Серотонин
 | |
| Клинические данные | |
|---|---|
| Другие имена | 5-HT, 5-гидрокситриптамин, энтерамин, тромбоцитин, 3-(β-аминоэтил)-5-гидроксииндол, тромботонин |
| Физиологические данные | |
| Исходные ткани | ядра шва , энтерохромаффинные клетки |
| Целевые ткани | общесистемный |
| Рецепторы | 5-НТ 1 , 5-НТ 2 , 5-НТ 3 , 5-НТ 4 , 5-НТ 5 , 5-НТ 6 , 5-НТ 7 |
| Agonists | Indirectly: SSRIs, MAOIs |
| Precursor | 5-HTP |
| Biosynthesis | Aromatic L-amino acid decarboxylase |
| Metabolism | MAO |
| Identifiers | |
| CAS Number | |
| PubChem CID | |
| IUPHAR/BPS | |
| ChemSpider | |
| KEGG | |
| PDB ligand | |
| CompTox Dashboard (EPA) | |
| ECHA InfoCard | 100.000.054 |
 | |
| Names | |
|---|---|
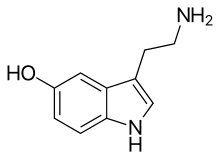
| IUPAC name 5-Hydroxytryptamine | |
| Preferred IUPAC name 3-(2-Aminoethyl)-1H-indol-5-ol | |
| Other names 5-Hydroxytryptamine, 5-HT, Enteramine; Thrombocytin, 3-(β-Aminoethyl)-5-hydroxyindole, 3-(2-Aminoethyl)indol-5-ol, Thrombotonin | |
| Identifiers | |
3D model (JSmol) | |
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| ECHA InfoCard | 100.000.054 |
| KEGG | |
| MeSH | Serotonin |
PubChem CID | |
| UNII | |
CompTox Dashboard (EPA) | |
| Properties | |
| C10H12N2O | |
| Molar mass | 176.215 g/mol |
| Appearance | White powder |
| Melting point | 167.7 °C (333.9 °F; 440.8 K) 121–122 °C (ligroin)[3] |
| Boiling point | 416 ± 30 °C (at 760 Torr)[1] |
| slightly soluble | |
| Acidity (pKa) | 10.16 in water at 23.5 °C[2] |
| 2.98 D | |
| Hazards | |
| Lethal dose or concentration (LD, LC): | |
LD50 (median dose) | 750 mg/kg (subcutaneous, rat),[4] 4500 mg/kg (intraperitoneal, rat),[5] 60 mg/kg (oral, rat) |
| Safety data sheet (SDS) | External MSDS |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
Серотонин ( / ˌ s ɛr ə ˈ toʊ n ɪ n , ə ˌ s ɪər - / ) [6] [7] [8] или 5-гидрокситриптамин ( 5-НТ ) — моноаминовый нейромедиатор . Его биологическая функция сложна и затрагивает разнообразные функции, включая настроение , познание , вознаграждение , обучение , память и многочисленные физиологические процессы, такие как рвота и вазоконстрикция . [9]
Серотонин вырабатывается в центральной нервной системе (ЦНС), особенно в мозга ствола ядрах шва кожи , клетках Меркеля , легочных нейроэндокринных клетках языка и клетках вкусовых рецепторов . Примерно 90% серотонина, который производит человеческий организм , находится в желудочно-кишечного тракта , энтерохромаффинных клетках где он регулирует перистальтику кишечника. [10] [11] [12] Кроме того, он сохраняется в тромбоцитах и высвобождается при возбуждении и сужении сосудов, где затем действует как агонист других тромбоцитов. [13] Около 8% обнаруживается в тромбоцитах и 1–2% в ЦНС. [14]
The serotonin is secreted luminally and basolaterally, which leads to increased serotonin uptake by circulating platelets and activation after stimulation, which gives increased stimulation of myenteric neurons and gastrointestinal motility.[15] Остальная часть синтезируется в серотонинергических нейронах ЦНС, где выполняет различные функции, включая регуляцию настроения, аппетита и сна .
Serotonin secreted from the enterochromaffin cells eventually finds its way out of tissues into the blood. There, it is actively taken up by blood platelets, which store it. When the platelets bind to a clot, they release serotonin, where it can serve as a vasoconstrictor or a vasodilator while regulating hemostasis and blood clotting. In high concentrations, serotonin acts as a vasoconstrictor by contracting endothelial smooth muscle directly or by potentiating the effects of other vasoconstrictors (e.g. angiotensin II and norepinephrine). The vasoconstrictive property is mostly seen in pathologic states affecting the endothelium – such as atherosclerosis or chronic hypertension. In normal physiologic states, vasodilation occurs through the serotonin mediated release of nitric oxide from endothelial cells, and the inhibition of release of norepinephrine from adrenergic nerves.[16] Serotonin is also a growth factor for some types of cells, which may give it a role in wound healing. There are various serotonin receptors.
Biochemically, the indoleamine molecule derives from the amino acid tryptophan. Serotonin is metabolized mainly to 5-hydroxyindoleacetic acid, chiefly by the liver.Several classes of antidepressants, such as selective serotonin reuptake inhibitors (SSRIs) and serotonin–norepinephrine reuptake inhibitors (SNRIs), interfere with the normal reabsorption of serotonin after it is done with the transmission of the signal, therefore augmenting the neurotransmitter levels in the synapses.
Besides mammals, serotonin is found in all bilateral animals including worms and insects,[17] as well as in fungi and in plants.[18] Serotonin's presence in insect venoms and plant spines serves to cause pain, which is a side-effect of serotonin injection.[19][20] Serotonin is produced by pathogenic amoebae, causing diarrhea in the human gut.[21] Its widespread presence in many seeds and fruits may serve to stimulate the digestive tract into expelling the seeds.[22][failed verification]
Molecular structure[edit]
Biochemically, the indoleamine molecule derives from the amino acid tryptophan, via the (rate-limiting) hydroxylation of the 5 position on the ring (forming the intermediate 5-hydroxytryptophan), and then decarboxylation to produce serotonin.[23] Preferable conformations are defined via ethylamine chain, resulting in six different conformations.[24]
Crystal structure[edit]
Serotonin crystallizes in P212121 chiral space group forming different hydrogen-bonding interactions between serotonin molecules via N-H...O and O-H...N intermolecular bonds.[25] Serotonin also forms several salts, including pharmaceutical formulation of serotonin adipate.[26]
Biological role[edit]
Serotonin is involved in numerous physiological processes,[27] including sleep, thermoregulation, learning and memory, pain, (social) behavior,[28] sexual activity, feeding, motor activity, neural development,[29] and biological rhythms.[30] In less complex animals, such as some invertebrates, serotonin regulates feeding and other processes.[31] In plants serotonin synthesis seems to be associated with stress signals.[18][32] Despite its longstanding prominence in pharmaceutical advertising, the claim that low serotonin levels cause depression is not supported by scientific evidence.[33][34][35]
Cellular effects[edit]
Serotonin primarily acts through its receptors and its effects depend on which cells and tissues express these receptors.[30]
Metabolism involves first oxidation by monoamine oxidase to the corresponding aldehyde. The rate-limiting step is hydride transfer from serotonin to the flavin cofactor.[36] There follows oxidation by aldehyde dehydrogenase to 5-hydroxyindoleacetic acid (5-HIAA), the indole acetic-acid derivative. The latter is then excreted by the kidneys.
Receptors[edit]
The 5-HT receptors, the receptors for serotonin, are located on the cell membrane of nerve cells and other cell types in animals, and mediate the effects of serotonin as the endogenous ligand and of a broad range of pharmaceutical and psychedelic drugs. Except for the 5-HT3 receptor, a ligand-gated ion channel, all other 5-HT receptors are G-protein-coupled receptors (also called seven-transmembrane, or heptahelical receptors) that activate an intracellular second messenger cascade.[37]
Termination[edit]
Serotonergic action is terminated primarily via uptake of 5-HT from the synapse. This is accomplished through the specific monoamine transporter for 5-HT, SERT, on the presynaptic neuron. Various agents can inhibit 5-HT reuptake, including cocaine, dextromethorphan (an antitussive), tricyclic antidepressants and selective serotonin reuptake inhibitors (SSRIs). A 2006 study found that a significant portion of 5-HT's synaptic clearance is due to the selective activity of the plasma membrane monoamine transporter (PMAT) which actively transports the molecule across the membrane and back into the presynaptic cell.[38]
In contrast to the high affinity of SERT, the PMAT has been identified as a low-affinity transporter, with an apparent Km of 114 micromoles/l for serotonin, which is approximately 230 times higher than that of SERT. However, the PMAT, despite its relatively low serotonergic affinity, has a considerably higher transport "capacity" than SERT, "resulting in roughly comparable uptake efficiencies to SERT ... in heterologous expression systems."[38] The study also suggests that the administration of SSRIs such as fluoxetine and sertraline may be associated with an inhibitory effect on PMAT activity when used at higher than normal dosages (IC50 test values used in trials were 3–4 fold higher than typical prescriptive dosage).
Serotonylation[edit]
Serotonin can also signal through a nonreceptor mechanism called serotonylation, in which serotonin modifies proteins.[39] This process underlies serotonin's effects upon platelet-forming cells (thrombocytes) in which it links to the modification of signaling enzymes called GTPases that then trigger the release of vesicle contents by exocytosis.[40] A similar process underlies the pancreatic release of insulin.[39]
The effects of serotonin upon vascular smooth muscle tone – the biological function after which serotonin was originally named – depend upon the serotonylation of proteins involved in the contractile apparatus of muscle cells.[41]
| Receptor | Ki (nM)[42] | Receptor function[Note 1] |
|---|---|---|
| 5-HT1 receptor family signals via Gi/o inhibition of adenylyl cyclase. | ||
| 5-HT1A | 3.17 | Memory[vague] (agonists ↓); learning[vague] (agonists ↓); anxiety (agonists ↓); depression (agonists ↓); positive, negative, and cognitive symptoms of schizophrenia (partial agonists ↓); analgesia (agonists ↑); aggression (agonists ↓); dopamine release in the prefrontal cortex (agonists ↑); serotonin release and synthesis (agonists ↓) |
| 5-HT1B | 4.32 | Vasoconstriction (agonists ↑); aggression (agonists ↓); bone mass (↓). Serotonin autoreceptor. |
| 5-HT1D | 5.03 | Vasoconstriction (agonists ↑) |
| 5-HT1E | 7.53 | |
| 5-HT1F | 10 | |
| 5-HT2 receptor family signals via Gq activation of phospholipase C. | ||
| 5-HT2A | 11.55 | Psychedelia (agonists ↑); depression (agonists & antagonists ↓); anxiety (antagonists ↓); positive and negative symptoms of schizophrenia (antagonists ↓); norepinephrine release from the locus coeruleus (antagonists ↑); glutamate release in the prefrontal cortex (agonists ↑); dopamine in the prefrontal cortex (agonists ↑);[43] urinary bladder contractions (agonists ↑)[44] |
| 5-HT2B | 8.71 | Cardiovascular functioning (agonists increase risk of pulmonary hypertension), empathy (via von Economo neurons[45]) |
| 5-HT2C | 5.02 | Dopamine release into the mesocorticolimbic pathway (agonists ↓); acetylcholine release in the prefrontal cortex (agonists ↑); dopaminergic and noradrenergic activity in the frontal cortex (antagonists ↑);[46] appetite (agonists ↓); antipsychotic effects (agonists ↑); antidepressant effects (agonists & antagonists ↑) |
| Other 5-HT receptors | ||
| 5-HT3 | 593 | Emesis (agonists ↑); anxiolysis (antagonists ↑). |
| 5-HT4 | 125.89 | Movement of food across the GI tract (agonists ↑); memory & learning (agonists ↑); antidepressant effects (agonists ↑). Signalling via Gαs activation of adenylyl cyclase. |
| 5-HT5A | 251.2 | Memory consolidation.[47] Signals via Gi/o inhibition of adenylyl cyclase. |
| 5-HT6 | 98.41 | Cognition (antagonists ↑); antidepressant effects (agonists & antagonists ↑); anxiogenic effects (antagonists ↑[48]). Gs signalling via activating adenylyl cyclase. |
| 5-HT7 | 8.11 | Cognition (antagonists ↑); antidepressant effects (antagonists ↑). Acts by Gs signalling via activating adenylyl cyclase. |
Nervous system[edit]

The neurons of the raphe nuclei are the principal source of 5-HT release in the brain.[49] There are nine raphe nuclei, designated B1–B9, which contain the majority of serotonin-containing neurons (some scientists chose to group the nuclei raphes lineares into one nucleus), all of which are located along the midline of the brainstem, and centered on the reticular formation.[50][51] Axons from the neurons of the raphe nuclei form a neurotransmitter system reaching almost every part of the central nervous system. Axons of neurons in the lower raphe nuclei terminate in the cerebellum and spinal cord, while the axons of the higher nuclei spread out in the entire brain.
Ultrastructure and function[edit]
The serotonin nuclei may also be divided into two main groups, the rostral and caudal containing three and four nuclei respectively. The rostral group consists of the caudal linear nuclei (B8), the dorsal raphe nuclei (B6 and B7) and the median raphe nuclei (B5, B8 and B9), that project into multiple cortical and subcortical structures. The caudal group consists of the nucleus raphe magnus (B3), raphe obscurus nucleus (B2), raphe pallidus nucleus (B1), and lateral medullary reticular formation, that project into the brainstem.[52]
The serotonergic pathway is involved in sensorimotor function, with pathways projecting both into cortical (Dorsal and Median Raphe Nuclei), subcortical, and spinal areas involved in motor activity. Pharmacological manipulation suggests that serotonergic activity increases with motor activity while firing rates of serotonergic neurons increase with intense visual stimuli. Animal models suggest that kainate signaling negatively regulates serotonin actions in the retina, with possible implications for the control of the visual system.[53] The descending projections form a pathway that inhibits pain called the "descending inhibitory pathway" that may be relevant to a disorder such as fibromyalgia, migraine, and other pain disorders, and the efficacy of antidepressants in them.[54]
Serotonergic projections from the caudal nuclei are involved in regulating mood and emotion, and hypo-[55] or hyper-serotonergic[56] states may be involved in depression and sickness behavior.
Microanatomy[edit]
Serotonin is released into the synapse, or space between neurons, and diffuses over a relatively wide gap (>20 nm) to activate 5-HT receptors located on the dendrites, cell bodies, and presynaptic terminals of adjacent neurons.
When humans smell food, dopamine is released to increase the appetite. But, unlike in worms, serotonin does not increase anticipatory behaviour in humans; instead, the serotonin released while consuming activates 5-HT2C receptors on dopamine-producing cells. This halts their dopamine release, and thereby serotonin decreases appetite. Drugs that block 5-HT2C receptors make the body unable to recognize when it is no longer hungry or otherwise in need of nutrients, and are associated with weight gain,[57] especially in people with a low number of receptors.[58] The expression of 5-HT2C receptors in the hippocampus follows a diurnal rhythm,[59] just as the serotonin release in the ventromedial nucleus, which is characterised by a peak at morning when the motivation to eat is strongest.[60]
In macaques, alpha males have twice the level of serotonin in the brain as subordinate males and females (measured by the concentration of 5-HIAA in the cerebrospinal fluid (CSF)). Dominance status and CSF serotonin levels appear to be positively correlated. When dominant males were removed from such groups, subordinate males begin competing for dominance. Once new dominance hierarchies were established, serotonin levels of the new dominant individuals also increased to double those in subordinate males and females. The reason why serotonin levels are only high in dominant males, but not dominant females has not yet been established.[61]
In humans, levels of 5-HT1A receptor inhibition in the brain show negative correlation with aggression,[62] and a mutation in the gene that codes for the 5-HT2A receptor may double the risk of suicide for those with that genotype.[63] Serotonin in the brain is not usually degraded after use, but is collected by serotonergic neurons by serotonin transporters on their cell surfaces. Studies have revealed nearly 10% of total variance in anxiety-related personality depends on variations in the description of where, when and how many serotonin transporters the neurons should deploy.[64]
Outside the nervous system[edit]
Digestive tract (emetic)[edit]
Serotonin regulates gastrointestinal (GI) function. The gut is surrounded by enterochromaffin cells, which release serotonin in response to food in the lumen. This makes the gut contract around the food. Platelets in the veins draining the gut collect excess serotonin. There are often serotonin abnormalities in gastrointestinal disorders such as constipation and irritable bowel syndrome.[65]
If irritants are present in the food, the enterochromaffin cells release more serotonin to make the gut move faster, i.e., to cause diarrhea, so the gut is emptied of the noxious substance. If serotonin is released in the blood faster than the platelets can absorb it, the level of free serotonin in the blood is increased. This activates 5-HT3 receptors in the chemoreceptor trigger zone that stimulate vomiting.[66] Thus, drugs and toxins stimulate serotonin release from enterochromaffin cells in the gut wall. The enterochromaffin cells not only react to bad food but are also very sensitive to irradiation and cancer chemotherapy. Drugs that block 5HT3 are very effective in controlling the nausea and vomiting produced by cancer treatment, and are considered the gold standard for this purpose.[67]
Lungs[edit]
The lung,[68] including that of reptiles,[69] contains specialized epithelial cells that occur as solitary cells or as clusters called neuroepithelial bodies or bronchial Kulchitsky cells or alternatively K cells.[70] These are enterochromaffin cells that like those in the gut release serotonin.[70] Their function is probably vasoconstriction during hypoxia.[68]
Skin[edit]
Serotonin is also produced by Merkel cells which are part of the somatosensory system.[71]
Bone metabolism[edit]
In mice and humans, alterations in serotonin levels and signalling have been shown to regulate bone mass.[72][73][74][75] Mice that lack brain serotonin have osteopenia, while mice that lack gut serotonin have high bone density. In humans, increased blood serotonin levels have been shown to be a significant negative predictor of low bone density. Serotonin can also be synthesized, albeit at very low levels, in the bone cells. It mediates its actions on bone cells using three different receptors. Through 5-HT1B receptors, it negatively regulates bone mass, while it does so positively through 5-HT2B receptors and 5-HT2C receptors. There is very delicate balance between physiological role of gut serotonin and its pathology. Increase in the extracellular content of serotonin results in a complex relay of signals in the osteoblasts culminating in FoxO1/ Creb and ATF4 dependent transcriptional events.[76] Following the 2008 findings that gut serotonin regulates bone mass, the mechanistic investigations into what regulates serotonin synthesis from the gut in the regulation of bone mass have started. Piezo1 has been shown to sense RNA in the gut and relay this information through serotonin synthesis to the bone by acting as a sensor of single-stranded RNA (ssRNA) governing 5-HT production. Intestinal epithelium-specific deletion of mouse Piezo1 profoundly disturbed gut peristalsis, impeded experimental colitis, and suppressed serum 5-HT levels. Because of systemic 5-HT deficiency, conditional knockout of Piezo1 increased bone formation. Notably, fecal ssRNA was identified as a natural Piezo1 ligand, and ssRNA-stimulated 5-HT synthesis from the gut was evoked in a MyD88/TRIF-independent manner. Colonic infusion of RNase A suppressed gut motility and increased bone mass. These findings suggest gut ssRNA as a master determinant of systemic 5-HT levels, indicating the ssRNA-Piezo1 axis as a potential prophylactic target for treatment of bone and gut disorders. Studies in 2008, 2010 and 2019 have opened the potential for serotonin research to treat bone mass disorders.[77][78]
Organ development[edit]
Since serotonin signals resource availability it is not surprising that it affects organ development. Many human and animal studies have shown that nutrition in early life can influence, in adulthood, such things as body fatness, blood lipids, blood pressure, atherosclerosis, behavior, learning, and longevity.[79][80][81] Rodent experiment shows that neonatal exposure to SSRIs makes persistent changes in the serotonergic transmission of the brain resulting in behavioral changes,[82][83] which are reversed by treatment with antidepressants.[84] By treating normal and knockout mice lacking the serotonin transporter with fluoxetine scientists showed that normal emotional reactions in adulthood, like a short latency to escape foot shocks and inclination to explore new environments were dependent on active serotonin transporters during the neonatal period.[85][86]
Human serotonin can also act as a growth factor directly. Liver damage increases cellular expression of 5-HT2A and 5-HT2B receptors, mediating liver compensatory regrowth (see Liver § Regeneration and transplantation)[87] Serotonin present in the blood then stimulates cellular growth to repair liver damage.[88]5HT2B receptors also activate osteocytes, which build up bone[89] However, serotonin also inhibits osteoblasts, through 5-HT1B receptors.[90]
Cardiovascular growth factor[edit]
Serotonin, in addition, evokes endothelial nitric oxide synthase activation and stimulates, through a 5-HT1B receptor-mediated mechanism, the phosphorylation of p44/p42 mitogen-activated protein kinase activation in bovine aortic endothelial cell cultures.[clarification needed][91] In blood, serotonin is collected from plasma by platelets, which store it. It is thus active wherever platelets bind in damaged tissue, as a vasoconstrictor to stop bleeding, and also as a fibrocyte mitotic (growth factor), to aid healing.[92]
Pharmacology[edit]
Several classes of drugs target the 5-HT system, including some antidepressants, antipsychotics, anxiolytics, antiemetics, and antimigraine drugs, as well as, the psychedelic drugs and empathogens.
Mechanism of action[edit]
At rest, serotonin is stored within the vesicles of presynaptic neurons. When stimulated by nerve impulses, serotonin is released as a neurotransmitter into the synapse, reversibly binding to the postsynaptic receptor to induce a nerve impulse on the postsynaptic neuron. Serotonin can also bind to auto-receptors on the presynaptic neuron to regulate the synthesis and release of serotonin. Normally serotonin is taken back into the presynaptic neuron to stop its action, then reused or broken down by monoamine oxidase.[93]
Psychedelic drugs[edit]
The serotonergic psychedelic drugs psilocin/psilocybin, DMT, mescaline, psychedelic mushroom and LSD are agonists, primarily at 5HT2A/2C receptors.[94][95][96] The empathogen-entactogen MDMA releases serotonin from synaptic vesicles of neurons.[97]
Antidepressants[edit]
Drugs that alter serotonin levels are used in treating depression, generalized anxiety disorder, and social phobia. Monoamine oxidase inhibitors (MAOIs) prevent the breakdown of monoamine neurotransmitters (including serotonin), and therefore increase concentrations of the neurotransmitter in the brain. MAOI therapy is associated with many adverse drug reactions, and patients are at risk of hypertensive emergency triggered by foods with high tyramine content, and certain drugs. Some drugs inhibit the re-uptake of serotonin, making it stay in the synaptic cleft longer. The tricyclic antidepressants (TCAs) inhibit the reuptake of both serotonin and norepinephrine. The newer selective serotonin reuptake inhibitors (SSRIs) have fewer side-effects and fewer interactions with other drugs.[98]
Certain SSRI medications have been shown to lower serotonin levels below the baseline after chronic use, despite initial increases.[99] The 5-HTTLPR gene codes for the number of serotonin transporters in the brain, with more serotonin transporters causing decreased duration and magnitude of serotonergic signaling.[100] The 5-HTTLPR polymorphism (l/l) causing more serotonin transporters to be formed is also found to be more resilient against depression and anxiety.[101][102]
Serotonin syndrome[edit]
Extremely high levels of serotonin can cause a condition known as serotonin syndrome, with toxic and potentially fatal effects. In practice, such toxic levels are essentially impossible to reach through an overdose of a single antidepressant drug, but require a combination of serotonergic agents, such as an SSRI with a MAOI, which may occur in therapeutic doses.[103][104] The intensity of the symptoms of serotonin syndrome vary over a wide spectrum, and the milder forms are seen even at nontoxic levels.[105] It is estimated that 14% of patients experiencing serotonin syndrome overdose on SSRIs; meanwhile the fatality rate is between 2% and 12%.[103][106][107]
Antiemetics[edit]
Some 5-HT3 antagonists, such as ondansetron, granisetron, and tropisetron, are important antiemetic agents. They are particularly important in treating the nausea and vomiting that occur during anticancer chemotherapy using cytotoxic drugs. Another application is in the treatment of postoperative nausea and vomiting.
Other[edit]
Some serotonergic agonist drugs cause fibrosis anywhere in the body, particularly the syndrome of retroperitoneal fibrosis, as well as cardiac valve fibrosis.[108]In the past, three groups of serotonergic drugs have been epidemiologically linked with these syndromes. These are the serotonergic vasoconstrictive antimigraine drugs (ergotamine and methysergide),[108] the serotonergic appetite suppressant drugs (fenfluramine, chlorphentermine, and aminorex), and certain anti-Parkinsonian dopaminergic agonists, which also stimulate serotonergic 5-HT2B receptors. These include pergolide and cabergoline, but not the more dopamine-specific lisuride.[109]
As with fenfluramine, some of these drugs have been withdrawn from the market after groups taking them showed a statistical increase of one or more of the side effects described. An example is pergolide. The drug was declining in use since it was reported in 2003 to be associated with cardiac fibrosis.[110]
Two independent studies published in The New England Journal of Medicine in January 2007 implicated pergolide, along with cabergoline, in causing valvular heart disease.[111][112] As a result of this, the FDA removed pergolide from the United States market in March 2007.[113] (Since cabergoline is not approved in the United States for Parkinson's Disease, but for hyperprolactinemia, the drug remains on the market. Treatment for hyperprolactinemia requires lower doses than that for Parkinson's Disease, diminishing the risk of valvular heart disease).[114]
Methyl-tryptamines and hallucinogens[edit]
Several plants contain serotonin together with a family of related tryptamines that are methylated at the amino (NH2) and (OH) groups, are N-oxides, or miss the OH group. These compounds do reach the brain, although some portion of them are metabolized by monoamine oxidase enzymes (mainly MAO-A) in the liver. Examples are plants from the genus Anadenanthera that are used in the hallucinogenic yopo snuff. These compounds are widely present in the leaves of many plants, and may serve as deterrents for animal ingestion. Serotonin occurs in several mushrooms of the genus Panaeolus.[115]
Comparative biology and evolution[edit]
Unicellular organisms[edit]
Serotonin is used by a variety of single-cell organisms for various purposes. SSRIs have been found to be toxic to algae.[116] The gastrointestinal parasite Entamoeba histolytica secretes serotonin, causing a sustained secretory diarrhea in some people.[21][117] Patients infected with E. histolytica have been found to have highly elevated serum serotonin levels, which returned to normal following resolution of the infection.[118] E. histolytica also responds to the presence of serotonin by becoming more virulent.[119] This means serotonin secretion not only serves to increase the spread of enteamoebas by giving the host diarrhea but also serves to coordinate their behaviour according to their population density, a phenomenon known as quorum sensing. Outside the gut of a host, there is nothing that the entoamoebas provoke to release serotonin, hence the serotonin concentration is very low. Low serotonin signals to the entoamoebas they are outside a host and they become less virulent to conserve energy. When they enter a new host, they multiply in the gut, and become more virulent as the enterochromaffine cells get provoked by them and the serotonin concentration increases.
Edible plants and mushrooms[edit]
In drying seeds, serotonin production is a way to get rid of the buildup of poisonous ammonia. The ammonia is collected and placed in the indole part of L-tryptophan, which is then decarboxylated by tryptophan decarboxylase to give tryptamine, which is then hydroxylated by a cytochrome P450 monooxygenase, yielding serotonin.[120]
However, since serotonin is a major gastrointestinal tract modulator, it may be produced in the fruits of plants as a way of speeding the passage of seeds through the digestive tract, in the same way as many well-known seed and fruit associated laxatives. Serotonin is found in mushrooms, fruits, and vegetables. The highest values of 25–400 mg/kg have been found in nuts of the walnut (Juglans) and hickory (Carya) genera. Serotonin concentrations of 3–30 mg/kg have been found in plantains, pineapples, banana, kiwifruit, plums, and tomatoes. Moderate levels from 0.1–3 mg/kg have been found in a wide range of tested vegetables.[22][18]
Serotonin is one compound of the poison contained in stinging nettles (Urtica dioica), where it causes pain on injection in the same manner as its presence in insect venoms.[20] It is also naturally found in Paramuricea clavata, or the Red Sea Fan.[121]
Serotonin and tryptophan have been found in chocolate with varying cocoa contents. The highest serotonin content (2.93 μg/g) was found in chocolate with 85% cocoa, and the highest tryptophan content (13.27–13.34 μg/g) was found in 70–85% cocoa. The intermediate in the synthesis from tryptophan to serotonin, 5-hydroxytryptophan, was not found.[122]
Root development in Arabidopsis thaliana is stimulated and modulated by serotonin – in various ways at various concentrations.[123]
Serotonin serves as a plant defense chemical against fungi. When infected with Fusarium crown rot (Fusarium pseudograminearum), wheat (Triticum aestivum) greatly increases its production of tryptophan to synthesize new serotonin.[124] The function of this is poorly understood[124] but wheat also produces serotonin when infected by Stagonospora nodorum – in that case to retard spore production.[125] The model cereal Brachypodium distachyon – used as a research substitute for wheat and other production cereals – also produces serotonin, coumaroyl-serotonin, and feruloyl-serotonin in response to F. graminearum. This produces a slight antimicrobial effect. B. distachyon produces more serotonin (and conjugates) in response to deoxynivalenol (DON)-producing F. graminearum than non-DON-producing.[126] Solanum lycopersicum produces many AA conjugates – including several of serotonin – in its leaves, stems, and roots in response to Ralstonia solanacearum infection.[127]
Invertebrates[edit]
Serotonin functions as a neurotransmitter in the nervous systems of most animals.
Nematodes[edit]
For example, in the roundworm Caenorhabditis elegans, which feeds on bacteria, serotonin is released as a signal in response to positive events, such as finding a new source of food or in male animals finding a female with which to mate.[128] When a well-fed worm feels bacteria on its cuticle, dopamine is released, which slows it down; if it is starved, serotonin also is released, which slows the animal down further. This mechanism increases the amount of time animals spend in the presence of food.[129] The released serotonin activates the muscles used for feeding, while octopamine suppresses them.[130][131] Serotonin diffuses to serotonin-sensitive neurons, which control the animal's perception of nutrient availability.
Decapods[edit]
If lobsters are injected with serotonin, they behave like dominant individuals whereas octopamine causes subordinate behavior.[28] A crayfish that is frightened may flip its tail to flee, and the effect of serotonin on this behavior depends largely on the animal's social status. Serotonin inhibits the fleeing reaction in subordinates, but enhances it in socially dominant or isolated individuals. The reason for this is social experience alters the proportion between serotonin receptors (5-HT receptors) that have opposing effects on the fight-or-flight response.[clarification needed] The effect of 5-HT1 receptors predominates in subordinate animals, while 5-HT2 receptors predominates in dominants.[132]
In venoms[edit]
Serotonin is a common component of invertebrate venoms, salivary glands, nervous tissues, and various other tissues, across molluscs, insects, crustaceans, scorpions, various kinds of worms, and jellyfish.[20] Adult Rhodnius prolixus – hematophagous on vertebrates – secrete lipocalins into the wound during feeding. In 2003 these lipocalins were demonstrated to sequester serotonin to prevent vasoconstriction (and possibly coagulation) in the host.[133]
Insects[edit]
Serotonin is evolutionarily conserved and appears across the animal kingdom. It is seen in insect processes in roles similar to in the human central nervous system, such as memory, appetite, sleep, and behavior.[134][17] Some circuits in mushroom bodies are serotonergic.[135] (See specific Drosophila example below, §Dipterans.)
Acrididae[edit]
Locust swarming is initiated but not maintained by serotonin,[136] with release being triggered by tactile contact between individuals.[137] This transforms social preference from aversion to a gregarious state that enables coherent groups.[138][137][136] Learning in flies and honeybees is affected by the presence of serotonin.[139][140]
Role in insecticides[edit]
Insect 5-HT receptors have similar sequences to the vertebrate versions, but pharmacological differences have been seen. Invertebrate drug response has been far less characterized than mammalian pharmacology and the potential for species selective insecticides has been discussed.[141]
Hymenopterans[edit]
Wasps and hornets have serotonin in their venom,[142] which causes pain and inflammation[19][20] as do scorpions.[143][20] Pheidole dentata takes on more and more tasks in the colony as it gets older, which requires it to respond to more and more olfactory cues in the course of performing them. This olfactory response broadening was demonstrated to go along with increased serotonin and dopamine, but not octopamine in 2006.[144]
Dipterans[edit]
If flies are fed serotonin, they are more aggressive; flies depleted of serotonin still exhibit aggression, but they do so much less frequently.[145] In their crops it plays a vital role in digestive motility produced by contraction. Serotonin that acts on the crop is exogenous to the crop itself and 2012 research suggested that it probably originated in the serotonin neural plexus in the thoracic-abdominal synganglion.[146] In 2011 a Drosophila serotonergic mushroom body was found to work in concert with Amnesiac to form memories.[135] In 2007 serotonin was found to promote aggression in Diptera, which was counteracted by neuropeptide F – a surprising find given that they both promote courtship, which is usually similar to aggression in most respects.[135]
Vertebrates[edit]
Serotonin, also referred to as 5-hydroxytryptamine (5-HT), is a neurotransmitter most known for its involvement in mood disorders in humans. It is also a widely present neuromodulator among vertebrates and invertebrates.[147] Serotonin has been found having associations with many physiological systems such as cardiovascular, thermoregulation, and behavioral functions, including: circadian rhythm, appetite, aggressive and sexual behavior, sensorimotor reactivity and learning, and pain sensitivity.[148] Serotonin's function in neurological systems along with specific behaviors among vertebrates found to be strongly associated with serotonin will be further discussed. Two relevant case studies are also mentioned regarding serotonin development involving teleost fish and mice.
In mammals, 5-HT is highly concentrated in the substantia nigra, ventral tegmental area and raphe nuclei. Lesser concentrated areas include other brain regions and the spinal cord.[147] 5-HT neurons are also shown to be highly branched, indicating that they are structurally prominent for influencing multiple areas of the CNS at the same time, although this trend is exclusive solely to mammals.[148]
5-HT system in vertebrates[edit]
Vertebrates are multicellular organisms in the phylum Chordata that possess a backbone and a nervous system. This includes mammals, fish, reptiles, birds, etc. In humans, the nervous system is composed of the central and peripheral nervous system, with little known about the specific mechanisms of neurotransmitters in most other vertebrates. However, it is known that while serotonin is involved in stress and behavioral responses, it is also important in cognitive functions.[147] Brain organization in most vertebrates includes 5-HT cells in the hindbrain.[147] In addition to this, 5-HT is often found in other sections of the brain in non-placental vertebrates, including the basal forebrain and pretectum.[149] Since location of serotonin receptors contribute to behavioral responses, this suggests serotonin is part of specific pathways in non-placental vertebrates that are not present in amniotic organisms.[150] Teleost fish and mice are organisms most often used to study the connection between serotonin and vertebrate behavior. Both organisms show similarities in the effect of serotonin on behavior, but differ in the mechanism in which the responses occur.
Dogs / canine species[edit]
There are few studies of serotonin in dogs. One study reported serotonin values were higher at dawn than at dusk.[151] In another study, serum 5-HT levels did not seem to be associated with dogs' behavioural response to a stressful situation.[152] Urinary serotonin/creatinine ratio in bitches tended to be higher 4 weeks after surgery. In addition, serotonin was positively correlated with both cortisol and progesterone but not with testosterone after ovariohysterectomy.[153]
Teleost fish[edit]
Like non-placental vertebrates, teleost fish also possess 5-HT cells in other sections of the brain, including the basal forebrain.[149] Danio rerio (zebra fish) are a species of teleost fish often used for studying serotonin within the brain. Despite much being unknown about serotonergic systems in vertebrates, the importance in moderating stress and social interaction is known.[154] It is hypothesized that AVT and CRF cooperate with serotonin in the hypothalamic-pituitary-interrenal axis.[149] These neuropeptides influence the plasticity of the teleost, affecting its ability to change and respond to its environment. Subordinate fish in social settings show a drastic increase in 5-HT concentrations.[154] High levels of 5-HT long term influence the inhibition of aggression in subordinate fish.[154]
Mice[edit]
Researchers at the Department of Pharmacology and Medical Chemistry used serotonergic drugs on male mice to study the effects of selected drugs on their behavior.[155] Mice in isolation exhibit increased levels of agonistic behavior towards one another. Results found that serotonergic drugs reduce aggression in isolated mice while simultaneously increasing social interaction.[155] Each of the treatments use a different mechanism for targeting aggression, but ultimately all have the same outcome. While the study shows that serotonergic drugs successfully target serotonin receptors, it does not show specifics of the mechanisms that affect behavior, as all types of drugs tended to reduce aggression in isolated male mice.[155] Aggressive mice kept out of isolation may respond differently to changes in serotonin reuptake.
Behavior[edit]
Как и у людей, серотонин активно участвует в регуляции поведения большинства других позвоночных. Это включает в себя не только реакцию и социальное поведение, но и влияние на настроение. Дефекты в путях серотонина могут привести к резким изменениям настроения, а также к симптомам расстройств настроения, которые могут присутствовать не только у людей.
Социальное взаимодействие [ править ]
Одним из наиболее изученных аспектов социального взаимодействия, в котором участвует серотонин, является агрессия. Агрессия регулируется системой 5-HT, поскольку уровни серотонина могут как вызывать, так и подавлять агрессивное поведение, как это наблюдается у мышей (см. раздел «Мыши») и крабов. [155] Хотя это широко признано, неизвестно, взаимодействует ли серотонин прямо или косвенно с частями мозга, влияющими на агрессию и другое поведение. [147] Исследования уровней серотонина показывают, что они резко повышаются и снижаются во время социальных взаимодействий и обычно коррелируют с подавлением или возбуждением агрессивного поведения. [156] Точный механизм влияния серотонина на социальное поведение неизвестен, поскольку пути в системе 5-НТ у разных позвоночных могут сильно различаться. [147]
Реакция на раздражители [ править ]
Серотонин играет важную роль в путях реакции окружающей среды наряду с другими нейротрансмиттерами . [157] В частности, было обнаружено, что он участвует в слуховой обработке в социальных условиях, поскольку первичные сенсорные системы связаны с социальными взаимодействиями. [158] Серотонин обнаружен в структуре IC среднего мозга, которая обрабатывает видоспецифичные и неспецифические социальные взаимодействия и вокализации. [158] Он также получает акустические проекции, которые передают сигналы в области слуховой обработки. [158] Исследования показали, что серотонин формирует слуховую информацию, получаемую ИЦ, и, следовательно, влияет на реакцию на слуховые стимулы. [158] Это может влиять на то, как организм реагирует на звуки хищных или других агрессивных видов в окружающей среде, поскольку поглощение серотонина может влиять на агрессию и/или социальное взаимодействие.
Настроение [ править ]
Мы можем описывать настроение не как специфическое для эмоционального статуса, а как связанное с относительно длительным эмоциональным состоянием. Связь серотонина с настроением наиболее известна при различных формах депрессии и биполярных расстройствах у людей. [148] Расстройства, вызванные серотонинергической активностью, потенциально способствуют появлению многих симптомов большой депрессии, таких как общее настроение, активность, суицидальные мысли, а также сексуальная и когнитивная дисфункция . Селективные ингибиторы обратного захвата серотонина (СИОЗС) представляют собой класс препаратов, продемонстрировавших свою эффективность при лечении большого депрессивного расстройства, и являются наиболее назначаемым классом антидепрессантов. Функция СИОЗС заключается в блокировании обратного захвата серотонина, делая возможным поглощение большего количества серотонина принимающим нейроном. Животные изучались на протяжении десятилетий, чтобы понять депрессивное поведение среди видов. Одно из самых известных исследований, тест принудительного плавания (FST), было проведено для измерения потенциальной антидепрессивной активности. [148] Крыс помещали в неизбежный контейнер с водой, после чего сравнивали время, проведенное в неподвижном состоянии, и количество активных действий (таких как плескание или лазание) до и после введения группы антидепрессантов. Было показано, что антидепрессанты, которые избирательно ингибируют обратный захват NE, уменьшают неподвижность и избирательно увеличивают способность лазать, не влияя на плавание. Однако результаты применения СИОЗС также показывают снижение обездвиженности, но увеличение плавания без ущерба для лазания. Это исследование продемонстрировало важность поведенческих тестов на антидепрессанты, поскольку они могут обнаруживать лекарства, влияющие на основное поведение наряду с поведенческими компонентами вида. [148]
Рост и размножение [ править ]
У нематоды C. elegans искусственное истощение серотонина или повышение уровня октопамина сигнализирует о поведении, типичном для среды с низким содержанием пищи: C. elegans становится более активной, а спаривание и яйцекладка подавляются, тогда как при повышении серотонина происходит обратное. или октопамина у этого животного снижено. [31] Серотонин необходим для нормального спаривания самцов нематод. [159] и склонность оставлять еду в поисках партнера. [160] Серотонинергическая передача сигналов, используемая для адаптации поведения червя к быстрым изменениям окружающей среды, влияет на инсулиноподобную передачу сигналов и путь передачи сигналов TGF-бета . [161] которые контролируют долгосрочную адаптацию.
У плодовых мух инсулин регулирует уровень сахара в крови , а также действует как фактор роста . Таким образом, у плодовой мухи серотонинергические нейроны регулируют размер тела взрослой особи, влияя на секрецию инсулина. [162] [163] Серотонин также был идентифицирован как триггер стайного поведения саранчи. [138] У людей, хотя инсулин регулирует уровень сахара в крови, а ИФР регулирует рост, серотонин контролирует высвобождение обоих гормонов, модулируя высвобождение инсулина из бета-клеток поджелудочной железы посредством серотонилирования сигнальных белков ГТФазы. [39] Воздействие СИОЗС во время беременности замедляет рост плода. [164]
Генетически измененные черви C. elegans , у которых отсутствует серотонин, имеют увеличенную продолжительность репродуктивной жизни, могут страдать ожирением и иногда имеют задержку развития в состоянии покоя личинки . [165] [166]
фенотипы Старение и
Известно, что серотонин регулирует старение, обучение и память. Первые доказательства получены в результате изучения долголетия C. elegans . [161] На ранней стадии старения [ нечеткий ] , повышается уровень серотонина, что изменяет локомоторное поведение и ассоциативную память. [167] Эффект восстанавливается мутациями и препаратами (в том числе миансерином и метиотепином ), ингибирующими серотониновые рецепторы . Это наблюдение не противоречит представлению о снижении уровня серотонина у млекопитающих и человека, что обычно наблюдается в поздние, а не ранние сроки. [ нечеткий ] фаза старения.
Биохимические механизмы [ править ]
Биосинтез [ править ]

У животных и человека серотонин синтезируется из аминокислоты L — триптофана по короткому метаболическому пути, состоящему из двух ферментов : гидроксилазы триптофана (TPH) и декарбоксилазы ароматических аминокислот (DDC), а также кофермента пиридоксальфосфата . Реакция, опосредованная TPH, является лимитирующей стадией пути.Было показано, что TPH существует в двух формах: TPH1 , обнаруженный в нескольких тканях , и TPH2 , который является нейрон-специфической изоформой . [168]
Серотонин можно синтезировать из триптофана в лаборатории, используя Aspergillus niger и Psilocybe coprophila в качестве катализаторов. На первом этапе получения 5-гидрокситриптофана потребуется оставить триптофан в этаноле и воде в течение 7 дней, затем добавить достаточное количество HCl (или другой кислоты), чтобы довести pH до 3, а затем добавить NaOH, чтобы довести pH до 13, в течение 1 часа. . Aspergillus niger станет катализатором этой первой фазы. Второй этап синтеза самого триптофана из промежуточного продукта 5-гидрокситриптофана потребует добавления этанола и воды и на этот раз оставления на 30 дней. Следующие два шага будут такими же, как и первый этап: добавление HCl для достижения pH = 3, а затем добавление NaOH для достижения очень основного pH на уровне 13 в течение 1 часа. используется Psilocybe coprophila . На этой фазе в качестве катализатора реакции [169]

Серотонин, принимаемый перорально, не попадает в серотонинергические пути центральной нервной системы, поскольку не проникает через гематоэнцефалический барьер . [9] Однако триптофан и его метаболит 5-гидрокситриптофан (5-HTP), из которого синтезируется серотонин, проникают через гематоэнцефалический барьер. Эти агенты доступны в виде пищевых добавок и в различных продуктах питания и могут быть эффективными серотонинергическими агентами.Одним из продуктов распада серотонина является 5-гидроксииндолуксусная кислота (5-HIAA), которая выводится с мочой . Серотонин и 5-HIAA иногда производятся в избыточных количествах некоторыми опухолями или раковыми заболеваниями , и уровни этих веществ можно измерить в моче для проверки на наличие этих опухолей.
Аналитическая химия [ править ]
Оксид индия и олова рекомендуется использовать в качестве материала электрода при электрохимическом исследовании концентраций, производимых, обнаруживаемых или потребляемых микробами . [170] В 1994 году был разработан метод масс-спектрометрии для измерения молекулярной массы как природных, так и синтетических серотонинов. [171]
История и этимология [ править ]
Физиологам уже более столетия было известно, что сосудосуживающие вещества появляются в сыворотке крови при свертывании крови. [172] В 1935 году итальянец Витторио Эрспамер продемонстрировал, что экстракт энтерохромаффинных клеток заставляет кишечник сокращаться. Некоторые считали, что он содержит адреналин , но два года спустя Эрспамер смог доказать, что это был ранее неизвестный амин , который он назвал «энтерамином». [173] [174] В 1948 году Морис М. Раппорт , Арда Грин и Ирвин Пейдж из Кливлендской клиники сосудосуживающее вещество обнаружили в сыворотке крови , и, поскольку это был сывороточный агент, влияющий на тонус сосудов, они назвали его серотонином. [175]
В 1952 году было показано, что энтерамин является тем же веществом, что и серотонин, и по мере того, как был выяснен широкий спектр физиологических ролей, аббревиатура 5-HT от собственного химического названия 5-гидрокситриптамин стала предпочтительным названием в области фармакологии. [176] Синонимы серотонина включают: 5-гидрокситриптамин, тромботин, энтерамин, вещество DS и 3-(β-аминоэтил)-5-гидроксииндол. [177] В 1953 году Бетти Тварог и Пейдж обнаружили серотонин в центральной нервной системе. [178] Пейдж считал работу Эрспамера над Octopus vulgaris , Discoglossus pictus , Hexaplex trunculus , Bolinus Brandaris , Sepia , Mytilus и Ostrea как достоверную и фундаментальную для понимания этого недавно идентифицированного вещества, но считал свои более ранние результаты в различных моделях, особенно на основе крысиной крови, быть слишком сбитым с толку присутствием других биологически активных химических веществ, включая некоторые другие вазоактивные вещества. [179]
См. также [ править ]
Примечания [ править ]
- ^ Ссылки на функции этих рецепторов доступны на страницах Википедии для конкретного рассматриваемого рецептора.
Ссылки [ править ]
- ^ Рассчитано с использованием программного обеспечения Advanced Chemistry Development (ACD/Labs) V11.02 (© 1994–2011 ACD/Labs).
- ^ Мазак К., Дочи В., Кёкёши Дж., Носал Б. (апрель 2009 г.). «Протонное видообразование и микровидообразование серотонина и 5-гидрокситриптофана». Химия и биоразнообразие . 6 (4): 578–590. дои : 10.1002/cbdv.200800087 . ПМИД 19353542 . S2CID 20543931 .
- ^ Стоун С (1958). «[Индольные производные. II. Новый способ синтеза серотонина]». Медицина; Научное издание (на итальянском языке). 13 (1): 75–79. ПМИД 13524273 .
- ^ Эрспамер V (1952). «Предварительные исследования индолалкиламинов и фенилалкиламинов из экстрактов кожи амфибий». Научные исследования . 22 :694–702.
- ^ Таммисто Т. (1967). «Повышение токсичности 5-гидрокситриптамина этанолом у крыс и мышей». Annales Medicinae Experimentalis et Biologiae Fenniae . 46 (3): 382–384. ПМИД 5734241 .
- ^ Джонс Д. (2003) [1917]. Роуч П., Хартманн Дж., Сеттер Дж. (ред.). Словарь английского произношения . Кембридж: Издательство Кембриджского университета. ISBN 978-3-12-539683-8 .
- ^ «Серотонин» . Dictionary.com Полный (онлайн). nd
- ^ «Серотонин» . Словарь Merriam-Webster.com .
- ^ Перейти обратно: а б Молодой СН (ноябрь 2007 г.). «Как повысить серотонин в мозгу человека без лекарств» . Журнал психиатрии и неврологии . 32 (6): 394–399. ПМК 2077351 . ПМИД 18043762 .
- ^ «Микробы помогают производить серотонин в кишечнике» . Калифорнийский технологический институт . 9 апреля 2015 года . Проверено 3 июня 2022 г.
- ^ Кинг МВ. «Серотонин» . Страница медицинской биохимии . Медицинский факультет Университета Индианы . Проверено 1 декабря 2009 г.
- ^ Бергер М., Грей Дж.А., Рот Б.Л. (2009). «Расширенная биология серотонина» . Ежегодный обзор медицины . 60 : 355–366. дои : 10.1146/annurev.med.60.042307.110802 . ПМЦ 5864293 . ПМИД 19630576 .
- ^ Шлингер Р.Г., Мейер Ч.Р. (2003). «Влияние селективных ингибиторов обратного захвата серотонина на активацию тромбоцитов: могут ли они предотвратить острый инфаркт миокарда?». Американский журнал сердечно-сосудистых препаратов . 3 (3): 149–162. дои : 10.2165/00129784-200303030-00001 . ПМИД 14727927 . S2CID 23986530 .
- ^ Клинг А (2013). 5-HT2A: рецептор серотонина, который может играть роль в заболеваниях суставов (PDF) (Диссертация). Университет Умео. ISBN 978-91-7459-549-9 .
- ^ Яно Дж.М., Ю.К., Дональдсон Г.П., Шастри Г.Г., Энн П., Ма Л. и др. (апрель 2015 г.). «Аборигенные бактерии кишечной микробиоты регулируют биосинтез серотонина хозяина» . Клетка . 161 (2): 264–276. дои : 10.1016/j.cell.2015.02.047 . ПМЦ 4393509 . ПМИД 25860609 .
- ^ Ванхутт П.М. (февраль 1987 г.). «Серотонин и сосудистая стенка». Международный журнал кардиологии . 14 (2): 189–203. дои : 10.1016/0167-5273(87)90008-8 . ПМИД 3818135 .
- ^ Перейти обратно: а б Хузер А., Роведдер А., Апостолопулу А.А., Видманн А., Пфиценмайер Дж.Е., Майоло Э.М. и др. (2012). Зарс Т. (ред.). «Серотонинергическая центральная нервная система личинки дрозофилы: анатомия и поведенческая функция» . ПЛОС ОДИН . 7 (10): е47518. Бибкод : 2012PLoSO...747518H . дои : 10.1371/journal.pone.0047518 . ПМЦ 3474743 . ПМИД 23082175 .
- ^ Перейти обратно: а б с Рамакришна А., Гиридхар П., Равишанкар Г.А. (июнь 2011 г.). «Фитосеротонин: обзор» . Сигнализация и поведение растений . 6 (6): 800–809. Бибкод : 2011PlSiB...6..800A . дои : 10.4161/psb.6.6.15242 . ПМК 3218476 . ПМИД 21617371 .
- ^ Перейти обратно: а б Чен Дж., Ларивьер В.Р. (октябрь 2010 г.). «Ноцицептивные и антиноцицептивные эффекты инъекции и терапии пчелиным ядом: палка о двух концах» . Прогресс нейробиологии . 92 (2): 151–183. doi : 10.1016/j.pneurobio.2010.06.006 . ПМЦ 2946189 . ПМИД 20558236 .
- ^ Перейти обратно: а б с д и Эрспамер V (1966). «Присутствие индолалкиламинов в природе». 5-Гидрокситриптамин и родственные индолалкиламины . Берлин , Гейдельберг : Springer Berlin Heidelberg . стр. 132–181. дои : 10.1007/978-3-642-85467-5_4 . ISBN 978-3-642-85469-9 .
- ^ Перейти обратно: а б Макгоуэн К., Кейн А., Асаркоф Н., Уикс Дж., Герина В., Келлум Дж. и др. (август 1983 г.). «Entamoeba histolytica вызывает кишечную секрецию: роль серотонина». Наука . 221 (4612): 762–764. Бибкод : 1983Sci...221..762M . дои : 10.1126/science.6308760 . ПМИД 6308760 .
- ^ Перейти обратно: а б Фельдман Дж. М., Ли Э. М. (октябрь 1985 г.). «Содержание серотонина в пищевых продуктах: влияние на выведение 5-гидроксииндолуксусной кислоты с мочой» . Американский журнал клинического питания . 42 (4): 639–643. дои : 10.1093/ajcn/42.4.639 . ПМИД 2413754 .
- ^ Гонсалес-Флорес Д., Велардо Б., Гарридо М. и др. (2011). «Употребление в пищу сливы японской (Prunus salicina Lindl. сорт Crimson Globe) увеличивает уровень 6-сульфатоксимелатонина в моче и общий уровень антиоксидантной способности у молодых, средних и пожилых людей: пищевая и функциональная характеристика их содержания» . Журнал исследований продуктов питания и питания . 50 (4): 229–236.
- ^ Рычков Д.А., Хантер С., Ковальский В.Я., Ломзов А.А., Пулхэм Ч.Р., Болдырева Е.В. (июль 2016 г.). «К пониманию кристаллизации из раствора. ТФФ-исследования многокомпонентных кристаллов серотонина». Вычислительная и теоретическая химия . 1088 : 52–61. дои : 10.1016/j.comptc.2016.04.027 .
- ^ Наим М., Чедейн А.Р., Голен Дж.А., Манке Д.Р. (апрель 2022 г.). «Кристаллическая структура серотонина» . Acta Crystallographica Раздел E. 78 (Часть 4): 365–368. Бибкод : 2022AcCrE..78..365N . дои : 10.1107/S2056989022002559 . ПМЦ 8983975 . ПМИД 35492269 .
- ^ Рычков Д., Болдырева Е.В., Туманов Н.А. (сентябрь 2013 г.). «Новая структура соли серотонина: сравнение и конформационный анализ всех известных серотониновых комплексов». Acta Crystallographica Раздел C. 69 (Часть 9): 1055–1061. Бибкод : 2013AcCrC..69.1055R . дои : 10.1107/S0108270113019823 . ПМИД 24005521 .
- ^ Мохаммад-Заде Л.Ф., Мозес Л., Гуолтни-Брант С.М. (июнь 2008 г.). «Серотонин: обзор» . Журнал ветеринарной фармакологии и терапии . 31 (3): 187–199. дои : 10.1111/j.1365-2885.2008.00944.x . ПМИД 18471139 .
- ^ Перейти обратно: а б Кравиц Э.А. (сентябрь 1988 г.). «Гормональный контроль поведения: амины и смещение поведенческих реакций у омаров». Наука . 241 (4874): 1775–1781. Бибкод : 1988Sci...241.1775K . дои : 10.1126/science.2902685 . ПМИД 2902685 .
- ^ Синклер-Уилсон А., Лоуренс А., Ферезу И., Картоннет Х., Майлес С., Гарел С. и др. (август 2023 г.). «Пластичность таламокортикальных аксонов регулируется уровнем серотонина, модулируемым преждевременными родами» . Труды Национальной академии наук Соединенных Штатов Америки . 120 (33): e2301644120. Бибкод : 2023PNAS..12001644S . дои : 10.1073/pnas.2301644120 . ПМЦ 10438379 . ПМИД 37549297 .
- ^ Перейти обратно: а б Зифа Э., Филлион Дж. (сентябрь 1992 г.). «5-гидрокситриптаминовые рецепторы» . Фармакологические обзоры . 44 (3): 401–458. ПМИД 1359584 .
- ^ Перейти обратно: а б Шринивасан С., Садег Л., Элль И.С., Кристенсен А.Г., Фагерман Н.Дж., Ашрафи К. (июнь 2008 г.). «Серотонин регулирует жир и питание C. elegans посредством независимых молекулярных механизмов» . Клеточный метаболизм . 7 (6): 533–544. дои : 10.1016/j.cmet.2008.04.012 . ПМК 2495008 . ПМИД 18522834 .
- ^ Рамакришна А., Равишанкар Г.А. (ноябрь 2011 г.). «Влияние сигналов абиотического стресса на вторичные метаболиты растений» . Сигнализация и поведение растений . 6 (11). Информация : 1720–1731. Бибкод : 2011PlSiB...6.1720A . дои : 10.4161/psb.6.11.17613 . ПМЦ 3329344 . ПМИД 22041989 .
- ^ Уитакер Р., Косгроув Л. (2015). Психиатрия под влиянием: институциональная коррупция, социальный ущерб и рецепты реформ . Спрингер. стр. 55–56. ISBN 978-1-137-51602-2 .
- ^ Монкрифф Дж., Купер Р.Э., Стокманн Т., Амендола С., Хенгартнер М.П., Горовиц М.А. (август 2023 г.). «Серотониновая теория депрессии: систематический обзор доказательств» . Молекулярная психиатрия . 28 (8). Издательская группа «Природа»: 3243–3256. дои : 10.1038/s41380-022-01661-0 . ПМЦ 10618090 . ПМИД 35854107 . S2CID 250646781 .
- ^ Гаеми Н. (2022), Была ли опровергнута гипотеза серотонина? , получено 2 мая 2023 г.
- ^ Прах А., Пург М., Старе Дж., Вианелло Р., Маври Дж. (сентябрь 2020 г.). «Как моноаминоксидаза А разлагает серотонин: эмпирическое моделирование валентной связи на реакционной стадии» . Журнал физической химии. Б. 124 (38): 8259–8265. doi : 10.1021/acs.jpcb.0c06502 . ПМЦ 7520887 . ПМИД 32845149 .
- ^ Хэннон Дж., Хойер Д. (декабрь 2008 г.). «Молекулярная биология 5-НТ-рецепторов». Поведенческие исследования мозга . 195 (1): 198–213. дои : 10.1016/j.bbr.2008.03.020 . ПМИД 18571247 . S2CID 46043982 .
- ^ Перейти обратно: а б Чжоу М., Энгель К., Ван Дж. (январь 2007 г.). «Доказательства значительного вклада недавно идентифицированного переносчика моноаминов (PMAT) в поглощение серотонина в мозге человека» . Биохимическая фармакология . 73 (1): 147–154. дои : 10.1016/j.bcp.2006.09.008 . ПМК 1828907 . ПМИД 17046718 .
- ^ Перейти обратно: а б с Паульманн Н., Громанн М., Фойгт Дж.П., Берт Б., Вовинкель Дж., Бадер М. и др. (октябрь 2009 г.). О'Рахили С. (ред.). «Внутриклеточный серотонин модулирует секрецию инсулина бета-клетками поджелудочной железы путем серотонилирования белка» . ПЛОС Биология . 7 (10): e1000229. дои : 10.1371/journal.pbio.1000229 . ПМК 2760755 . ПМИД 19859528 .
- ^ Walther DJ, Peter JU, Winter S, Höltje M, Paulmann N, Grohmann M и др. (декабрь 2003 г.). «Серотонилирование малых ГТФаз является путем передачи сигнала, который запускает высвобождение альфа-гранул тромбоцитами» . Клетка . 115 (7): 851–862. дои : 10.1016/S0092-8674(03)01014-6 . ПМИД 14697203 . S2CID 16847296 .
- ^ Уоттс С.В., Пристли-младший, Томпсон Дж.М. (май 2009 г.). «Серотонилирование сосудистых белков, важных для сокращения» . ПЛОС ОДИН . 4 (5): е5682. Бибкод : 2009PLoSO...4.5682W . дои : 10.1371/journal.pone.0005682 . ПМЦ 2682564 . ПМИД 19479059 .
- ^ Рот Б.Л., Дрискол Дж. (12 января 2011 г.). «База данных ПДСП К и » . Программа скрининга психоактивных веществ (PDSP) . Университет Северной Каролины в Чапел-Хилл и Национальный институт психического здоровья США. Архивировано из оригинала 8 ноября 2013 года . Проверено 17 декабря 2013 г.
- ^ Бортолоцци А., Диас-Матайкс Л., Скорца М.К., Селада П., Артигас Ф. (декабрь 2005 г.). «Активация 5-НТ-рецепторов в префронтальной коре усиливает дофаминергическую активность». Журнал нейрохимии . 95 (6): 1597–1607. дои : 10.1111/j.1471-4159.2005.03485.x . hdl : 10261/33026 . ПМИД 16277612 . S2CID 18350703 .
- ^ Моро С., Эдвардс Л., Чесс-Уильямс Р. (ноябрь 2016 г.). 5-HT 2A « Усиление сократительной активности уротелия и собственной пластинки свиной рецептора » . Международный журнал урологии . 23 (11): 946–951. дои : 10.1111/iju.13172 . ПМИД 27531585 .
- ^ «Нейрон Фон Экономо – НейронБанк» . Neuronbank.org . [ ненадежный медицинский источник? ]
- ^ Миллан М.Дж., Гобер А., Лежен Ф., Декейн А., Ньюман-Танкреди А., Пасто В. и др. (сентябрь 2003 г.). «Новый агонист мелатонина агомелатин (S20098) является антагонистом рецепторов 5-гидрокситриптамина2C, блокада которых усиливает активность лобнокортикальных дофаминергических и адренергических путей». Журнал фармакологии и экспериментальной терапии . 306 (3): 954–964. дои : 10.1124/jpet.103.051797 . ПМИД 12750432 . S2CID 18753440 .
- ^ Гонсалес Р., Чавес-Паскасио К., Менесес А. (сентябрь 2013 г.). «Роль рецепторов 5-HT5A в консолидации памяти». Поведенческие исследования мозга . 252 : 246–251. дои : 10.1016/j.bbr.2013.05.051 . ПМИД 23735322 . S2CID 140204585 .
- ^ Наутиял К.М., Хен Р. (2017). «Серотониновые рецепторы при депрессии: от А к Б» . F1000Исследования . 6 : 123. дои : 10.12688/f1000research.9736.1 . ПМК 5302148 . ПМИД 28232871 .
- ^ Фрейзер А., Хенслер Дж.Г. (1999). «Понимание нейроанатомической организации серотонинергических клеток головного мозга дает представление о функциях этого нейромедиатора» . В: Сигел Г.Дж. , Агранов, Бернард В., Фишер С.К., Альберс Р.В., Улер, доктор медицинских наук (ред.). Основная нейрохимия (Шестое изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0-397-51820-3 .
В 1964 году Дальстрем и Фукс (обсуждается в [2]), используя метод гистофлуоресценции Фалька-Хилларпа , заметили, что большая часть серотонинергической сомы обнаруживается в группах клеточных тел, которые ранее обозначались как ядра шва.
- ^ Биндер, доктор медицинских наук, Хирокава Н. (2009). энциклопедия нейробиологии . Берлин: Шпрингер. п. 705. ИСБН 978-3-540-23735-8 .
- ^ Группа нейронов ядер шва расположена вдоль ствола мозга от меток « Средний мозг » до « Продолговатый мозг», с центром на мосту . ( См. соответствующее изображение .)
- ^ Мюллер К.П., Джейкобс Б.Л., ред. (2009). Справочник по поведенческой нейробиологии серотонина (1-е изд.). Лондон: Академик. стр. 51–59. ISBN 978-0-12-374634-4 .
- ^ Пассос А.Д., Эркулано А.М., Оливейра К.Р., де Лима С.М., Роча Ф.А., Фрейтас Х.Р. и др. (октябрь 2019 г.). «Регуляция серотонинергической системы каинатом в сетчатке птиц». Клеточная и молекулярная нейробиология . 39 (7): 1039–1049. дои : 10.1007/s10571-019-00701-8 . ПМИД 31197744 . S2CID 189763144 .
- ^ Соммер С (2009). «Серотонин в боли и контроле боли». В Мюллер КП, Джейкобс Б.Л. (ред.). Справочник по поведенческой нейробиологии серотонина (1-е изд.). Лондон: Академик. стр. 457–460. ISBN 978-0-12-374634-4 .
- ^ Хенслер Дж.Г. (2009). «Серотонин в режиме и эмоциях». В Мюллер КП, Джейкобс Б.Л. (ред.). Справочник по поведенческой нейробиологии серотонина (1-е изд.). Лондон: Академик. стр. 367–399. ISBN 978-0-12-374634-4 .
- ^ Эндрюс П.В., Бхарвани А., Ли КР, Фокс М., Томсон Дж.А. (апрель 2015 г.). «Является ли серотонин верхним или нижним? Эволюция серотонинергической системы и ее роль в депрессии и реакции на антидепрессанты». Неврологические и биоповеденческие обзоры . 51 : 164–188. doi : 10.1016/j.neubiorev.2015.01.018 . ПМИД 25625874 . S2CID 23980182 .
- ^ Шталь С.М., Миньон Л., Мейер Дж.М. (март 2009 г.). «Что на первом месте: атипичная антипсихотическая терапия или кардиометаболический риск?» . Acta Psychiatrica Scandinavica . 119 (3): 171–179. дои : 10.1111/j.1600-0447.2008.01334.x . ПМИД 19178394 . S2CID 24035040 .
- ^ Бакленд П.Р., Хугендорн Б., Гай К.А., Смит С.К., Коулман С.Л., О'Донован MC (март 2005 г.). «Низкая экспрессия генов, обусловленная ассоциацией аллели гена рецептора 5-HT2C с увеличением веса, вызванным антипсихотиками». Американский журнал психиатрии . 162 (3): 613–615. дои : 10.1176/appi.ajp.162.3.613 . ПМИД 15741483 .
- ^ Холмс MC, Френч К.Л., Секль-младший (июнь 1997 г.). «Нарушение регуляции суточных ритмов серотонина 5-HT2C и экспрессии генов рецепторов кортикостероидов в гиппокампе при ограничении пищи и глюкокортикоидах» . Журнал неврологии . 17 (11): 4056–4065. doi : 10.1523/JNEUROSCI.17-11-04056.1997 . ПМК 6573558 . ПМИД 9151722 .
- ^ Лейбовиц С.Ф. (1990). «Роль серотонина в расстройствах пищевого поведения». Наркотики . 39 (Приложение 3): 33–48. дои : 10.2165/00003495-199000393-00005 . ПМИД 2197074 . S2CID 8612545 .
- ^ Макгуайр, Майкл (2013) «Вера, нейробиология фантазий, страхов и противоречий» (Prometius Books)
- ^ Каспи Н., Модаи И., Барак П., Вайсборд А., Збарски Х., Хиршманн С. и др. (март 2001 г.). «Аугментация пиндолола у агрессивных пациентов с шизофренией: двойное слепое перекрестное рандомизированное исследование». Международная клиническая психофармакология . 16 (2): 111–115. дои : 10.1097/00004850-200103000-00006 . ПМИД 11236069 . S2CID 24822810 .
- ^ Ито З., Айзава И., Такеучи М., Табе М., Накамура Т. (декабрь 1975 г.). «[Труды: Исследование моторики желудочно-кишечного тракта с использованием датчика внепросветной силы. 6. Наблюдение моторики желудка и двенадцатиперстной кишки с использованием синтетического мотилина]». Нихон Хэйкацукин Гаккай Засши . 11 (4): 244–246. ПМИД 1232434 .
- ^ Леш К.П., Бенгель Д., Хейлс А., Сабол С.З., Гринберг Б.Д., Петри С. и др. (ноябрь 1996 г.). «Связь черт, связанных с тревогой, с полиморфизмом в регуляторной области гена переносчика серотонина». Наука . 274 (5292): 1527–1531. Бибкод : 1996Sci...274.1527L . дои : 10.1126/science.274.5292.1527 . ПМИД 8929413 . S2CID 35503987 .
- ^ Битти Д.Т., Смит Дж.А. (май 2008 г.). «Фармакология серотонина в желудочно-кишечном тракте: обзор». Архив фармакологии Наунина-Шмидеберга . 377 (3): 181–203. дои : 10.1007/s00210-008-0276-9 . ПМИД 18398601 . S2CID 32820765 .
- ^ Позвонил HP (2003). Фармакология . Эдинбург: Черчилль Ливингстон. п. 187. ИСБН 978-0-443-07145-4 .
- ^ де Вит Р., Аапро М., Блоуэр П.Р. (сентябрь 2005 г.). «Есть ли фармакологическая основа для различий в эффективности антагонистов 5-HT3-рецепторов у рефрактерных пациентов?». Химиотерапия и фармакология рака . 56 (3): 231–238. дои : 10.1007/s00280-005-1033-0 . ПМИД 15838653 . S2CID 27576150 .
- ^ Перейти обратно: а б Лауверинс Дж. М., Кокеларе Дж., Теунинк П. (апрель 1973 г.). «Нейроэпителиальные тельца, продуцирующие серотонин, в слизистой оболочке дыхательных путей кролика». Наука . 180 (4084): 410–413. Бибкод : 1973Sci...180..410L . дои : 10.1126/science.180.4084.410 . ПМИД 4121716 . S2CID 2809307 .
- ^ Пастор Л.М., Бальеста Х., Перес-Томас Р., Марин Х.А., Эрнандес Ф., Мадрид Х.Ф. (июнь 1987 г.). «Иммуноцитохимическая локализация серотонина в легких рептилий». Исследования клеток и тканей . 248 (3). ООО «Спрингер Сайенс энд Бизнес Медиа»: 713–715. дои : 10.1007/bf00216504 . ПМИД 3301000 . S2CID 9871728 .
- ^ Перейти обратно: а б Сонстегард К.С., Мейлман Р.Б., Чик Дж.М., Томлин Т.Е., ДиОгустин Р.П. (ноябрь 1982 г.). «Морфо-цитохимическая характеристика нейроэпителиальных тел легких плода кролика. I. Исследование изолированных нейроэпителиальных тел». Экспериментальное исследование легких . 3 (3–4): 349–377. дои : 10.3109/01902148209069663 . ПМИД 6132813 .
- ^ Чанг В., Канда Х., Икеда Р., Линг Дж., ДеБерри Дж.Дж., Гу Дж.Г. (сентябрь 2016 г.). «Диск Меркеля представляет собой серотонинергический синапс в эпидермисе для передачи тактильных сигналов у млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (37): E5491–E5500. Бибкод : 2016PNAS..113E5491C . дои : 10.1073/pnas.1610176113 . ПМК 5027443 . ПМИД 27573850 .
- ^ Фрост М., Андерсен Т.Е., Ядав В., Бриксен К., Карсенти Г., Кассем М. (март 2010 г.). «Пациенты с фенотипом высокой костной массы из-за мутации Lrp5-T253I имеют низкие уровни серотонина в плазме» . Журнал исследований костей и минералов . 25 (3): 673–675. дои : 10.1002/jbmr.44 . ПМИД 20200960 . S2CID 24280062 .
- ^ Розен CJ (февраль 2009 г.). «Прорыв в биологию костей: секреты серотонина». Природная медицина . 15 (2): 145–146. дои : 10.1038/nm0209-145 . ПМИД 19197289 . S2CID 5489589 .
- ^ Мёддер У.И., Ахенбах С.Дж., Амин С., Риггс Б.Л., Мелтон Л.Дж., Хосла С. (февраль 2010 г.). «Связь уровня серотонина в сыворотке крови с плотностью костей и структурными параметрами у женщин» . Журнал исследований костей и минералов . 25 (2): 415–422. дои : 10.1359/jbmr.090721 . ПМК 3153390 . ПМИД 19594297 .
- ^ Фрост М., Андерсен Т., Госсиел Ф., Хансен С., Боллерслев Дж., ван Хул В. и др. (август 2011 г.). «Уровни серотонина, склеростина, маркеров костного обмена, а также плотность и микроархитектура костной ткани у пациентов с фенотипом высокой костной массы из-за мутации Lrp5» . Журнал исследований костей и минералов . 26 (8): 1721–1728. дои : 10.1002/jbmr.376 . ПМИД 21351148 .
- ^ Коде А., Мосиалоу И., Силва Б.С., Рачед М.Т., Чжоу Б., Ван Дж. и др. (октябрь 2012 г.). «FOXO1 управляет функцией серотонина, полученного из кишечника, по подавлению костей» . Журнал клинических исследований . 122 (10): 3490–3503. дои : 10.1172/JCI64906 . ПМК 3461930 . ПМИД 22945629 .
- ^ Ядав В.К., Баладжи С., Суреш П.С., Лю XS, Лу X, Ли З и др. (март 2010 г.). «Фармакологическое ингибирование синтеза серотонина в кишечнике является потенциальным костным анаболическим средством лечения остеопороза» . Природная медицина . 16 (3): 308–312. дои : 10.1038/нм.2098 . ПМЦ 2836724 . ПМИД 20139991 .
- ^ Сугисава Э., Такаяма Ю., Такемура Н., Кондо Т., Хатакеяма С., Кумагай Ю. и др. (август 2020 г.). «Ощущение РНК с помощью Gut Piezo1 необходимо для системного синтеза серотонина» . Клетка . 182 (3): 609–624.e21. дои : 10.1016/j.cell.2020.06.022 . ПМИД 32640190 .
- ^ Озанн SE, Hales CN (январь 2004 г.). «Продолжительность жизни: догоняющий рост и ожирение у мышей-самцов». Природа . 427 (6973): 411–412. Бибкод : 2004Natur.427..411O . дои : 10.1038/427411b . ПМИД 14749819 . S2CID 40256021 .
- ^ Льюис Д.С., Бертран Х.А., МакМахан К.А., Макгилл Х.К., Кэри К.Д., Масоро Э.Дж. (октябрь 1986 г.). «Прием пищи перед отъемом влияет на ожирение молодых взрослых бабуинов» . Журнал клинических исследований . 78 (4): 899–905. дои : 10.1172/JCI112678 . ПМК 423712 . ПМИД 3760191 .
- ^ Хан П. (июль 1984 г.). «Влияние размера помета на уровень холестерина и инсулина в плазме, а также некоторых ферментов печени и жировой ткани у взрослых грызунов». Журнал питания . 114 (7): 1231–1234. дои : 10.1093/jn/114.7.1231 . ПМИД 6376732 .
- ^ Попа Д., Лена С., Александр С., Адриен Дж. (апрель 2008 г.). «Длительный синдром депрессии, вызванный снижением поглощения серотонина во время постнатального развития: данные сна, стресса и поведения» . Журнал неврологии . 28 (14): 3546–3554. doi : 10.1523/JNEUROSCI.4006-07.2008 . ПМК 6671102 . ПМИД 18385313 .
- ^ Макиаг Д., Симпсон К.Л., Коппингер Д., Лу Ю., Ван Ю., Лин Р.К. и др. (январь 2006 г.). «Воздействие неонатальных антидепрессантов оказывает долгосрочное воздействие на поведение и серотониновую систему» . Нейропсихофармакология . 31 (1): 47–57. дои : 10.1038/sj.npp.1300823 . ПМК 3118509 . ПМИД 16012532 .
- ^ Макиаг Д., Уильямс Л., Коппингер Д., Пол И.А. (февраль 2006 г.). «Воздействие циталопрама у новорожденных вызывает долговременные изменения в поведении, которые обращаются вспять при лечении имипрамином у взрослых» . Европейский журнал фармакологии . 532 (3): 265–269. дои : 10.1016/j.ejphar.2005.12.081 . ПМЦ 2921633 . ПМИД 16483567 .
- ^ Холден С. (октябрь 2004 г.). «Нейронаука. Лечение новорожденных мышей прозаком вызывает беспокойство» . Наука . 306 (5697): 792. doi : 10.1126/science.306.5697.792 . ПМИД 15514122 .
- ^ Ансорж М.С., Чжоу М., Лира А., Хен Р., Гингрич Дж.А. (октябрь 2004 г.). «Блокада транспортера 5-HT в раннем возрасте меняет эмоциональное поведение взрослых мышей» . Наука . 306 (5697): 879–881. Бибкод : 2004Sci...306..879A . дои : 10.1126/science.1101678 . ПМИД 15514160 .
- ^ Лесуртель М., Граф Р., Алейл Б., Вальтер Дж., Тиан Ю., Йохум В. и др. (апрель 2006 г.). «Продуцируемый тромбоцитами серотонин опосредует регенерацию печени». Наука . 312 (5770): 104–107. Бибкод : 2006Sci...312..104L . дои : 10.1126/science.1123842 . ПМИД 16601191 . S2CID 43189753 .
- ^ Матондо Р.Б., Пунт С., Хомберг Дж., Туссен М.Дж., Кисес Р., Корпораал С.Дж. и др. (апрель 2009 г.). «Удаление переносчика серотонина у крыс нарушает гомеостаз серотонина без нарушения регенерации печени» . Американский журнал физиологии. Физиология желудочно-кишечного тракта и печени . 296 (4): G963–G968. дои : 10.1152/ajpgi.90709.2008 . ПМИД 19246633 . Архивировано из оригинала 28 декабря 2019 года . Проверено 5 декабря 2019 г.
- ^ Колле С., Шильц С., Жоффруа В., Марото Л., Лоне Ж.М., де Вернежуль М.К. (февраль 2008 г.). «Рецептор серотонина 5-HT2B контролирует костную массу посредством рекрутирования и пролиферации остеобластов» . Журнал ФАСЭБ . 22 (2): 418–427. дои : 10.1096/fj.07-9209com . ПМК 5409955 . ПМИД 17846081 .
- ^ Ядав В.К., Рю Дж.Х., Суда Н., Танака К.Ф., Гингрич Дж.А., Шютц Г. и др. (ноябрь 2008 г.). «Lrp5 контролирует образование костной ткани, ингибируя синтез серотонина в двенадцатиперстной кишке» . Клетка . 135 (5): 825–837. дои : 10.1016/j.cell.2008.09.059 . ПМЦ 2614332 . ПМИД 19041748 .
- «Чтобы построить кости, нужна смелость, выяснили ученые» . ScienceDaily (пресс-релиз). 1 декабря 2008 г.
- ^ Макдаффи Дж. Э., Мотли Э.Д., Лимберд Л.Е., Малек М.А. (март 2000 г.). «5-гидрокситриптамин стимулирует фосфорилирование митоген-активируемой протеинкиназы p44/p42 в культурах эндотелиальных клеток аорты крупного рогатого скота» . Журнал сердечно-сосудистой фармакологии . 35 (3): 398–402. дои : 10.1097/00005344-200003000-00008 . ПМИД 10710124 .
- ^ Ногучи М., Фурукава К.Т., Моримото М. (декабрь 2020 г.). «Легочные нейроэндокринные клетки: физиология, тканевый гомеостаз и заболевания» . Модели и механизмы заболеваний . 13 (12): dmm046920. дои : 10.1242/dmm.046920 . ПМЦ 7774893 . ПМИД 33355253 .
- ^ Фуллер Р.В. (1980). «Фармакология центральных серотониновых нейронов». Ежегодный обзор фармакологии и токсикологии . 20 : 111–127. дои : 10.1146/annurev.pa.20.040180.000551 . ПМИД 6992697 .
- ^ Название М., Лион Р.А., Гленнон Р.А. (1988). «Доказательства связывания радиолигандов указывают на то, что рецептор 5-HT2 мозга является местом действия галлюциногенов ЛСД и фенилизопропиламина». Психофармакология . 94 (2): 213–216. дои : 10.1007/BF00176847 . ПМИД 3127847 . S2CID 24179554 .
- ^ Николс Д.Е. (2000). «Роль серотонинергических нейронов и 5-НТ-рецепторов в действии галлюциногенов». В Баумгартене Х.Г., Готерте М. (ред.). Серотонинергические нейроны и 5-НТ-рецепторы в ЦНС . Санта-Клара, Калифорния: Springer-Verlag TELOS. ISBN 978-3-540-66715-5 .
- ^ Капур С., Симан П. (2002). «Антагонисты рецепторов NMDA кетамин и PCP оказывают прямое воздействие на рецепторы дофамина D (2) и серотонина 5-HT (2) - последствия для моделей шизофрении» . Молекулярная психиатрия . 7 (8): 837–844. дои : 10.1038/sj.mp.4001093 . ПМИД 12232776 .
- ^ Член парламента Джонсона, Хоффман А.Дж., Николс Д.Е. (декабрь 1986 г.). «Влияние энантиомеров МДА, МДМА и родственных аналогов на высвобождение [3H] серотонина и [3H] дофамина из перелитых срезов мозга крыс». Европейский журнал фармакологии . 132 (2–3): 269–276. дои : 10.1016/0014-2999(86)90615-1 . ПМИД 2880735 .
- ^ Гудман Л.С., Брантон Л.Л., Чабнер Б., Ноллманн Б.К. (2001). Фармакологические основы терапии Гудмана и Гилмана . Нью-Йорк: МакГроу-Хилл. стр. 459–461. ISBN 978-0-07-162442-8 .
- ^ Бенмансур С., Чекки М., Морилак Д.А., Герхардт Г.А., Яворс М.А., Гулд Г.Г. и др. (декабрь 1999 г.). «Влияние хронического лечения антидепрессантами на функцию переносчика серотонина, плотность и уровень мРНК» . Журнал неврологии . 19 (23): 10494–10501. doi : 10.1523/JNEUROSCI.19-23-10494.1999 . ПМК 6782424 . ПМИД 10575045 .
- ^ Бейчман Дж. Х., Бальдассарра Л., Мик Х., Де Лука В., Кинг Н., Бендер Д. и др. (июнь 2006 г.). «Полиморфизмы транспортера серотонина и стойкая, всеобъемлющая детская агрессия». Американский журнал психиатрии . 163 (6): 1103–1105. дои : 10.1176/appi.ajp.163.6.1103 . ПМИД 16741214 .
- ^ Пезавас Л., Мейер-Линденберг А., Драбант Э.М., Верчински Б.А., Муньос К.Е., Колачана Б.С. и др. (июнь 2005 г.). «Полиморфизм 5-HTTLPR влияет на взаимодействие поясной извилины и миндалевидного тела человека: генетический механизм предрасположенности к депрессии». Природная неврология . 8 (6): 828–834. дои : 10.1038/nn1463 . ПМИД 15880108 . S2CID 1864631 .
- ^ Шинка Дж.А., Буш Р.М., Робишо-Кин Н. (февраль 2004 г.). «Метаанализ связи между полиморфизмом гена переносчика серотонина (5-HTTLPR) и личностной тревожностью» . Молекулярная психиатрия . 9 (2): 197–202. дои : 10.1038/sj.mp.4001405 . ПМИД 14966478 .
- ^ Перейти обратно: а б Нью А.М., Нельсон С., Люнг Дж.Г. (1 октября 2015 г.). Александр Э., Сусла GM (ред.). «Неотложная психиатрическая помощь в отделении интенсивной терапии». Расширенная интенсивная терапия AACN . 26 (4): 285–293, викторина 294–295. doi : 10.4037/NCI.0000000000000104 . ПМИД 26484986 .
- ^ Исбистер Г.К., Боу С.Дж., Доусон А., Уайт И.М. (2004). «Относительная токсичность селективных ингибиторов обратного захвата серотонина (СИОЗС) при передозировке». Журнал токсикологии. Клиническая токсикология . 42 (3): 277–285. дои : 10.1081/CLT-120037428 . ПМИД 15362595 . S2CID 43121327 .
- ^ Данкли Э.Дж., Исбистер Г.К., Сиббритт Д., Доусон А.Х., Уайт И.М. (сентябрь 2003 г.). «Критерии токсичности серотонина Хантера: простые и точные правила диагностического принятия решения о токсичности серотонина» . КДЖМ . 96 (9): 635–642. дои : 10.1093/qjmed/hcg109 . ПМИД 12925718 .
- ^ Фрэнк С. (июль 2008 г.). «Распознавание и лечение серотонинового синдрома» . Канадский семейный врач . 54 (7): 988–992. ПМК 2464814 . ПМИД 18625822 .
- ^ Бойер Э.В., Шеннон М. (март 2005 г.). «Серотониновый синдром». Медицинский журнал Новой Англии . 352 (11): 1112–1120. дои : 10.1056/NEJMra041867 . ПМИД 15784664 .
- ^ Перейти обратно: а б Баскин С.И. (1991). Основы кардиотоксикологии . Бока-Ратон: CRC Press. ISBN 978-0-8493-8809-5 . Проверено 3 февраля 2010 г.
- ^ Йахнихен С., Хоровски Р., Перц Х. «Перголид и каберголин, но не лисурид, проявляют эффективность агонистов в отношении рецепторов серотонина 5-HT 2B » (PDF) . Проверено 3 февраля 2010 г.
- ^ Консультативный комитет по нежелательным реакциям на лекарства, Австралия (2004 г.). «Сердечная вальвулопатия с перголидом» . Aust Adv Drug React Bull . 23 (4). Архивировано из оригинала 27 июня 2012 года.
- ^ Шаде Р., Андерсон Ф., Суисса С., Хаверкамп В., Гарбе Э. (январь 2007 г.). «Агонисты дофамина и риск регургитации сердечного клапана» . Медицинский журнал Новой Англии . 356 (1): 29–38. doi : 10.1056/NEJMoa062222 . ПМИД 17202453 .
- ^ Занеттини Р., Антонини А., Гатто Дж., Джентиле Р., Тесей С., Пеццоли Г. (январь 2007 г.). «Клапанные пороки сердца и применение агонистов дофамина при болезни Паркинсона» . Медицинский журнал Новой Англии . 356 (1): 39–46. doi : 10.1056/NEJMoa054830 . ПМИД 17202454 .
- ^ «Консультации по общественному здравоохранению Управления по санитарному надзору за качеством пищевых продуктов и медикаментов» . Управление по контролю за продуктами и лекарствами . 29 марта 2007 года . Проверено 7 февраля 2010 г.
- ^ «MedWatch – Информационные предупреждения по безопасности 2007 г. Пермакс (перголид) и дженерики» . США Управление по санитарному надзору за качеством пищевых продуктов и медикаментов . 29 марта 2007 года . Проверено 30 марта 2007 г.
- ^ Тайлер В.Е. (сентябрь 1958 г.). «Появление серотонина в галлюциногенном грибе». Наука . 128 (3326): 718. Бибкод : 1958Sci...128..718T . дои : 10.1126/science.128.3326.718 . ПМИД 13580242 .
- ^ Джонсон DJ, Сандерсон Х., Брейн Р.А., Уилсон С.Дж., Соломон К.Р. (май 2007 г.). «Токсичность и опасность антидепрессантов, селективных ингибиторов обратного захвата серотонина, флуоксетина, флувоксамина и сертралина для водорослей». Экотоксикология и экологическая безопасность . 67 (1): 128–139. Бибкод : 2007ЭкоЭС..67..128J . дои : 10.1016/j.ecoenv.2006.03.016 . ПМИД 16753215 .
- ^ Макгоуэн К., Герина В., Уикс Дж., Доновиц М. (1985). «Секреторные гормоны Entamoeba histolytica ». Симпозиум Фонда Ciba . Симпозиумы Фонда Новартис. Том. 112. С. 139–154. дои : 10.1002/9780470720936.ch8 . ISBN 978-0-470-72093-6 . ПМИД 2861068 .
- ^ Бану Н., Заиди К.Р., Мехди Г., Мансур Т. (июль 2005 г.). «Нейрогуморальные изменения и их роль при амебиазе» . Индийский журнал клинической биохимии . 20 (2): 142–145. дои : 10.1007/BF02867414 . ПМЦ 3453840 . ПМИД 23105547 .
- ^ Ачарья Д.П., Сен М.Р., Сен ПК (август 1989 г.). «Влияние экзогенного 5-гидрокситриптамина на патогенность Entamoeba histolytica у экспериментальных животных». Индийский журнал экспериментальной биологии . 27 (8): 718–720. ПМИД 2561282 .
- ^ Шредер П., Абеле С., Гор П., Штульфаут-Ройш У., Гросс В. (1999). «Последние энзимологии биосинтеза серотонина в семенах грецкого ореха». Триптофан, Серотонин и Мелатонин . Достижения экспериментальной медицины и биологии. Том. 467. стр. 637–644. дои : 10.1007/978-1-4615-4709-9_81 . ISBN 978-0-306-46204-7 . ПМИД 10721112 .
- ^ Пенез Н., Кулиоли Г., Перес Т., Бриан Ж.Ф., Томас О.П., Блаш И. (октябрь 2011 г.). «Противообрастающие свойства простых индольных и пуриновых алкалоидов средиземноморской горгонии Paramuricea clavata». Журнал натуральных продуктов . 74 (10): 2304–2308. дои : 10.1021/np200537v . ПМИД 21939218 .
- ^ Гильен-Касла В., Росалес-Конрадо Н., Леон-Гонсалес М.Э., Перес-Аррибас Л.В., Поло-Диес Л.М. (апрель 2012 г.). «Определение серотонина и его предшественников в образцах шоколада методом капиллярной жидкостной хроматографии с масс-спектрометрическим детектированием». Журнал хроматографии А. 1232 : 158–165. дои : 10.1016/j.chroma.2011.11.037 . ПМИД 22186492 .
- ^ Пелахио-Флорес Р., Ортис-Кастро Р., Мендес-Браво А., Масиас-Родригес Л., Лопес-Бусио Х. (март 2011 г.). «Серотонин, сигнал триптофана, сохраняющийся у растений и животных, регулирует архитектуру корневой системы, вероятно, действуя как естественный ингибитор ауксина у Arabidopsis thaliana» . Физиология растений и клеток . 52 (3). Издательство Оксфордского университета (OUP): 490–508. дои : 10.1093/pcp/pcr006 . ПМИД 21252298 .
- ^ Перейти обратно: а б Пауэлл Дж. Дж., Карер Дж., Фицджеральд Т.Л., Стиллер Дж., Коварелли Л., Сюй Кью и др. (март 2017 г.). «Возбудитель фузариозной кроновой гнили Fusarium pseudograminearum вызывает ряд транскрипционных и метаболических изменений в мягкой пшенице (Triticum aestivum L.)» . Анналы ботаники . 119 (5). Издательство Оксфордского университета (OUP): 853–867. дои : 10.1093/aob/mcw207 . ПМК 5604588 . ПМИД 27941094 . S2CID 3823345 .
- ^ Дю Фаль Л.А., Соломон П.С. (октябрь 2013 г.). «Некротрофический эффектор SnToxA индуцирует синтез нового фитоалексина в пшенице» . Новый фитолог . 200 (1). Уайли : 185–200. дои : 10.1111/nph.12356 . ПМИД 23782173 .
- ^ Паске Ж.К., Шауш С., Макадре С., Бальзерг С., Юге С., Мартин-Маньетт М.Л. и др. (июль 2014 г.). «Дифференциальная экспрессия генов и метаболомный анализ Brachypodium distachyon, зараженного продуцирующими и непродуцирующими дезоксиниваленол штаммами Fusarium graminearum» . БМК Геномика . 15 (1). BioMed Central : 629. doi : 10.1186/1471-2164-15-629 . ПМК 4124148 . ПМИД 25063396 .
- ^ Цейсс Д.Р., Пиатер Л.А., Дубери И.А. (февраль 2021 г.). «Гидроксикоричные амиды: интересные конъюгаты защитных метаболитов растений». Тенденции в науке о растениях . 26 (2). Cell Press : 184–195. doi : 10.1016/j.tplants.2020.09.011 . ПМИД 33036915 . S2CID 222256660 .
- ^ Йонц М.Г., ЕкатериниМерсье А., ЖоффрПоттер Дж.В. (2001). «Влияние 5-HT (серотонина) на репродуктивное поведение Heterodera Schachtii (Nematoda)». Канадский журнал зоологии . 79 (9): 1727. doi : 10.1139/z01-135 .
- ^ Савин Э.Р., Ранганатан Р., Хорвиц Х.Р. (июнь 2000 г.). «Скорость локомоторики C. elegans модулируется окружающей средой через дофаминергический путь и опытом через серотонинергический путь» . Нейрон . 26 (3): 619–631. doi : 10.1016/S0896-6273(00)81199-X . ПМИД 10896158 . S2CID 9247380 .
- ^ Ниакарис Т., Эйвери Л. (январь 2003 г.). «Серотонин регулирует реполяризацию глоточной мышцы C. elegans» . Журнал экспериментальной биологии . 206 (Часть 2): 223–231. дои : 10.1242/jeb.00101 . ПМЦ 4441752 . ПМИД 12477893 .
- ^ Россо М.Н., Джонс Дж.Т., Абад П. (2009). «РНКи и функциональная геномика растительных паразитических нематод». Ежегодный обзор фитопатологии . 47 (1). Годовые обзоры : 207–232. дои : 10.1146/annurev.phyto.112408.132605 . ПМИД 19400649 . п. 218:
Октопамин и серотонин регулируют активность нейронов М3, которые управляют сокращением глотки во время кормления C. elegans ... Замачивание Meloidogyne J2 в дцРНК в присутствии... резорцина плюс серотонин приводило к поглощению растворов и подавлению генов. экспрессируется в кишечнике и пищеводных железах.
- ^ Йе С.Р., Фрике Р.А., Эдвардс Д.Х. (январь 1996 г.). «Влияние социального опыта на серотонинергическую модуляцию схемы побега раков». Наука . 271 (5247): 366–369. Бибкод : 1996Sci...271..366Y . CiteSeerX 10.1.1.470.6528 . дои : 10.1126/science.271.5247.366 . ПМИД 8553075 . S2CID 1575533 .
- ^ Фрай Б.Г., Рулантс К., Шампань Д.Э., Шейб Х., Тиндалл Дж.Д., Кинг Г.Ф. и др. (2009). «Токсигеномная мультивселенная: конвергентное пополнение белков в яды животных» . Ежегодный обзор геномики и генетики человека . 10 (1). Годовые обзоры : 483–511. дои : 10.1146/annurev.genom.9.081307.164356 . ПМИД 19640225 .
- ^ «Серотонин, серотониновые рецепторы и их действие у насекомых» . Нейромедиатор . 2 : 1–14. 2015. дои : 10.14800/nt.314 .
- ^ Перейти обратно: а б с Шуфс Л., Де Луф А., Ван Хил М.Б. (январь 2017 г.). «Нейропептиды как регуляторы поведения насекомых» . Ежегодный обзор энтомологии . 62 (1). Годовые обзоры : 35–52. doi : 10.1146/annurev-ento-031616-035500 . ПМИД 27813667 .
- ^ Перейти обратно: а б Ван Х, Кан Л (7 января 2014 г.). «Молекулярные механизмы изменения фаз у саранчи» . Ежегодный обзор энтомологии . 59 (1). Годовые обзоры : 225–244. doi : 10.1146/annurev-ento-011613-162019 . ПМИД 24160426 .
п. 231,
Изменение количества нескольких потенциальных нейротрансмиттеров... таких как серотонин... может играть важную роль в ремоделировании ЦНС во время фазовых изменений (26, 56, 80).
п. 233,
У саранчи S. gregaria количество серотонина в грудных ганглиях положительно коррелировало со степенью стадного поведения, вызванного различными периодами скученности. Серия фармакологических и поведенческих экспериментов продемонстрировала, что серотонин играет ключевую роль в индукции начальной поведенческой грегаризации (2, 80). Однако серотонин не отвечает за поддержание стадного поведения, поскольку его количество у долгоживущих стадных саранчовых составляет менее половины от этого количества у долгоживущих одиночных особей (80). У L. migratoria инъекция серотонина также может в некоторой степени инициировать стадное поведение, но серотонин в сочетании с лечением скученности вызывал более одиночное поведение, чем одна только инъекция серотонина (48). Значительных различий в уровнях серотонина в тканях головного мозга между двумя фазами L. migratoria не обнаружено . В недавнем отчете Танаки и Нишиде (97) измерялось поведение влечения/избегания у S. gregaria после однократных и многократных инъекций серотонина в различных концентрациях. Серотонин оказывал лишь кратковременное влияние на уровень некоторых двигательных функций и не участвовал в контроле стадного поведения (97). Кроме того, неясно, как нейромедиатор влияет на это уникальное поведение, поскольку модель бинарной логистической регрессии, используемая в этих исследованиях для поведенческого анализа, фокусировалась в основном только на одном поведенческом параметре, представляющем общее фазовое состояние. Очевидно, что изменение фазы поведения может включать альтернативные механизмы регулирования у разных видов саранчи. Таким образом, эти исследования показывают, что регуляторные механизмы ЦНС, регулирующие инициирование и поддержание фазовых изменений, являются видоспецифичными и включают взаимодействие между этими нейротрансмиттерами.
Учитывая ключевую роль аминергической передачи сигналов, какие последующие пути участвуют в формировании долговременной памяти? Отт и др. (63) исследовали роль [] протеинкиназы[] в фазовом изменении у S. gregaria : ... цАМФ-зависимая протеинкиназа А (PKA). Используя фармакологическое вмешательство и RNAi, эти авторы продемонстрировали, что PKA... играет решающую роль в модуляции склонности саранчи к приобретению и выражению стадного поведения. ... К сожалению, хотя была выдвинута гипотеза о корреляции между серотонином и ПКА, прямых доказательств предоставлено не было. - ^ Перейти обратно: а б Чжан Л., Лекок М., Лачининский А., Хантер Д. (январь 2019 г.). «Борьба с саранчой и кузнечиками» . Ежегодный обзор энтомологии . 64 (1). Годовые обзоры : 15–34. doi : 10.1146/annurev-ento-011118-112500 . ПМИД 30256665 . S2CID 52843907 . п. 20:
...грегаризация вызывается... тактильной стимуляцией... Тактильные раздражители вызывают увеличение биогенных аминов, особенно серотонина, в нервной системе саранчи (1, 116); эти амины играют решающую роль в нейрофизиологии смены фаз поведения саранчи.
- ^ Перейти обратно: а б Ансти М.Л., Роджерс С.М., Отт С.Р., Берроуз М., Симпсон С.Дж. (январь 2009 г.). «Серотонин опосредует поведенческую сплоченность, лежащую в основе формирования стаи пустынной саранчи». Наука . 323 (5914): 627–630. Бибкод : 2009Sci...323..627A . дои : 10.1126/science.1165939 . ПМИД 19179529 . S2CID 5448884 .
- Морган Дж. (29 января 2009 г.). «Саранча полна серотонина» . Новости Би-би-си .
- ^ Ситараман Д., Лаферьер Х., Бирман С., Зарс Т. (июнь 2012 г.). «Серотонин имеет решающее значение для вознаграждения обонятельной кратковременной памяти у дрозофилы». Журнал нейрогенетики . 26 (2): 238–244. дои : 10.3109/01677063.2012.666298 . ПМИД 22436011 . S2CID 23639918 .
- ^ Бикер Дж., Мензель Р. (январь 1989 г.). «Химические коды контроля поведения членистоногих». Природа . 337 (6202): 33–39. Бибкод : 1989Natur.337...33B . дои : 10.1038/337033a0 . ПМИД 2562906 . S2CID 223750 .
- ^ Цай М, Ли З, Фань Ф, Хуан Ц, Шао Икс, Сун Г (март 2010 г.). «Разработка и синтез новых инсектицидов на основе серотонинергического лиганда 1-[(4-аминофенил)этил]-4-[3-(трифторметил)фенил]пиперазина (ПАПП)». Журнал сельскохозяйственной и пищевой химии . 58 (5): 2624–2629. дои : 10.1021/jf902640u . ПМИД 20000410 .
- ^ Манахан С.Э. (2002). Токсикологическая химия и биохимия (3-е изд.). ЦРК Пресс. п. 393. ИСБН 978-1-4200-3212-3 .
- ^ Постма ТЛ (2009). «Нейротоксичные яды и яды животных» . В Доббсе MR (ред.). Клиническая нейротоксикология . УБ Сондерс. стр. 463–489. дои : 10.1016/B978-032305260-3.50049-6 . ISBN 978-0-323-05260-3 .
- ^ Гаденн С., Баррозо Р.Б., Антон С. (11 марта 2016 г.). «Пластичность обоняния насекомых: пахнуть или не пахнуть?». Ежегодный обзор энтомологии . 61 (1). Годовые обзоры : 317–333. doi : 10.1146/annurev-ento-010715-023523 . hdl : 11336/19586 . ПМИД 26982441 . S2CID 207568844 .
- ^ Дирик Х.А., Гринспен Р.Дж. (май 2007 г.). «Серотонин и нейропептид F оказывают противоположное модулирующее воздействие на агрессию мух». Природная генетика . 39 (5): 678–682. дои : 10.1038/ng2029 . ПМИД 17450142 . S2CID 33768246 .
- ^ Стоффолано Дж. Г. , Хаселтон А.Т. (7 января 2013 г.). «Взрослые двукрылые: уникальный и игнорируемый орган». Ежегодный обзор энтомологии . 58 (1). Годовые обзоры : 205–225. doi : 10.1146/annurev-ento-120811-153653 . ПМИД 23317042 .
- ^ Перейти обратно: а б с д и ж Бакке-Казенав Ж., Бхаратия Р., Барьер Г., Дельбек Ж.П., Бугию Н., Ди Джованни Г. и др. (февраль 2020 г.). «Серотонин в познании и поведении животных» . Международный журнал молекулярных наук . 21 (5): 1649. doi : 10.3390/ijms21051649 . ПМК 7084567 . ПМИД 32121267 .
- ^ Перейти обратно: а б с д и Лаки I (август 1998 г.). «Спектр поведения, на который влияет серотонин» . Биологическая психиатрия . 44 (3): 151–162. дои : 10.1016/s0006-3223(98)00139-5 . ПМИД 9693387 . S2CID 3001666 .
- ^ Перейти обратно: а б с Бэкстрем Т., Винберг С. (25 октября 2017 г.). «Серотонин координирует реакцию на социальный стресс: чему мы можем научиться у рыбы» . Границы в неврологии . 11 : 595. дои : 10.3389/fnins.2017.00595 . ПМЦ 5669303 . ПМИД 29163002 .
- ^ Бергер М., Грей Дж.А., Рот Б.Л. (1 февраля 2009 г.). «Расширенная биология серотонина» . Ежегодный обзор медицины . 60 (1): 355–366. дои : 10.1146/annurev.med.60.042307.110802 . ПМЦ 5864293 . ПМИД 19630576 .
- ^ Альбергина Д., Пиччионе Г., Пумилия Г., Джоэ М., Риццо М., Раффо П. и др. (июль 2019 г.). «Суточные колебания серотонина и кортизола в моче у здоровых приютских собак и влияние внутривидового социального воздействия». Физиология и поведение . 206 : 1–6. дои : 10.1016/j.physbeh.2019.03.016 . ПМИД 30898540 . S2CID 81965422 .
- ^ Риджио Дж., Марити С., Серджи В., Диверио С., Гаццано А. (декабрь 2020 г.). «Концентрация серотонина и триптофана в сыворотке у собак из приюта, демонстрирующих разные поведенческие реакции на потенциально стрессовую процедуру» . Ветеринарные науки . 8 (1): 1. doi : 10.3390/vetsci8010001 . ПМЦ 7824451 . ПМИД 33374183 .
- ^ Хидбринг-Сандберг Э., Ларссон Э., Мадей А., Хёглунд О.В. (июль 2021 г.). «Кратковременное влияние овариогистерэктомии на серотонин, кортизол, тестостерон и прогестерон в моче у сук» . Исследовательские заметки BMC . 14 (1): 265. doi : 10.1186/s13104-021-05680-y . ПМЦ 8272283 . ПМИД 34246304 .
- ^ Перейти обратно: а б с Винберг С., Торнквист П.О. (июнь 2016 г.). «Роль серотонина мозга в модуляции поведения рыб» . Современная зоология . 62 (3): 317–323. дои : 10.1093/cz/zow037 . ПМК 5804243 . ПМИД 29491919 .
- ^ Перейти обратно: а б с д Оливье Б., Мос Дж., Ван дер Хейден Дж., Хартог Дж. (1 февраля 1989 г.). «Серотонинергическая модуляция социальных взаимодействий у изолированных мышей-самцов». Психофармакология . 97 (2): 154–156. дои : 10.1007/BF00442239 . ПМИД 2498921 . S2CID 37170174 .
- ^ Хубер Р., Смит К., Делаго А., Исакссон К., Кравиц Э.А. (май 1997 г.). «Серотонин и агрессивная мотивация ракообразных: изменение решения об отступлении» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (11): 5939–5942. Бибкод : 1997PNAS...94.5939H . дои : 10.1073/pnas.94.11.5939 . ПМК 20885 . ПМИД 9159179 .
- ^ Санчес КЛ, Бискап К.С., Герперц С., Габер Т.Дж., Кун СМ, Худ С.Х. и др. (май 2015 г.). «Роль серотонина (5-НТ) в поведенческом контроле: результаты исследований на животных и клинические последствия» . Международный журнал нейропсихофармакологии . 18 (10): pyv050. дои : 10.1093/ijnp/pyv050 . ПМЦ 4648158 . ПМИД 25991656 .
- ^ Перейти обратно: а б с д Петерсен CL, Херли LM (октябрь 2017 г.). «В контексте: связь слуховой обработки с цепями социального поведения в мозге позвоночных» . Интегративная и сравнительная биология . 57 (4): 865–877. дои : 10.1093/icb/icx055 . ПМК 6251620 . ПМИД 28985384 .
- ^ Лоер CM, Кеньон CJ (декабрь 1993 г.). «Мутанты с дефицитом серотонина и брачное поведение самцов нематоды Caenorhabditis elegans» . Журнал неврологии . 13 (12): 5407–5417. doi : 10.1523/JNEUROSCI.13-12-05407.1993 . ПМК 6576401 . ПМИД 8254383 .
- ^ Липтон Дж., Климанн Г., Гош Р., Линтс Р., Эммонс С.В. (август 2004 г.). «Поиск партнера у Caenorhabditis elegans: генетическая модель полового влечения у простого беспозвоночного» . Журнал неврологии . 24 (34): 7427–7434. doi : 10.1523/JNEUROSCI.1746-04.2004 . ПМК 6729642 . ПМИД 15329389 .
- ^ Перейти обратно: а б Мураками Х., Мураками С. (август 2007 г.). «Рецепторы серотонина антагонистически модулируют продолжительность жизни Caenorhabditis elegans» . Стареющая клетка . 6 (4): 483–488. дои : 10.1111/j.1474-9726.2007.00303.x . ПМИД 17559503 . S2CID 8345654 .
- ^ Каплан Д.Д., Циммерманн Г., Суяма К., Мейер Т., Скотт MP (июль 2008 г.). «ГТФаза семейства нуклеостеминов, NS3, действует в серотонинергических нейронах, регулируя передачу сигналов инсулина и контролируя размер тела» . Гены и развитие . 22 (14): 1877–1893. дои : 10.1101/gad.1670508 . ПМЦ 2492735 . ПМИД 18628395 .
- ^ Руауд А.Ф., Thummel CS (июль 2008 г.). «Серотонин и инсулиновая сигнализация объединяются, чтобы контролировать рост дрозофилы» . Гены и развитие . 22 (14): 1851–1855. дои : 10.1101/gad.1700708 . ПМЦ 2735276 . ПМИД 18628391 .
- ^ Дэвидсон С., Проконов Д., Талер М., Мааян Р., Харелл Д., Гил-Ад И. и др. (февраль 2009 г.). «Влияние воздействия селективных ингибиторов обратного захвата серотонина внутриутробно на рост плода: потенциальная роль осей IGF-I и HPA» . Педиатрические исследования . 65 (2): 236–241. дои : 10.1203/PDR.0b013e318193594a . ПМИД 19262294 .
- ^ Бен Арус Дж., Лаффон С., Шатене Д. (октябрь 2009 г.). Брезина В. (ред.). «Молекулярные и сенсорные основы пищевого поведения двух состояний C. elegans» . ПЛОС ОДИН . 4 (10): е7584. Бибкод : 2009PLoSO...4.7584B . дои : 10.1371/journal.pone.0007584 . ПМК 2762077 . ПМИД 19851507 .
- ^ Сзе Ю., Виктор М., Лоер С., Ши Ю., Рувкун Г. (февраль 2000 г.). «Дефекты пищевой и метаболической передачи сигналов у мутанта Caenorhabditis elegans, синтезирующего серотонин». Природа . 403 (6769): 560–564. Бибкод : 2000Natur.403..560S . дои : 10.1038/35000609 . ПМИД 10676966 . S2CID 4394553 .
- ^ Мураками Х., Бессингер К., Хеллманн Дж., Мураками С. (июль 2008 г.). «Манипулирование сигналом серотонина подавляет раннюю фазу поведенческого старения у Caenorhabditis elegans». Нейробиология старения . 29 (7): 1093–1100. doi : 10.1016/j.neurobiolaging.2007.01.013 . ПМИД 17336425 . S2CID 37671716 .
- ^ Коте Ф., Тевено Э., Флиньи С., Фромс Ю., Дармон М., Рипош М.А. и др. (ноябрь 2003 г.). «Нарушение ненейронального гена tph1 демонстрирует важность периферического серотонина в сердечной функции» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (23): 13525–13530. Бибкод : 2003PNAS..10013525C . дои : 10.1073/pnas.2233056100 . ПМЦ 263847 . ПМИД 14597720 .
- ^ Аларкон Дж., Сид Э., Лилло Л., Сеспедеса К., Агила С., Альдерете Дж.Б. (2008). «Биотрансформация производных индола мицелиальными культурами» . Zeitschrift Fur Naturforschung. C, Журнал биологических наук . 63 (1–2): 82–84. дои : 10.1515/znc-2008-1-215 . ПМИД 18386493 . S2CID 29472174 .
- ^ Сисмает Х.Дж., Голуч Э.Д. (июнь 2018 г.). «Электрохимические исследования поведения микробных сообществ» . Ежегодный обзор аналитической химии . 11 (1). Годовые обзоры : 441–461. Бибкод : 2018ARAC...11..441S . doi : 10.1146/annurev-anchem-061417-125627 . ПМИД 29490192 . S2CID 3632265 . п. 449:
Таблица 1. Соответствующие пики потенциала для различных электроактивных биомолекул, которые производятся или потребляются микробами, о которых сообщается в литературе. а ... Серотонин | Оксид индия и олова | +0,67 | 66
- ^ Хенсон Дж. М., Батлер М. Дж., Дэй AW (1999). «ТЕМНАЯ СТОРОНА МИЦЕЛИЯ: меланины фитопатогенных грибов». Ежегодный обзор фитопатологии . 37 (1). Годовые обзоры : 447–471. дои : 10.1146/annurev.phyto.37.1.447 . ПМИД 11701831 .
- ^ Энтони М. (декабрь 1984 г.). «Антагонисты серотонина». Австралийский и новозеландский медицинский журнал . 14 (6): 888–895. дои : 10.1111/j.1445-5994.1984.tb03802.x . ПМИД 6398056 . S2CID 28327178 .
- ^ Эрспамер V (декабрь 1954 г.). «Фармакология индолалкиламинов». Фармакологические обзоры . 6 (4): 425–487. ПМИД 13236482 .
- ^ Негри Л (2006). «[Витторио Эрспамер (1909–1999)]» . Медицина сквозь века . 18 (1): 97–113. ПМИД 17526278 .
- ^ Раппорт М.М., Грин А.А., Страница IH (декабрь 1948 г.). «Сывороточный сосудосуживающий препарат серотонин; выделение и характеристика» . Журнал биологической химии . 176 (3): 1243–1251. дои : 10.1016/S0021-9258(18)57137-4 . ПМИД 18100415 .
- ^ Фельдберг В., Тох CC (февраль 1953 г.). «Распределение 5-гидрокситриптамина (серотонина, энтерамина) в стенке пищеварительного тракта» . Журнал физиологии . 119 (2–3): 352–362. дои : 10.1113/jphysicalol.1953.sp004850 . ПМЦ 1392800 . ПМИД 13035756 .
- ^ SciFinder - Подробности о веществе серотонина. Доступ (4 ноября 2012 г.). [ нужна полная цитата ]
- ^ Тварог Б.М., Пейдж IH (октябрь 1953 г.). «Содержание серотонина в тканях и моче некоторых млекопитающих и метод его определения» . Американский журнал физиологии . 175 (1): 157–161. дои : 10.1152/ajplegacy.1953.175.1.157 . ПМИД 13114371 .
- ^ Страница IH (июль 1954 г.). «Серотонин (5-гидрокситриптамин)». Физиологические обзоры . 34 (3). Американское физиологическое общество : 563–588. дои : 10.1152/physrev.1954.34.3.563 . ПМИД 13185755 .
Дальнейшее чтение [ править ]
- Гуткнехт Л., Джейкоб С., Штробель А., Кригебаум С., Мюллер Дж., Цзэн Ю. и др. (июнь 2007 г.). «Вариации гена триптофангидроксилазы-2 влияют на черты личности и расстройства, связанные с эмоциональной дисрегуляцией» . Международный журнал нейропсихофармакологии . 10 (3): 309–320. дои : 10.1017/S1461145706007437 . ПМИД 17176492 .
Внешние ссылки [ править ]
- МС-спектр 5-гидрокситриптамина
- Серотонин связан с белками в PDB
- PsychoTropicalResearch Обширные обзоры серотонинергических препаратов и серотонинового синдрома.
- Молекула месяца: серотонин из Бристольского университета
- 60-секундный психолог: это несправедливо! У меня низкий уровень серотонина , Scientific American
- Интерпретация теста на серотонин в ClinLab Navigator .