные отклонения использовались для биометрической идентификации и раннего выявления начала заболевания. Картирование сосудистых бифуркаций является одним из основных этапов биометрической идентификации. [48] Результаты такого анализа структуры кровеносных сосудов сетчатки можно оценить на основе фактических данных. [49] сосудистых бифуркаций изображений глазного дна, полученных из набора данных DRIVE. [50] Кроме того, были определены классы судов набора данных DRIVE, [51] Также доступен автоматизированный метод точного выделения этих бифуркаций. [52] Изменения кровообращения сетчатки наблюдаются с возрастом. [53] и воздействие загрязнения воздуха, [54] и может указывать на сердечно-сосудистые заболевания, такие как гипертония и атеросклероз. [55] [56] [57] Определение эквивалентной ширины артериол и венул вблизи диска зрительного нерва также является широко используемым методом выявления сердечно-сосудистых рисков. [58]
Функция [ править ]
Сетчатка преобразует оптическое изображение в нервные импульсы, начиная с закономерного возбуждения цветочувствительных пигментов палочек и колбочек — фоторецепторных клеток сетчатки . Возбуждение обрабатывается нервной системой и различными частями мозга, работающими параллельно, формируя в мозгу представление о внешней среде.
Колбочки реагируют на яркий свет и обеспечивают цветовое зрение высокого разрешения при дневном освещении (также называемое фотопическим зрением ). Реакция палочек насыщена при дневном освещении и не способствует формированию видения. Однако палочки реагируют на тусклый свет и обеспечивают монохроматическое зрение с более низким разрешением при очень низком уровне освещенности (так называемое скотопическое зрение ). Освещенность в большинстве офисов находится между этими двумя уровнями и называется мезопическим зрением . На мезопическом уровне освещенности и палочки, и колбочки активно передают информацию о закономерностях. Какой вклад вносит информация палочек в видение образов в этих обстоятельствах, неясно.
Реакция колбочек на свет различной длины называется их спектральной чувствительностью. В нормальном человеческом зрении спектральная чувствительность колбочки попадает в один из трех подтипов, часто называемых синим, зеленым и красным, но более точно известных как подтипы колбочек, чувствительных к короткой, средней и длинной волне. Отсутствие одного или нескольких подтипов колбочек приводит к тому, что у людей возникают нарушения цветового зрения или различные виды дальтонизма . Эти люди не слепы к объектам определенного цвета, но не способны различать цвета, которые могут различать люди с нормальным зрением. Люди обладают этим трехцветным зрением , в то время как у большинства других млекопитающих отсутствуют колбочки с чувствительным к красному пигменту, и поэтому у них хуже дихроматическое цветовое зрение. Однако у некоторых животных есть четыре спектральных подтипа, например, у форели к коротким, средним и длинным подтипам, сходным с человеком, добавляется ультрафиолетовая подгруппа. Некоторые рыбы также чувствительны к поляризации света.
В фоторецепторах воздействие света гиперполяризует мембрану в виде серии последовательных сдвигов. Внешний сегмент клетки содержит фотопигмент . Внутри клетки нормальный уровень циклического гуанозинмонофосфата (цГМФ) поддерживает канал Na+ открытым, и, таким образом, в состоянии покоя клетка деполяризуется. Фотон изомеризоваться заставляет сетчатку , связанную с белком-рецептором, в транс -ретиналь . Это заставляет рецептор активировать несколько G-белков . Это, в свою очередь, заставляет Ga-субъединицу белка активировать фосфодиэстеразу (PDE6), которая разрушает цГМФ, что приводит к закрытию ионных каналов, управляемых циклическими нуклеотидами Na+ (CNG). Таким образом, клетка гиперполяризуется. Количество высвобождаемого нейромедиатора уменьшается при ярком свете и увеличивается при падении уровня освещенности. Фактический фотопигмент обесцвечивается при ярком свете и заменяется только в результате химического процесса, поэтому при переходе от яркого света к темноте глазу может потребоваться до тридцати минут, чтобы достичь полной чувствительности.
При таком возбуждении светом фоторецептор посылает пропорциональный синаптический ответ биполярным клеткам, которые, в свою очередь, сигнализируют ганглиозным клеткам сетчатки . Фоторецепторы также перекрестно связаны горизонтальными клетками и амакриновыми клетками , которые модифицируют синаптический сигнал до того, как он достигнет ганглиозных клеток, при этом нервные сигналы смешиваются и объединяются. Из нервных клеток сетчатки только ганглиозные клетки сетчатки и несколько амакриновых клеток создают потенциалы действия .
В ганглиозных клетках сетчатки наблюдаются два типа ответа в зависимости от рецептивного поля клетки. Рецептивные поля ганглиозных клеток сетчатки состоят из центральной, примерно круглой области, где свет оказывает одно влияние на возбуждение клетки, и кольцевой области, где свет оказывает противоположный эффект. В ON-клетках увеличение интенсивности света в центре рецептивного поля приводит к увеличению скорости стрельбы. В выключенных ячейках он уменьшается. В линейной модели этот профиль отклика хорошо описывается разностью гауссиан и является основой алгоритмов обнаружения границ . Помимо этого простого различия, ганглиозные клетки также различаются по хроматической чувствительности и типу пространственной суммации. Клетки, демонстрирующие линейное пространственное суммирование, называются X-клетками (также называемыми парвоцеллюлярными, P или карликовыми ганглиозными клетками), а клетки, демонстрирующие нелинейное суммирование, называются Y-клетками (также называемыми магноцеллюлярными, M или парасольными ганглиозными клетками сетчатки), хотя соответствие между Х- и Y-клетки (в сетчатке кошки) и Р- и М-клетки (в сетчатке приматов) не так просты, как когда-то казалось.
При передаче зрительных сигналов в мозг, зрительный путь , сетчатка вертикально разделена на две части: височную (ближе к виску) и носовую (ближе к носу) половину. Аксоны носовой половины пересекают мозг в перекресте зрительных нервов и соединяются с аксонами височной половины другого глаза, прежде чем перейти в латеральное коленчатое тело .
Хотя в сетчатке имеется более 130 миллионов рецепторов, в зрительном нерве всего около 1,2 миллиона волокон (аксонов). Таким образом, внутри сетчатки выполняется большой объем предварительной обработки. Ямка . дает наиболее точную информацию Несмотря на то, что зрительный нерв занимает около 0,01% поля зрения (менее 2° угла зрения ), около 10% аксонов зрительного нерва сосредоточены в ямке. Предел разрешения фовеа был определен примерно в 10 000 точек. Информационная емкость оценивается в 500 000 бит в секунду (подробнее о битах см. в теории информации ) без цвета или около 600 000 бит в секунду, включая цвет. [59]
Пространственное кодирование [ править ]

Когда сетчатка посылает в мозг нервные импульсы, представляющие изображение, она пространственно кодирует (сжимает) эти импульсы, чтобы они соответствовали ограниченным возможностям зрительного нерва. Сжатие необходимо, поскольку фоторецепторных клеток в 100 раз больше, чем ганглиозных клеток . Это осуществляется путем « декорреляции », которая осуществляется «структурами центр-окружение», реализуемыми биполярными и ганглиозными клетками.
В сетчатке существует два типа центрально-окружающих структур – центральные и внецентровые. У центральных элементов есть положительно взвешенный центр и отрицательно взвешенный объем. Оффцентры – это полная противоположность. Положительное воздействие более известно как возбуждающее , а отрицательное – как тормозящее .
Эти центрально-окружающие структуры не являются физически очевидными в том смысле, что их нельзя увидеть, окрашивая образцы ткани и исследуя анатомию сетчатки. Структуры центр-окружение логичны (т.е. математически абстрактны) в том смысле, что они зависят от силы связей между биполярными и ганглиозными клетками. Считается, что сила связи между клетками обусловлена количеством и типами ионных каналов, встроенных в синапсы между биполярными и ганглиозными клетками.
Структуры центр-объем математически эквивалентны алгоритмам обнаружения краев , используемым компьютерными программистами для выделения или улучшения краев на цифровой фотографии. Таким образом, сетчатка выполняет операции с импульсами, представляющими изображение, чтобы улучшить края объектов в поле зрения. Например, на изображении собаки, кошки и автомобиля именно края этих объектов содержат больше всего информации. Чтобы высшие функции мозга (или компьютера, если уж на то пошло) могли извлекать и классифицировать такие объекты, как собака и кошка, сетчатка является первым шагом к разделению различных объектов на сцене.
Например, следующая матрица лежит в основе компьютерного алгоритма , реализующего обнаружение границ. Эта матрица является компьютерным эквивалентом структуры центр-объем. В этом примере каждый блок (элемент) внутри этой матрицы будет подключен к одному фоторецептору. Фоторецептор в центре — это обрабатываемый рецептор тока. Центральный фоторецептор умножается на весовой коэффициент +1. Окружающие фоторецепторы являются «ближайшими соседями» к центру и умножаются на значение -1/8. Наконец вычисляется сумма всех девяти этих элементов. Это суммирование повторяется для каждого фоторецептора на изображении путем смещения влево до конца строки, а затем вниз до следующей строки.
| -1/8 | -1/8 | -1/8 |
| -1/8 | +1 | -1/8 |
| -1/8 | -1/8 | -1/8 |
Общая сумма этой матрицы равна нулю, если все входные сигналы от девяти фоторецепторов имеют одинаковое значение. Нулевой результат означает, что изображение было однородным (не меняющимся) в пределах этого небольшого участка. Отрицательные или положительные суммы означают, что изображение менялось в пределах этого небольшого участка из девяти фоторецепторов.
Приведенная выше матрица является лишь приближением к тому, что на самом деле происходит внутри сетчатки. Отличия заключаются в следующем:
- Приведенный выше пример называется «сбалансированным». Термин «сбалансированный» означает, что сумма отрицательных весов равна сумме положительных весов, поэтому они полностью компенсируются. Ганглиозные клетки сетчатки почти никогда не бывают идеально сбалансированными.
- Стол имеет квадратную форму, а центрально-окружающие структуры сетчатки имеют круглую форму.
- Нейроны работают на цепочках спайков, нервных клеток движущихся по аксонам . Компьютеры оперируют одним числом с плавающей запятой , которое по существу является постоянным для каждого входного пикселя . (Компьютерный пиксель по сути является эквивалентом биологического фоторецептора.)
- Сетчатка выполняет все эти вычисления параллельно, в то время как компьютер обрабатывает каждый пиксель по отдельности. Сетчатка не производит повторяющихся суммаций и сдвигов, как компьютер.
- Наконец, горизонтальные и амакриновые клетки , но здесь они не представлены. значительную роль в этом процессе играют
Вот пример входного изображения и то, как обнаружение краев может его изменить.

Как только изображение пространственно кодируется структурами центр-окружение, сигнал отправляется по зрительному нерву (через аксоны ганглиозных клеток) через перекрест зрительных нервов в LGN ( латеральное коленчатое ядро ). Точная функция LGN в настоящее время неизвестна. Затем выходные данные LGN отправляются в заднюю часть мозга. В частности, выходной сигнал LGN «излучается» в первичную зрительную кору V1 .
Упрощенный поток сигналов: Фоторецепторы → Биполярный → Ганглий → Хиазма → LGN → Кора V1.

Клиническое значение
Существует множество наследственных и приобретенных заболеваний или нарушений, которые могут поражать сетчатку. Некоторые из них включают в себя:
- Пигментный ретинит — группа генетических заболеваний, поражающих сетчатку и вызывающих потерю ночного и периферического зрения.
- Дегенерация желтого пятна описывает группу заболеваний, характеризующихся потерей центрального зрения из-за гибели или повреждения клеток макулы .
- Колбочково-стержневая дистрофия (CORD) описывает ряд заболеваний, при которых потеря зрения вызвана повреждением колбочек и /или палочек сетчатки.
- При отслоении сетчатки сетчатка отделяется от задней части глазного яблока. Игнипунктура – устаревший метод лечения. Термин «отслойка сетчатки» используется для описания отделения нейросенсорной сетчатки от пигментного эпителия сетчатки . [60] Существует несколько современных методов лечения отслойки сетчатки: пневматическая ретинопексия , склеральная пряжка , криотерапия , лазерная фотокоагуляция pars plana и витрэктомия .
- И гипертония , и сахарный диабет могут вызвать повреждение крошечных кровеносных сосудов, снабжающих сетчатку, что приводит к гипертонической ретинопатии и диабетической ретинопатии .
- Ретинобластома – это рак сетчатки.
- Заболевания сетчатки у собак включают дисплазию сетчатки , прогрессирующую атрофию сетчатки и внезапную приобретенную дегенерацию сетчатки .
- Липемия сетчатки — это белое появление сетчатки, которое может возникать вследствие отложения липидов при дефиците липопротеинлипазы .
- Отслойка сетчатки . Нервная сетчатка иногда отслаивается от пигментного эпителия. В некоторых случаях причиной такого отслоения является травма глазного яблока, из-за которой жидкость или кровь скапливаются между нервной сетчаткой и пигментным эпителием. Отслойка иногда вызывается контрактурой тонких коллагеновых фибрилл в стекловидном теле, которые тянут участки сетчатки внутрь глазного яблока. [23]
- Куриная слепота : Куриная слепота возникает у любого человека с тяжелым дефицитом витамина А. Причина этого в том, что без витамина А количество образующихся ретиналя и родопсина сильно снижается. Это состояние называется куриной слепотой, поскольку количество света в ночное время слишком мало, чтобы обеспечить адекватное зрение людям с дефицитом витамина А. [18]
Кроме того, сетчатку называют «окном» в мозг и тело, поскольку отклонения, обнаруженные при исследовании сетчатки, могут выявить как неврологические, так и системные заболевания. [61]
Диагностика [ править ]
Для диагностики заболеваний и нарушений, поражающих сетчатку, доступен ряд различных инструментов. Офтальмоскопия и фотография глазного дна уже давно используются для исследования сетчатки. Недавно адаптивная оптика была использована для получения изображений отдельных палочек и колбочек в сетчатке живого человека, а компания из Шотландии разработала технологию, которая позволяет врачам наблюдать всю сетчатку без какого-либо дискомфорта для пациентов. [62]
Электроретинограмма . используется для неинвазивного измерения электрической активности сетчатки, на которую влияют некоторые заболевания Относительно новая технология, которая в настоящее время становится широко доступной, — это оптическая когерентная томография (ОКТ). Этот неинвазивный метод позволяет получить объемную 3D- поперечного сечения высокого разрешения томограмму или томограмму тонких структур сетчатки гистологического качества. Анализ сосудов сетчатки — это неинвазивный метод исследования мелких артерий и вен сетчатки, который позволяет сделать выводы о морфологии и функции мелких сосудов в других частях тела человека. Установлено, что он является предиктором сердечно-сосудистых заболеваний. [63] и, согласно исследованию, опубликованному в 2019 году, похоже, имеет потенциал для раннего выявления болезни Альцгеймера. [64]
Лечение [ править ]
Лечение зависит от характера заболевания или расстройства.
лечения Общие методы
Ниже приведены распространенные методы лечения заболеваний сетчатки:
- Интравитреальные препараты, такие как анти-VEGF или кортикостероидные препараты.
- Витреоретинальная хирургия
- Использование пищевых добавок
- Модификация системных факторов риска заболеваний сетчатки
лечения Необычные методы
Генная терапия сетчатки
Генная терапия обещает стать потенциальным средством лечения широкого спектра заболеваний сетчатки. Это предполагает использование неинфекционного вируса для доставки гена в часть сетчатки. Векторы рекомбинантного аденоассоциированного вируса (rAAV) обладают рядом особенностей, которые делают их идеально подходящими для генной терапии сетчатки, включая отсутствие патогенности, минимальную иммуногенность и способность стабильно и эффективно трансдуцировать постмитотические клетки. [65] Векторы rAAV все чаще используются из-за их способности опосредовать эффективную трансдукцию пигментного эпителия сетчатки (RPE), фоторецепторных клеток и ганглиозных клеток сетчатки . Каждый тип клеток может быть специфически нацелен путем выбора подходящей комбинации серотипа AAV , промотора и места внутриглазной инъекции.
Несколько клинических испытаний уже сообщили о положительных результатах использования rAAV для лечения врожденного амавроза Лебера , показывая, что терапия была безопасной и эффективной. [66] [67] Серьезных нежелательных явлений отмечено не было, и во всех трех исследованиях у пациентов наблюдалось улучшение зрительных функций, что оценивалось рядом методов. Используемые методы различались в трех исследованиях, но включали оба функциональных метода, таких как определение остроты зрения. [67] [68] [69] и функциональная мобильность [68] [69] [70] а также объективные показатели, которые менее подвержены предвзятости, такие как способность зрачка реагировать на свет. [66] [71] и улучшения функциональной МРТ. [72] Улучшения сохранялись в долгосрочной перспективе, и пациенты продолжали чувствовать себя хорошо даже спустя более чем 1,5 года. [66] [67]
Уникальная архитектура сетчатки и ее относительно защищенная иммунная среда способствуют этому процессу. [73] Плотные соединения , образующие гемато-ретинальный барьер, отделяют субретинальное пространство от кровоснабжения, тем самым защищая его от микробов и большинства иммуноопосредованных повреждений, а также повышая его способность реагировать на векторную терапию. Чрезвычайно разделенная анатомия глаза облегчает точную доставку суспензий терапевтических векторов к конкретным тканям под прямой визуализацией с использованием микрохирургических методов. [74] В защищенной среде сетчатки векторы AAV способны поддерживать высокий уровень экспрессии трансгена в пигментном эпителии сетчатки (RPE), фоторецепторах или ганглиозных клетках в течение длительных периодов времени после однократной обработки. Кроме того, глаз и зрительную систему можно регулярно и легко контролировать на предмет зрительных функций и структурных изменений сетчатки после инъекций с помощью неинвазивных передовых технологий, таких как острота зрения, контрастная чувствительность , глазного дна автофлуоресценция (FAF), адаптированные к темноте пороги зрения. , диаметр сосудов, пупиллометрия, электроретинография (ЭРГ), мультифокальная ЭРГ и оптическая когерентная томография (ОКТ). [75]
Эта стратегия эффективна против ряда изученных заболеваний сетчатки, включая неоваскулярные заболевания, которые являются признаками возрастной макулярной дегенерации , диабетической ретинопатии и ретинопатии недоношенных . Поскольку регуляция васкуляризации в зрелой сетчатке включает баланс между эндогенными положительными факторами роста , такими как фактор роста эндотелия сосудов (VEGF), и ингибиторами ангиогенеза , такими как фактор пигментного эпителия ( PEDF ), rAAV-опосредованная экспрессия PEDF, Было показано, что ангиостатин и растворимый рецептор VEGF sFlt-1, которые являются антиангиогенными белками, уменьшают аберрантное образование сосудов на животных моделях. [76] Поскольку специфическая генная терапия не может быть легко использована для лечения значительной части пациентов с дистрофией сетчатки, существует большой интерес к разработке более широко применимой терапии факторами выживания. Нейротрофические факторы обладают способностью модулировать рост нейронов во время развития, поддерживая существующие клетки и обеспечивая восстановление поврежденных популяций нейронов в глазу. AAV, кодирующий нейротрофические факторы, такие как члены семейства факторов роста фибробластов (FGF) и GDNF, либо защищают фоторецепторы от апоптоза, либо замедляют гибель клеток. [76]
Трансплантация органов трансплантации Были попытки сетчатки, но без особого успеха. В Массачусетском технологическом институте , Университете Южной Калифорнии, Университете RWTH Ахена и Университете Нового Южного Уэльса разрабатывается «искусственная сетчатка»: имплантат, который будет обходить фоторецепторы сетчатки и напрямую стимулировать прикрепленные нервные клетки сигналами. с цифровой камеры.
История [ править ]
Около 300 г. до н. э . Герофил идентифицировал сетчатку по вскрытиям трупных глаз. Он назвал его паутинным слоем из-за сходства с паутиной и сетчатым из-за сходства с кастинговой сетью. Термин «паутинный» стал обозначать слой вокруг мозга; термин ретиформ стал относиться к сетчатке . [77]
Между 1011 и 1021 годами нашей эры Ибн аль-Хайсам опубликовал многочисленные эксперименты, демонстрирующие, что зрение возникает за счет света, отражающегося от объектов в глаз. Это согласуется с теорией интромиссии и противоречит теории эмиссии , теории, согласно которой зрение возникает за счет лучей, испускаемых глазами. Однако Ибн Аль-Хайсам решил, что сетчатка не может быть ответственной за начало зрения, поскольку изображение, формируемое на ней, является перевернутым. Вместо этого он решил, что все должно начинаться с поверхности линзы. [78]
В 1604 году Иоганн Кеплер разработал оптику глаза и пришел к выводу, что сетчатка должна быть местом начала зрения. Он предоставил другим ученым возможность согласовать перевернутое изображение на сетчатке с нашим восприятием мира как вертикального. [79]
В 1894 году Сантьяго Рамон-и-Кахаль опубликовал первую важную характеристику нейронов сетчатки в книге Retina der Wirbelthiere ( «Сетчатка позвоночных »). [80]
Джордж Уолд , Холдан Кеффер Хартлайн и Рагнар Гранит получили Нобелевскую премию по физиологии и медицине 1967 года за научные исследования сетчатки. [81]
Недавнее исследование Пенсильванского университета подсчитало, что приблизительная пропускная способность сетчатки человека составляет 8,75 мегабит в секунду, тогда как морской свинки составляет 875 килобит в секунду. скорость передачи данных через сетчатку [82]
Макларен и Пирсон и коллеги из Университетского колледжа Лондона и офтальмологической больницы Мурфилдс в Лондоне в 2006 году показали, что фоторецепторные клетки можно успешно трансплантировать в сетчатку мыши, если донорские клетки находятся на критической стадии развития. [83] Недавно Адер и его коллеги из Дублина показали с помощью электронного микроскопа, что трансплантированные фоторецепторы образуют синаптические связи. [84]
В 2012 году Себастьян Сын и его лаборатория в Массачусетском технологическом институте запустили EyeWire , гражданскую онлайн- игру, в которой игроки отслеживают нейроны сетчатки. [85] Цели проекта EyeWire — идентифицировать конкретные типы клеток среди известных широких классов клеток сетчатки и составить карту связей между нейронами сетчатки, что поможет определить, как работает зрение. [86] [87]
Дополнительные изображения [ править ]
 Структуры глаза, обозначенные
Структуры глаза, обозначенные Другой вид глаза и его структур с надписью
Другой вид глаза и его структур с надписью Иллюстрация изображения, «видимого» сетчаткой, независимо от обработки зрительного нерва и полосатой коры.
Иллюстрация изображения, «видимого» сетчаткой, независимо от обработки зрительного нерва и полосатой коры.



См. также [ править ]
- Аденоассоциированный вирус и генная терапия сетчатки человека
- Чарльз Шепенс – «отец современной хирургии сетчатки»
- Эволюция глаза
- Дуплекс сетчатки
- Сканирование сетчатки
- Окклюзия вен сетчатки
- Список вариантов ксантом, связанных с подтипами гиперлипопротеинемии
- Родопсин
- Стойкость зрения
Ссылки [ править ]
- ^ Дж, Краузе Уильям (2005). Основная гистология человека Краузе для студентов-медиков . Бока-Ратон, Флорида: Universal Publishers. ISBN 978-1-58112-468-2 .
- ^ Jump up to: Перейти обратно: а б с д и ж г «Сенсорная рецепция: Человеческое зрение: Структура и функции человеческого глаза», том. 27, Британская энциклопедия, 1987 г.
- ^ «Исследователи из Пенсильванского университета подсчитали, как много глаз сообщает мозгу» (пресс-релиз). ПЕНН Медицина . 26 июля 2006 г. Архивировано из оригинала 11 марта 2013 г. . Проверено 22 апреля 2022 г.
- ^ Jump up to: Перейти обратно: а б с Вьегас, Филипе О.; Нойхаусс, Стефан CF (2021). «Метаболический ландшафт для поддержания целостности и функции сетчатки» . Границы молекулярной нейронауки . 14 . дои : 10.3389/fnmol.2021.656000 . ISSN 1662-5099 . ПМК 8081888 . ПМИД 33935647 .
- ^ Jump up to: Перейти обратно: а б Колб, Хельга (1995). «Простая анатомия сетчатки» . Вебвидение . ПМИД 21413391 . Проверено 1 января 2018 г.
- ^ Колб, Хельга. «Фоторецепторы» . Вебвидение . Проверено 11 января 2018 г.
- ^ Франц К., Гроше Дж., Скачков С.Н., Шинкингер С., Фойя С., Шильд Д., Укерманн О., Трэвис К., Райхенбах А., Гак Дж. (2007). «Клетки Мюллера — это живые оптические волокна в сетчатке позвоночных» . Учеб. Натл. акад. наук. США . 104 (20): 8287–8292. Бибкод : 2007PNAS..104.8287F . дои : 10.1073/pnas.0611180104 . ЧВК 1895942 . ПМИД 17485670 .
- ^ Бейкер, Оливер (23 апреля 2010 г.). «В центре внимания: клетки глаза как световоды». Фокус физического обзора . Том. 25, нет. 15. дои : 10.1103/physrevfocus.25.15 .
- ^ Jump up to: Перейти обратно: а б Брингманн А, Сюрбе С, Гёрнер К, Кача Дж, Франке М, Видеманн П, Райхенбах А (2018). «Ямка примата: строение, функции и развитие». Прога Retin Eye Res . 66 : 49–84. doi : 10.1016/j.preteyeres.2018.03.006 . ПМИД 29609042 . S2CID 5045660 .
- ^ Сперлинг, Л.; Хаббард, Р. (1 февраля 1975 г.). «Ретинохром кальмара» . Журнал общей физиологии . 65 (2): 235–251. дои : 10.1085/jgp.65.2.235 . ISSN 0022-1295 . ПМК 2214869 . ПМИД 235007 .
- ^ «Как пауки видят мир – Австралийский музей» . www.австралийский.музей . Архивировано из оригинала 12 сентября 2017 года . Проверено 5 декабря 2017 г.
- ^ Дражен, Дж. К.; Да, Дж.; Фридман Дж.; Кондон, Н. (июнь 2011 г.). «Метаболизм и ферментативная активность миксины мелководных и глубоководных вод Тихого океана». Сравнительная биохимия и физиология. Часть A: Молекулярная и интегративная физиология . 159 (2): 182–187. дои : 10.1016/j.cbpa.2011.02.018 . ПМИД 21356325 .
- ^ Лабин, А.М.; Рыбак, Э.Н. (16 апреля 2010 г.). «Глиальные клетки сетчатки повышают остроту зрения человека» . Письма о физических отзывах . 104 (15): 158102. Бибкод : 2010PhRvL.104o8102L . doi : 10.1103/PhysRevLett.104.158102 . ПМИД 20482021 .
- ^ Основы видения. Архивировано 3 декабря 2013 года в Wayback Machine , Брайан А. Ванделл.
- ^ «Ретинальная туника» . Региональный колледж ветеринарной медицины Вирджинии-Мэриленда . Архивировано из оригинала 18 мая 2007 года.
- ^ Гольдберг А.Ф., Мориц О.Л., Уильямс Д.С. (2016). «Молекулярные основы архитектуры внешнего сегмента фоторецепторов» . Прога Retin Eye Res . 55 : 52–81. doi : 10.1016/j.preteyeres.2016.05.003 . ПМК 5112118 . ПМИД 27260426 .
- ^ Аршавский В.Ю., Бернс М.Е. (2012). «Передача сигналов фоторецепторов: поддержка зрения в широком диапазоне интенсивности света» . J Биол Хим . 287 (3): 1620–1626. дои : 10.1074/jbc.R111.305243 . ПМЦ 3265842 . ПМИД 22074925 .
- ^ Jump up to: Перейти обратно: а б Физиология Гайтона и Холла . п. 612.
- ^ Воробей-младший, Хикс Д., Хэмел С.П. (2010). «Пигментный эпителий сетчатки в норме и патологии» . Карр Мол Мед . 10 (9): 802–823. дои : 10.2174/156652410793937813 . ПМК 4120883 . ПМИД 21091424 .
- ^ Летелье Х., Боволента П., Мартинес-Моралес Х.Р. (2017). «Пигментированный эпителий, яркий партнер в борьбе с дегенерацией фоторецепторов». Дж. Нейрогенет . 31 (4): 203–215. дои : 10.1080/01677063.2017.1395876 . ПМИД 29113536 . S2CID 1351539 .
- ^ Шепард, Гордон (2004). Синаптическая организация мозга . Нью-Йорк: Издательство Оксфордского университета. стр. 217–225 . ISBN 978-0-19-515956-1 .
- ^ Ромер, Альфред Шервуд; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. п. 465. ИСБН 978-0-03-910284-5 .
- ^ Jump up to: Перейти обратно: а б Физиология Гайтона и Холла . п. 609.
- ^ Jump up to: Перейти обратно: а б с Куэнка, Николас; Ортуньо-Лизаран, Изабель; Пинилла, Изабель (март 2018 г.). «Клеточная характеристика ОКТ и внешних полос сетчатки с использованием специфических иммуногистохимических маркеров и клинические последствия» (PDF) . Офтальмология . 125 (3): 407–422. дои : 10.1016/j.ophtha.2017.09.016 . hdl : 10045/74474 . ПМИД 29037595 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л м н тот п д Ставеренги, Джованни; Садда, Шринивас; Чакраварти, Уша; Спейд, Ричард Ф. (2014). «Предлагаемый словарь анатомических ориентиров в нормальной спектральной когерентной томографии заднего сегмента». Офтальмология . 121 (8): 1572–1578. дои : 10.1016/j.ophtha.2014.02.023 . ПМИД 24755005 .
- ^ Мейер, Карстен Х.; Саксена, Сандип; Садда, Шринивас Р. (2017). Оптическая когерентная томография спектральной области при заболеваниях желтого пятна . Нью-Дели: Спрингер. ISBN 978-8132236108 . OCLC 964379175 .
- ^ Jump up to: Перейти обратно: а б Хильдебранд, Йоран Дариус; Филдер, Алистер Р. (2011). «Анатомия и физиология сетчатки». Детская сетчатка . Шпрингер, Берлин, Гейдельберг. стр. 39–65. дои : 10.1007/978-3-642-12041-1_2 . ISBN 978-3642120404 .
- ^ Тургут, Бурак; Университет Фырат; Медицина, Школа; Офтальмологическое отделение; Элязыг; Турция (2017). «Прошлое и настоящее Терминология для структур сетчатки и хориоидеи в оптической когерентной томографии» . Европейский офтальмологический обзор . 11 (1): 59. doi : 10.17925/eor.2017.11.01.59 .
- ^ «Наружные слои сетчатки как предикторы потери зрения» . Обзор офтальмологии .
- ^ «Азбука ОКТ» . Обзор оптометрии .
- ^ Шерман, Дж (июнь 2009 г.). «Линия целостности фоторецепторов, соединяющая слой нервных волокон, является ключом к клиническому диагнозу». Оптометрия . 80 (6): 277–278. дои : 10.1016/j.optm.2008.12.006 . ПМИД 19465337 .
- ^ Бостон, Марко А. Бонини Фильо, доктор медицинских наук, и Андре Дж. Уиткин, доктор медицинских наук. «Наружные слои сетчатки как предикторы потери зрения» . Проверено 7 апреля 2018 г.
{{cite news}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: Перейти обратно: а б с Хивнер, В; Певный, Л. (1 декабря 2012 г.). «Развитие глаз и ретиногенез» . Перспективы Колд-Спринг-Харбор в биологии . 4 (12): а008391. doi : 10.1101/cshperspect.a008391 . ПМЦ 3504437 . ПМИД 23071378 .
- ^ Гальдер, Г; Каллаертс, П; Геринг, WJ (24 марта 1995 г.). «Индукция эктопических глаз путем целевой экспрессии безглазого гена у дрозофилы». Наука . 267 (5205): 1788–1792. Бибкод : 1995Sci...267.1788H . дои : 10.1126/science.7892602 . ПМИД 7892602 .
- ^ Чепко, Конни (сентябрь 2014 г.). «Совершенно разные клетки-предшественники сетчатки производят определенные типы потомства». Обзоры природы Неврология . 15 (9): 615–627. дои : 10.1038/nrn3767 . ISSN 1471-003X . ПМИД 25096185 . S2CID 15038502 .
- ^ Хатакеяма, Дж; Кагеяма, Р. (февраль 2004 г.). «Определение судьбы клеток сетчатки и факторы bHLH». Семинары по клеточной биологии и биологии развития . 15 (1): 83–89. дои : 10.1016/j.semcdb.2003.09.005 . ПМИД 15036211 .
- ^ Jump up to: Перейти обратно: а б Ло Джудис, Квентин; Леле, Мэрион; Ла Манно, Джоэле; Фабр, Пьер Ж. (1 сентября 2019 г.). «Одноклеточная транскрипционная логика спецификации клеточной судьбы и руководство аксонами в нейронах сетчатки раннего рождения» . Разработка . 146 (17): dev178103. дои : 10.1242/dev.178103 . ISSN 0950-1991 . ПМИД 31399471 .
- ^ Ремингтон, Ли Энн (2012). Клиническая анатомия и физиология зрительной системы (3-е изд.). Сент-Луис: Эльзевир/Баттерворт-Хайнеманн. ISBN 978-1-4377-1926-0 . OCLC 745905738 .
- ^ Ю, ДЯ; Ю, ПК; Крингл, С.Дж.; Канг, Миннесота; Вс, RU (май 2014 г.). «Функциональные и морфологические характеристики сосудов сетчатки и хориоидеи». Прогресс в исследованиях сетчатки и глаз . 40 : 53–93. doi : 10.1016/j.preteyeres.2014.02.001 . ПМИД 24583621 . S2CID 21312546 .
- ^ Киль, Джеффри В. Анатомия . Морган и Клейпул Науки о жизни. Архивировано из оригинала 5 декабря 2017 года . Проверено 17 апреля 2017 г.
- ^ Штраус, Олаф. «Пигментный эпителий сетчатки» . Вебвидение . Проверено 1 января 2018 г.
- ^ Кайнежад, Пардис; Тахцидис, Илиас; Шивапрасад, Собха; Джеффри, Глен (2023). «Наблюдение за дыханием сетчатки человека в реальном времени и замедлением митохондриального дыхания с возрастом» . Научные отчеты . 13 (1): 6445. Бибкод : 2023НатСР..13.6445К . дои : 10.1038/s41598-023-32897-7 . ISSN 2045-2322 . ПМЦ 10119193 . ПМИД 37081065 .
- ^ «СВЕТОВОЕ ПОВРЕЖДЕНИЕ СЕТЧАТКИ» . photobiology.info . Проверено 23 февраля 2023 г.
- ^ «Схематическое изображение отделения диска и извлечения фагосом в пигментные эпителиальные клетки» . Архивировано из оригинала 21 сентября 2012 года . Проверено 22 апреля 2022 г.
- ^ Бава С.Р.; ЯшРой РЦ (1972). «Влияние темновой и световой адаптации на сетчатку и гребешок курицы» . Экспериментальное исследование глаз . 13 (1): 92–97. дои : 10.1016/0014-4835(72)90129-7 . ПМИД 5060117 . Архивировано из оригинала 9 октября 2014 года.
- ^ Бава, СР; ЯшРой, RC (1974). «Строение и функции гребешка грифа» . Клетки Ткани Органы . 89 (3): 473–480. doi : 10.1159/000144308 (неактивен 12 июня 2024 г.). ПМИД 4428954 . Архивировано из оригинала 14 июля 2015 года.
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на июнь 2024 г. ( ссылка ) - ^ Шерман, Т (1981). «О соединении больших судов с малыми – смысл закона Мюррея» . Журнал общей физиологии . 78 (4): 431–453. дои : 10.1085/jgp.78.4.431 . ПМК 2228620 . ПМИД 7288393 .
- ^ Аззопарди Г.; Петков Н. (2011). «Обнаружение бифуркаций сосудов сетчатки с помощью обучаемых фильтров типа V4». Компьютерный анализ изображений и закономерностей (PDF) . Конспекты лекций по информатике. Том. 6854. стр. 451–459. дои : 10.1007/978-3-642-23672-3_55 . ISBN 978-3-642-23671-6 . Архивировано (PDF) из оригинала 9 августа 2017 года.
- ^ «Изображения глазного дна сетчатки – основная истина о сосудистых бифуркациях и пересечениях» . Университет Гронингена . Проверено 20 апреля 2018 г.
- ^ «ДРАЙВ: Цифровые изображения сетчатки для извлечения сосудов» . Изображение Института наук, Утрехтский университет . Архивировано из оригинала 6 августа 2020 года . Проверено 20 апреля 2018 г.
- ^ Куреши, Т.А.; Хабиб, М.; Хантер, А.; Аль-Дири, Б. (июнь 2013 г.). «Помеченный вручную эталонный показатель артерий/вен для набора данных DRIVE». Материалы 26-го Международного симпозиума IEEE по компьютерным медицинским системам . стр. 485–488. дои : 10.1109/cbms.2013.6627847 . ISBN 978-1-4799-1053-3 . S2CID 7705121 .
- ^ Куреши, Т.А.; Хантер, А.; Аль-Дири, Б. (июнь 2014 г.). «Байесовский подход к локальной конфигурации соединений сетчатки». Конференция IEEE 2014 по компьютерному зрению и распознаванию образов . стр. 3105–3110. CiteSeerX 10.1.1.1026.949 . дои : 10.1109/cvpr.2014.397 . ISBN 978-1-4799-5118-5 . S2CID 14654500 .
- ^ Адар С.Д., Кляйн Р., Кляйн Б.Е., Шпиро А.А., Котч М.Ф., Вонг Т.Ю. и др. (2010). «Загрязнение воздуха и микроциркуляторное русло: перекрестная оценка изображений сетчатки in vivo в рамках популяционного многоэтнического исследования атеросклероза (MESA)» . ПЛОС Мед . 7 (11): e1000372. дои : 10.1371/journal.pmed.1000372 . ПМЦ 2994677 . ПМИД 21152417 .
- ^ Лоувис, Тиджс; Панис, Люк Инт; Кичински, Михал; Бовер, Патрик Де; Наврот, Тим С. (2013). «Реакция микрососудистой системы сетчатки на кратковременные изменения загрязнения воздуха твердыми частицами у здоровых взрослых» . Перспективы гигиены окружающей среды . 121 (9): 1011–1016. дои : 10.1289/ehp.1205721 . ПМК 3764070 . ПМИД 23777785 .
- ^ Цо, Марк ОМ; Джампол, Ли М. (1982). «Патофизиология гипертонической ретинопатии». Офтальмология . 89 (10): 1132–1145. дои : 10.1016/s0161-6420(82)34663-1 . ПМИД 7155524 .
- ^ Чепмен, Н.; Делль'омо, Г.; Сартини, MS; Витт, Н.; Хьюз, А.; Том, С.; Педринелли, Р. (1 августа 2002 г.). «Заболевания периферических сосудов связаны с аномальным соотношением диаметров артериол в бифуркациях сетчатки человека». Клиническая наука . 103 (2): 111–116. дои : 10.1042/cs1030111 . ISSN 0143-5221 . ПМИД 12149100 .
- ^ Паттон, Н.; Аслам, Т.; МакГилливрей, Т.; Дорогой, И.; Диллон, Б.; Эйкельбум, Р.; Йогесан, К.; Констебль, И. (2006). «Анализ изображений сетчатки: концепции, приложения и потенциал». Прогресс в исследованиях сетчатки и глаз . 25 (1): 99–127. doi : 10.1016/j.preteyeres.2005.07.001 . ПМИД 16154379 . S2CID 7434103 .
- ^ Вонг Т., Кнудтсон, М.Д., Кляйн Р., Кляйн Б.Е., Мейер С.М., Хаббард Л.Д. (2004). «Компьютерное измерение диаметра сосудов сетчатки в исследовании глаз Бивер-Дэм: методология, корреляция между глазами и влияние аномалий рефракции». Офтальмология . 111 (6): 1183–1190. дои : 10.1016/j.ophtha.2003.09.039 . ПМИД 15177969 .
- ^ Чен, Джанглин; Крэнтон, Уэйн; Фин, Марк (2016). Справочник по технологии визуального отображения (2-е изд.). Чам, Швейцария: Springer. ISBN 9783319143460 . ОСЛК 962009228 .
- ^ Сетчатка (4-е изд.). Филадельфия: Эльзевир/Мосби. 2006. С. 2013–2015. ISBN 978-0-323-02598-0 . OCLC 62034580 .
- ^ Фрит, Пегги; Мехта, Арпан Р. (ноябрь 2021 г.). «Сетчатка как окно в мозг» . Ланцет Неврология . 20 (11): 892. doi : 10.1016/S1474-4422(21)00332-X . ПМК 7611980 . ПМИД 34687632 .
- ^ Взгляд в будущее. Архивировано 12 февраля 2012 г. в Wayback Machine Ingenia , март 2007 г.
- ^ Зейдельманн, С.Б.; и др. (1 ноября 2016 г.). «Калибры сосудов сетчатки в прогнозировании долгосрочных сердечно-сосудистых исходов» . Тираж . 134 (18): 1328–1338. doi : 10.1161/CIRCULATIONAHA.116.023425 . ПМК 5219936 . ПМИД 27682886 .
- ^ Керкес, Г; и др. (11 января 2019 г.). «Функциональные и морфологические изменения сосудов сетчатки при болезни Альцгеймера и легких когнитивных нарушениях» . Научные отчеты . 9 (63): 63. Бибкод : 2019НацСР...9...63Q . дои : 10.1038/s41598-018-37271-6 . ПМК 6329813 . ПМИД 30635610 .
- ^ Динкулеску Астра; Глушакова Людмила; Сок-Хон Мин; Хаусвирт Уильям В. (2005). «Генная терапия аденоассоциированных вирусов при заболеваниях сетчатки». Генная терапия человека . 16 (6): 649–663. дои : 10.1089/hum.2005.16.649 . ПМИД 15960597 .
- ^ Jump up to: Перейти обратно: а б с Сидеджиян А.В.; Хаусвирт WW; Алеман Т.С.; Каушал С.; Шварц С.Б.; Бойе С.Л.; Виндзорский EAM; и др. (2009). «Генная терапия RPE65 человека при врожденном амаврозе Лебера: сохранение ранних улучшений зрения и безопасность в течение 1 года» . Генная терапия человека . 20 (9): 999–1004. дои : 10.1089/hum.2009.086 . ПМЦ 2829287 . ПМИД 19583479 .
- ^ Jump up to: Перейти обратно: а б с Симонелли Ф.; Магуайр AM; Теста Ф.; Пирс Э.А.; Мингоцци Ф.; Бенничелли Дж.Л.; Росси С.; и др. (2010). «Генная терапия врожденного амавроза Лебера безопасна и эффективна в течение 1,5 лет после введения вектора» . Молекулярная терапия . 18 (3): 643–650. дои : 10.1038/mt.2009.277 . ПМЦ 2839440 . ПМИД 19953081 .
- ^ Jump up to: Перейти обратно: а б Магуайр AM; Симонелли Ф.; Пирс Э.А.; Пью ЭН; Мингоцци Ф.; Бенничелли Дж.; Банфи С.; и др. (2008). «Безопасность и эффективность переноса генов при врожденном амаврозе Лебера» . Медицинский журнал Новой Англии . 358 (21): 2240–2248. doi : 10.1056/NEJMoa0802315 . ПМЦ 2829748 . ПМИД 18441370 .
- ^ Jump up to: Перейти обратно: а б Магуайр AM; Высокий КА; Ауриккио А.; Райт Дж. Ф.; Пирс Э.А.; Теста Ф.; Мингоцци Ф.; и др. (2009). «Возрастные эффекты генной терапии RPE65 при врожденном амаврозе Лебера: исследование фазы 1 с увеличением дозы» . Ланцет . 374 (9701): 1597–1605. дои : 10.1016/S0140-6736(09)61836-5 . ПМЦ 4492302 . ПМИД 19854499 .
- ^ Бейнбридж JWB; Смит А.Дж.; Баркер СС; Робби С.; Хендерсон Р.; Балагган К.; Вишванатан А.; и др. (2008). «Влияние генной терапии на зрительные функции при врожденном амаврозе Лебера» (PDF) . Медицинский журнал Новой Англии . 358 (21): 2231–2239. CiteSeerX 10.1.1.574.4003 . doi : 10.1056/NEJMoa0802268 . hdl : 10261/271174 . ПМИД 18441371 . Архивировано (PDF) из оригинала 11 августа 2017 года.
- ^ Хаусвирт WW; Алеман Т.С.; Каушал С.; Сидеджиян А.В.; Шварц С.Б.; Ван Л.; Конлон Ти Джей; и др. (2008). «Лечение врожденного амавроза Лебера, вызванного мутациями RPE65, путем субретинальной инъекции вектора гена аденоассоциированного вируса: краткосрочные результаты исследования фазы I» . Генная терапия человека . 19 (10): 979–990. дои : 10.1089/hum.2008.107 . ПМЦ 2940541 . ПМИД 18774912 .
- ^ Аштари М.; Цыковски Л.Л.; Монро Дж. Ф.; Маршалл К.А.; Чунг, округ Колумбия; Ауриккио А.; Симонелли Ф.; и др. (2011). «Зрительная кора человека реагирует на восстановление функции сетчатки, опосредованное генной терапией» . Журнал клинических исследований . 121 (6): 2160–2168. дои : 10.1172/JCI57377 . ПМК 3104779 . ПМИД 21606598 .
- ^ Беннетт Дж (2003). «Иммунный ответ после внутриглазной доставки рекомбинантных вирусных векторов» . Генная терапия . 10 (11): 977–982. дои : 10.1038/sj.gt.3302030 . ПМИД 12756418 .
- ^ Кураче Энрико М.; Ауриккио Альберто (2008). «Универсальность векторов AAV для переноса генов сетчатки» . Исследование зрения . 48 (3): 353–359. дои : 10.1016/j.visres.2007.07.027 . ПМИД 17923143 . S2CID 9926758 .
- ^ ден Холландер, Аннеке И.; Ропман, Рональд; Кенекуп, Роберт К.; Кремерс, Франс П.М. (2008). «Врожденный амавроз Лебера: гены, белки и механизмы заболевания». Прогресс в исследованиях сетчатки и глаз . 27 (4): 391–419. doi : 10.1016/j.preteyeres.2008.05.003 . ПМИД 18632300 . S2CID 30202286 .
- ^ Jump up to: Перейти обратно: а б Роллинг, Ф. (2004). «Перенос генов, опосредованный рекомбинантным AAV, в сетчатку: перспективы генной терапии» . Генная терапия . 11 (С1): С26–С32. дои : 10.1038/sj.gt.3302366 . ISSN 0969-7128 . ПМИД 15454954 .
- ^ Добсон, Дж. Ф. (март 1925 г.). «Герофил Александрийский» . Труды Королевского медицинского общества . 18 (Sect_Hist_Med): 19–32. дои : 10.1177/003591572501801704 . ISSN 0035-9157 . ПМК 2201994 . ПМИД 19984605 .
- ^ Сабра, AI (Ред.). (1011–1021/1989). Оптика Ибн Аль-Хайсама: Книги I-III: О прямом зрении (А.И. Сабра, Пер.). Институт Варбурга.
- ^ Фишман, Р.С. (1973). «Открытие Кеплером изображения на сетчатке» . Архив офтальмологии . 89 (1): 59–61. doi : 10.1001/archopht.1973.01000040061014 . ПМИД 4567856 . Проверено 23 февраля 2023 г.
- ^ «Сантьяго Рамон и Кахаль - Биографический» . www.nobelprize.org . Архивировано из оригинала 6 октября 2015 года . Проверено 20 октября 2015 г.
- ^ «Нобелпремия.орг» . nobelprize.org . Архивировано из оригинала 30 июня 2017 года . Проверено 5 декабря 2017 г.
- ^ Рейли, Майкл. «Расчет скорости зрения» . Новый учёный . Архивировано из оригинала 31 мая 2015 года . Проверено 5 декабря 2017 г.
- ^ Макларен, RE; Пирсон, РА; МакНил, А; и др. (ноябрь 2006 г.). «Восстановление сетчатки путем трансплантации предшественников фоторецепторов» (PDF) . Природа . 444 (7116): 203–207. Бибкод : 2006Natur.444..203M . дои : 10.1038/nature05161 . hdl : 2027.42/62596 . ПМИД 17093405 . S2CID 4415311 .
- ^ Барч, У.; Орияхель, В.; Кенна, ПФ; Линке, С.; Ричард, Г.; Петровиц, Б.; Хамфрис, П.; Фаррар, Дж.Дж.; Адер, М. (2008). «Клетки сетчатки интегрируются во внешний ядерный слой и дифференцируются в зрелые фоторецепторы после субретинальной трансплантации взрослым мышам». Экспериментальное исследование глаз . 86 (4): 691–700. дои : 10.1016/j.exer.2008.01.018 . ПМИД 18329018 .
- ^ «О: EyeWire» . Архивировано из оригинала 13 февраля 2012 года . Проверено 26 марта 2012 г.
- ^ «Сетчатка << EyeWire» . Архивировано из оригинала 24 марта 2012 года . Проверено 27 марта 2012 г.
- ^ «ОкоВайр» . Архивировано из оригинала 24 апреля 2012 года . Проверено 27 марта 2012 г.
Дальнейшее чтение [ править ]
- С. Рамон-и-Кахаль , Гистология нервной системы человека и позвоночных , Малоин, Париж, 1911.
- Родик Р.В. (1965). «Количественный анализ реакции ганглиозных клеток сетчатки кошки на зрительные стимулы». Видение Рез . 5 (11): 583–601. дои : 10.1016/0042-6989(65)90033-7 . ПМИД 5862581 .
- Ванделл, Брайан А. (1995). Основы видения . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-853-7 .
- Вассле Х., Бойкот BB (1991). «Функциональная архитектура сетчатки млекопитающих». Физиол. Преподобный . 71 (2): 447–480. дои : 10.1152/physrev.1991.71.2.447 . ПМИД 2006220 .
- Шульц Х.Л., Гетц Т., Кашкото Дж., Вебер Б.Х. (2004). «Ретином – определение эталонного транскриптома сетчатки/пигментного эпителия сетчатки взрослых млекопитающих» . BMC Genomics (о транскриптоме цвета глаз). 5 (1): 50. дои : 10.1186/1471-2164-5-50 . ПМК 512282 . ПМИД 15283859 .
- Даулинг, Джон (2007). «Ретина» . Схоларпедия . 2 (12): 3487. Бибкод : 2007SchpJ...2.3487D . дои : 10.4249/scholarpedia.3487 .
Внешние ссылки [ править ]
- Гистология глаза , под редакцией Уильяма Краузе, кафедра патологии и анатомии, Медицинский факультет Университета Миссури.
- Глаз, мозг и зрение - онлайн-книга - Дэвид Хьюбел
- Колб Х., Фернандес Э. и Нельсон Р. (2003). Webvision: Нейронная организация сетчатки позвоночных . Солт-Лейк-Сити, Юта: Глазной центр Джона Морана, Университет Юты . Проверено 22 июля 2014 г.
- Изображение слоев сетчатки. Неврология, 2-е издание, Национальная медицинская библиотека США.
- Семинары Джереми Натанса: «Сетчатка позвоночных: структура, функции и эволюция»
- Сетчатка – клеточно-центрированная база данных
- Гистологическое изображение: 07901loa - Система обучения гистологии в Бостонском университете.
- Энциклопедия MedlinePlus : 002291
