Эндоцитоз
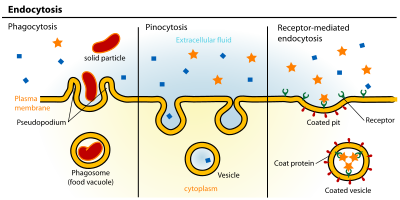
Эндоцитоз – это клеточный процесс , при котором вещества доставляются в клетку. Материал, подлежащий интернализации, окружен участком клеточной мембраны , который затем отпочковывается внутри клетки, образуя везикулу, содержащую проглоченные материалы. Эндоцитоз включает пиноцитоз (питание клеток) и фагоцитоз (поедание клеток). Это форма активного транспорта.
История
[ редактировать ]Термин был предложен Де Дювом в 1963 году. [1] Фагоцитоз был открыт Эли Мечниковым в 1882 году. [2]
Пути
[ редактировать ]
Пути эндоцитоза можно разделить на четыре категории: а именно, рецептор-опосредованный эндоцитоз (также известный как клатрин-опосредованный эндоцитоз), кавеолы , пиноцитоз и фагоцитоз . [3]
- Клатрин-опосредованный эндоцитоз опосредуется образованием небольших (диаметром около 100 нм) везикул, имеющих морфологически характерную оболочку, состоящую из цитозольного белка клатрина . [4] Покрытые клатрином везикулы (CCV) обнаруживаются практически во всех клетках и образуют домены плазматической мембраны, называемые ямками, покрытыми клатрином. Покрытые ямки могут концентрировать крупные внеклеточные молекулы, которые имеют различные рецепторы , ответственные за рецептор-опосредованный эндоцитоз лигандов, например липопротеинов низкой плотности , трансферрина , факторов роста , антител и многих других. [5]
- Изучать [6] в клетках млекопитающих подтверждают уменьшение размера клатриновой оболочки в среде повышенного напряжения. Кроме того, это предполагает, что два очевидно различных способа сборки клатрина, а именно покрытые ямки и покрытые бляшки, наблюдаемые в экспериментальных исследованиях, могут быть следствием различных напряжений в плазматической мембране.
- Кавеолы представляют собой наиболее часто встречающиеся зачатки плазматической мембраны, не покрытые клатрином, которые существуют на поверхности многих, но не всех типов клеток. Они состоят из холестерин-связывающего белка кавеолина (Vip21) с двойным слоем, обогащенным холестерином и гликолипидами . Кавеолы представляют собой небольшие (диаметром около 50 нм) ямки в мембране колбовидной формы, напоминающие форму пещеры (отсюда и название кавеолы). Они могут составлять до трети площади плазматической мембраны клеток некоторых тканей, особенно много их в гладкомышечных клетках I типа , пневмоцитах , фибробластах , адипоцитах и эндотелиальных клетках . [7] Считается также, что поглощение внеклеточных молекул специфически опосредовано через рецепторы в кавеолах.

Слева направо: фагоцитоз, пиноцитоз, рецептор-опосредованный эндоцитоз. - Потоцитоз - это форма рецептор-опосредованного эндоцитоза, при которой кавеолы используются для доставки молекул различного размера в клетку. В отличие от большинства эндоцитозов, при которых кавеолы используются для доставки содержимого везикул к лизосомам или другим органеллам, материал, эндоцитозированный посредством потоцитоза, высвобождается в цитозоль. [8]
- Пиноцитоз , который обычно возникает в сильно взъерошенных участках плазматической мембраны, представляет собой инвагинацию клеточной мембраны с образованием кармана, который затем защемляется в клетку, образуя везикулу (диаметром 0,5–5 мкм), заполненную большим объемом. внеклеточной жидкости и молекул внутри нее (эквивалентно ~ 100 CCV). Заполнение кармана происходит неспецифическим образом. Затем везикула перемещается в цитозоль и сливается с другими везикулами, такими как эндосомы и лизосомы . [9]
- Фагоцитоз — это процесс, посредством которого клетки связывают и усваивают твердые частицы диаметром более 0,75 мкм, такие как мелкие частицы пыли, клеточный мусор, микроорганизмы и апоптотические клетки. Эти процессы включают поглощение более крупных площадей мембраны, чем клатрин-опосредованный эндоцитоз и кавеол . путь

Более поздние эксперименты показали, что эти морфологические описания эндоцитарных событий могут быть неадекватными, и более подходящий метод классификации может быть основан на том, зависят ли определенные пути от клатрина и динамина .
Динаминозависимые и клатриннезависимые пути включают FEME , UFE , ADBE , EGFR-NCE и IL2Rβ. поглощение [10]
Динаминонезависимые и клатриннезависимые пути включают путь CLIC/GEEC (регулируемый Graf1 ), [11] а также MEND и макропиноцитоз . [10]
Клатрин-опосредованный эндоцитоз — единственный путь, зависящий как от клатрина, так и от динамина.
Основные компоненты
[ редактировать ]Эндоцитарный путь клеток млекопитающих состоит из отдельных мембранных компартментов, которые усваивают молекулы из плазматической мембраны и возвращают их обратно на поверхность (как в ранних эндосомах и рециркулирующих эндосомах) или сортируют их для деградации (как в поздних эндосомах и лизосомах). Основными компонентами эндоцитарного пути являются: [3]
- Ранние эндосомы представляют собой первый компартмент эндоцитарного пути. Ранние эндосомы часто расположены на периферии клетки и принимают большинство типов везикул, поступающих с поверхности клетки. Они имеют характерную тубуло-везикулярную структуру (везикулы диаметром до 1 мкм с соединенными канальцами диаметром около 50 нм) и слабокислый pH. В основном они представляют собой сортирующие органеллы, в которых многие эндоцитированные лиганды диссоциируют от своих рецепторов при кислом pH компартмента и из которых многие рецепторы возвращаются на поверхность клетки (через трубочки). [12] [13] Это также место сортировки трансцитотического пути к более поздним компартментам (например, поздним эндосомам или лизосомам) через трансвезикулярные компартменты (например, мультивезикулярные тельца (MVB) или эндосомальные везикулы-носители (ECV)).
- Поздние эндосомы получают эндоцитированный материал по пути к лизосомам , обычно из ранних эндосом эндоцитозного пути, из транс-сети Гольджи (TGN) в биосинтетическом пути и из фагосом в фагоцитарном пути. [14] Поздние эндосомы часто содержат белки, характерные для нуклеосом, митохондрий и мРНК, включая лизосомальные мембранные гликопротеины и кислые гидролазы. Они кислые (приблизительно pH 5,5) и являются частью пути транспортировки рецепторов маннозо-6-фосфата . Считается, что поздние эндосомы опосредуют финальный набор событий сортировки перед доставкой материала в лизосомы.
- Лизосомы являются последним отделом эндоцитарного пути. Их основная функция — расщепление продуктов клеточных отходов, жиров, углеводов, белков и других макромолекул на простые соединения. Затем они возвращаются в цитоплазму в качестве новых материалов для строительства клеток. Для этого лизосомы используют около 40 различных типов гидролитических ферментов, все из которых производятся в эндоплазматическом ретикулуме, модифицируются в аппарате Гольджи и функционируют в кислой среде. [15] Приблизительный pH лизосомы составляет 4,8, и при электронной микроскопии (ЭМ) они обычно выглядят как большие вакуоли (диаметром 1-2 мкм), содержащие электронно-плотный материал. У них высокое содержание лизосомальных мембранных белков и активных лизосомальных гидролаз, но отсутствует маннозо-6-фосфатный рецептор. Их обычно считают основным гидролитическим отделом клетки. [16] [17]
Недавно было обнаружено, что эйзосома служит воротами эндоцитоза у дрожжей. [18]
Клатрин-опосредованный
[ редактировать ]Основным путем эндоцитоза в большинстве клеток и наиболее изученным является путь, опосредованный молекулой клатрина . [19] [20] Этот крупный белок способствует образованию покрытой ямки на внутренней поверхности плазматической мембраны клетки. Затем эта ямка отрастает в клетку, образуя покрытый везикулы в цитоплазме клетки. При этом он вносит в клетку не только небольшой участок поверхности клетки, но и небольшой объем жидкости извне клетки. [21] [22] [23]
Функция пальто деформирует донорскую мембрану для образования пузырька, а также участвует в отборе груза везикул. Комплексы оболочки, которые к настоящему времени хорошо охарактеризованы, включают белок оболочки-I (COP-I), COP-II и клатрин. [24] [25] Клатриновые оболочки участвуют в двух важнейших этапах транспорта: (i) рецептор-опосредованном и жидкофазном эндоцитозе от плазматической мембраны к ранней эндосоме и (ii) транспорте от TGN к эндосомам. При эндоцитозе клатриновая оболочка собирается на цитоплазматической стороне плазматической мембраны, образуя ямки, которые инвагинируют, чтобы отщипнуться (разрезать) и стать свободными CCV. В культивируемых клетках сборка CCV занимает ~1 мин, а каждую минуту может образовываться от нескольких сотен до тысячи и более. [26] Основным компонентом каркаса клатриновой оболочки является белок массой 190 кДа, называемый тяжелой цепью клатрина (CHC), который связан с белком массой 25 кДа, называемым легкой цепью клатрина (CLC), образуя трехногие тримеры, называемые трискелионами.
Везикулы избирательно концентрируют и исключают определенные белки во время формирования и не являются репрезентативными для мембраны в целом. Адаптеры AP2 представляют собой мультисубъединичные комплексы, выполняющие эту функцию на плазматической мембране. Наиболее изученными рецепторами, которые сосредоточены в покрытых оболочкой везикулах клеток млекопитающих, являются рецептор ЛПНП (который удаляет ЛПНП из циркулирующей крови), рецептор трансферрина (который доставляет ионы трехвалентного железа, связанные с трансферрином, в клетку) и некоторые гормональные рецепторы (такие как это для ЭФР ).
В любой момент около 25% плазматической мембраны фибробласта состоит из ямок, покрытых оболочкой. Поскольку покрытая ямка живет около минуты, прежде чем она прорастет в клетку, фибробласт занимает ее поверхность этим путем примерно раз в 50 минут. Покрытые везикулы, образующиеся из плазматической мембраны, имеют диаметр около 100 нм и время жизни, измеряемое несколькими секундами. После того, как оболочка сброшена, оставшийся везикула сливается с эндосомами и продвигается по эндоцитарному пути. Фактический процесс отпочкования, при котором ямка превращается в везикулу, осуществляется клатрином; Ему помогает набор цитоплазматических белков, в который входят динамин и адаптеры, такие как адаптин .
Покрытые ямки и везикулы были впервые обнаружены в тонких срезах ткани в электронном микроскопе Томасом Ф. Ротом и Китом Р. Портером . [27] Их важность для выведения ЛПНП из крови была обнаружена Ричардом Г. Андерсоном, Майклом С. Брауном и Джозефом Л. Гольдштейном в 1977 году. [28] Покрытые оболочкой везикулы были впервые очищены Барбарой Пирс , которая открыла молекулу клатриновой оболочки в 1976 году. [29]
Процессы и компоненты
[ редактировать ]Белки кавеолина, такие как кавеолин-1 ( CAV1 ), кавеолин-2 ( CAV2 ) и кавеолин-3 ( CAV3 ), играют важную роль в процессе формирования кавеол. Более конкретно, CAV1 и CAV2 ответственны за образование кавеол в немышечных клетках, тогда как CAV3 функционирует в мышечных клетках. Процесс начинается с синтеза CAV1 в ЭР , где он образует устойчивые к детергентам олигомеры . Затем эти олигомеры проходят через комплекс Гольджи , прежде чем достичь поверхности клетки, способствуя образованию кавеол. Образование кавеол также обратимо за счет разборки при определенных условиях, таких как повышенное натяжение плазматической мембраны. Эти определенные условия затем зависят от типа тканей, которые выражают кавеолярную функцию. Например, не все ткани, содержащие кавеолярные белки, имеют кавеолярную структуру, т.е. гематоэнцефалический барьер . [30] Хотя среди кавеол сохраняется множество морфологических особенностей, функции каждого белка CAV разнообразны. Одной из общих черт кавеолинов являются их гидрофобные участки потенциальных шпилечных структур, состоящих из α-спиралей . Вставка этих шпилеобразных α-спиралей образует оболочку кавеол, что приводит к искривлению мембраны. Помимо вставки, кавеолины также способны к олигомеризации, что в дальнейшем играет роль в искривлении мембраны. Недавние исследования также обнаружили, что полимераза I, фактор высвобождения транскрипта и реакция белка депривации сыворотки также играют роль в сборке кавеол. Помимо сборки кавеол, исследователи также обнаружили, что белки CAV1 могут также влиять на другие пути эндоцитоза. Когда CAV1 связывается с Cdc42 , CAV1 инактивирует его и регулирует активность Cdc42 во время мембранного транспорта. [31]
Механизмы
[ редактировать ]Процесс поглощения клетками зависит от наклона и хиральности составляющих молекул, вызывающих почкование мембраны. Поскольку такие хиральные и наклонные липидные молекулы, вероятно, находятся в форме «плота», исследователи предполагают, что образование кавеол также следует этому механизму, поскольку кавеолы также обогащены компонентами плота. Когда белки кавеолина связываются с внутренним листком через холестерин , мембрана начинает изгибаться, что приводит к спонтанному искривлению. Этот эффект обусловлен распределением силы, возникающей при связывании олигомера кавеолина с мембраной. Распределение силы затем изменяет натяжение мембраны, что приводит к отпочкованию и, в конечном итоге, к образованию пузырьков. [32]
Галерея
[ редактировать ]- Эндоцитоз. Например, коронавирус SARS-CoV-2 связывается с рецептором ACE2 эпителиальной клетки .
 Этап 1
Этап 1 Этап 2
Этап 2 Этап 3
Этап 3- Анимация эндоцитоза (1)
- Анимация эндоцитоза (2)



См. также
[ редактировать ]- Активный транспорт
- Эмпериполез
- RAP6 (белок 6, активирующий Rab5)
- Экзоцитоз
- Фагоцитоз
- Пиноцитоз
- Трансэндоцитоз
Ссылки
[ редактировать ]- ^ Михаэлис А., Грин М.М., Ригер Р. (1991). Глоссарий генетики: классический и молекулярный (Пятое изд.). Берлин: Springer-Verlag. ISBN 978-3-642-75333-6 .
- ^ «Илья Мечников — Биографическое» . www.nobelprize.org . Архивировано из оригинала 10 октября 2016 г. Проверено 10 октября 2016 г.
- ^ Jump up to: а б Марш М. (2001). Эндоцитоз . Издательство Оксфордского университета. п. VII. ISBN 978-0-19-963851-2 .
- ^ МакМахон Х.Т., Букро Э. (июль 2011 г.). «Молекулярный механизм и физиологические функции клатрин-опосредованного эндоцитоза». Обзоры природы. Молекулярно-клеточная биология . 12 (8): 517–33. дои : 10.1038/nrm3151 . ПМИД 21779028 . S2CID 15235357 .
- ^ Марш М., МакМахон Х.Т. (июль 1999 г.). «Структурная эра эндоцитоза». Наука . 285 (5425): 215–220. дои : 10.1126/science.285.5425.215 . ПМИД 10398591 .
- ^ Ираджизад Э., Валани Н., Витч С.Л., Лю А.П., Агравал А. (февраль 2017 г.). «Полимеризация клатрина проявляет высокую механо-геометрическую чувствительность» . Мягкая материя . 13 (7): 1455–1462. Бибкод : 2017SMat...13.1455I . дои : 10.1039/C6SM02623K . ПМК 5452080 . ПМИД 28124714 .
- ^ Партон Р.Г., Саймонс К. (март 2007 г.). «Множественные лица кавеол». Обзоры природы. Молекулярно-клеточная биология . 8 (3): 185–194. дои : 10.1038/nrm2122 . ПМИД 17318224 . S2CID 10830810 .
- ^ Минео С., Андерсон Р.Г. (август 2001 г.). «Потоцитоз. Лекция Роберта Фельгена». Гистохимия и клеточная биология . 116 (2): 109–118. дои : 10.1007/s004180100289 . ПМИД 11685539 .
- ^ Фальконе С., Кокуччи Э., Подини П., Кирххаузен Т., Клементи Э., Мелдолези Дж. (ноябрь 2006 г.). «Макропиноцитоз: регулируемая координация эндоцитарных и экзоцитарных мембранных событий». Журнал клеточной науки . 119 (Часть 22): 4758–4769. дои : 10.1242/jcs.03238 . ПМИД 17077125 . S2CID 14303429 .
- ^ Jump up to: а б Казаменто А, Букро Е (июнь 2020 г.). «Молекулярный механизм быстрого эндофилин-опосредованного эндоцитоза» . Биохимический журнал . 477 (12): 2327–2345. дои : 10.1042/bcj20190342 . ПМЦ 7319585 . ПМИД 32589750 .
- ^ Лундмарк Р., Доэрти Г.Дж., Хоуз М.Т., Кортезе К., Валлис Ю., Партон Р.Г., МакМахон Х.Т. (ноябрь 2008 г.). «Белок GRAF1, активирующий ГТФазу, регулирует эндоцитарный путь CLIC/GEEC» . Современная биология . 18 (22): 1802–1808. дои : 10.1016/j.cub.2008.10.044 . ПМЦ 2726289 . ПМИД 19036340 .
- ^ Меллман I (1996). «Эндоцитоз и молекулярная сортировка». Ежегодный обзор клеточной биологии и биологии развития . 12 : 575–625. дои : 10.1146/annurev.cellbio.12.1.575 . ПМИД 8970738 .
- ^ Мукерджи С., Гош Р.Н., Максфилд Ф.Р. (июль 1997 г.). «Эндоцитоз». Физиологические обзоры . 77 (3): 759–803. дои : 10.1152/physrev.1997.77.3.759 . ПМИД 9234965 .
- ^ Стурфогель В., Строус Г.Дж., Гез Х.Дж., Ооршот В., Шварц А.Л. (май 1991 г.). «Поздние эндосомы происходят из ранних эндосом в результате созревания». Клетка . 65 (3): 417–427. дои : 10.1016/0092-8674(91)90459-C . ПМИД 1850321 . S2CID 31539542 .
- ^ Вайсманн Г. (ноябрь 1965 г.). «Лизосома». Медицинский журнал Новой Англии . 273 (20): 1084–90 (продолжение). дои : 10.1056/NEJM196511112732006 . ПМИД 5319614 .
- ^ Грюнберг Дж. Максфилд Ф.Р. (август 1995 г.). «Мембранный транспорт по эндоцитарному пути». Современное мнение в области клеточной биологии . 7 (4): 552–563. дои : 10.1016/0955-0674(95)80013-1 . ПМИД 7495576 .
- ^ Луцио Дж.П., Роус Б.А., Брайт Н.А., Прайор П.Р., Маллок Б.М., Пайпер Р.К. (май 2000 г.). «Слияние лизосом-эндосом и биогенез лизосом» . Журнал клеточной науки . 113 (9): 1515–1524. дои : 10.1242/jcs.113.9.1515 . ПМИД 10751143 . [ постоянная мертвая ссылка ]
- ^ Вальтер Т.К., Брикнер Дж.Х., Агилар П.С., Берналес С., Пантоха С., Уолтер П. (февраль 2006 г.). «Эизосомы отмечают статические участки эндоцитоза». Природа . 439 (7079): 998–1003. Бибкод : 2006Natur.439..998W . дои : 10.1038/nature04472 . ПМИД 16496001 . S2CID 2838121 .
- ^ Кирххаузен Т., Оуэн Д., Харрисон С.С. (май 2014 г.). «Молекулярная структура, функции и динамика клатрин-опосредованного мембранного трафика» . Перспективы Колд-Спринг-Харбор в биологии . 6 (5): а016725. doi : 10.1101/cshperspect.a016725 . ПМЦ 3996469 . ПМИД 24789820 .
- ^ Бицикас В., Корреа И.Р., Николс Б.Дж. (сентябрь 2014 г.). «Клатрин-независимые пути не вносят существенного вклада в поток эндоцитов» . электронная жизнь . 3 : e03970. doi : 10.7554/eLife.03970 . ПМЦ 4185422 . ПМИД 25232658 .
- ^ Бенмера А., Ламаз С. (август 2007 г.). «Ямы, покрытые клатрином: vive la différence?». Трафик . 8 (8): 970–982. дои : 10.1111/j.1600-0854.2007.00585.x . ПМИД 17547704 . S2CID 12685926 .
- ^ Раппопорт Дж. З. (июнь 2008 г.). «Сосредоточение внимания на клатрин-опосредованном эндоцитозе». Биохимический журнал . 412 (3): 415–423. дои : 10.1042/BJ20080474 . ПМИД 18498251 . S2CID 24174632 .
- ^ Грансет Б., Одерматт Б., Ройл С.Дж., Лагнадо Л. (декабрь 2007 г.). «Клатрин-опосредованный эндоцитоз: физиологический механизм извлечения везикул в синапсах гиппокампа» . Журнал физиологии . 585 (Часть 3): 681–686. дои : 10.1113/jphysicalol.2007.139022 . ПМК 2375507 . ПМИД 17599959 .
- ^ Робинсон М.С. (март 1997 г.). «Пальто и почкование пузырьков». Тенденции в клеточной биологии . 7 (3): 99–102. дои : 10.1016/S0962-8924(96)10048-9 . ПМИД 17708916 .
- ^ Глик Б.С., Малхотра В. (декабрь 1998 г.). «Любопытный статус аппарата Гольджи» . Клетка . 95 (7): 883–889. дои : 10.1016/S0092-8674(00)81713-4 . ПМИД 9875843 .
- ^ Гайдаров И., Сантини Ф., Уоррен Р.А., Кин Дж.Х. (май 1999 г.). «Пространственный контроль динамики покрытых ямок в живых клетках». Природная клеточная биология . 1 (1): 1–7. дои : 10.1038/8971 . ПМИД 10559856 . S2CID 12553151 .
- ^ РОТ Т.Ф., ПОРТЕР КР (февраль 1964 г.). «Поглощение белка желтка в ооците комара Aedes Aegypti. L» . J Клеточная Биол . 20 (2): 313–32. дои : 10.1083/jcb.20.2.313 . ПМК 2106398 . PMID 14126875 .
- ^ Андерсон Р.Г., Браун М.С., Гольдштейн Дж.Л. (март 1977 г.). «Роль эндоцитарных пузырьков с покрытием в поглощении связанного с рецептором липопротеина низкой плотности в фибробластах человека». Клетка . 10 (3): 351–364. дои : 10.1016/0092-8674(77)90022-8 . ПМИД 191195 . S2CID 25657719 .
- ^ Пирс Б.М. (апрель 1976 г.). «Клатрин: уникальный белок, связанный с внутриклеточным переносом мембран с помощью везикул, покрытых оболочкой» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (4): 1255–1259. Бибкод : 1976PNAS...73.1255P . дои : 10.1073/pnas.73.4.1255 . ПМК 430241 . ПМИД 1063406 .
- ^ Партон Р.Г., Тиллу В.А., Коллинз Б.М. (апрель 2018 г.). «Кавеолы» . Современная биология . 28 (8): Р402–Р405. дои : 10.1016/j.cub.2017.11.075 . ПМИД 29689223 . S2CID 235331463 .
- ^ Кумари С., Мг С., мэр С. (март 2010 г.). «Эндоцитоз отключен: несколько способов проникновения в клетку» . Клеточные исследования . 20 (3): 256–75. дои : 10.1038/cr.2010.19 . ПМК 7091825 . ПМИД 20125123 .
- ^ Сарасидж Р.К., мэр С., Рао М. (май 2007 г.). «Вызванное хиральностью почкование: механизм эндоцитоза и морфологии кавеол, опосредованный рафтами?» . Биофизический журнал . 92 (9): 3140–58. Бибкод : 2007BpJ....92.3140S . дои : 10.1529/biophysj.106.085662 . ПМЦ 1852369 . ПМИД 17237196 .
Дальнейшее чтение
[ редактировать ]- Доэрти Дж.Дж., МакМахон Х.Т. (2009). «Механизмы эндоцитоза». Ежегодный обзор биохимии . 78 : 857–902. doi : 10.1146/annurev.biochem.78.081307.110540 . ПМИД 19317650 .