нейротрансмиссия
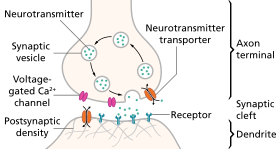
Нейротрансмиссия (лат. transmissio «прохождение, пересечение» от « трансмиттере » «отправлять, пропускать») — это процесс, посредством которого сигнальные молекулы, называемые нейротрансмиттерами, ( пресинаптическим нейроном высвобождаются окончанием аксона нейрона ), связываются с ним и реагируют с ним. рецепторы на дендритах другого нейрона (постсинаптического нейрона), находящегося на небольшом расстоянии. Аналогичный процесс происходит при ретроградной нейротрансмиссии , когда дендриты постсинаптического нейрона высвобождают ретроградные нейротрансмиттеры (например, эндоканнабиноиды ; синтезируются в ответ на повышение внутриклеточных уровней кальция ), которые передают сигналы через рецепторы, расположенные на окончаниях аксонов пресинаптического нейрона. главным образом в ГАМКергических и глутаматергических синапсах . [1] [2] [3] [4]
Нейротрансмиссия регулируется несколькими различными факторами: доступностью и скоростью синтеза нейромедиатора, высвобождением этого нейромедиатора, базовой активностью постсинаптической клетки, количеством доступных постсинаптических рецепторов, с которыми нейромедиатор может связываться, и последующими удаление или дезактивация нейромедиатора ферментами или пресинаптическим обратным захватом. [5] [6]
В ответ на пороговый потенциал действия или градуированный электрический потенциал высвобождается нейромедиатор в пресинаптическом терминале . Высвобожденный нейротрансмиттер может затем перемещаться через синапс, чтобы быть обнаруженным и связываться с рецепторами постсинаптического нейрона. Связывание нейротрансмиттеров может влиять на постсинаптический нейрон как тормозным , так и возбуждающим образом. Связывание нейротрансмиттеров с рецепторами постсинаптического нейрона может вызвать либо кратковременные изменения, такие как изменения мембранного потенциала , называемые постсинаптическими потенциалами , либо долгосрочные изменения за счет активации сигнальных каскадов .
Нейроны образуют сложные биологические нейронные сети, по которым проходят нервные импульсы (потенциалы действия). Нейроны не соприкасаются друг с другом (за исключением случая электрического синапса через щелевой переход ); вместо этого нейроны взаимодействуют в точках тесного контакта, называемых синапсами. Нейрон передает информацию посредством потенциала действия. Когда нервный импульс достигает синапса, он может вызвать высвобождение нейромедиаторов, которые влияют на другой (постсинаптический) нейрон. Постсинаптический нейрон может получать входные сигналы от многих дополнительных нейронов, как возбуждающих, так и тормозных. Возбуждающее и тормозящее влияния суммируются, и если суммарный эффект является тормозящим, то нейрон с меньшей вероятностью «сработает» (т. е. создаст потенциал действия), а если суммарный эффект является возбуждающим, нейрон с большей вероятностью сработает. огонь. Насколько вероятно, что нейрон сработает, зависит от того, насколько далеко его мембранный потенциал находится от порогового потенциала , напряжения, при котором срабатывает потенциал действия, поскольку он достаточно зависит от напряжения. натриевые каналы активируются так, что чистый входящий натриевый ток превышает все внешние токи. [7] Возбуждающие входы приближают нейрон к порогу, тогда как тормозящие входы приближают нейрон к порогу. Потенциал действия — это событие «все или ничего»; нейроны, мембраны которых не достигли порога, не сработают, а те, которые достигли порога, должны сработать. Как только потенциал действия инициируется (традиционно в бугорке аксона ), он распространяется вдоль аксона, что приводит к высвобождению нейротрансмиттеров в синаптическом бутоне для передачи информации еще одному соседнему нейрону.
Этапы нейротрансмиссии в синапсе
[ редактировать ]- Синтез нейромедиатора. Это может происходить в теле клетки , в аксоне или в конце аксона .
- Хранение нейромедиатора в накопительных гранулах или везикулах в окончаниях аксонов.
- Кальций поступает в терминаль аксона во время потенциала действия, вызывая высвобождение нейромедиатора в синаптическую щель.
- После высвобождения медиатор связывается и активирует рецептор в постсинаптической мембране.
- Деактивация нейромедиатора. Нейромедиатор либо разрушается ферментативно, либо возвращается в терминал, из которого он пришел, где его можно использовать повторно, либо разлагается и удаляется. [8]
Общее описание
[ редактировать ]Нейромедиаторы спонтанно упаковываются в везикулы и высвобождаются отдельными квантовыми пакетами независимо от пресинаптических потенциалов действия. Это медленное высвобождение заметно и оказывает микротормозное или микровозбуждающее воздействие на постсинаптический нейрон. Потенциал действия кратковременно усиливает этот процесс. Нейротрансмиттеры, содержащие везикулы, группируются вокруг активных центров и после высвобождения могут быть переработаны с помощью одного из трех предложенных механизмов. Первый предложенный механизм включает частичное открытие, а затем повторное закрытие пузырька. Вторые два предусматривают полное слияние пузырька с мембраной с последующей рециркуляцией или рециркуляцией в эндосому. Везикулярное слияние во многом обусловлено концентрацией кальция в микродоменах, расположенных вблизи кальциевых каналов, что обеспечивает высвобождение нейротрансмиттера всего лишь в течение микросекунд, тогда как возвращение к нормальной концентрации кальция занимает пару сотен микросекунд. Считается, что экзоцитоз везикул осуществляется белковым комплексом, называемым SNARE – мишень для ботулинических токсинов . После высвобождения нейромедиатор попадает в синапс и встречает рецепторы. Рецепторы нейромедиаторов могут быть либо ионотропными, либо связанными с g-белком. Ионотропные рецепторы пропускают ионы при воздействии лиганда. Основная модель включает в себя рецептор, состоящий из нескольких субъединиц, которые позволяют координировать предпочтение ионов. Рецепторы, связанные с G-белком, также называемые метаботропными рецепторами, при связывании с лигандом претерпевают конформационные изменения, приводящие к внутриклеточному ответу. Прекращение активности нейромедиатора обычно осуществляется переносчиком, однако возможна и ферментативная дезактивация. [9]
Суммирование
[ редактировать ]Каждый нейрон соединяется с множеством других нейронов, получая от них множество импульсов. Суммирование – это сложение этих импульсов на аксонном бугорке. Если нейрон получает только возбуждающие импульсы, он генерирует потенциал действия. Если вместо этого нейрон получит столько же тормозных импульсов, сколько и возбуждающих, торможение нейтрализует возбуждение, и нервный импульс на этом остановится. [10] Генерация потенциала действия пропорциональна вероятности и характеру высвобождения нейротрансмиттера, а также сенсибилизации постсинаптических рецепторов. [11] [12] [13]
Пространственное суммирование означает, что эффекты импульсов, полученных в разных местах нейрона, суммируются, так что нейрон может сработать, когда такие импульсы получены одновременно, даже если каждого импульса по отдельности будет недостаточно, чтобы вызвать срабатывание.
Временное суммирование означает, что эффекты импульсов, полученных в одном и том же месте, могут суммироваться, если импульсы получены в тесной временной последовательности. Таким образом, нейрон может сработать при получении нескольких импульсов, даже если каждого импульса по отдельности будет недостаточно, чтобы вызвать срабатывание. [14]
Конвергенция и дивергенция
[ редактировать ]Нейротрансмиссия предполагает как конвергенцию, так и расхождение информации. Сначала на один нейрон влияют многие другие, что приводит к конвергенции входных данных. Когда нейрон срабатывает, сигнал отправляется многим другим нейронам, что приводит к расхождению выходных сигналов. Многие другие нейроны находятся под влиянием этого нейрона. [ нужна ссылка ]
Котрансмиссия
[ редактировать ]Котрансмиссия — это высвобождение нескольких типов нейромедиаторов из одного нервного окончания .
В нервном окончании нейротрансмиттеры присутствуют в мембранных везикулах размером 35–50 нм, называемых синаптическими везикулами . Чтобы высвободить нейротрансмиттеры, синаптические пузырьки временно стыкуются и сливаются у основания специализированных липопротеиновых структур чашеобразной формы размером 10–15 нм на пресинаптической мембране, называемых поросомами . [15] нейрональных поросом Протеом был решен, обеспечивая молекулярную архитектуру и полный состав механизмов. [16]
Недавние исследования множества систем показали, что большинство, если не все, нейроны выделяют несколько различных химических посланников. [17] Котрансмиссия позволяет оказывать более сложные эффекты на постсинаптические рецепторы и, таким образом, обеспечивает более сложную связь между нейронами.
В современной нейробиологии нейроны часто классифицируются по их котрансмиттеру. Например, стриарные «ГАМКергические нейроны» используют опиоидные пептиды или вещество Р в качестве основного котрансмиттера.
Некоторые нейроны могут высвобождать по крайней мере два нейротрансмиттера одновременно, причем другой является котрансмиттером, чтобы обеспечить стабилизирующую отрицательную обратную связь, необходимую для значимого кодирования, в отсутствие тормозных интернейронов . [18] Примеры включают в себя:
- ГАМК – глицина . совместное высвобождение
- Дофамин – глутамата . совместное высвобождение
- ацетилхолина (АХ) и глутамата. Совместное высвобождение
- ACh – вазоактивного кишечного пептида (VIP). совместное высвобождение
- ACh – пептида, связанного с геном кальцитонина (CGRP). совместный высвобождение
- глутамата и динорфина Совместное высвобождение (в гиппокампе ).
Норадреналин и АТФ являются симпатическими комедиаторами. Обнаружено, что эндоканнабиноид анадамид и каннабиноид WIN 55,212-2 могут изменять общий ответ на стимуляцию симпатических нервов и указывают на то, что пресинаптические рецепторы CB1 опосредуют симпато -ингибирующее действие. Таким образом, каннабиноиды могут ингибировать как норадренергический, так и пуринергический компоненты симпатической нейротрансмиссии. [19]
Одной из необычных пар котрансмиттеров являются ГАМК и глутамат, которые высвобождаются из одних и тех же окончаний аксонов нейронов, происходящих из вентральной покрышки (ВТА), внутреннего бледного шара и супрамаммилярного ядра . [20] Первые два выступают на хабенулу , тогда как известно, что проекции надсосцевидного ядра нацелены на зубчатую извилину гиппокампа. [20]
Генетическая ассоциация
[ редактировать ]Нейротрансмиссия генетически связана с другими характеристиками или особенностями. Например, анализ обогащения различных сигнальных путей привел к открытию генетической связи с внутричерепным объемом. [21]
См. также
[ редактировать ]- Ауторецептор
- Модель биологического нейрона § Синаптическая передача (Кох и Сегев)
- Электрофизиология
- Рецептор, связанный с G-белком
- Молекулярная нейрофармакология
- Нервно-мышечная передача
- Нейропсихофармакология
Ссылки
[ редактировать ]- ^ Мелис М., Пистис М. (декабрь 2007 г.). «Передача сигналов эндоканнабиноидов в дофаминовых нейронах среднего мозга: больше, чем физиология?» . Современная нейрофармакология . 5 (4): 268–77. дои : 10.2174/157015907782793612 . ПМЦ 2644494 . ПМИД 19305743 .
Таким образом, вполне возможно, что низкие уровни рецепторов CB1 расположены на глутаматергических и ГАМКергических окончаниях, воздействующих на DA-нейроны [127, 214], где они могут точно регулировать высвобождение тормозных и возбуждающих нейротрансмиттеров и регулировать активацию DA-нейронов.
Таким образом, электрофизиологические эксперименты in vitro, проведенные независимыми лабораториями, предоставили доказательства локализации рецептора CB1 на глутаматергических и ГАМКергических окончаниях аксонов в VTA и SNc. - ^ Флорес А., Мальдонадо Р., Беррендеро Ф. (декабрь 2013 г.). «Перекрестные помехи каннабиноидов и гипокретина в центральной нервной системе: что мы знаем на данный момент» . Границы в неврологии . 7 : 256. дои : 10.3389/fnins.2013.00256 . ПМЦ 3868890 . ПМИД 24391536 .
Прямое взаимодействие CB1-HcrtR1 было впервые предложено в 2003 г. (Hilairet et al., 2003). Действительно, 100-кратное увеличение способности гипокретина-1 активировать передачу сигналов ERK наблюдалось при совместной экспрессии CB1 и HcrtR1... В этом исследовании более высокая эффективность гипокретина-1 по регуляции гетеромера CB1-HcrtR1 по сравнению с гомомером HcrtR1-HcrtR1 сообщалось (Ward et al., 2011b). Эти данные позволяют однозначно идентифицировать гетеромеризацию CB1-HcrtR1, имеющую существенное функциональное значение. ... Существование перекрестных помех между гипокретинергической и эндоканнабиноидной системами убедительно подтверждается их частично перекрывающимся анатомическим распределением и общей ролью в нескольких физиологических и патологических процессах. Однако мало что известно о механизмах, лежащих в основе этого взаимодействия. ... Действуя как ретроградный мессенджер, эндоканнабиноиды модулируют глутаматергические возбуждающие и ГАМКергические тормозные синаптические входы в дофаминергические нейроны VTA и передачу глутамата в NAc. Таким образом, активация рецепторов CB1, присутствующих на окончаниях аксонов ГАМКергических нейронов в ВТА, ингибирует передачу ГАМК, устраняя это ингибирующее воздействие на дофаминергические нейроны (Riegel and Lupica, 2004). Глутаматная синаптическая передача в VTA и NAc, преимущественно от нейронов PFC, аналогичным образом модулируется активацией рецепторов CB1 (Melis et al., 2004).
• Рисунок 1: Схема экспрессии CB1 головного мозга и орексинергических нейронов, экспрессирующих OX1 (HcrtR1) или OX2 (HcrtR2).
• Рисунок 2: Механизмы синаптической передачи сигналов в системах каннабиноидов и орексина.
• Рисунок 3: Схема путей мозга, участвующих в приеме пищи. - ^ Фрейнд Т.Ф., Катона И., Пиомелли Д. (июль 2003 г.). «Роль эндогенных каннабиноидов в синаптической передаче сигналов» . Физиологические обзоры . 83 (3): 1017–66. doi : 10.1152/physrev.00004.2003 . ПМИД 12843414 .
- ^ Аяканну, Тангесверан; Тейлор, Энтони Х.; Марсило, Тимоти Х.; Уиллетс, Джонатон М.; Конье, Джастин С. (2013). «Эндоканнабиноидная система и рак, зависимый от половых стероидов» . Международный журнал эндокринологии . 2013 : 259676. doi : 10.1155/2013/259676 . ISSN 1687-8337 . ПМЦ 3863507 . ПМИД 24369462 .
- ^ Нагацу, Т. (декабрь 2000 г.). «[Молекулярные механизмы нейротрансмиссии]». Ринсё Синкейгаку = Клиническая неврология . 40 (12): 1185–1188. ISSN 0009-918X . ПМИД 11464453 .
- ^ Андреа, Лаура С.; Бурроне, Хуан (март 2018 г.). «Роль спонтанной нейротрансмиссии в развитии синапсов и цепей» . Журнал нейробиологических исследований . 96 (3): 354–359. дои : 10.1002/jnr.24154 . ISSN 0360-4012 . ПМЦ 5813191 . ПМИД 29034487 .
- ^ Холден А., Уинлоу В. (1984). Нейробиология боли: симпозиум Северной группы нейробиологов, состоявшийся в Лидсе 18 апреля 1983 г. (1-е изд.). Манчестерский университет Пр. п. 111. ИСБН 978-0-7190-1061-3 .
- ^ Колб Б., Уишоу IQ (2003). Основы нейропсихологии человека (5-е изд.). Ценность. стр. 102–104. ISBN 978-0-7167-5300-1 . (ссылка на все пять этапов)
- ^ Сквайр Л., Берг Д., Блум Ф.Е., Дю Лак С., Гош А., Спитцер Северная Каролина (2013). Фундаментальная нейронаука (4-е изд.). Амстердам: Elsevier/Academic Press. стр. 133–181. ISBN 978-0-12-385870-2 .
- ^ Уильямс С.М., Макнамара Дж.О., Ламантия А.С., Кац Л.К., Фитцпатрик Д., Августин Г.Дж., Первес Д. (2001). «Суммирование синаптических потенциалов» . В Purves D, Augustine GJ, Fitzpatrick D, et al. (ред.). Нейронаука (2-е изд.). Сандерленд, Массачусетс: Sinauer Associates.
- ^ Ван Дж. Х., Вэй Дж., Чен X, Ю Дж., Чен Н., Ши Дж. (сентябрь 2008 г.). «Усиление и точность паттернов передачи в корковых возбуждающих унитарных синапсах улучшают кодирование спайков». Журнал клеточной науки . 121 (Часть 17): 2951–60. дои : 10.1242/jcs.025684 . ПМИД 18697836 . S2CID 31747181 .
- ^ Ю Дж, Цянь Х, Чен Н, Ван Дж Х (2011). «Квантовое высвобождение глутамата необходимо для надежного кодирования нейронов в церебральных сетях» . ПЛОС ОДИН . 6 (9): e25219. Бибкод : 2011PLoSO...625219Y . дои : 10.1371/journal.pone.0025219 . ПМК 3176814 . ПМИД 21949885 .
- ^ Ю Дж, Цянь Х, Ван Дж. Х. (август 2012 г.). «Повышение вероятности высвобождения передатчика улучшает преобразование синаптических аналоговых сигналов в цифровые спайки нейронов» . Молекулярный мозг . 5 (26): 26. дои : 10.1186/1756-6606-5-26 . ПМЦ 3497613 . ПМИД 22852823 .
- ^ Хеверн Фольксваген. «PSY 340 Мозг и поведение» . Архивировано из оригинала 19 февраля 2006 года.
- ^ Андерсон Л.Л. (2006). «Открытие поросомы: универсального секреторного механизма в клетках» . Журнал клеточной и молекулярной медицины . 10 (1): 126–31. дои : 10.1111/j.1582-4934.2006.tb00294.x . ПМЦ 3933105 . ПМИД 16563225 .
- ^ Ли Дж.С., Джеремик А., Шин Л., Чо В.Дж., Чен Икс, Йена Б.П. (июль 2012 г.). «Нейрональный поросомный протеом: молекулярная динамика и архитектура» . Журнал протеомики . 75 (13): 3952–62. дои : 10.1016/j.jprot.2012.05.017 . ПМК 4580231 . ПМИД 22659300 .
- ^ Трюдо Л.Е., Гутьеррес Р. (июнь 2007 г.). «О котрансмиссии и пластичности фенотипа нейромедиатора». Молекулярные вмешательства . 7 (3): 138–46. дои : 10.1124/ми.7.3.5 . ПМИД 17609520 .
- ^ Томас Э.А., Борнштейн Дж.К. (2003). «Тормозящая котрансмиссия или постгиперполяризующие потенциалы могут регулировать возбуждение в рекуррентных сетях с возбуждающей метаботропной передачей». Нейронаука . 120 (2): 333–51. дои : 10.1016/S0306-4522(03)00039-3 . ПМИД 12890506 . S2CID 26851745 .
- ^ Пакдичоте П., Данн В.Р., Ралевич В. (ноябрь 2007 г.). «Каннабиноиды ингибируют норадренергическую и пуринергическую симпатическую котрансмиссию в изолированном брыжеечном артериальном русле крысы» . Британский журнал фармакологии . 152 (5): 725–33. дои : 10.1038/sj.bjp.0707397 . ПМК 2190027 . ПМИД 17641668 .
- ^ Перейти обратно: а б Дх, Корень; С, Чжан; диджей, Баркер; Дж. Миранда-Барриентос; Б, Лю; Хл, Ван; М, Моралес (19 июня 2018 г.). «Избирательное распределение в мозгу и отличительная синаптическая архитектура двойных глутаматергических-ГАМКергических нейронов» . Отчеты по ячейкам . 23 (12): 3465–3479. дои : 10.1016/j.celrep.2018.05.063 . ПМЦ 7534802 . ПМИД 29924991 .
- ^ Адамс Х.Х., Хибар Д.П., Чураки В., Стейн Дж.Л., Найквист П.А., Рентерия М.Е. и др. (декабрь 2016 г.). «Новые генетические локусы, лежащие в основе внутричерепного объема человека, выявлены посредством общегеномной ассоциации» . Природная неврология . 19 (12): 1569–1582. дои : 10.1038/nn.4398 . ПМК 5227112 . ПМИД 27694991 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|