Амакриновая клетка
Вы можете помочь дополнить эту статью текстом, переведенным из соответствующей статьи на чешском языке . (Апрель 2012 г.) Нажмите [показать], чтобы просмотреть важные инструкции по переводу. |
| Амакриновая клетка | |
|---|---|
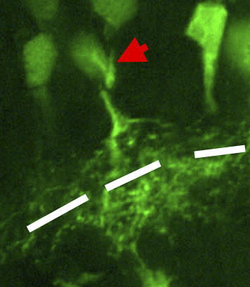 Клетки сетчатки Xenopus , окрашенные на cdk2/циклин2, красная стрелка указывает на амакриновые клетки. IPL показан в черно-белом цвете. | |
| Подробности | |
| Расположение | Внутренний ядерный слой и слой ганглиозных клеток сетчатки . |
| Форма | Варьируется |
| Функция | тормозные или нейромодулирующие интернейроны |
| Нейромедиатор | гамма-аминомасляная кислота , глицин , DA или 5-HT |
| Пресинаптические связи | Биполярные клетки |
| Постсинаптические связи | Биполярные клетки и ганглиозные клетки |
| Идентификаторы | |
| МеШ | D025042 |
| НейроЛекс ID | нифекст_36 |
| ФМА | 67766 |
| Анатомические термины нейроанатомии | |

В анатомии глаза . клетки представляют интернейроны сетчатки собой амакриновые [1] Они названы от греческого a– «нон» makr – «длинный» и in – «волокно» из-за коротких нейронных отростков . Амакриновые клетки являются тормозными нейронами и проецируют свои дендритные ветви на внутренний плексиформный слой (IPL), они взаимодействуют с ганглиозными клетками сетчатки и биполярными клетками или с ними обоими. [2]
Структура
[ редактировать ]Амакриновые клетки действуют во внутреннем плексиформном слое (IPL), втором синаптическом слое сетчатки, где биполярные клетки и ганглиозные клетки сетчатки образуют синапсы . Существует по крайней мере 33 различных подтипа амакриновых клеток, основанных только на их морфологии и стратификации дендритов. Как и горизонтальные клетки , амакриновые клетки работают латерально, но в то время как горизонтальные клетки связаны с выходными данными палочек и колбочек, амакриновые клетки влияют на выходные данные биполярных клеток и часто более специализированы. Каждый тип амакриновых клеток высвобождает один или несколько нейромедиаторов там, где он соединяется с другими клетками. [2]
Их часто классифицируют по ширине поля связи, в каком слое (слоях) IPL они находятся, а также по типу нейромедиатора. Большинство из них являются ингибирующими, используя гамма-аминомасляную кислоту или глицин в качестве нейромедиаторов .
Типы
[ редактировать ]Как упоминалось выше, существует несколько разных способов разделить множество различных типов амакриновых клеток на подтипы.
ГАМКергическая, глицинергическая или ни одна из них :Амакриновые клетки могут быть ГАМКергическими , глицинергическими или ни одним из них, в зависимости от того, какой тормозной нейромедиатор они экспрессируют (ГАМК, глицин или ни один из них). ГАМКергические амакриновые клетки обычно представляют собой амакриновые клетки с широким полем поля и обнаруживаются в слое ганглиозных клеток (GCL) и внутреннем ядерном слое (INL). Одним из типов ГАМКергических амакриновых клеток, который достаточно хорошо изучен, являются амакриновые клетки звездообразования . Эти амакриновые клетки обычно характеризуются экспрессией холин-ацетилтрансферазы или ChAT и, как известно, играют роль в избирательности направления и обнаружении направленного движения. [2] Ацетилхолин также высвобождается из этих амакриновых клеток, но его функция до конца не изучена. [3] Другим подтипом ГАМКергических амакриновых клеток являются дофаминергические. Все они экспрессируют ТГ, и эти амакриновые клетки модулируют адаптацию к свету и циркадный ритм. [2] Это широко распространенные амакриновые клетки, которые диффузно выделяют дофамин, одновременно выделяя ГАМК и осуществляя все нормальные синаптические выделения. [3] Было отмечено множество других делений ГАМКергических амакриновых клеток, но перечисленные выше являются одними из наиболее широко исследованных и обсуждаемых.
Глицинергические амакриновые клетки не так подробно охарактеризованы, как ГАМКергические амакриновые клетки. Однако все глицинергические амакриновые клетки отмечены транспортером глицина GlyT1 . Одной из очень хорошо охарактеризованных глицинергических амакриновых клеток являются амакриновые клетки AII . Эти клетки присутствуют в INL. [2] Одной из важных функций амакриновых клеток AII является то, что они захватывают входной сигнал от палочковидных биполярных клеток и перераспределяют его в конусные биполярные клетки, используя синаптические окончания конусных биполярных клеток в качестве адаптеров. [4]
Около 15% амакриновых клеток не являются ни ГАМКергическими, ни глицинергическими. [2] Эти амакриновые клетки иногда называют амакриновыми клетками nGnG, и считается, что факторы транскрипции, действующие на предшественников, решают судьбу амакриновых клеток. Было обнаружено, что одним из факторов транскрипции, который избирательно экспрессируется в амакриновых клетках nGnG, является Neurod6. [5]
Длина дендритных беседок :В зависимости от длины и распространения дендритных ветвей амакриновые клетки можно разделить на амакриновые клетки с узким полем (около 70 микрометров в диаметре), амакриновые клетки со средним полем (около 170 микрометров в диаметре) и амакриновые клетки с широким полем (около 350 микрометров в диаметре). . [2] Эта разная длина обеспечивает разные специфические функции, которые могут выполнять амакриновые клетки. Амакриновые клетки с узким полем поля обеспечивают вертикальную связь между различными уровнями сетчатки. Они также помогают в создании функциональных субъединиц в рецептивном поле ганглиозных клеток. Эти амакриновые клетки с узким полем поля и их перекрытие в этих субъединицах могут позволить определенным ганглиозным клеткам обнаруживать небольшие движения очень маленького пятна в поле зрения. Одним из типов клеток с узким полем, который делает это, является амакриновая клетка звездообразования. [3]
Амакриновые клетки среднего поля также способствуют вертикальной коммуникации в клетках сетчатки, но большая часть их общей функции до сих пор неизвестна. Поскольку размер их дендритных стволов очень похож на размер ганглиозных клеток, они могут размывать края поля зрения ганглиозных клеток. Точно так же амакриновые клетки с широким полем зрения трудно исследовать и даже обнаружить, поскольку они охватывают всю сетчатку и их не так много. Однако, учитывая их размер, одной из их основных функций является боковая связь внутри слоя, хотя некоторые из них также осуществляют вертикальную связь между слоями. [3]
Организация
[ редактировать ]Амакриновые клетки и другие интернейронные клетки сетчатки с меньшей вероятностью окажутся рядом с соседями одного и того же подтипа, чем это могло бы произойти случайно, что приводит к образованию «зон отчуждения», разделяющих их. Мозаичное расположение обеспечивает механизм равномерного распределения каждого типа клеток по сетчатке, гарантируя, что все части поля зрения имеют доступ к полному набору элементов обработки. [6] Трансмембранные белки MEGF10 и MEGF11 играют решающую роль в формировании мозаики амакриновых клеток звездообразования и горизонтальных клеток у мышей. [7]
Функция
[ редактировать ]Во многих случаях подтип амакриновых клеток говорит о их функции (форма определяет функцию), но можно выделить некоторые специфические функции амакриновых клеток сетчатки.
- Перехват ганглиозных клеток сетчатки и/или биполярных клеток в IPL [2]
- Создайте функциональные субъединицы в рецептивных полях многих ганглиозных клеток.
- Способствуют вертикальной связи внутри слоев сетчатки.
- Выполняют паракринные функции, такие как высвобождение дофамина, ацетилхолина и оксида азота. [3] [8]
- Благодаря своим связям с другими клетками сетчатки в синапсах и высвобождению нейротрансмиттеров они способствуют обнаружению направленного движения, модулируют адаптацию к свету и циркадный ритм. [2] и контролировать высокую чувствительность скотопического зрения посредством связей с биполярными клетками палочек и колбочек. [4]
Нам еще многое предстоит узнать о различных функциях различных амакриновых клеток. Считается, что амакриновые клетки с обширными дендритными деревьями вносят вклад в тормозное окружение за счет обратной связи как на уровне биполярных клеток, так и на уровне ганглиозных клеток. Считается, что в этой роли они дополняют действие горизонтальных клеток.
Другие формы амакриновых клеток, вероятно, играют модулирующую роль, позволяя регулировать чувствительность фотопического и скотопического зрения . Амакриновая клетка AII является медиатором сигналов от палочек в скотопических условиях. [4]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Колб, Х; Нельсон, Р; Фернандес, Э (1995). «Роль амакриновых клеток». ПМИД 21413397 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Перейти обратно: а б с д и ж г час я Баласубраманян, Р.; Ган, Л. (2014). «Развитие амакриновых клеток сетчатки и их дендритная стратификация» . Текущие отчеты офтальмологии . 2 (3): 100–106. дои : 10.1007/s40135-014-0048-2 . ПМЦ 4142557 . ПМИД 25170430 .
- ^ Перейти обратно: а б с д и Масланд, Р.Х. (2012). «Задачи амакриновых клеток» . Визуальная нейронаука . 29 (1): 3–9. дои : 10.1017/s0952523811000344 . ПМЦ 3652807 . ПМИД 22416289 .
- ^ Перейти обратно: а б с Марк, RE; Андерсон-младший; Джонс, BW; Сигулинский, К.Л.; Лауритцен, Дж. С. (2014). «Коннектом амакриновых клеток AII: узел плотной сети» . Границы в нейронных цепях . 8 : 104. doi : 10.3389/fncir.2014.00104 . ПМЦ 4154443 . ПМИД 25237297 .
- ^ Кей, Дж. Н.; Войнеску, ЧП; Чу, МВт; Санес, младший (2011). «Экспрессия Neurod6 определяет новые подтипы амакриновых клеток сетчатки и регулирует их судьбу» . Природная неврология . 14 (8): 965–72. дои : 10.1038/nn.2859 . ПМК 3144989 . ПМИД 21743471 .
- ^ Вассле, Х.; Риман, HJ (22 марта 1978 г.). «Мозаика нервных клеток сетчатки млекопитающих». Труды Королевского общества B: Биологические науки . 200 (1141): 441–461. Бибкод : 1978RSPSB.200..441W . дои : 10.1098/rspb.1978.0026 . ПМИД 26058 . S2CID 28724457 .
- ^ Кей, Джереми Н.; Чу, Моника В.; Санес, Джошуа Р. (март 2012 г.). «MEGF10 и MEGF11 опосредуют гомотипические взаимодействия, необходимые для мозаичного расположения нейронов сетчатки» . Природа . 483 (7390): 465–9. Бибкод : 2012Natur.483..465K . дои : 10.1038/nature10877 . ПМК 3310952 . ПМИД 22407321 .
- ^ Джейкоби, Джейсон (5 декабря 2018 г.). «Саморегулирующаяся сеть щелевых соединений амакриновых клеток контролирует высвобождение оксида азота в сетчатке» . Нейрон . 100 (5): 1149–1162. дои : 10.1016/j.neuron.2018.09.047 . ПМК 6317889 . ПМИД 30482690 .
- Николлс, Джон Г.; А. Роберт Мартин; Пол А. Фукс; Дэвид А. Браун; Мэтью Э. Даймонд; Дэвид А. Вайсблат (2012). От нейрона к мозгу, пятое издание . Бостон, Массачусетс: Sinauer Associates, Inc. ISBN 978-0-87893-609-0 .
- Масланд Р.Х. (2001). «Основной план сетчатки». Нат. Нейроски . 4 (9): 877–86. дои : 10.1038/nn0901-877 . ПМИД 11528418 . S2CID 205429773 .
Внешние ссылки
[ редактировать ]- Статья Webvision об амакриновых клетках
- Амакриновые клетки Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- Книга «Глазной мозг и зрение». Архивировано 5 октября 2020 г. в Wayback Machine Hubel D (1988). «Глазной мозг и зрение», вся книга доступна в Интернете.
- Поиск NIF — амакриновые клетки , заархивировано 5 марта 2016 г. в Wayback Machine через информационную структуру нейробиологии.
