Нейрон
| Нейрон | |
|---|---|
 Анатомия мультиполярного нейрона | |
| Идентификаторы | |
| МеШ | D009474 |
| НейроЛекс ID | sao1417703748 |
| ТА98 | A14.0.00.002 |
| ТД | Х2.00.06.1.00002 |
| ФМА | 54527 |
| Анатомические термины нейроанатомии | |
В нервной системе нейрон через , нейрон или нервная клетка — это электрически возбудимая клетка , которая излучает электрические сигналы, называемые потенциалами действия, нейронную сеть . Нейроны общаются с другими клетками через синапсы , которые представляют собой специализированные соединения, которые обычно используют незначительное количество химических нейротрансмиттеров для передачи электрического сигнала от пресинаптического нейрона к клетке-мишени через синаптическую щель.
Нейроны — основные компоненты нервной ткани всех животных, кроме губок и плакозоа . Неживотные, такие как растения и грибы, не имеют нервных клеток. Молекулярные данные свидетельствуют о том, что способность генерировать электрические сигналы впервые появилась в эволюции примерно 700–800 миллионов лет назад, в тонианский период. Предшественниками нейронов были пептидергические секреторные клетки. В конечном итоге они получили новые генные модули, которые позволили клеткам создавать постсинаптические каркасы и ионные каналы, генерирующие быстрые электрические сигналы. Способность генерировать электрические сигналы была ключевым нововведением в эволюции нервной системы. [1]
Нейроны обычно делятся на три типа в зависимости от их функции. Сенсорные нейроны реагируют на раздражители, такие как прикосновение, звук или свет, которые влияют на клетки органов чувств , и посылают сигналы в спинной или головной мозг . Мотонейроны получают сигналы от головного и спинного мозга, чтобы контролировать все, начиная с мышечных сокращений. [2] к железистому выходу . Интернейроны соединяют нейроны с другими нейронами в той же области головного или спинного мозга. Когда несколько нейронов функционально связаны друг с другом, они образуют так называемую нейронную цепь .
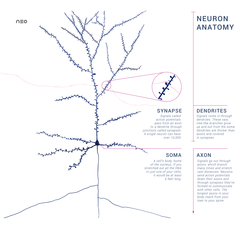
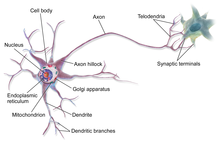
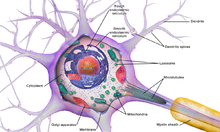
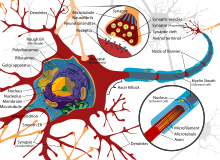
Нейроны — это особые клетки, которые состоят из некоторых структур, общих для всех других эукариотических клеток, таких как тело клетки (сома), ядро, гладкая и шероховатая эндоплазматическая сеть , аппарат Гольджи , митохондрии и другие клеточные компоненты. [3] Кроме того, нейроны имеют другие уникальные структуры, такие как дендриты и одиночный аксон . [3] Сома представляет собой компактную структуру, а аксоны и дендриты представляют собой нити, выходящие из сомы. Дендриты обычно сильно ветвятся и простираются на несколько сотен микрометров от сомы. Аксон покидает сому в месте вздутия, называемого аксонным бугорком , и проходит расстояние до 1 метра у людей или более у других видов. Он разветвляется, но обычно сохраняет постоянный диаметр. На самом дальнем конце ветвей аксона находятся окончания аксона , где нейрон может передавать сигнал через синапс в другую клетку. Нейроны могут не иметь дендритов или аксонов. Термин «нейрит» используется для описания дендрита или аксона, особенно когда клетка недифференцирована .
Большинство нейронов получают сигналы через дендриты и сому и отправляют сигналы по аксону. В большинстве синапсов сигналы переходят от аксона одного нейрона к дендриту другого. Однако синапсы могут соединять аксон с другим аксоном или дендрит с другим дендритом. Процесс передачи сигналов частично электрический, частично химический. Нейроны электрически возбудимы из-за поддержания градиентов напряжения на их мембранах . Если напряжение изменяется на достаточно большую величину в течение короткого интервала времени, нейрон генерирует импульс по принципу «все или ничего», электрохимический называемый потенциалом действия . Этот потенциал быстро распространяется по аксону и по мере достижения активирует синаптические связи. Синаптические сигналы могут быть возбуждающими или тормозящими , увеличивая или уменьшая суммарное напряжение, достигающее сомы.
В большинстве случаев нейроны генерируются нервными стволовыми клетками во время развития мозга и в детстве. Нейрогенез в большинстве областей мозга в значительной степени прекращается во взрослом возрасте.
Нервная система [ править ]
Нейроны являются основными компонентами нервной системы наряду с глиальными клетками , которые обеспечивают ей структурную и метаболическую поддержку. [4] Нервная система состоит из центральной нервной системы , включающей головной и спинной мозг , и периферической нервной системы , включающей вегетативную , кишечную и соматическую нервные системы . [5] У позвоночных большинство нейронов принадлежит центральной нервной системе , но некоторые из них расположены в периферических ганглиях , а многие сенсорные нейроны расположены в органах чувств, таких как сетчатка и улитка .
Аксоны могут объединяться в пучки , образующие нервы периферической нервной системы (как нити проводов составляют кабели). Пучки аксонов в центральной нервной системе называются трактами .
Анатомия и гистология [ править ]
Нейроны узкоспециализированы для обработки и передачи клеточных сигналов. Учитывая разнообразие функций, выполняемых в разных отделах нервной системы, существует большое разнообразие их формы, размеров и электрохимических свойств. Например, сома нейрона может иметь от 4 до 100 микрометров . диаметр [6]
- Сома – тело нейрона. Поскольку он содержит ядро , большая часть синтеза белка здесь происходит . Ядро может иметь диаметр от 3 до 18 микрометров. [7]
- Дендриты . нейрона представляют собой клеточные расширения со многими ответвлениями Эту общую форму и структуру метафорически называют дендритным деревом. Именно здесь большая часть входных данных в нейрон происходит через дендритный шип .
- Аксон представляет собой более тонкий, похожий на кабель выступ, длина которого может превышать диаметр сомы в десятки, сотни или даже десятки тысяч раз. Аксон в первую очередь передает нервные сигналы от сомы и возвращает к ней некоторые виды информации. Многие нейроны имеют только один аксон, но этот аксон может (и обычно будет) подвергаться обширному разветвлению, обеспечивая связь со многими клетками-мишенями. Часть аксона, где он выходит из сомы, называется аксонным бугорком . Помимо того, что аксонный бугорок является анатомической структурой, он также имеет наибольшую плотность потенциалзависимых натриевых каналов . Это делает его наиболее легко возбуждаемой частью нейрона и зоной инициации спайков аксона. В электрофизиологическом отношении он имеет наиболее отрицательный пороговый потенциал .
- Хотя аксон и аксонный бугорок обычно участвуют в оттоке информации, эта область также может получать входные данные от других нейронов.
- Терминал аксона находится на конце аксона, наиболее удаленном от сомы, и содержит синапсы . Синаптические бутоны представляют собой специализированные структуры, в которых химические вещества -нейромедиаторы высвобождаются для связи с целевыми нейронами. Помимо синаптических бутонов на конце аксона, нейрон может иметь проходные бутоны, расположенные по длине аксона.
Принятый взгляд на нейрон приписывает его различным анатомическим компонентам определенные функции; однако дендриты и аксоны часто действуют вопреки своей так называемой основной функции. [8]
Аксоны и дендриты в центральной нервной системе обычно имеют толщину всего около одного микрометра, тогда как некоторые в периферической нервной системе намного толще. Сома обычно имеет диаметр около 10–25 микрометров и часто ненамного больше содержащегося в ней клеточного ядра. человека Самый длинный аксон мотонейрона может достигать длины более метра и достигать от основания позвоночника до пальцев ног.
Сенсорные нейроны могут иметь аксоны, идущие от пальцев ног до заднего столба спинного мозга, длина которых у взрослых превышает 1,5 метра. У жирафов одиночные аксоны длиной несколько метров проходят по всей длине шеи. Большая часть того, что известно о функции аксонов, получено в результате изучения гигантского аксона кальмара , идеального экспериментального препарата из-за его относительно огромных размеров (толщина 0,5–1 миллиметр, длина несколько сантиметров).
Полностью дифференцированные нейроны постоянно постмитотические. [9] однако стволовые клетки, присутствующие во взрослом мозге, могут регенерировать функциональные нейроны на протяжении всей жизни организма (см. нейрогенез ). Астроциты звездчатой формы — глиальные клетки . Было замечено, что они превращаются в нейроны благодаря своей характеристике плюрипотентности , подобной стволовым клеткам .
Мембрана [ править ]
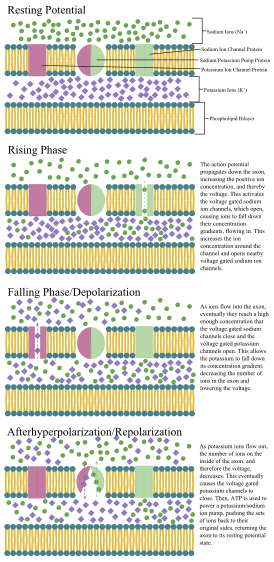
Как и все клетки животных, тело каждого нейрона окружено плазматической мембраной — двойным слоем липидных молекул, в который встроено множество типов белковых структур. [10] Липидный бислой является мощным электрическим изолятором , но в нейронах многие белковые структуры, встроенные в мембрану, электрически активны. К ним относятся ионные каналы, которые позволяют электрически заряженным ионам проходить через мембрану, и ионные насосы, которые химически транспортируют ионы с одной стороны мембраны на другую. Большинство ионных каналов проницаемы только для определенных типов ионов. Некоторые ионные каналы являются потенциалозависимыми , что означает, что их можно переключать между открытым и закрытым состояниями, изменяя разность потенциалов на мембране. Другие являются химически закрытыми, то есть их можно переключать между открытым и закрытым состояниями за счет взаимодействия с химическими веществами, которые диффундируют через внеклеточную жидкость. Ионные калий включают натрий , кальций , хлорид и . материалы Взаимодействие между ионными каналами и ионными насосами приводит к разнице потенциалов на мембране, обычно чуть менее 1/10 вольт в исходном состоянии. Это напряжение выполняет две функции: во-первых, оно обеспечивает источник питания для ряда зависимых от напряжения белковых механизмов, встроенных в мембрану; во-вторых, он обеспечивает основу для передачи электрического сигнала между различными частями мембраны.
Гистология и внутреннее строение [ править ]


Многочисленные микроскопические комки, называемые тельцами Ниссля (или веществом Ниссля), можно увидеть, когда тела нервных клеток окрашиваются базофильным («любящим основания») красителем. Эти структуры состоят из шероховатой эндоплазматической сети и связанной с ней рибосомальной РНК . Названные в честь немецкого психиатра и невропатолога Франца Ниссля (1860–1919), они участвуют в синтезе белка, и их известность можно объяснить тем фактом, что нервные клетки очень метаболически активны. Базофильные красители, такие как анилин или (слабый) гематоксилин. [11] выделяют отрицательно заряженные компоненты и таким образом связываются с фосфатным остовом рибосомальной РНК.
Тело клетки нейрона поддерживается сложной сеткой структурных белков, называемых нейрофиламентами , которые вместе с нейротрубочками (нейрональными микротрубочками) собираются в более крупные нейрофибриллы. [12] Некоторые нейроны также содержат пигментные гранулы, такие как нейромеланин (коричнево-черный пигмент, являющийся побочным продуктом синтеза катехоламинов ) и липофусцин (желтовато-коричневый пигмент), оба из которых накапливаются с возрастом. [13] [14] [15] важными для функции нейронов, являются актин и тубулин микротрубочек Другими структурными белками , . β-тубулин класса III обнаруживается почти исключительно в нейронах. Актин преимущественно обнаруживается на кончиках аксонов и дендритов во время развития нейронов. Там динамику актина можно модулировать посредством взаимодействия с микротрубочками. [16]
Аксоны и дендриты имеют разные внутренние структурные характеристики. Типичные аксоны почти никогда не содержат рибосом , за исключением некоторых в начальном сегменте. Дендриты содержат гранулярную эндоплазматическую сеть или рибосомы, количество которых уменьшается по мере увеличения расстояния от тела клетки.
Классификация [ править ]

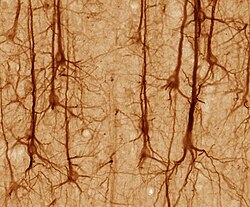
Нейроны различаются по форме и размеру, и их можно классифицировать по морфологии и функциям. [18] Анатом Камилло Гольджи сгруппировал нейроны на два типа; тип I с длинными аксонами, используемый для перемещения сигналов на большие расстояния, и тип II с короткими аксонами, которые часто можно спутать с дендритами. Клетки типа I можно дополнительно классифицировать по расположению сомы. Основная морфология нейронов I типа, представленных спинальными мотонейронами , состоит из тела клетки, называемого сомой, и длинного тонкого аксона, покрытого миелиновой оболочкой . Дендритное дерево окружает тело клетки и получает сигналы от других нейронов. Конец аксона имеет ветвящиеся окончания аксона , которые высвобождают нейротрансмиттеры в щель, называемую синаптической щелью , между терминалями и дендритами следующего нейрона. [ нужна ссылка ]
классификация Структурная
Полярность [ править ]
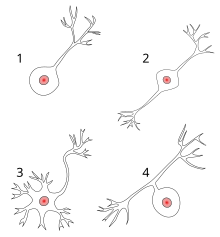
1 униполярный нейрон
2 Биполярный нейрон
3 Мультиполярный нейрон
4 Псевдоуниполярный нейрон
Большинство нейронов можно анатомически охарактеризовать как: [19]
- Униполярный : одиночный процесс. Униполярные клетки представляют собой исключительно сенсорные нейроны. Их дендриты получают сенсорную информацию, иногда непосредственно от самого стимула. Клеточные тела униполярных нейронов всегда находятся в ганглиях. Сенсорная рецепция является периферической функцией, поэтому тело клетки находится на периферии, хотя и ближе к ЦНС в ганглии. Аксон выступает из окончаний дендритов мимо тела клетки в ганглии и в центральную нервную систему.
- Биполярный : 1 аксон и 1 дендрит. Они встречаются главным образом в обонятельном эпителии и в составе сетчатки.
- Мультиполярный : 1 аксон и 2 или более дендритов.
- Гольджи I : нейроны с длинновыступающими аксональными отростками; примерами являются пирамидные клетки, клетки Пуркинье и клетки переднего рога.
- Гольджи II : нейроны, аксональные отростки которых проецируются локально; лучший пример - гранулярная клетка
- Анаксонический : аксон невозможно отличить от дендритов.
- Псевдоуниполярный : 1 отросток, который затем служит одновременно аксоном и дендритом.
Другое [ править ]
Некоторые уникальные типы нейронов можно идентифицировать по их расположению в нервной системе и особой форме. Некоторые примеры: [ нужна ссылка ]
- Корзинчатые клетки , интернейроны, образующие плотное сплетение терминалей вокруг сомы клеток-мишеней, встречаются в коре и мозжечке.
- Клетки Бетца , крупные мотонейроны
- Клетки Лугаро , интернейроны мозжечка
- Средние шипистые нейроны , большинство нейронов полосатого тела.
- Клетки Пуркинье , огромные нейроны мозжечка, тип мультиполярного нейрона Гольджи I.
- Пирамидные клетки , нейроны с треугольной сомой, тип Гольджи I.
- Клетки шиповника , уникальные тормозные нейроны человека, которые соединяются с пирамидными клетками.
- Клетки Реншоу , нейроны, оба конца которых связаны с альфа-мотонейронами.
- Униполярные щеточные клетки , интернейроны с уникальным дендритом, заканчивающимся щеткообразным пучком.
- Гранульные клетки , тип нейрона Гольджи II.
- Клетки переднего рога , мотонейроны, расположенные в спинном мозге.
- Веретенообразные клетки , интернейроны, которые соединяют широко разнесенные области мозга.
Функциональная классификация
Направление [ править ]
- Афферентные нейроны передают информацию от тканей и органов в центральную нервную систему и также называются сенсорными нейронами .
- Эфферентные нейроны (мотонейроны) передают сигналы от центральной нервной системы к эффекторным клеткам.
- Интернейроны соединяют нейроны в определенных областях центральной нервной системы.
Афферентные и эфферентные также обычно относятся к нейронам, которые, соответственно, передают информацию в мозг или отправляют информацию из него.
Действие на другие нейроны [ править ]
Нейрон влияет на другие нейроны, высвобождая нейромедиатор, который связывается с химическими рецепторами . Воздействие на постсинаптический нейрон определяется типом активируемого рецептора, а не пресинаптическим нейроном или нейромедиатором. Нейромедиатор можно рассматривать как ключ, а рецептор — как замок: один и тот же нейромедиатор может активировать несколько типов рецепторов. Рецепторы можно широко классифицировать как возбуждающие (вызывающие увеличение скорости срабатывания), тормозящие (вызывающие снижение скорости срабатывания) или модуляторные (вызывающие долговременные эффекты, не связанные напрямую с частотой срабатывания). [ нужна ссылка ]
Два наиболее распространенных (90%+) нейротрансмиттера в мозге, глутамат и ГАМК , имеют в значительной степени согласованное действие. Глутамат действует на несколько типов рецепторов и оказывает возбуждающее действие на ионотропные рецепторы и модулирующее действие на метаботропные рецепторы . Аналогичным образом ГАМК действует на несколько типов рецепторов, но все они обладают ингибирующим действием (по крайней мере, у взрослых животных). Из-за такой последовательности нейробиологи обычно называют клетки, выделяющие глутамат, «возбуждающими нейронами», а клетки, выделяющие ГАМК, «тормозными нейронами». Некоторые другие типы нейронов оказывают последовательное воздействие, например, «возбуждающие» мотонейроны спинного мозга, выделяющие ацетилхолин , и «тормозные» спинальные нейроны , выделяющие глицин . [ нужна ссылка ]
Различие между возбуждающими и тормозными нейромедиаторами не является абсолютным. Скорее, это зависит от класса химических рецепторов, присутствующих на постсинаптическом нейроне. В принципе, один нейрон, высвобождающий один нейромедиатор, может оказывать возбуждающее воздействие на одни цели, тормозящее воздействие на другие и модулирующее воздействие на другие. Например, фоторецепторные клетки сетчатки постоянно выделяют нейромедиатор глутамат в отсутствие света. Так называемые OFF- биполярные клетки , как и большинство нейронов, возбуждаются высвобождаемым глутаматом. Однако соседние нейроны-мишени, называемые биполярными клетками ON, вместо этого ингибируются глутаматом, поскольку у них отсутствуют типичные ионотропные рецепторы глутамата , и вместо этого они экспрессируют класс ингибирующих метаботропных рецепторов глутамата. [20] При наличии света фоторецепторы перестают выделять глутамат, который освобождает биполярные клетки ON от торможения, активируя их; это одновременно снимает возбуждение с выключенных биполярных клеток, заставляя их замолчать. [ нужна ссылка ]
Определить тип тормозящего эффекта, который пресинаптический нейрон будет оказывать на постсинаптический нейрон, можно на основе белков, которые экспрессирует пресинаптический нейрон. Нейроны, экспрессирующие парвальбумин, обычно ослабляют выходной сигнал постсинаптического нейрона зрительной коры , тогда как нейроны, экспрессирующие соматостатин, обычно блокируют дендритные входы к постсинаптическому нейрону. [21]
Характер разряда [ править ]
Нейроны обладают внутренними электрореактивными свойствами, такими как собственные паттерны колебаний трансмембранного напряжения . [22] Итак, нейроны можно классифицировать по их электрофизиологическим характеристикам:
- Тоник или обычный прием. Некоторые нейроны обычно постоянно (тонически) активны и обычно срабатывают с постоянной частотой. Пример: интернейроны в нейростриатуме .
- Фазический или взрывной . Нейроны, которые активируются очередями, называются фазическими.
- Быстродействующий. Некоторые нейроны отличаются высокой скоростью срабатывания, например, некоторые типы корковых тормозных интернейронов, клетки бледного шара , ганглиозные клетки сетчатки . [23] [24]
Нейромедиатор [ править ]
Нейротрансмиттеры — это химические посланники, передаваемые от одного нейрона к другому нейрону или к мышечной или железистой клетке .
- Холинергические нейроны – ацетилхолин. Ацетилхолин высвобождается из пресинаптических нейронов в синаптическую щель. Он действует как лиганд как для лиганд-управляемых ионных каналов, так и для метаботропных (GPCR) мускариновых рецепторов . Никотиновые рецепторы представляют собой пентамерные лиганд-управляемые ионные каналы, состоящие из альфа- и бета-субъединиц, которые связывают никотин . Связывание лиганда открывает канал, вызывая приток Na. + деполяризации и увеличивает вероятность пресинаптического высвобождения нейромедиатора. Ацетилхолин синтезируется из холина и ацетилкофермента А.
- Адренергические нейроны – норадреналин. Норадреналин (норадреналин) высвобождается из большинства постганглионарных нейронов симпатической нервной системы на два набора GPCR: альфа-адренорецепторы и бета-адренорецепторы . Норадреналин является одним из трех распространенных нейротрансмиттеров катехоламинов и наиболее распространенным из них в периферической нервной системе ; как и другие катехоламины, он синтезируется из тирозина .
- ГАМКергические нейроны – гамма-аминомасляная кислота . ГАМК является одним из двух нейроингибиторов центральной нервной системы (ЦНС), наряду с глицином. ГАМК имеет гомологичную функцию с АХ , открывая анионные каналы, которые позволяют Cl − ионы проникают в постсинаптический нейрон. кл. − вызывает гиперполяризацию внутри нейрона, уменьшая вероятность срабатывания потенциала действия, поскольку напряжение становится более отрицательным (для срабатывания потенциала действия должен быть достигнут порог положительного напряжения). ГАМК синтезируется из глутаматных нейротрансмиттеров с помощью фермента глутаматдекарбоксилазы .
- Глутаматергические нейроны – глутамат. Глутамат является одним из двух основных нейромедиаторов возбуждающих аминокислот, наряду с аспартатом . Глутаматные рецепторы относятся к одной из четырех категорий, три из которых представляют собой лиганд-управляемые ионные каналы, а одна из них представляет собой рецептор, связанный с G-белком (часто называемый GPCR).
- АМРА- и каинатные рецепторы функционируют как катионные каналы, проницаемые для Na. + катионные каналы, обеспечивающие быструю возбуждающую синаптическую передачу.
- NMDA- рецепторы — еще один катионный канал, более проницаемый для кальция. 2+ . Функция NMDA-рецепторов зависит от связывания глицинового рецептора в качестве коагониста внутри поры канала. Рецепторы NMDA не функционируют без присутствия обоих лигандов.
- Метаботропные рецепторы, GPCR модулируют синаптическую передачу и постсинаптическую возбудимость.
- Глутамат может вызвать эксайтотоксичность, когда приток крови к мозгу прерывается, что приводит к повреждению мозга . Когда кровоток подавляется, глутамат высвобождается из пресинаптических нейронов, вызывая большую активацию рецепторов NMDA и AMPA, чем обычно, вне стрессовых условий, что приводит к повышению уровня кальция. 2+ и На + попадание в постсинаптический нейрон и повреждение клеток. Глутамат синтезируется из аминокислоты глютамин с помощью фермента глутаматсинтазы .
- Дофаминергические нейроны — дофамин . Дофамин является нейротрансмиттером, который действует на Gs-сопряженные рецепторы типа D1 (D1 и D5), которые увеличивают цАМФ и PKA, и рецепторы типа D2 (D2, D3 и D4), которые активируют Gi-связанные рецепторы, которые уменьшают цАМФ и PKA. Дофамин связан с настроением и поведением и модулирует как пре-, так и постсинаптическую нейротрансмиссию. Потеря дофаминовых нейронов в черной субстанции связана с болезнью Паркинсона . Дофамин синтезируется из аминокислоты тирозина . Тирозин катализируется в леводопа (или L-ДОФА ) тирозингидроксилазой ароматических аминокислот , а затем леводопа превращается в дофамин под действием декарбоксилазы .
- Серотонинергические нейроны — серотонин . Серотонин (5-гидрокситриптамин, 5-НТ) может действовать как возбуждающе, так и тормозно. Из четырех классов рецепторов 5-HT 3 представляют собой GPCR и 1 — катионный канал, управляемый лигандом. Серотонин синтезируется из триптофана триптофангидроксилазой , а затем далее декарбоксилазой. Недостаток 5-HT в постсинаптических нейронах связан с депрессией. препараты, блокирующие пресинаптический переносчик серотонина Для лечения используются , такие как Прозак и Золофт .
- Пуринергические нейроны — АТФ. АТФ является нейромедиатором, действующим как на лиганд-управляемые ионные каналы ( рецепторы P2X ), так и на рецепторы GPCR ( P2Y ). Однако АТФ наиболее известен как котрансмиттер . Такая пуринергическая передача сигналов также может быть опосредована другими пуринами , такими как аденозин , который особенно действует на рецепторы P2Y.
- Гистаминергические нейроны — гистамин . Гистамин является моноаминовым нейротрансмиттером и нейромодулятором . находятся в туберомаммиллярном ядре гипоталамуса Нейроны, продуцирующие гистамин , . [25] Гистамин участвует в возбуждении и регуляции режима сна/бодрствования.
Мультимодельная классификация
пытается С 2012 года сообщество клеточных и вычислительных нейробиологов разработать универсальную классификацию нейронов, которая будет применяться ко всем нейронам мозга, а также к представителям разных видов. Это делается путем рассмотрения трех основных качеств всех нейронов: электрофизиологии, морфологии и индивидуального транскриптома клеток. Помимо универсальности, эта классификация имеет то преимущество, что позволяет также классифицировать астроциты. Метод, называемый патч-секвенированием , при котором можно измерить все три качества одновременно, широко используется Институтом наук о мозге Аллена . [26] В 2023 году в результате международного сотрудничества исследователей, использующих самые передовые подходы молекулярной биологии, был создан комплексный атлас клеток взрослого и развивающегося человеческого мозга на транскрипционном, эпигенетическом и функциональном уровнях. [27]
Связь [ править ]


Нейроны общаются друг с другом через синапсы , где окончание аксона одной клетки контактирует с дендритом, сомой или, реже, аксоном другого нейрона. Нейроны, такие как клетки Пуркинье в мозжечке, могут иметь более 1000 дендритных ветвей, образующих связи с десятками тысяч других клеток; другие нейроны, например магноцеллюлярные нейроны супраоптического ядра , имеют только один или два дендрита, каждый из которых получает тысячи синапсов.
Синапсы могут быть возбуждающими или тормозящими, повышающими или понижающими активность целевого нейрона соответственно. Некоторые нейроны также общаются через электрические синапсы, которые представляют собой прямые электропроводящие соединения между клетками. [28]
Когда потенциал действия достигает окончания аксона, он открывает потенциалзависимые кальциевые каналы , позволяя ионам кальция проникать в терминаль. Кальций заставляет синаптические пузырьки, наполненные молекулами нейромедиаторов, сливаться с мембраной, высвобождая их содержимое в синаптическую щель. Нейромедиаторы диффундируют через синаптическую щель и активируют рецепторы постсинаптического нейрона. Высокий уровень цитозольного кальция в окончаниях аксонов запускает поглощение кальция митохондриями, что, в свою очередь, активирует энергетический метаболизм митохондрий для производства АТФ для поддержания непрерывной нейротрансмиссии. [29]
Аутапс — это синапс, в котором аксон нейрона соединяется с собственными дендритами.
Человеческий мозг имеет примерно 8,6 х 10 10 (восемьдесят шесть миллиардов) нейронов. [30] [31] Каждый нейрон имеет в среднем 7000 синаптических связей с другими нейронами. Подсчитано, что мозг трехлетнего ребенка имеет около 10 15 синапсы (1 квадриллион). Это число снижается с возрастом и стабилизируется к взрослому возрасту. Оценки для взрослого варьируются от 10 14 до 5 х 10 14 синапсы (от 100 до 500 триллионов). [32]
Неэлектрохимическая передача сигналов [ править ]
Исследования показывают, что помимо электрической и химической передачи сигналов нейроны мозга здорового человека также могут общаться посредством:
- сила, создаваемая увеличением дендритных шипов [33]
- перенос белков – белки, транспортируемые транснейронально (TNTP) [34] [35]
Они также могут модулироваться под воздействием окружающей среды и гормонов, выделяемых из других частей организма. [36] на которые могут более или менее непосредственно влиять нейроны. Это также относится к нейротрофинам, таким как BDNF . Микробиом кишечника также связан с мозгом. [37] Нейроны также взаимодействуют с микроглией , основными иммунными клетками мозга, через специальные контактные площадки, называемые «соматическим соединением». Эти связи позволяют микроглии постоянно контролировать и регулировать функции нейронов и при необходимости оказывать нейропротекцию. [38]
распространения действия потенциалов Механизмы
В 1937 году Джон Закари Янг предположил, что гигантский аксон кальмара можно использовать для изучения электрических свойств нейронов. [39] Он больше человеческих нейронов, но похож на него, что облегчает изучение. Вставив электроды в аксоны гигантских кальмаров, были проведены точные измерения мембранного потенциала .
Клеточная мембрана аксона и сомы содержит потенциалзависимые ионные каналы, которые позволяют нейрону генерировать и распространять электрический сигнал (потенциал действия). Некоторые нейроны также генерируют подпороговые колебания мембранного потенциала . Эти сигналы генерируются и распространяются ионами, несущими заряд , включая натрий (Na + ), калий (К + ), хлорид (Cl − ) и кальций (Ca 2+ ) .
Несколько стимулов могут активировать нейрон, приводя к электрической активности, включая давление , растяжение, химические передатчики и изменения электрического потенциала на клеточной мембране. [40] Стимулы вызывают открытие определенных ионных каналов внутри клеточной мембраны, что приводит к потоку ионов через клеточную мембрану, изменяя мембранный потенциал. Нейроны должны сохранять определенные электрические свойства, определяющие их тип нейронов. [41]
Тонкие нейроны и аксоны требуют меньших метаболических затрат для производства и проведения потенциалов действия, но более толстые аксоны передают импульсы быстрее. оболочку Чтобы свести к минимуму метаболические затраты при сохранении быстрой проводимости, многие нейроны имеют изолирующую миелиновую вокруг своих аксонов. Оболочки образованы глиальными клетками: олигодендроцитами в ЦНС и шванновскими клетками в периферической нервной системе. Оболочка позволяет потенциалам действия перемещаться быстрее, чем в безмиелиновых аксонах того же диаметра, используя при этом меньше энергии. Миелиновая оболочка периферических нервов обычно проходит вдоль аксона участками длиной около 1 мм, перемежающимися непокрытыми узлами Ранвье , которые содержат высокую плотность потенциалзависимых ионных каналов. Рассеянный склероз — неврологическое заболевание, возникающее в результате демиелинизации аксонов центральной нервной системы.
Некоторые нейроны не генерируют потенциалы действия, а вместо этого генерируют градуированный электрический сигнал , который, в свою очередь, вызывает постепенное высвобождение нейромедиаторов. Такие нейроны без спайков обычно являются сенсорными нейронами или интернейронами, поскольку они не могут передавать сигналы на большие расстояния.
Нейронное кодирование [ править ]
Нейронное кодирование связано с тем, как сенсорная и другая информация представляется в мозге нейронами. Основная цель изучения нейронного кодирования — охарактеризовать взаимосвязь между стимулом и реакциями отдельных нейронов или ансамбля , а также взаимосвязь между электрической активностью нейронов внутри ансамбля. [42] Считается, что нейроны могут кодировать как цифровую , так и аналоговую информацию. [43]
Принцип «все или ничего» [ править ]
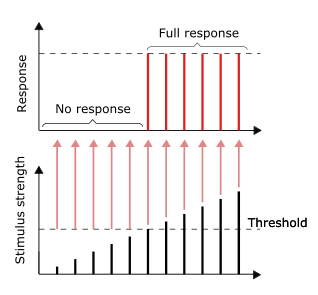
Проведение нервных импульсов является примером реакции «все или ничего» . Другими словами, если нейрон вообще реагирует, то он должен реагировать полностью. Большая интенсивность стимуляции, например, более яркое изображение или более громкий звук, не создает более сильный сигнал, но может увеличить частоту импульсов. [44] : 31 Рецепторы по-разному реагируют на раздражители. Медленно адаптирующиеся или тонические рецепторы реагируют на устойчивый стимул и производят постоянную частоту импульсов. Тонические рецепторы чаще всего реагируют на повышенную интенсивность стимула увеличением частоты срабатывания, обычно как степенную функцию стимула, отображаемую в зависимости от количества импульсов в секунду. Это можно сравнить с внутренним свойством света: для большей интенсивности определенной частоты (цвета) требуется больше фотонов, поскольку фотоны не могут стать «сильнее» для определенной частоты.
Другие типы рецепторов включают быстро адаптирующиеся или фазовые рецепторы, у которых возбуждение уменьшается или прекращается при постоянном стимуле; примеры включают кожу , при прикосновении к которой нейроны активируются, но если объект сохраняет равномерное давление, нейроны перестают активироваться. Нейроны кожи и мышц, реагирующие на давление и вибрацию, имеют дополнительные фильтрующие структуры, помогающие их функциям.
Одной из таких структур является тельце Пачини . Он имеет концентрические слои, подобные луковице, которые формируются вокруг окончания аксона. Когда оказывается давление и корпускула деформируется, механический стимул передается на аксон, который срабатывает. Если давление устойчиво, стимуляция прекращается; таким образом, эти нейроны обычно реагируют временной деполяризацией во время начальной деформации и снова, когда давление снимается, что заставляет корпускулу снова менять форму. Другие типы адаптации важны для расширения функции ряда других нейронов. [45]
Этимология и орфография [ править ]
Немецкий анатом Генрих Вильгельм Вальдейер ввел термин «нейрон» в 1891 году. [46] на основе древнегреческого νεῦρον нейрон «сухожилие, шнур, нерв». [47]
Слово было принято во французском языке с орфографическим нейроном . Это написание также использовалось многими авторами на английском языке. [48] но теперь оно стало редким в использовании в Америке и необычным в использовании в Великобритании. [49] [47]
История [ править ]

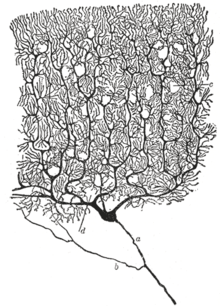
Место нейрона как основной функциональной единицы нервной системы было впервые признано в конце 19 века благодаря работам испанского анатома Сантьяго Рамона-и-Кахаля . [50]
Чтобы сделать структуру отдельных нейронов видимой, Рамон-и-Кахаль усовершенствовал процесс окрашивания серебром , разработанный Камилло Гольджи . [50] Усовершенствованный процесс включает в себя технику под названием «двойная пропитка», которая используется до сих пор.
В 1888 году Рамон-и-Кахаль опубликовал статью о мозжечке птиц. В этой статье он заявил, что не может найти доказательств наличия анастомозов между аксонами и дендритами, и назвал каждый нервный элемент «абсолютно автономным кантоном». [50] [46] Это стало известно как доктрина нейронов , один из центральных принципов современной нейробиологии . [50]
В 1891 году немецкий анатом Генрих Вильгельм Вальдейер написал весьма влиятельный обзор доктрины нейронов, в котором ввел термин «нейрон» для описания анатомо-физиологической единицы нервной системы. [51] [52]
Окрашивание импрегнацией серебром является полезным методом для нейроанатомических исследований, поскольку по неизвестным причинам оно окрашивает лишь небольшой процент клеток в ткани, обнажая полную микроструктуру отдельных нейронов без большого перекрытия с другими клетками. [53]
Учение о нейронах [ править ]

Учение о нейронах — это ныне фундаментальная идея о том, что нейроны являются основными структурными и функциональными единицами нервной системы. Теория была выдвинута Сантьяго Рамоном-и-Кахалем в конце 19 века. Он считал, что нейроны представляют собой отдельные клетки (не связанные в сеть), действующие как метаболически различные единицы.
Более поздние открытия внесли уточнения в эту доктрину. Например, глиальные клетки , которые не являются нейронами, играют важную роль в обработке информации. [54] Кроме того, электрические синапсы встречаются чаще, чем считалось ранее. [55] включающий прямые цитоплазматические связи между нейронами. На самом деле нейроны могут образовывать еще более тесные связи: гигантский аксон кальмара возникает в результате слияния нескольких аксонов. [56]
Рамон-и-Кахаль также постулировал закон динамической поляризации, который гласит, что нейрон получает сигналы от своих дендритов и тела клетки и передает их в виде потенциалов действия вдоль аксона в одном направлении: от тела клетки. [57] Закон динамической поляризации имеет важные исключения; дендриты могут служить синаптическим выходом нейронов [58] и аксоны могут получать синаптические входы. [59]
Компартментальное моделирование нейронов [ править ]
Хотя нейроны часто называют «фундаментальными единицами» мозга, они выполняют внутренние вычисления. Нейроны интегрируют входные данные внутри дендритов, и эта сложность теряется в моделях, которые предполагают, что нейроны являются фундаментальной единицей. Дендритные ветви можно моделировать как пространственные компартменты, активность которых связана с пассивными свойствами мембраны, но также может различаться в зависимости от входных сигналов от синапсов. Компартментальное моделирование дендритов особенно полезно для понимания поведения нейронов, которые слишком малы для регистрации с помощью электродов, как в случае с Drosophila melanogaster . [60]
Нейроны головного мозга [ править ]
Число нейронов в мозге резко варьируется от вида к виду. [61] По оценкам, у человека насчитывается 10–20 миллиардов нейронов в коре головного мозга и 55–70 миллиардов нейронов в мозжечке . [62] Напротив, нематода червь- Caenorhabditis elegans имеет всего 302 нейрона, что делает его идеальным модельным организмом , поскольку ученые смогли нанести на карту все его нейроны. Плодовая мушка Drosophila melanogaster , частый объект биологических экспериментов, имеет около 100 000 нейронов и демонстрирует множество сложных моделей поведения. Многие свойства нейронов, от типа используемых нейротрансмиттеров до состава ионных каналов, сохраняются у разных видов, что позволяет ученым изучать процессы, происходящие в более сложных организмах, в гораздо более простых экспериментальных системах.
Неврологические расстройства [ править ]
Эта статья нуждается в дополнительных цитатах для проверки . ( май 2018 г. ) |
Болезнь Шарко-Мари-Тута (ШМТ) — гетерогенное наследственное заболевание нервов ( нейропатия ), которое характеризуется потерей мышечной ткани и чувствительности к прикосновению, преимущественно в стопах и ногах, на поздних стадиях распространяется на кисти и руки. В настоящее время это неизлечимое заболевание является одним из наиболее распространенных наследственных неврологических заболеваний , которым страдают 36 из 100 000 человек. [63]
Болезнь Альцгеймера (БА), также известная как болезнь Альцгеймера , представляет собой нейродегенеративное заболевание , характеризующееся прогрессирующим ухудшением когнитивных функций , а также снижением повседневной активности и нейропсихиатрическими симптомами или поведенческими изменениями. [64] Наиболее ярким ранним симптомом является потеря кратковременной памяти ( амнезия ), которая обычно проявляется в виде незначительной забывчивости, которая становится все более выраженной по мере прогрессирования заболевания, при относительной сохранности старых воспоминаний. По мере прогрессирования расстройства когнитивные (интеллектуальные) нарушения распространяются на область речи ( афазия ), навыков движений ( апраксия ) и распознавания ( агнозия ), а такие функции, как принятие решений и планирование, нарушаются. [65] [66]
Болезнь Паркинсона (БП), также известная как Паркинсон , представляет собой дегенеративное заболевание центральной нервной системы, которое часто нарушает моторику и речь. [67] Болезнь Паркинсона принадлежит к группе состояний, называемых двигательными расстройствами . [68] Для него характерна ригидность мышц, тремор , замедление физических движений ( брадикинезия ), а в крайних случаях — потеря физических движений ( акинезия ). Первичные симптомы являются результатом снижения стимуляции моторной коры со стороны базальных ганглиев , обычно вызванного недостаточным образованием и действием дофамина, который вырабатывается в дофаминергических нейронах головного мозга. Вторичные симптомы могут включать когнитивную дисфункцию высокого уровня и легкие языковые проблемы. БП бывает как хроническим, так и прогрессирующим.
Миастения гравис – нервно-мышечное заболевание, приводящее к нестабильной мышечной слабости и утомляемости при выполнении простых действий. Слабость обычно вызвана циркулирующими антителами , которые блокируют рецепторы ацетилхолина в постсинаптическом нервно-мышечном соединении, подавляя стимулирующий эффект нейромедиатора ацетилхолина. Миастению лечат иммунодепрессантами , ингибиторами холинэстеразы и, в отдельных случаях, тимэктомией .
Демиелинизация [ править ]

Демиелинизация — процесс, характеризующийся постепенной потерей миелиновой оболочки, окружающей нервные волокна. Когда миелин разрушается, проводимость сигнала по нервам может быть значительно нарушена или потеряна, и нерв в конечном итоге увядает. Демиелинизация может поражать как центральную, так и периферическую нервную систему, способствуя различным неврологическим расстройствам, таким как рассеянный склероз , синдром Гийена-Барре и хроническая воспалительная демиелинизирующая полинейропатия . Хотя демиелинизация часто вызывается аутоиммунной реакцией, она также может быть вызвана вирусными инфекциями, нарушениями обмена веществ, травмой и приемом некоторых лекарств.
дегенерация Аксональная
Хотя большинство реакций на травму включают в себя приток кальция, способствующий повторному заживлению оторванных частей, повреждения аксонов первоначально приводят к острой аксональной дегенерации , которая представляет собой быстрое разделение проксимального и дистального концов, происходящее в течение 30 минут после повреждения. [69] Дегенерация сопровождается отеком аксолеммы и в конечном итоге приводит к образованию бусинок. Гранулярный распад аксонального цитоскелета и внутренних органелл происходит после деградации аксолеммы. Ранние изменения включают накопление митохондрий в паранодальных областях в месте повреждения. Эндоплазматическая сеть разрушается, митохондрии разбухают и в конечном итоге распадаются. Распад зависит от убиквитина и кальпаина протеаз (вызванный притоком ионов кальция), что позволяет предположить, что аксональная дегенерация является активным процессом, вызывающим полную фрагментацию. Этот процесс занимает около 24 часов в ПНС и дольше в ЦНС. Сигнальные пути, ведущие к дегенерации аксолеммы, неизвестны.
Нейрогенез [ править ]
Нейроны рождаются в процессе нейрогенеза , при котором нервные стволовые клетки делятся, образуя дифференцированные нейроны. Как только полностью дифференцированные нейроны сформированы, они больше не способны подвергаться митозу . Нейрогенез преимущественно происходит в эмбрионах большинства организмов.
Нейрогенез у взрослых может происходить, и исследования возраста нейронов человека показывают, что этот процесс происходит только для меньшинства клеток и что подавляющее большинство нейронов в неокортексе формируется до рождения и сохраняется без замены. Степень существования взрослого нейрогенеза у людей и его вклад в познание являются спорными: в 2018 году были опубликованы противоречивые отчеты. [70]
В организме содержатся различные типы стволовых клеток, которые способны дифференцироваться в нейроны. Исследователи нашли способ превратить клетки кожи человека в нервные клетки с помощью трансдифференцировки , при которой «клетки вынуждены приобретать новую идентичность». [71]
Во время нейрогенеза в мозге млекопитающих клетки-предшественники и стволовые клетки переходят от пролиферативных делений к дифференциативным. Эта прогрессия приводит к нейронам и глии, которые заселяют корковые слои. Эпигенетические модификации играют ключевую роль в регуляции экспрессии генов при дифференцировке нервных стволовых клеток и имеют решающее значение для определения судьбы клеток в развивающемся и взрослом мозге млекопитающих. Эпигенетические модификации включают метилирование цитозина ДНК с образованием 5-метилцитозина и деметилирование 5-метилцитозина . [72] Эти модификации имеют решающее значение для определения судьбы клеток в мозге развивающихся и взрослых млекопитающих. Метилирование ДНК-цитозина катализируется ДНК-метилтрансферазами (DNMT) . Деметилирование метилцитозина катализируется в несколько стадий ферментами ТЕТ , которые осуществляют окислительные реакции (например, превращение 5-метилцитозина в 5-гидроксиметилцитозин ), и ферментами пути эксцизионной репарации оснований ДНК (BER). [72]
На разных стадиях развития нервной системы млекопитающих используются два процесса репарации ДНК для восстановления двухцепочечных разрывов ДНК. Этими путями являются гомологичная рекомбинационная репарация, используемая в пролиферации нервных клеток-предшественников, и негомологичное соединение концов, используемое главным образом на более поздних стадиях развития. [73]
Межклеточная связь между развивающимися нейронами и микроглией также необходима для правильного нейрогенеза и развития мозга. [74]
Регенерация нервов [ править ]
Периферические аксоны могут вырасти заново, если их разорвать. [75] но один нейрон не может быть функционально заменен нейроном другого типа ( закон Ллинаса ). [22]
См. также [ править ]
Ссылки [ править ]
- ^ Найл, Себастьян Р.; Грау-Бове, Ксавье; Элек, Анамария; Наваррете, Кристина; Чианферони, Дамиано; Чива, Кристина; Каньяс-Арментерос, Дидак; Маллабиабаррена, Аррате; Камм, Кай; Сабидо, Эдвард; Грубер-Водика, Харальд; Шируотер, Бернд; Серрано, Луис; Себе-Педрос, Арнау (октябрь 2023 г.). «Поэтапное возникновение программы экспрессии нейрональных генов на ранней стадии эволюции животных» . Клетка . 186 (21): 4676–4693.e29. дои : 10.1016/j.cell.2023.08.027 . ПМЦ 10580291 . ПМИД 37729907 . Архивировано из оригинала 17 октября 2023 года.
- ^ Зайя LC, Тади П. Нейроанатомия, Моторный нейрон . [Обновлено 25 июля 2022 г.]. В: StatPearls [Интернет]. Остров сокровищ (Флорида): StatPearls Publishing; 2023 январь-.
- ↑ Перейти обратно: Перейти обратно: а б Зедалис Дж. и Эггебрехт Дж. (8 марта 2018 г.) Биология для курсов AP® 26.1 Нейроны и глиальные клетки. OpenStax https://openstax.org/books/biology-ap-courses/pages/26-1-neurons-and-glial-cells (по состоянию на 2023 г., 15 августа).
- ^ Мур, Кейт; Далли, Артур (2005). Клинически-ориентированная анатомия (5-е изд.). ДВВ. стр. 47 . ISBN 0-7817-3639-0 .
Пучок нервных волокон (аксонов), соединяющих соседние или отдаленные ядра ЦНС, представляет собой тракт.
- ^ «Какие части нервной системы?» . Октябрь 2018 года . Проверено 8 июля 2022 г.
- ^ Дэвис, Мелисса (9 апреля 2002 г.). «Нейрон: сравнение размеров» . Нейронаука: путешествие по мозгу . Проверено 20 июня 2009 г.
- ^ Чудлер Э.Х. «Факты и цифры о мозге» . Нейронаука для детей . Проверено 20 июня 2009 г.
- ^ «16.7: Нервная система» . Свободные тексты по биологии . 14 января 2021 г. Проверено 28 февраля 2022 г.
- ^ Херруп К., Ян Ю (май 2007 г.). «Регуляция клеточного цикла в постмитотическом нейроне: оксюморон или новая биология?». Обзоры природы. Нейронаука . 8 (5): 368–78. дои : 10.1038/nrn2124 . ПМИД 17453017 . S2CID 12908713 .
- ^ Хименес, К. (февраль 1998 г.). «[Состав и строение мембраны нейрона: молекулярные основы ее физиологии и патологии]» . Ревиста де Неврология . 26 (150): 232–239. ISSN 0210-0010 . ПМИД 9563093 .
- ^ Бюллетень государственных больниц . Государственная комиссия по безумию. 1897. с. 378.
- ^ «Медицинское определение нейротрубочек» . www.merriam-webster.com .
- ^ Зекка Л., Галлорини М., Шюнеманн В., Траутвейн А.Х., Герлах М., Ридерер П., Веццони П., Тампеллини Д. (март 2001 г.). «Содержание железа, нейромеланина и ферритина в черной субстанции нормальных людей в разном возрасте: последствия для хранения железа и нейродегенеративных процессов». Журнал нейрохимии . 76 (6): 1766–73. дои : 10.1046/j.1471-4159.2001.00186.x . ПМИД 11259494 . S2CID 31301135 .
- ^ Эрреро М.Т., Хирш Э.К., Кастнер А., Лукин М.Р., Джавой-Агид Ф., Гонсало Л.М., Обесо Дж.А., Агид Ю. (1993). «Накопление нейромеланина с возрастом в катехоламинергических нейронах ствола мозга Macaca fascicleis». Развивающая нейронаука . 15 (1): 37–48. дои : 10.1159/000111315 . ПМИД 7505739 .
- ^ Бранк UT, Терман А (сентябрь 2002 г.). «Липофусцин: механизмы возрастного накопления и влияние на функции клеток». Свободно-радикальная биология и медицина . 33 (5): 611–9. дои : 10.1016/s0891-5849(02)00959-0 . ПМИД 12208347 .
- ^ Чжао Б., Мека Д.П., Шарренберг Р., Кениг Т., Шванке Б., Коблер О., Виндхорст С., Кройц М.Р., Михайлова М., Кальдерон де Анда Ф (август 2017 г.). «Микротрубочки модулируют динамику F-актина во время поляризации нейронов» . Научные отчеты . 7 (1): 9583. Бибкод : 2017NatSR...7.9583Z . дои : 10.1038/s41598-017-09832-8 . ПМК 5575062 . ПМИД 28851982 .
- ^ Ли В.К., Хуан Х., Фэн Дж., Санес-младший, Браун Э.Н., Со П.Т., Недиви Е (февраль 2006 г.). «Динамическое ремоделирование дендритных ветвей в ГАМКергических интернейронах зрительной коры взрослых» . ПЛОС Биология . 4 (2): е29. дои : 10.1371/journal.pbio.0040029 . ПМЦ 1318477 . ПМИД 16366735 .
- ^ Эл, Мартини, Фредерик и др. (2005). Анатомия и физиология, издание 2007 г. , издание 2007 г. Книжный магазин «Рекс», Inc. 288. ИСБН 978-971-23-4807-5 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^
 В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 12.2 Нервная ткань. ISBN 978-1-947172-04-3 .
В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 12.2 Нервная ткань. ISBN 978-1-947172-04-3 . - ^ Гербер У (январь 2003 г.). «Метаботропные рецепторы глутамата в сетчатке позвоночных». Документа Офтальмологическая. Достижения офтальмологии . 106 (1): 83–7. дои : 10.1023/А:1022477203420 . ПМИД 12675489 . S2CID 22296630 .
- ^ Уилсон Н.Р., Рунян Калифорния, Ван Ф.Л., Сур М. (август 2012 г.). «Деление и вычитание с помощью отдельных корковых тормозных сетей in vivo» . Природа . 488 (7411): 343–8. Бибкод : 2012Natur.488..343W . дои : 10.1038/nature11347 . hdl : 1721.1/92709 . ПМЦ 3653570 . ПМИД 22878717 .
- ↑ Перейти обратно: Перейти обратно: а б Ллинас Р.Р. (01 января 2014 г.). «Внутренние электрические свойства нейронов млекопитающих и функции ЦНС: историческая перспектива» . Границы клеточной нейронауки . 8 : 320. дои : 10.3389/fncel.2014.00320 . ПМК 4219458 . ПМИД 25408634 .
- ^ Колодин Ю.О., Веселовская Н.Н., Веселовский Н.С., Федулова СА. Ионная проводимость связана с формированием повторяющихся импульсов в ганглиозных клетках сетчатки крысы . Конгресс Acta Physiologica. Архивировано из оригинала 7 октября 2012 г. Проверено 20 июня 2009 г.
- ^ «Ионная проводимость, лежащая в основе возбудимости тонических ганглиозных клеток сетчатки взрослой крысы» . Юколодин.50webs.com. 27 апреля 2008 г. Проверено 16 февраля 2013 г.
- ^ Скаммелл Т.Э., Джексон А.С., Фрэнкс Н.П., Висден В., Даувилье Ю. (январь 2019 г.). «Гистамин: нейронные цепи и новые лекарства» . Спать . 42 (1). doi : 10.1093/sleep/zsy183 . ПМЦ 6335869 . ПМИД 30239935 .
- ^ «Техника Patch-seq помогает отобразить изменения нервных клеток мозга» . Новости-medical.net . 3 декабря 2020 г. Проверено 26 августа 2021 г.
- ^ Наука АААС. «ПЕРЕПИСЬ КЛЕТОК МОЗГА» . Проверено 17 октября 2023 г.
- ^ Макферсон, Гордон (2002). Медицинский словарь Блэка (40-е изд.). Лэнхэм, Мэриленд: Scarecrow Press. стр. 431–434. ISBN 0810849844 .
- ^ Иванников М.В., Маклауд Г.Т. (июнь 2013 г.). «Уровни свободного Ca²⁺ в митохондриях и их влияние на энергетический обмен в окончаниях двигательных нервов дрозофилы» . Биофизический журнал . 104 (11): 2353–61. Бибкод : 2013BpJ...104.2353I . дои : 10.1016/j.bpj.2013.03.064 . ПМЦ 3672877 . ПМИД 23746507 .
- ^ Эркулано-Хаузель С (ноябрь 2009 г.). «Человеческий мозг в цифрах: мозг примата в линейном масштабе» . Границы человеческой неврологии . 3:31 . doi : 10.3389/neuro.09.031.2009 . ПМК 2776484 . ПМИД 19915731 .
- ^ «Почему человеческий мозг так сложно понять? Мы спросили четырех нейробиологов» . Институт Аллена . Проверено 17 октября 2023 г.
- ^ Драхман Д.А. (июнь 2005 г.). «Есть ли у нас лишние мозги?». Неврология . 64 (12): 2004–5. дои : 10.1212/01.WNL.0000166914.38327.BB . ПМИД 15985565 . S2CID 38482114 .
- ^ Укар, Хасан; Ватанабэ, Сатоши; Ногучи, Джун; Моримото, Юичи; Иино, Юсуке; Ягишита, Шо; Такахаси, Норико; Касаи, Харуо (декабрь 2021 г.). «Механическое воздействие расширения дендритных шипиков на пресинаптический экзоцитоз». Природа 600 (7890): 686–689. Бибкод : 2021Природа.600..686U . дои : 10.1038/ s41586-021-04125-7 ISSN 1476-4687 . ПМИД 34819666 . S2CID 244648506 .
Краткое содержание:
«Сильные синапсы выявляют механические взаимодействия в мозгу» . Природа . 24 ноября 2021 г. doi : 10.1038/d41586-021-03516-0 . Проверено 21 февраля 2022 г. - ^ «Исследователи открывают новый тип клеточной связи в мозге» . Исследовательский институт Скриппса . Проверено 12 февраля 2022 г.
- ^ Скиаппарелли, Лусио М.; Шарма, Пранав; Он, Хай-Янь; Ли, Цзяньли; Шах, Сахил Х.; МакКлатчи, Дэниел Б.; Ма, Юаньхуэй; Лю, Хань-Сюань; Голдберг, Джеффри Л.; Йейтс, Джон Р.; Клайн, Холлис Т. (25 января 2022 г.). «Протеомный скрининг выявляет разнообразный транспорт белков между соединенными нейронами зрительной системы» . Отчеты по ячейкам . 38 (4): 110287. doi : 10.1016/j.celrep.2021.110287 . ISSN 2211-1247 . ПМЦ 8906846 . ПМИД 35081342 .
- ^ Левитан, Ирвин Б.; Качмарек, Леонард К. (2015). «Электрическая сигнализация в нейронах». Нейрон . Издательство Оксфордского университета. стр. 41–62. дои : 10.1093/med/9780199773893.003.0003 . ISBN 978-0-19-977389-3 .
- ^ О'Лири, Оливия Ф.; Огбонная, Эбере С.; Феличе, Даниэла; Левоне, Брунно Р.; К. Конрой, Лотарингия; Фицджеральд, Патрик; Браво, Хавьер А.; Форсайт, Пол; Биненшток, Джон; Динан, Тимоти Г.; Крайан, Джон Ф. (1 февраля 2018 г.). «Блуждающий нерв модулирует экспрессию BDNF и нейрогенез в гиппокампе» . Европейская нейропсихофармакология . 28 (2): 307–316. дои : 10.1016/j.euroneuro.2017.12.004 . ISSN 0924-977X . ПМИД 29426666 . S2CID 46819013 .
- ^ Череп С., Посфаи Б., Ленарт Н., Фекете Р., Ласло З.И., Леле З. (январь 2020 г.). «Микроглия контролирует и защищает функции нейронов посредством специализированных соматических пуринергических соединений» . Наука . 367 (6477): 528–537. Бибкод : 2020Sci...367..528C . doi : 10.1126/science.aax6752 . ПМИД 31831638 . S2CID 209343260 .
- ^ Чудлер Э.Х. «Вехи нейробиологических исследований» . Нейронаука для детей . Проверено 20 июня 2009 г.
- ^ Патлак Дж., Гиббонс Р. (1 ноября 2000 г.). «Электрическая активность нервов» . Потенциалы действия в нервных клетках . Архивировано из оригинала 27 августа 2009 года . Проверено 20 июня 2009 г.
- ^ Харрис-Уоррик, РМ (октябрь 2011 г.). «Нейромодуляция и гибкость в сетях центрального генератора образов» . Современное мнение в нейробиологии . 21 (5): 685–92. дои : 10.1016/j.conb.2011.05.011 . ПМК 3171584 . ПМИД 21646013 .
- ^ Браун Э.Н., Касс Р.Э., Митра П.П. (май 2004 г.). «Анализ данных множественных нейронных импульсов: современное состояние и проблемы будущего». Природная неврология . 7 (5): 456–61. дои : 10.1038/nn1228 . ПМИД 15114358 . S2CID 562815 .
- ^ Торп С.Дж. (1990). «Время прибытия пиков: высокоэффективная схема кодирования для нейронных сетей» (PDF) . Экмиллер Р., Хартманн Г., Хауске Г. (ред.). Параллельная обработка в нейронных системах и компьютерах . Северная Голландия. стр. 91–94. ISBN 9780444883902 . Архивировано из оригинала (PDF) 15 февраля 2012 г.
- ↑ Перейти обратно: Перейти обратно: а б Калат, Джеймс В. (2016). Биологическая психология (12-е изд.). Австралия: Cengage Learning. ISBN 9781305105409 . OCLC 898154491 .
- ^ Эккерт Р., Рэндалл Д. (1983). Физиология животных: механизмы и приспособления . Сан-Франциско: WH Freeman. п. 239 . ISBN 978-0-7167-1423-1 .
- ↑ Перейти обратно: Перейти обратно: а б Фингер, Стэнли (1994). Истоки нейробиологии: история исследований функций мозга . Издательство Оксфордского университета. п. 47. ИСБН 9780195146943 . ОСЛК 27151391 .
Первая статья Рамона-и-Кахаля о пятне Гольджи была посвящена мозжечку птиц и появилась в журнале Revista в 1888 году. Он признал, что обнаружил, что нервные волокна очень сложны, но заявил, что не может найти никаких доказательств существования ни аксонов, ни дендритов. накладывают анастомоз и формируют сети. Каждый нервный элемент он называл «абсолютно автономным кантоном».
- ↑ Перейти обратно: Перейти обратно: а б Оксфордский словарь английского языка , 3-е издание, 2003 г., св.
- ^ Мехта А.Р., Мехта П.Р., Андерсон С.П., Маккиннон Б.Л., Компстон А. (январь 2020 г.). «Этимология серого вещества и нейрон (е)» . Мозг . 143 (1): 374–379. дои : 10.1093/brain/awz367 . ПМЦ 6935745 . ПМИД 31844876 .
- ^ «Программа просмотра Ngram Google Книг» . book.google.com . Проверено 19 декабря 2020 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Лопес-Муньос Ф., Бойя Дж., Аламо К. (октябрь 2006 г.). «Теория нейронов, краеугольный камень нейробиологии, к столетию со дня вручения Нобелевской премии Сантьяго Рамону-и-Кахалю». Бюллетень исследований мозга . 70 (4–6): 391–405. дои : 10.1016/j.brainresbull.2006.07.010 . ПМИД 17027775 . S2CID 11273256 .
- ^ Фингер, Стэнли (1994). Истоки нейробиологии: история исследований функций мозга . Издательство Оксфордского университета. п. 47. ИСБН 9780195146943 . ОСЛК 27151391 .
...человек, который два года спустя напишет очень влиятельный обзор доказательств в пользу доктрины нейронов. В своей статье Вальдейер (1891)... писал, что нервные клетки свободно оканчиваются разветвлениями на концах и что «нейрон» является анатомической и физиологической единицей нервной системы. Слово «нейрон» родилось именно так.
- ^ «Ктонамедит — словарь медицинских эпонимов» . www.whonamedit.com .
Сегодня Вильгельма фон Вальдейера-Гарца помнят как основателя теории нейронов, придумавшего термин «нейрон» для описания клеточной функциональной единицы нервной системы, а также сформулировавшего и разъяснившего эту концепцию в 1891 году.
- ^ Грант G (октябрь 2007 г.). «Как Нобелевская премия по физиологии и медицине 1906 года была разделена между Гольджи и Кахалем». Обзоры исследований мозга . 55 (2): 490–8. дои : 10.1016/j.brainresrev.2006.11.004 . ПМИД 17306375 . S2CID 24331507 .
- ^ Ведьмак М.Р., Киров С.А., Харрис К.М. (январь 2007 г.). «Пластичность перисинаптической астроглии во время синаптогенеза в гиппокампе зрелой крысы». Глия . 55 (1): 13–23. CiteSeerX 10.1.1.598.7002 . дои : 10.1002/glia.20415 . ПМИД 17001633 . S2CID 10664003 .
- ^ Коннорс Б.В., Лонг, Массачусетс (2004). «Электрические синапсы в мозге млекопитающих» . Ежегодный обзор неврологии . 27 (1): 393–418. дои : 10.1146/annurev.neuro.26.041002.131128 . ПМИД 15217338 .
- ^ Гиллери Р.В. (июнь 2005 г.). «Наблюдения за синаптическими структурами: истоки учения о нейронах и его современное состояние» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 360 (1458): 1281–307. дои : 10.1098/rstb.2003.1459 . ПМК 1569502 . ПМИД 16147523 .
- ^ Саббатини Р.М. (апрель – июль 2003 г.). «Нейроны и синапсы: история открытия» . Журнал «Мозг и разум» : 17.
- ^ Джурисич М., Антич С., Чен В.Р., Зечевич Д. (июль 2004 г.). «Визуализация напряжения от дендритов митральных клеток: затухание ВПСП и триггерные зоны спайков» . Журнал неврологии . 24 (30): 6703–14. doi : 10.1523/JNEUROSCI.0307-04.2004 . HDL : 1912/2958 . ПМК 6729725 . ПМИД 15282273 .
- ^ Кочилла А.Дж., Алфорд С. (март 1997 г.). «Синапсическое возбуждение, опосредованное рецептором глутамата, в аксонах миноги» . Журнал физиологии . 499 (Часть 2): 443–57. дои : 10.1113/jphysicalol.1997.sp021940 . ПМК 1159318 . ПМИД 9080373 .
- ^ Гувенс Н.В., Уилсон Р.И. (2009). «Распространение сигнала в центральных нейронах дрозофилы» . Журнал неврологии . 29 (19): 6239–6249. doi : 10.1523/jneurosci.0764-09.2009 . ПМК 2709801 . ПМИД 19439602 .
- ^ Уильямс Р.В., Херруп К. (1988). «Контроль количества нейронов». Ежегодный обзор неврологии . 11 (1): 423–53. дои : 10.1146/annurev.ne.11.030188.002231 . ПМИД 3284447 .
- ^ фон Бартельд К.С., Бахни Дж., Эркулано-Хаузель С. (декабрь 2016 г.). «Поиск истинного количества нейронов и глиальных клеток в человеческом мозге: обзор 150-летнего подсчета клеток» . Журнал сравнительной неврологии . 524 (18): 3865–3895. дои : 10.1002/cne.24040 . ПМК 5063692 . ПМИД 27187682 .
- ^ Краевски К.М., Льюис Р.А., Фюрст Д.Р., Туранский С., Хиндерер С.Р., Гарберн Дж., Камхольц Дж., Шай М.Е. (июль 2000 г.). «Неврологическая дисфункция и аксональная дегенерация при болезни Шарко-Мари-Тута типа 1А». Мозг . 123 (7): 1516–27. дои : 10.1093/мозг/123.7.1516 . ПМИД 10869062 .
- ^ «О болезни Альцгеймера: Симптомы» . Национальный институт старения. Архивировано из оригинала 15 января 2012 года . Проверено 28 декабря 2011 г.
- ^ Бернс А., Илифф С. (февраль 2009 г.). «Болезнь Альцгеймера». БМЖ . 338 : б158. дои : 10.1136/bmj.b158 . ПМИД 19196745 . S2CID 8570146 .
- ^ Querfurth HW, LaFerla FM (январь 2010 г.). «Болезнь Альцгеймера». Медицинский журнал Новой Англии . 362 (4): 329–44. дои : 10.1056/NEJMra0909142 . ПМИД 20107219 . S2CID 205115756 .
- ^ «Информационная страница о болезни Паркинсона» . НИНДС . 30 июня 2016 года. Архивировано из оригинала 4 января 2017 года . Проверено 18 июля 2016 г.
- ^ «Двигательные расстройства» . Международное общество нейромодуляции .
- ^ Кершенштейнер М., Шваб М.Е., Лихтман Дж.В., Мисгельд Т. (май 2005 г.). «Визуализация аксональной дегенерации и регенерации в поврежденном спинном мозге in vivo». Природная медицина . 11 (5): 572–7. дои : 10.1038/nm1229 . ПМИД 15821747 . S2CID 25287010 .
- ^ Кемперманн Дж., Гейдж Ф.Х., Айгнер Л., Сонг Х., Кертис М.А., Тюрет С., Кун Х.Г., Джессбергер С., Франкланд П.В., Кэмерон Х.А., Гулд Э., Хен Р., Абрус Д.Н., Тони Н., Шиндер А.Ф., Чжао X, Лукассен П.Дж. , Фрисен Дж. (июль 2018 г.). «Нейрогенез взрослого человека: данные и оставшиеся вопросы» . Клеточная стволовая клетка . 23 (1): 25–30. дои : 10.1016/j.stem.2018.04.004 . ПМК 6035081 . ПМИД 29681514 .
- ^ Каллауэй, Юэн (26 мая 2011 г.). «Как сделать человеческий нейрон». Природа . дои : 10.1038/news.2011.328 .
Преобразуя клетки человеческой кожи в работающие нервные клетки, исследователи, возможно, разработали модель заболеваний нервной системы и, возможно, даже регенеративную терапию, основанную на трансплантации клеток. Достижение, о котором сегодня сообщается в журнале Nature , является последним в быстро развивающейся области, называемой трансдифференциацией, в которой клетки вынуждены принимать новые идентичности. В прошлом году исследователи превратили клетки соединительной ткани, обнаруженные в коже, в клетки сердца, клетки крови и клетки печени.
- ↑ Перейти обратно: Перейти обратно: а б Ван З, Тан Б, Хэ Ю, Джин П (март 2016 г.). «Динамика метилирования ДНК в нейрогенезе» . Эпигеномика . 8 (3): 401–14. дои : 10.2217/эпи.15.119 . ПМЦ 4864063 . ПМИД 26950681 .
- ^ Ории К.Е., Ли Ю., Кондо Н., Маккиннон П.Дж. (июнь 2006 г.). «Селективное использование негомологичных путей соединения концов и гомологичной рекомбинации ДНК во время развития нервной системы» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (26): 10017–22. Бибкод : 2006PNAS..10310017O . дои : 10.1073/pnas.0602436103 . ПМК 1502498 . ПМИД 16777961 .
- ^ Цереп, Чаба; Шварц, Анетт Д.; Посфаи, Балаж; Ласло, Жофия И.; Келлермайер, Анна; Кёрней, Жужанна; Кисфали, Мате; Ньергес, Миклош; Леле, Жолт; Катона, Иштван (сентябрь 2022 г.). «Микроглиальный контроль развития нейронов через соматические пуринергические соединения» . Отчеты по ячейкам . 40 (12): 111369. doi : 10.1016/j.celrep.2022.111369 . ПМЦ 9513806 . ПМИД 36130488 . S2CID 252416407 .
- ^ Ю Г, Хэ Зи (август 2006 г.). «Глиальное ингибирование регенерации аксонов ЦНС» . Обзоры природы. Нейронаука . 7 (8): 617–27. дои : 10.1038/nrn1956 . ПМЦ 2693386 . ПМИД 16858390 .
Дальнейшее чтение [ править ]
- Баллок Т.Х., Беннетт М.В., Джонстон Д., Джозефсон Р., Мардер Э., Филдс Р.Д. (ноябрь 2005 г.). «Нейронаука. Нейронная доктрина, редукс». Наука . 310 (5749): 791–3. дои : 10.1126/science.1114394 . ПМИД 16272104 . S2CID 170670241 .
- Кандель Э.Р., Шварц Дж.Х., Джесселл Т.М. (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 0-8385-7701-6 .
- Питерс А., Палай С.Л., Вебстер Х.С. (1991). Тонкая структура нервной системы (3-е изд.). Нью-Йорк: Издательство Оксфордского университета. ISBN 0-19-506571-9 .
- Рамон-и-Кахаль С. (1933). Гистология (10-е изд.). Балтимор: Вуд.
- Робертс А., Буш Б.М. (1981). Нейроны без импульсов . Кембридж: Издательство Кембриджского университета. ISBN 0-521-29935-7 .
- Снелл РС (2010). Клиническая нейроанатомия . Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-9427-5 .
Внешние ссылки [ править ]
 Определения из Викисловаря
Определения из Викисловаря  СМИ из Commons
СМИ из Commons  Цитаты из Wikiquote
Цитаты из Wikiquote  Учебники из Wikibooks
Учебники из Wikibooks  Данные из Викиданных
Данные из Викиданных
- Нейробиология в Керли
- IBRO (Международная организация по исследованию мозга) . Содействие исследованиям в области нейробиологии, особенно в менее хорошо финансируемых странах.
- NeuronBank — онлайн-инструмент нейромики для каталогизации типов нейронов и синаптических связей.
- Нейроанатомические изображения высокого разрешения мозга приматов и не-приматов .
- Кафедра неврологии Викиверситета , которая в настоящее время предлагает два курса: «Основы неврологии» и «Сравнительная неврология» .
- Поиск NIF - Neuron. Архивировано 22 января 2015 г. на Wayback Machine через Neuroscience Information Framework.
- База данных, ориентированная на клетки – нейрон
- Полный список типов нейронов согласно конвенции Петиллы на сайте NeuroLex .
- NeuroMorpho.Org - онлайн-база данных цифровых реконструкций морфологии нейронов.
- Галерея изображений иммуногистохимии: Нейрон
- Академия Хана: Анатомия нейрона
- Изображения нейронов