Химический синапс

Химические синапсы представляют собой биологические соединения, через которые сигналы нейронов могут передаваться друг другу и ненейронным клеткам, например, клеткам мышц или желез . Химические синапсы позволяют нейронам образовывать цепи внутри центральной нервной системы . Они имеют решающее значение для биологических вычислений , лежащих в основе восприятия и мышления . Они позволяют нервной системе подключаться к другим системам организма и управлять ими.
В химическом синапсе один нейрон высвобождает нейромедиатора молекулы в небольшое пространство ( синаптическую щель ), прилегающее к другому нейрону. Нейромедиаторы содержатся в небольших мешочках, называемых синаптическими пузырьками , и высвобождаются в синаптическую щель путем экзоцитоза . Эти молекулы затем связываются с рецепторами нейромедиаторов на постсинаптической клетке. Наконец, нейротрансмиттеры выводятся из синапса с помощью одного из нескольких потенциальных механизмов, включая ферментативную деградацию или повторный захват либо специфическими переносчиками на пресинаптической клетке, либо на какой-либо другой нейроглии, чтобы прекратить действие нейромедиатора.
взрослого человека содержит По оценкам , мозг от 10 14 до 5 × 10 14 (100–500 триллионов) синапсов. [1] Каждый кубический миллиметр коры головного мозга содержит примерно миллиард ( короткий масштаб , т.е. 10 9 ) из них. [2] Количество синапсов в коре головного мозга человека оценивается отдельно в 0,15 квадриллиона (150 триллионов). [3]
Слово «синапс» было введено сэром Чарльзом Скоттом Шеррингтоном в 1897 году. [4] Химические синапсы — не единственный тип биологических синапсов : электрические и иммунологические синапсы существуют также . Однако без уточнения «синапс» обычно относится к химическим синапсам.
Структура
[ редактировать ]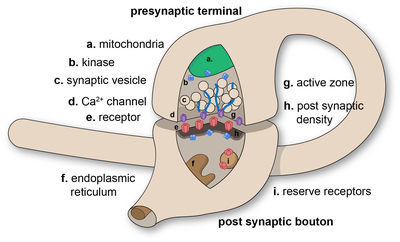
Синапсы — это функциональные связи между нейронами или между нейронами и другими типами клеток. [5] [6] Типичный нейрон дает несколько тысяч синапсов, хотя есть типы, которые образуют гораздо меньше синапсов. [7] Большинство синапсов соединяют аксоны с дендритами . [8] [9] но существуют и другие типы связей, в том числе между аксоном и телом клетки, [10] [11] аксон к аксону, [10] [11] и дендрит-дендрит . [9] Синапсы, как правило, слишком малы, чтобы их можно было распознать с помощью светового микроскопа, за исключением точек, где кажется, что мембраны двух клеток соприкасаются, но их клеточные элементы можно четко визуализировать с помощью электронного микроскопа .
Химические синапсы передают информацию направленно от пресинаптической клетки к постсинаптической и поэтому асимметричны по структуре и функциям. Пресинаптическое окончание аксона , или синаптический бутон, представляет собой специализированную область внутри аксона пресинаптической клетки, содержащую нейромедиаторы, заключенные в небольшие мембраносвязанные сферы, называемые синаптическими пузырьками (а также ряд других поддерживающих структур и органелл, таких как митохондрии и эндоплазматическая сеть ). Синаптические пузырьки прикрепляются к пресинаптической плазматической мембране в областях, называемых активными зонами .
Непосредственно напротив находится область постсинаптической клетки, содержащая рецепторы нейромедиаторов ; для синапсов между двумя нейронами постсинаптическая область может быть обнаружена на дендритах или теле клетки. Сразу за постсинаптической мембраной находится сложный комплекс взаимосвязанных белков, называемый постсинаптической плотностью (PSD).
Белки PSD участвуют в закреплении и транспортировке рецепторов нейромедиаторов, а также в модуляции активности этих рецепторов. Рецепторы и PSD часто обнаруживаются в специализированных выступах главного дендритного стержня, называемых дендритными шипами .
Синапсы можно описать как симметричные и асимметричные. При исследовании под электронным микроскопом асимметричные синапсы характеризуются округлыми пузырьками в пресинаптической клетке и выраженной постсинаптической плотностью. Асимметричные синапсы обычно являются возбуждающими. Симметричные синапсы, напротив, имеют уплощенные или удлиненные везикулы и не содержат заметной постсинаптической плотности. Симметричные синапсы обычно являются тормозными.
Синаптическая щель , также называемая синаптической щелью , представляет собой щель между пре- и постсинаптическими клетками шириной около 20 нм (0,02 мкм). [12] Небольшой объем расщелины позволяет быстро повышать и понижать концентрацию нейромедиаторов. [13]
Аутапс — это химический (или электрический) синапс, образующийся, когда аксон одного нейрона синапсирует с собственными дендритами.
Передача сигналов в химических синапсах
[ редактировать ]Обзор
[ редактировать ]Вот краткое изложение последовательности событий, которые происходят при синаптической передаче от пресинаптического нейрона к постсинаптической клетке. Каждый шаг описан более подробно ниже. Обратите внимание, что, за исключением последнего этапа, весь процесс может длиться всего несколько сотен микросекунд в самых быстрых синапсах. [14]
- Процесс начинается с волны электрохимического возбуждения, называемой потенциалом действия, движущейся по мембране пресинаптической клетки, пока она не достигнет синапса.
- Электрическая деполяризация мембраны в синапсе вызывает открытие каналов, проницаемых для ионов кальция.
- Ионы кальция проходят через пресинаптическую мембрану, быстро увеличивая концентрацию кальция внутри.
- Высокая концентрация кальция активирует набор чувствительных к кальцию белков, прикрепленных к везикулам , содержащим химический нейромедиатор .
- Эти белки меняют форму, вызывая слияние мембран некоторых «состыкованных» везикул с мембраной пресинаптической клетки, тем самым открывая везикулы и сбрасывая их нейротрансмиттерное содержимое в синаптическую щель — узкое пространство между мембранами пре- и постсинаптической клетки. клетки.
- Нейромедиатор диффундирует внутри расщелины. Часть его ускользает, но часть связывается с молекулами химических рецепторов, расположенными на мембране постсинаптической клетки.
- Связывание нейромедиатора приводит к активации каким-то образом молекулы рецептора. Возможны несколько типов активации, которые более подробно описаны ниже. В любом случае это ключевой этап, посредством которого синаптический процесс влияет на поведение постсинаптической клетки.
- Из-за тепловой вибрации , движения атомов, колеблющихся около своих положений равновесия в кристаллическом твердом теле, молекулы нейромедиаторов в конечном итоге отрываются от рецепторов и уносятся прочь.
- Нейромедиатор либо реабсорбируется пресинаптической клеткой, а затем переупаковывается для будущего высвобождения, либо метаболически расщепляется.
Высвобождение нейротрансмиттера
[ редактировать ]
Высвобождение нейромедиатора запускается при поступлении нервного импульса (или потенциала действия ) и происходит посредством необычайно быстрого процесса клеточной секреции ( экзоцитоза ). Внутри пресинаптического нервного окончания везикулы , содержащие нейромедиатор, локализуются вблизи синаптической мембраны. Поступающий потенциал действия вызывает приток ионов кальция через потенциал-зависимые, кальций-селективные ионные каналы при движении вниз потенциала действия (хвостовой ток). [15] Ионы кальция затем связываются с белками синаптотагмина , обнаруженными в мембранах синаптических везикул, позволяя везикулам сливаться с пресинаптической мембраной. [16] Слияние пузырька — стохастический процесс, приводящий к частым сбоям синаптической передачи в очень мелких синапсах, типичных для центральной нервной системы . большие химические синапсы (например, нервно-мышечные соединения С другой стороны, ) имеют вероятность синаптического высвобождения, по сути, равную 1. Слияние пузырьков управляется действием набора белков в пресинаптическом терминале, известных как SNARE . В целом белковый комплекс или структура, которая обеспечивает стыковку и слияние пресинаптических везикул, называется активной зоной. [17] Мембрана, добавленная в процессе слияния, позже извлекается путем эндоцитоза и перерабатывается для образования свежих везикул, наполненных нейротрансмиттерами.
Исключение из общей тенденции высвобождения нейромедиаторов путем слияния везикул обнаружено в рецепторных клетках типа II вкусовых рецепторов млекопитающих . Здесь нейротрансмиттер АТФ высвобождается непосредственно из цитоплазмы в синаптическую щель через потенциалзависимые каналы. [18]
Связывание рецептора
[ редактировать ]Рецепторы на противоположной стороне синаптической щели связывают молекулы нейромедиаторов. Рецепторы могут реагировать одним из двух основных способов. Во-первых, рецепторы могут напрямую открывать лиганд-управляемые ионные каналы в постсинаптической клеточной мембране, заставляя ионы проникать в клетку или выходить из нее и изменять локальный трансмембранный потенциал . [14] Результирующее изменение напряжения называется постсинаптическим потенциалом . В общем случае результат является возбуждающим в случае деполяризующих токов и тормозящим в случае гиперполяризующих токов. Является ли синапс возбуждающим или тормозящим, зависит от того, какой тип ионных каналов проводит постсинаптический ток (токи), что, в свою очередь, зависит от типа рецепторов и нейромедиатора, используемых в синапсе. Второй способ, которым рецептор может влиять на мембранный потенциал, — это модуляция производства химических медиаторов внутри постсинаптического нейрона. Эти вторичные мессенджеры могут затем усиливать тормозящую или возбуждающую реакцию на нейротрансмиттеры. [14]
Прекращение действия
[ редактировать ]После того, как молекула нейромедиатора связывается с молекулой рецептора, ее необходимо удалить, чтобы постсинаптическая мембрана могла продолжать передавать последующие ВПСП и/или ТПСП . Это удаление может произойти посредством одного или нескольких процессов:
- Нейромедиатор может диффундировать из-за термически вызванных колебаний как его, так и рецептора, что делает его доступным для метаболического расщепления вне нейрона или для реабсорбции. [19]
- Ферменты внутри субсинаптической мембраны могут инактивировать/метаболизировать нейромедиатор.
- Насосы обратного захвата могут активно перекачивать нейротрансмиттер обратно в пресинаптическое окончание аксона для переработки и повторного высвобождения после более позднего потенциала действия. [19]
Синаптическая сила
[ редактировать ]Сила синапса была определена Бернардом Кацем как произведение вероятности (пресинаптического) высвобождения pr , размера квантов q (постсинаптический ответ на высвобождение одного пузырька нейромедиатора, «кванта») и n , числа сайты релизов. «Унитарное соединение» обычно относится к неизвестному количеству отдельных синапсов, соединяющих пресинаптический нейрон с постсинаптическим нейроном. Амплитуда постсинаптических потенциалов (ПСП) может составлять от 0,4 мВ до 20 мВ. [20] Амплитуда ПСП может модулироваться нейромодуляторами или изменяться в результате предыдущей активности. Изменения синаптической силы могут быть кратковременными, продолжающимися от нескольких секунд до минут, или долгосрочными ( долговременная потенциация , или ДП), продолжающимися часами. Считается, что обучение и память являются результатом долгосрочных изменений синаптической силы посредством механизма, известного как синаптическая пластичность .
Десенсибилизация рецепторов
[ редактировать ]Десенсибилизация постсинаптических рецепторов – это снижение реакции на один и тот же нейромедиаторный стимул. Это означает, что сила синапса может фактически уменьшаться по мере того, как серия потенциалов действия прибывает в быстрой последовательности – явление, которое приводит к так называемой частотной зависимости синапсов. Нервная система использует это свойство в вычислительных целях и может настраивать свои синапсы с помощью таких средств, как фосфорилирование задействованных белков.
Синаптическая пластичность
[ редактировать ]Синаптическая передача может быть изменена предыдущей деятельностью. Эти изменения называются синаптической пластичностью и могут привести либо к снижению эффективности синапса, называемому депрессией, либо к увеличению эффективности, называемому потенциацией. Эти изменения могут быть как долгосрочными, так и краткосрочными. Формы краткосрочной пластичности включают синаптическое утомление или депрессию и увеличение синапсов . Формы долгосрочной пластичности включают долговременную депрессию и долговременную потенциацию . Синаптическая пластичность может быть либо гомосинаптической (происходящей в одном синапсе), либо гетеросинаптической (происходящей в нескольких синапсах).
Гомосинаптическая пластичность
[ редактировать ]Гомосинаптическая пластичность (или также гомотропная модуляция) — это изменение синаптической силы, возникающее в результате истории активности конкретного синапса. Это может быть результатом изменений в пресинаптическом кальции, а также обратной связи с пресинаптическими рецепторами, т.е. формой аутокринной передачи сигналов . Гомосинаптическая пластичность может влиять на количество и скорость пополнения везикул или на взаимосвязь между высвобождением кальция и везикул. Гомосинаптическая пластичность может иметь и постсинаптическую природу. Это может привести как к увеличению, так и к уменьшению синаптической силы.
Одним из примеров являются нейроны симпатической нервной системы (СНС), выделяющие норадреналин , который, помимо воздействия на постсинаптические рецепторы, влияет и на пресинаптические α2-адренорецепторы , ингибируя дальнейшее высвобождение норадреналина. [21] Этот эффект используется вместе с клонидином для оказания ингибирующего воздействия на СНС.
Гетеросинапсическая пластичность
[ редактировать ]Гетеросинаптическая пластичность (или также гетеротропная модуляция) — это изменение синаптической силы, возникающее в результате активности других нейронов. Опять же, пластичность может изменять количество везикул или скорость их пополнения, а также взаимосвязь между высвобождением кальция и везикул. Кроме того, это может напрямую влиять на приток кальция. Гетеросинаптическая пластичность может иметь и постсинаптический характер, влияя на чувствительность рецепторов.
Одним из примеров снова являются нейроны симпатической нервной системы , выделяющие норадреналин , который, кроме того, оказывает тормозящее действие на пресинаптические окончания нейронов парасимпатической нервной системы . [21]
Интеграция синаптических входов
[ редактировать ]В общем, если возбуждающий синапс достаточно силен, потенциал действия в пресинаптическом нейроне вызовет потенциал действия в постсинаптической клетке. Во многих случаях возбуждающий постсинаптический потенциал (ВПСП) не достигает порога возникновения потенциала действия. Когда потенциалы действия от нескольких пресинаптических нейронов срабатывают одновременно или если один пресинаптический нейрон срабатывает с достаточно высокой частотой, ВПСП могут перекрываться и суммироваться. Если перекрывается достаточное количество ВПСП, суммарное ВПСП может достичь порога инициации потенциала действия. Этот процесс известен как суммирование и может служить фильтром верхних частот для нейронов. [22]
С другой стороны, пресинаптический нейрон, высвобождающий тормозной нейромедиатор, такой как ГАМК , может вызвать тормозящий постсинаптический потенциал (ТПСП) в постсинаптическом нейроне, отодвигая мембранный потенциал дальше от порога, уменьшая его возбудимость и затрудняя его работу. нейрон инициирует потенциал действия. Если ТПСП перекрывается с ВПСП, то ТПСП во многих случаях может помешать нейрону запустить потенциал действия. Таким образом, выходной сигнал нейрона может зависеть от входного сигнала множества разных нейронов, каждый из которых может иметь разную степень влияния, в зависимости от силы и типа синапса с этим нейроном. Джон Кэрью Эклс выполнил некоторые важные ранние эксперименты по синаптической интеграции, за которые он получил Нобелевскую премию по физиологии и медицине в 1963 году.
Передача объема
[ редактировать ]Когда нейротрансмиттер высвобождается в синапсе, он достигает своей максимальной концентрации внутри узкого пространства синаптической щели, но некоторая часть его обязательно диффундирует, прежде чем реабсорбируется или разрушается. Если он диффундирует, он может активировать рецепторы, расположенные либо в других синапсах, либо на мембране вдали от синапсов. Внесинаптическая активность нейромедиатора известна как объемная передача . [23] Хорошо известно, что такие эффекты в некоторой степени имеют место, но их функциональное значение уже давно является предметом споров. [24]
Недавние работы показывают, что передача объема может быть преобладающим способом взаимодействия для некоторых особых типов нейронов. В коре головного мозга млекопитающих класс нейронов, называемых нейроглиаформными клетками, может ингибировать другие близлежащие кортикальные нейроны, высвобождая нейротрансмиттер ГАМК во внеклеточное пространство. [25] По тому же принципу ГАМК, высвобождаемая из нейроглиаформных клеток во внеклеточное пространство, также действует на окружающие астроциты , определяя роль объемной передачи в контроле гомеостаза ионов и нейромедиаторов. [26] Примерно 78% бутонов нейроглиаформных клеток не образуют классических синапсов. Это может быть первый окончательный пример химического взаимодействия нейронов там, где отсутствуют классические синапсы. [25]
Связь с электрическими синапсами
[ редактировать ]Электрический синапс — это электропроводящая связь между двумя прилегающими нейронами , которая образуется в узком промежутке между пре- и постсинаптическими клетками , известном как щелевое соединение . В щелевых соединениях клетки приближаются друг к другу на расстояние примерно 3,5 нм , а не на расстояние от 20 до 40 нм, которое разделяет клетки в химических синапсах. [27] [28] В отличие от химических синапсов, постсинаптический потенциал в электрических синапсах обусловлен не открытием ионных каналов химическими передатчиками, а скорее прямой электрической связью между обоими нейронами. Электрические синапсы работают быстрее, чем химические. [13] Электрические синапсы встречаются по всей нервной системе, в том числе в сетчатке , ретикулярном ядре таламуса , неокортексе и в гиппокампе . [29] Хотя химические синапсы обнаруживаются как между возбуждающими, так и между тормозными нейронами, электрические синапсы чаще всего обнаруживаются между более мелкими локальными тормозными нейронами. Электрические синапсы могут существовать между двумя аксонами, двумя дендритами или между аксоном и дендритом. [30] [31] У некоторых рыб и амфибий электрические синапсы можно обнаружить в пределах того же терминаля химического синапса, что и в клетках Маутнера . [32]
Эффекты наркотиков
[ редактировать ]Одной из важнейших особенностей химических синапсов является то, что они являются местом действия большинства психоактивных веществ . На синапсы влияют такие наркотики, как кураре, стрихнин, кокаин, морфин, алкоголь, ЛСД и многие другие. Эти препараты по-разному влияют на синаптическую функцию и часто ограничиваются синапсами, которые используют определенный нейромедиатор. Например, кураре — это яд, который не дает ацетилхолину деполяризовать постсинаптическую мембрану, вызывая паралич . Стрихнин блокирует тормозящее действие нейромедиатора глицина , который заставляет организм улавливать и реагировать на более слабые и ранее игнорированные раздражители, что приводит к неконтролируемым мышечным спазмам . Морфин действует на синапсы, которые используют нейротрансмиттеры эндорфины , а алкоголь усиливает тормозящее действие нейромедиатора ГАМК . ЛСД влияет на синапсы, которые используют нейромедиатор серотонин . Кокаин блокирует обратный захват дофамина и, следовательно, усиливает его действие.
История и этимология
[ редактировать ]В 1950-е годы Бернард Кац и Пол Фатт лягушки наблюдали спонтанные миниатюрные синаптические токи в нервно-мышечных соединениях . [33] На основе этих наблюдений они разработали «квантовую гипотезу», которая является основой нашего нынешнего понимания высвобождения нейромедиаторов как экзоцитоза и за которую Кац получил Нобелевскую премию по физиологии и медицине в 1970 году. [34] В конце 1960-х годов Рикардо Миледи и Кац выдвинули гипотезу о том, что вызванный деполяризацией приток ионов кальция запускает экзоцитоз .
Сэр Чарльз Скотт Шеррингтон придумал слово «синапс», а историю этого слова Шеррингтон изложил в письме, которое он написал Джону Фултону:
«Я чувствовал потребность в каком-то имени, чтобы назвать соединение между нервной клеткой и нервной клеткой... Я предложил использовать слово «синдесм»... Он [ сэр Майкл Фостер со своим другом по Троице Верраллом , ученым- еврипидом ] посоветовался по этому поводу . и Верралл предложил слово «синапс» (от греческого «застежка»).' – Чарльз Скотт Шеррингтон [4]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Драхман Д. (2005). «Есть ли у нас лишние мозги?». Неврология . 64 (12): 2004–5. дои : 10.1212/01.WNL.0000166914.38327.BB . ПМИД 15985565 . S2CID 38482114 .
- ^ Алонсо-Нанкларес Л., Гонсалес-Сориано Х., Родригес-младший, ДеФелипе Х. (сентябрь 2008 г.). «Гендерные различия в плотности корковых синапсов человека» . Учеб. Натл. акад. наук. США . 105 (38): 14615–9. Бибкод : 2008PNAS..10514615A . дои : 10.1073/pnas.0803652105 . ПМЦ 2567215 . ПМИД 18779570 .
- ^ Факты и цифры о мозге, Вашингтонский университет.
- ^ Перейти обратно: а б Коуэн, В. Максвелл; Зюдхоф, Томас К.; Стивенс, Чарльз Ф. (2003). Синапсы . Джу Пресс. п. 11. ISBN 9780801871184 . Проверено 9 июня 2020 г.
- ^ Раппорт, Ричард Л. (2005). Нервные окончания: открытие синапса . WW Нортон и компания. стр. 1–37. ISBN 978-0-393-06019-5 .
- ^ Сквайр, Ларри Р.; Флойд Блум; Николас Спитцер (2008). Фундаментальная нейронаука . Академическая пресса. стр. 425–6. ISBN 978-0-12-374019-9 .
- ^ Хайман, Стивен Э.; Эрик Джонатан Нестлер (1993). Молекулярные основы психиатрии . Американский психиатрический паб. стр. 425–6. ISBN 978-0-88048-353-7 .
- ^ Смилкштейн, Рита (2003). Мы рождены, чтобы учиться: использование естественного процесса обучения мозга для создания современной учебной программы . Корвин Пресс. п. 56. ИСБН 978-0-7619-4642-7 .
- ^ Перейти обратно: а б Литтон, Уильям В. (2002). От компьютера к мозгу: основы вычислительной нейронауки . Спрингер. п. 28. ISBN 978-0-387-95526-1 . Аксоны, соединяющие дендрит с дендритом, представляют собой дендродендритные синапсы. Аксоны, соединяющие аксон с дендритом, называются аксодендритическим синапсом.
- ^ Перейти обратно: а б Гарбер, Стивен Д. (2002). Биология: Самоучитель . Джон Уайли и сыновья. п. 175 . ISBN 978-0-471-22330-6 .
синапсы соединяют аксоны с телом клетки.
- ^ Перейти обратно: а б Вайс, Мирин; д-р Стивен М. Мирин; Доктор Роксана Бартель (1994). Кокаин . Американский психиатрический паб. п. 52. ИСБН 978-1-58562-138-5 . Проверено 26 декабря 2008 г. Аксоны, оканчивающиеся на теле постсинаптической клетки, представляют собой аксосоматические синапсы. Аксоны, оканчивающиеся на аксонах, представляют собой аксоаксонные синапсы.
- ^ Уидроу, Бернард; Ким, Янгсик; Пак, Дукан; Перин, Хосе Краузе (2019). «Правило обучения природы». Искусственный интеллект в эпоху нейронных сетей и мозговых вычислений . Эльзевир. стр. 1–30. дои : 10.1016/b978-0-12-815480-9.00001-3 . ISBN 978-0-12-815480-9 . S2CID 125516633 .
- ^ Перейти обратно: а б Кандел, Шварц и Джесселл 2000 , с. 182
- ^ Перейти обратно: а б с Медведь, Марк Ф; Коннорс, Барри В.; Парадизо, Майкл А. (2007). Нейронаука: исследование мозга . Филадельфия, Пенсильвания: Липпинкотт Уильямс и Уилкинс. С. 113–118 .
- ^ Ллинас Р., Стейнберг И.З., Уолтон К. (1981). «Связь между пресинаптическим током кальция и постсинаптическим потенциалом в гигантском синапсе кальмара» . Биофизический журнал . 33 (3): 323–351. Бибкод : 1981BpJ....33..323L . дои : 10.1016/S0006-3495(81)84899-0 . ПМЦ 1327434 . ПМИД 6261850 .
- ^ Чепмен, Эдвин Р. (2002). «Синаптотагмин: сенсор Ca2+, запускающий экзоцитоз?». Nature Reviews Молекулярно-клеточная биология . 3 (7): 498–508. дои : 10.1038/nrm855 . ISSN 1471-0080 . ПМИД 12094216 . S2CID 12384262 .
- ^ Крейг К. Гарнер и Кан Шен. Структура и функции активных зон позвоночных и беспозвоночных. Структура и функциональная организация синапса. Эд: Йоханнес Хелл и Майкл Элерс. Спрингер, 2008.
- ^ Романов Роман А.; Лэшер, Роберт С.; Высоко, Бриджит; Сэвидж, Логан Э.; Лоусон, Адам; Рогачевская Ольга Александровна; Чжао, Гаитян; Рогачевский Вадим Владимирович; Быстрова Марина Ф.; Чурбанов Глеб Д.; Адамейко Игорь; Харкани, Тибор; Ян, Жуйбяо; Кидд, Грэм Дж.; Марамбо, Филипп; Киннамон, Джон К.; Колесников Станислав С.; Фингер, Томас Э. (2018). «Химические синапсы без синаптических везикул: пуринергическая нейротрансмиссия через канал CALHM1-митохондриальный сигнальный комплекс» . Научная сигнализация . 11 (529): eaao1815. дои : 10.1126/scisignal.aao1815 . ISSN 1945-0877 . ПМК 5966022 . ПМИД 29739879 .
- ^ Перейти обратно: а б Шервуд Л., Стикавы (2007). Физиология человека 6e: от клеток к системам
- ^ Диас-Риос М., Миллер М.В. (июнь 2006 г.). «Целевая регуляция синаптической эффективности в питающем центральном генераторе паттернов аплизии: потенциальные субстраты для поведенческой пластичности?» . Биол. Бык . 210 (3): 215–29. дои : 10.2307/4134559 . JSTOR 4134559 . ПМИД 16801496 . S2CID 34154835 .
- ^ Перейти обратно: а б Ранг, HP; Дейл, ММ; Риттер, Дж. М. (2003). Фармакология (5-е изд.). Эдинбург: Черчилль Ливингстон. п. 129. ИСБН 978-0-443-07145-4 .
- ^ Брюс Альбертс; Александр Джонсон; Джулиан Льюис; Мартин Рафф; Кейт Робертс; Питер Уолтер, ред. (2002). «Гл. 11. Раздел: Одиночные нейроны — сложные вычислительные устройства» . Молекулярная биология клетки (4-е изд.). Гирляндная наука. ISBN 978-0-8153-3218-3 .
- ^ Золи М., Торри С., Феррари Р. и др. (1998). «Появление концепции объемной передачи». Мозговой Рес. Мозговой Рес. Преподобный . 26 (2–3): 136–47. дои : 10.1016/S0165-0173(97)00048-9 . ПМИД 9651506 . S2CID 20495134 .
- ^ Фуксе К., Дальстрем А., Хойстад М. и др. (2007). «От картирования Гольджи-Кахаля до характеристики нейронных сетей на основе передатчиков, ведущих к двум режимам коммуникации мозга: проводной и объемной передаче» (PDF) . Мозговой Res Rev. 55 (1): 17–54. дои : 10.1016/j.brainresrev.2007.02.009 . hdl : 10447/9980 . ПМИД 17433836 . S2CID 1323780 .
- ^ Перейти обратно: а б Ола С., Фюле М., Комлоши Г. и др. (2009). «Регуляция корковых микросхем путем унитарной ГАМК-опосредованной объемной передачи» . Природа . 461 (7268): 1278–81. Бибкод : 2009Natur.461.1278O . дои : 10.1038/nature08503 . ПМЦ 2771344 . ПМИД 19865171 .
- ^ Рожа М., Бака Дж., Борде С., Рожа Б., Катона Г., Тамаш Г. и др. (2015). «Унитарная ГАМКергическая передача объема от отдельных интернейронов к астроцитам в коре головного мозга» (PDF) . Структура и функции мозга . 222 (1): 651–659. дои : 10.1007/s00429-015-1166-9 . ПМИД 26683686 . S2CID 30728927 .
- ^ Кандель, Шварц и Джесселл 2000 , стр. 176
- ^ Хормузди и др. 2004 г.
- ^ Коннорс Б.В., Лонг, Массачусетс (2004). «Электрические синапсы в мозгу млекопитающих» . Анну. Преподобный Нейроски . 27 (1): 393–418. дои : 10.1146/annurev.neuro.26.041002.131128 . ПМИД 15217338 .
- ^ Веруки М.Л., Хартвейт Э. (декабрь 2002 г.). «Электрические синапсы опосредуют передачу сигналов по палочковидному пути сетчатки млекопитающих» . Дж. Нейроски . 22 (24): 10558–66. doi : 10.1523/JNEUROSCI.22-24-10558.2002 . ПМЦ 6758447 . ПМИД 12486148 .
- ^ Беннетт М.В., Паппас Г.Д., Алджуре Э., Накадзима Ю. (март 1967 г.). «Физиология и ультраструктура электротонических соединений. II. Спинальные и медуллярные электромоторные ядра мормирид». Дж. Нейрофизиология . 30 (2): 180–208. дои : 10.1152/jn.1967.30.2.180 . ПМИД 4167209 .
- ^ Переда А.Е., Раш Дж.Е., Надь Дж.И., Беннетт М.В. (декабрь 2004 г.). «Динамика электрической передачи на трефовых окончаниях клеток Маутнера». Мозговой Рес. Мозговой Рес. Преподобный . 47 (1–3): 227–44. CiteSeerX 10.1.1.662.9352 . дои : 10.1016/j.brainresrev.2004.06.010 . ПМИД 15572174 . S2CID 9527518 .
- ^ Августин, Джордж Дж.; Касаи, Харуо (1 февраля 2007 г.). «Бернард Кац, квантовое высвобождение медиатора и основы пресинаптической физиологии» . Журнал физиологии . 578 (Часть 3): 623–625. дои : 10.1113/jphysicalol.2006.123224 . ПМК 2151334 . ПМИД 17068096 .
- ^ «Нобелевская премия» . Британский медицинский журнал . 4 (5729): 190. 24 октября 1970 г. дои : 10.1136/bmj.4.5729.190 . ПМК 1819734 . ПМИД 4320287 .
Ссылки
[ редактировать ]- Карлсон, Нил Р. (2007). Физиология поведения (9-е изд.). Бостон, Массачусетс: Pearson Education. ISBN 978-0-205-59389-7 .
- Кандел, Эрик Р .; Шварц, Джеймс Х.; Джесселл, Томас М. (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-0-8385-7701-1 .
- Ллинас Р., Сугимори М., Саймон С.М. (апрель 1982 г.). «Передача посредством пресинаптической спайкообразной деполяризации в гигантском синапсе кальмара» . Учеб. Натл. акад. наук. США . 79 (7): 2415–9. Бибкод : 1982PNAS...79.2415L . дои : 10.1073/pnas.79.7.2415 . ПМК 346205 . ПМИД 6954549 .
- Ллинас Р., Стейнберг И.З., Уолтон К. (1981). «Связь между пресинаптическим током кальция и постсинаптическим потенциалом в гигантском синапсе кальмара» . Биофизический журнал . 33 (3): 323–352. Бибкод : 1981BpJ....33..323L . дои : 10.1016/S0006-3495(81)84899-0 . ПМЦ 1327434 . ПМИД 6261850 .
- Медведь, Марк Ф.; Коннорс, Барри В.; Парадизо, Майкл А. (2001). Нейронаука: исследование мозга . Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-3944-3 .
- Хормузди, С.Г.; Филиппов, М.А.; Митропулу, Дж; Моньер, Х; Бруззоне, Р. (март 2004 г.). «Электрические синапсы: динамическая сигнальная система, формирующая активность нейронных сетей». Биохим Биофиз Акта . 1662 (1–2): 113–137. дои : 10.1016/j.bbamem.2003.10.023 . ПМИД 15033583 .
- Карп, Джеральд (2005). Клеточная и молекулярная биология: концепции и эксперименты (4-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. ISBN 978-0-471-46580-5 .
- Николлс, Дж.Г.; Мартин, Арканзас; Уоллес, Б.Г.; Фукс, Пенсильвания (2001). От нейрона к мозгу (4-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-439-3 .
Внешние ссылки
[ редактировать ]- Обзор Synapse для детей
- Synapse – клеточно-центрированная база данных
- Атлас ультраструктурной нейроцитологии. Галерея изображений синапсов и других нейрональных структур электронного микроскопа, собранная лабораторией Кристен Харрис.