Ауторецептор
Ауторецептор расположенный – это тип рецептора, в мембранах нервных клеток . Он служит частью петли отрицательной обратной связи при передаче сигнала . Он чувствителен только к нейротрансмиттерам или гормонам, выделяемым нейроном, на котором расположен ауторецептор. Точно так же гетерорецептор чувствителен к нейротрансмиттерам и гормонам, которые не выделяются клеткой, на которой он расположен. Данный рецептор может действовать как ауторецептор или как гетерорецептор, в зависимости от типа передатчика, выделяемого клеткой, в которую он встроен.
Ауторецепторы могут располагаться в любой части клеточной мембраны: в дендритах , теле клетки , аксоне или окончаниях аксона . [1]
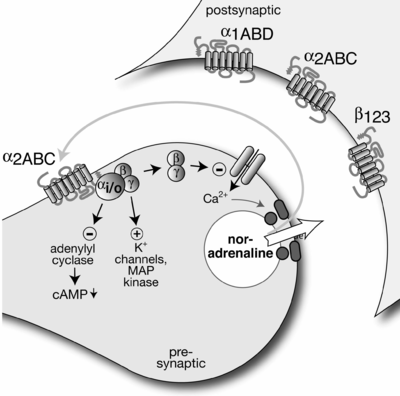
Канонически пресинаптический нейрон высвобождает нейромедиатор через синаптическую щель , который обнаруживается рецепторами постсинаптического нейрона. Ауторецепторы пресинаптического нейрона также обнаруживают этот нейромедиатор и часто контролируют внутренние клеточные процессы, обычно подавляя дальнейшее высвобождение или синтез нейромедиатора. Таким образом, высвобождение нейромедиатора регулируется отрицательной обратной связью. Ауторецепторы обычно представляют собой рецепторы, связанные с G-белком (а не ионные каналы, управляемые трансмиттером ), и действуют через вторичный мессенджер . [2]
Примеры
[ редактировать ]Ингибирование ауторецепторов приводит к увеличению высвобождения соответствующих нейромедиаторов. Основными клинически важными ауторецепторами являются альфа 2 ( подтип 2 адренергических рецепторов), H 3 ( подтип 3 гистаминовых рецепторов), 5 HT 1 ( подтип 1 серотониновых рецепторов). Соответствующими препаратами являются клонидин альфа-2 в качестве агониста, используемого при гипертонии , который снижает высвобождение норэпинефрина и адреналина из пресинаптических нейронов. Тизанидин, используемый в качестве релаксанта скелетных мышц центрального действия, также действует на альфа-2. 5HT 1A является мишенью буспирона , который действует как частичный агонист и используется как атипичный неседативный анксиолитик. Агонистами рецепторов 5HT 1B/1D являются триптаны и алкалоиды спорыньи , которые используются в лечение мигрени. Подтип рецептора 5 HT 1F. Агонист лекарственного препарата лесмидитан, также используемый при лечении мигрени. Антагонист H 3 -рецепторов Питолисант, используемый при нарколепсии. Например, норадреналин, высвобождаемый симпатическими нейронами, может взаимодействовать с адренорецепторами альфа-2А и альфа-2С, ингибируя дальнейшее высвобождение норадреналина. Сходным образом, ацетилхолин, высвобождаемый из парасимпатических нейронов, может взаимодействовать с рецепторами М 2 и М 4, ингибируя дальнейшее высвобождение ацетилхолина. Атипичным примером является β-адренергический ауторецептор в симпатической периферической нервной системе , который увеличивает высвобождение медиатора. [1]
рецептором микроэлементов Недавно было показано, что ауторецептор D2sh взаимодействует с 1 (TAAR1), G-сопряженным белковым рецептором GPCR , для регуляции моноаминергических систем в мозге. [3] Активный TAAR1 противодействует активности ауторецептора, инактивируя транспортер дофамина (DAT). [4] В своем обзоре TAAR1 в моноаминергических системах Се и Миллер предложили следующую схему: синаптический дофамин связывается с дофаминовым ауторецептором, который активирует DAT. Дофамин проникает в пресинаптические клетки и связывается с TAAR1, что увеличивает аденилатциклазы активность . В конечном итоге это позволяет транслировать следовые количества аминов в цитоплазме и активировать циклические нуклеотид-управляемые ионные каналы , которые дополнительно активируют TAAR1 и сбрасывают дофамин в синапс. Посредством серии событий фосфорилирования, связанных с PKA и PKC , активный TAAR1 инактивирует DAT, предотвращая захват дофамина из синапса. [5] Наличие двух постсинаптических рецепторов с противоположными способностями регулировать функцию переносчика моноаминов позволяет регулировать моноаминергическую систему.
Активность ауторецепторов также может снижать эффективность парных импульсов (PPF). [ нужна ссылка ] Клетка обратной связи активируется (частично) деполяризованным постсинаптическим нейроном. Клетка обратной связи высвобождает нейромедиатор, к которому восприимчив ауторецептор пресинаптического нейрона. Ауторецептор вызывает ингибирование кальциевых каналов (замедление притока ионов кальция) и открытие калиевых каналов (увеличение оттока ионов калия) в пресинаптической мембране. Эти изменения концентрации ионов эффективно уменьшают количество исходного нейромедиатора, высвобождаемого пресинаптическим окончанием в синаптическую щель. Это вызывает окончательное угнетение активности постсинаптического нейрона. Таким образом, цикл обратной связи завершен.

Ссылки
[ редактировать ]- ^ Перейти обратно: а б Сигел Г.Дж., Агранов Б.В., Альберс Р.В. и др., ред. (1999). «Катехоламиновые рецепторы». Базовая нейрохимия: молекулярные, клеточные и медицинские аспекты (6-е изд.). Липпинкотт-Рейвен.
- ^ Медведь; Коннорс; Парадизо (2006). Нейронаука: исследование мозга (3-е изд.). п. 119 .
- ^ Се Цз, WS (2007). «Передача сигналов аминоассоциированного рецептора 1 макаки-резуса: усиление переносчиками моноаминов и ослабление ауторецептором D2 in vitro». Журнал фармакологии и экспериментальной терапии . 321 (1): 116–127. дои : 10.1124/jpet.106.116863 . ПМИД 17234900 .
- ^ Се З, Уэстморленд С.В., Миллер Г.М. (2008). «Модуляция переносчиков моноаминов обычными биогенными аминами через следовые амин-ассоциированные рецепторы 1 и моноаминовые ауторецепторы в эмбриональных клетках почек 293 человека и синаптосомах мозга». Журнал фармакологии и экспериментальной терапии . 325 (2): 629–640. дои : 10.1124/jpet.107.135079 . ПМИД 18310473 .
- ^ Се З, Миллер GM (2009). «Отслеживание аминоассоциированного рецептора 1 как моноаминергического модулятора в мозге» . Биохимическая фармакология . 78 (9): 1095–1104. дои : 10.1016/j.bcp.2009.05.031 . ПМЦ 2748138 . ПМИД 19482011 .