Синаптический везикула
| Синаптический везикула | |
|---|---|
 Нейрон А (передающий) к нейрону Б (принимающий). 1 . Митохондрия ; 2 . Синаптический везикула с нейротрансмиттерами ; 3 . Ауторецептор 4 . Синапс с высвобожденным нейротрансмиттером ( серотонином ); 5 . Постсинаптические рецепторы активируются нейромедиатором (индукция постсинаптического потенциала ); 6 . Кальциевый канал ; 7 . Экзоцитоз пузырька; 8 . Восстановленный нейромедиатор. | |
| Подробности | |
| Система | Нервная система |
| Идентификаторы | |
| латинский | синаптический везикула  |
| МеШ | Д013572 |
| ТД | Х2.00.06.2.00004 |
| Анатомические термины микроанатомии | |
В нейроне синаптические везикулы (или везикулы нейротрансмиттеров ) хранят различные нейротрансмиттеры , которые высвобождаются в синапсе . Высвобождение регулируется потенциалзависимым кальциевым каналом . Везикулы необходимы для распространения нервных импульсов между нейронами и постоянно воссоздаются клеткой . Область аксона , содержащая группы пузырьков, представляет собой окончание аксона или «терминальный бутон». За один бутон можно высвободить до 130 везикул за десятиминутный период стимуляции с частотой 0,2 Гц. [1] В зрительной коре головного мозга человека синаптические пузырьки имеют средний диаметр 39,5 нанометров (нм) со стандартным отклонением 5,1 нм. [2]
Структура [ править ]
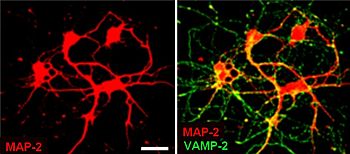
Синаптические везикулы относительно просты, поскольку лишь ограниченное количество белков умещается в сфере диаметром 40 нм. Очищенные везикулы имеют соотношение белок : фосфолипид 1:3, а липидный состав состоит из 40% фосфатидилхолина , 32% фосфатидилэтаноламина , 12% фосфатидилсерина , 5% фосфатидилинозитола и 10% холестерина . [4]
Синаптические везикулы содержат два класса обязательных компонентов: транспортные белки, участвующие в захвате нейротрансмиттеров, и транспортные белки, которые участвуют в экзоцитозе , эндоцитозе и рециклировании синаптических везикул.
- Транспортные белки состоят из протонных насосов , которые генерируют электрохимические градиенты , обеспечивающие захват нейротрансмиттеров, и транспортеров нейротрансмиттеров, которые регулируют фактическое поглощение нейротрансмиттеров. Необходимый протонный градиент создается V-АТФазой , которая расщепляет АТФ для получения энергии. Везикулярные транспортеры перемещают нейротрансмиттеры из цитоплазмы клеток в синаптические пузырьки. Везикулярные переносчики глутамата , например, посредством этого процесса изолируют глутамат в везикулы.
- Транспортировка белков более сложна. Они включают внутренние мембранные белки , периферически связанные белки и такие белки, как SNARE . Эти белки не имеют общих характеристик, которые позволили бы их идентифицировать как белки синаптических пузырьков, и мало что известно о том, как эти белки специфически откладываются в синаптических пузырьках. Многие, но не все известные белки синаптических везикул взаимодействуют с невезикулярными белками и связаны со специфическими функциями. [4]
Стехиометрия движения различных нейротрансмиттеров в везикулу представлена в следующей таблице.
| Тип(ы) нейротрансмиттера | Внутреннее движение | Движение наружу |
|---|---|---|
| норадреналин , дофамин , гистамин , серотонин и ацетилхолин. | нейромедиатор + | 2 часа + |
| ГАМК и глицин | нейромедиатор | 1 час + |
| глутамат | нейромедиатор − + Cl − | 1 час + |
Недавно было обнаружено, что синаптические везикулы также содержат небольшие молекулы РНК, включая фрагменты транспортной РНК , фрагменты Y-РНК и мирРНК . [5] Считается, что это открытие окажет большое влияние на изучение химических синапсов.
нейротоксинов Эффекты
некоторые нейротоксины , такие как батрахотоксин Известно, что , разрушают синаптические пузырьки. Столбнячный ( токсин повреждает мембранные белки, ассоциированные с везикулами VAMP), тип v-SNARE , тогда как ботулинические токсины повреждают t-SNARE S и v-SNARES и, таким образом, ингибируют синаптическую передачу. [6] Токсин паука , называемый альфа-латротоксином, связывается с нейрексинами , повреждая везикулы и вызывая массивное высвобождение нейротрансмиттеров. [ нужна ссылка ]
Пулы везикул [ править ]
Везикулы в нервном окончании сгруппированы в три пула: легковысвобождаемый пул, рециклирующий пул и резервный пул. [7] Эти бассейны различаются по своей функции и положению в нервном окончании. Легко высвобождаемые пулы прикрепляются к клеточной мембране , что делает их первой группой везикул, высвобождаемых при стимуляции. Легковысвобождаемый пул невелик и быстро исчерпывается. Пул рециркуляции находится вблизи клеточной мембраны и имеет тенденцию циклически повторяться при умеренной стимуляции, так что скорость высвобождения везикул равна или ниже скорости образования везикул. Этот пул больше, чем легковысвобождаемый пул, но для его мобилизации требуется больше времени. Резервный пул содержит везикулы, которые не выделяются в нормальных условиях. Этот резервный пул может быть довольно большим (~50%) в нейронах, выращенных на стеклянной подложке, но очень мал или отсутствует в зрелых синапсах интактной ткани мозга. [8] [9]
Физиология [ править ]
Цикл синаптических пузырьков [ править ]
События цикла синаптических пузырьков можно разделить на несколько ключевых этапов: [10]
- 1. Трафик в синапс
Компоненты синаптических пузырьков в пресинаптическом нейроне первоначально передаются в синапс с помощью членов семейства кинезиновых моторов. У C. elegans основным мотором синаптических везикул является UNC-104. [11] Есть также свидетельства того, что другие белки, такие как UNC-16/Sunday Driver, регулируют использование моторов для транспорта синаптических везикул. [12]
- 2. Загрузка передатчика
Попадая в синапс, синаптические пузырьки загружаются нейротрансмиттером. Загрузка трансмиттера — это активный процесс, требующий транспортера нейромедиатора и АТФазы протонной помпы, которая обеспечивает электрохимический градиент. Эти транспортеры селективны для разных классов передатчиков. характеристики unc-17 и unc-47, которые кодируют везикулярный переносчик ацетилхолина и везикулярный переносчик ГАМК . На сегодняшний день описаны [13]
- 3. Стыковка
Загруженные синаптические везикулы должны стыковаться вблизи мест высвобождения, однако стыковка — это этап цикла, о котором мы мало что знаем. Было идентифицировано множество белков на синаптических везикулах и в местах высвобождения, однако ни одно из идентифицированных белковых взаимодействий между белками везикул и белками мест высвобождения не может объяснить фазу стыковки цикла. Мутанты rab-3 и munc-18 изменяют стыковку везикул или организацию везикул в местах высвобождения, но они не нарушают полностью стыковку. [14] Белки SNARE теперь, по-видимому, также участвуют в этапе стыковки цикла. [15]
- 4. Грунтовка
После того как синаптические пузырьки первоначально стыкуются, их необходимо подготовить, прежде чем они смогут начать слияние. Прайминг подготавливает синаптические пузырьки к тому, чтобы они могли быстро сливаться в ответ на приток кальция. Считается, что этот этап прайминга включает образование частично собранных комплексов SNARE. В этом событии участвуют белки Munc13 , RIM и RIM-BP. [16] Считается, что Munc13 стимулирует изменение синтаксина t-SNARE из закрытой конформации в открытую, что стимулирует сборку комплексов v-SNARE/t-SNARE. [17] RIM также, по-видимому, регулирует прайминг, но не является обязательным для этого этапа. [ нужна ссылка ]
- 5. Фьюжн
Примированные везикулы очень быстро сливаются с клеточной мембраной в ответ на повышение уровня кальция в цитоплазме. При этом накопленный нейромедиатор высвобождается в синаптическую щель . Считается, что событие термоядерного синтеза происходит непосредственно через SNARE и управляется энергией, обеспечиваемой сборкой SNARE. Триггером этого события, чувствительным к кальцию, является белок синаптотагмин, связывающий кальций. Способность SNAREs опосредовать слияние кальций-зависимым образом недавно была восстановлена in vitro. В соответствии с тем, что SNARE необходимы для процесса слияния, мутанты v-SNARE и t-SNARE C. elegans смертельны. Сходным образом, мутанты у Drosophila и нокауты у мышей указывают на то, что эти SNARES играют критическую роль в синаптическом экзоцитозе. [10]
- 6. Эндоцитоз
Это объясняет повторный захват синаптических везикул в модели полного контактного слияния. Однако в других исследованиях собраны данные, свидетельствующие о том, что этот тип слияния и эндоцитоза не всегда имеет место. [ нужна ссылка ]
Переработка везикул [ править ]
Считается, что за рециркуляцию синаптических пузырьков ответственны два ведущих механизма действия: полное слияние коллапса и метод «поцелуй и беги». Оба механизма начинаются с образования синаптической поры, которая высвобождает медиатор во внеклеточное пространство. После высвобождения нейротрансмиттера пора может либо полностью расшириться, так что везикула полностью схлопывается в синаптическую мембрану, либо она может быстро закрыться и оторвать мембрану, вызывая слияние по принципу «поцелуй и беги». [18]
Полный коллапс [ править ]
Было показано, что периоды интенсивной стимуляции нервных синапсов истощают количество пузырьков, а также увеличивают клеточную емкость и площадь поверхности. [19] Это указывает на то, что после того, как синаптические везикулы высвобождают свою полезную нагрузку нейромедиатора, они сливаются с клеточной мембраной и становятся ее частью. Пометив синаптические везикулы с помощью HRP ( пероксидазы хрена ), Хойзер и Риз обнаружили, что части клеточной мембраны в нервно-мышечном соединении лягушки поглощались клеткой и превращались обратно в синаптические везикулы. [20] Исследования показывают, что весь цикл экзоцитоза, извлечения и реформирования синаптических везикул занимает менее 1 минуты. [21]
При полном слиянии синаптический везикула сливается и включается в клеточную мембрану. Формирование новой мембраны является белково-опосредованным процессом и может происходить только при определенных условиях. После потенциала действия Ca 2+ проникает в пресинаптическую мембрану. Калифорния 2+ связывается со специфическими белками цитоплазмы, одним из которых является синаптотагмин , который, в свою очередь, запускает полное слияние синаптического пузырька с клеточной мембраной. Полному слиянию пор способствуют белки SNARE . Это большое семейство белков обеспечивает стыковку синаптических везикул АТФ-зависимым образом. С помощью синаптобревина на синаптическом пузырьке комплекс t-SNARE на мембране, состоящий из синтаксина и SNAP-25 , может стыковать, примировать и сливать синаптический пузырь с мембраной. [22]
Было показано, что механизм полного коллапса слияния является мишенью ботулинического и столбнячного токсинов . Ботулинический токсин обладает протеазной активностью, которая разрушает белок SNAP-25 . Белок SNAP-25 необходим для слияния пузырьков, которые высвобождают нейротрансмиттеры, в частности ацетилхолин. [23] Ботулинический токсин по существу расщепляет эти белки SNARE и при этом предотвращает слияние синаптических везикул с клеточной синаптической мембраной и высвобождение их нейротрансмиттеров. Столбнячный токсин действует по аналогичному пути, но вместо этого атакует белок синаптобревин на синаптическом пузырьке. В свою очередь, эти нейротоксины препятствуют полному слиянию синаптических везикул. Без этого механизма могут возникнуть мышечные спазмы, паралич и смерть. [ нужна ссылка ]
«Поцелуй и беги» [ править ]
Второй механизм повторного использования синаптических пузырьков известен как слияние «поцелуй и беги» . В этом случае синаптическая везикула «целует» клеточную мембрану, открывая небольшую пору для высвобождения полезной нагрузки нейромедиатора, затем закрывает пору и возвращается обратно в клетку. [18] Механизм «поцелуй и беги» стал горячо обсуждаемой темой. Его эффекты наблюдались и записывались; однако причина его использования в отличие от полного слияния с коллапсом все еще исследуется. Было высказано предположение, что «поцелуй и беги» часто используется для сохранения дефицитных везикулярных ресурсов, а также для ответа на высокочастотные сигналы. [24] Эксперименты показали, что события «поцелуй и беги» действительно случаются. Впервые наблюдавшийся Кацем и дель Кастильо, позже было замечено, что механизм «поцелуй и беги» отличается от полного слияния коллапса тем, что клеточная емкость не увеличивается в событиях «поцелуй и беги». [24] Это подтверждает идею о том, что синаптический везикула высвобождает свою полезную нагрузку, а затем отделяется от мембраны.
Модуляция [ править ]
Таким образом, клетки, по-видимому, имеют по крайней мере два механизма рециркуляции мембран. При определенных условиях клетки могут переключаться с одного механизма на другой. Медленное, обычное, полное слияние преобладает над синаптической мембраной, когда Ca 2+ уровни низкие, и механизм быстрого поцелуя и беги применяется, когда Ca 2+ уровни высокие. [ нужна ссылка ]
Алесь и др. показали, что повышенные концентрации внеклеточных ионов кальция смещают предпочтительный способ рециркуляции и высвобождения синаптических везикул на механизм «поцелуй и беги» в зависимости от концентрации кальция. Было высказано предположение, что во время секреции нейромедиаторов в синапсах режим экзоцитоза модулируется кальцием для достижения оптимальных условий для сопряженного экзоцитоза и эндоцитоза в соответствии с синаптической активностью. [25]
Экспериментальные данные свидетельствуют о том, что «поцелуй и беги» является доминирующим способом синаптической высвобождения в начале серии стимулов. В этом контексте «поцелуй и беги» отражает высокую вероятность высвобождения везикул. Частота «поцелуй и беги» также увеличивается из-за быстрого возбуждения и стимуляции нейрона, что позволяет предположить, что кинетика этого типа высвобождения быстрее, чем у других форм высвобождения везикул. [26]
История [ править ]
С появлением электронного микроскопа в начале 1950-х годов было обнаружено, что нервные окончания содержат большое количество электронно-прозрачных (прозрачных для электронов) пузырьков. [27] [28] Термин «синаптический везикула» впервые был введен Де Робертисом и Беннеттом в 1954 году. [29] лягушки Это произошло вскоре после того, как было обнаружено, что высвобождение медиатора в нервно-мышечном соединении индуцирует постсинаптические миниатюрные потенциалы концевой пластинки , которые были приписаны высвобождению дискретных пакетов нейромедиаторов (квантов) из пресинаптического нервного окончания. [30] [31] Таким образом, было разумно предположить, что в таких везикулах содержится вещество-передатчик ( ацетилхолин ), которое по секреторному механизму высвобождает свое содержимое в синаптическую щель (гипотеза везикул). [32] [33]
Недостающим звеном была демонстрация того, что нейромедиатор ацетилхолин на самом деле содержится в синаптических везикулах. Примерно десять лет спустя применение методов субклеточного фракционирования к тканям мозга позволило выделить сначала нервные окончания ( синаптосомы ), [34] и впоследствии синаптических везикул головного мозга млекопитающих. В этой работе были задействованы две конкурирующие лаборатории: лаборатория Виктора П. Уиттакера в Институте физиологии животных Совета сельскохозяйственных исследований, Бабрахам, Кембридж, Великобритания, и лаборатория Эдуардо де Робертиса в Институте общей анатомии и эмбриологии медицинского факультета. Университет Буэнос-Айреса, Аргентина. [35] Работа Уиттекера, демонстрирующая ацетилхолин во фракциях везикул головного мозга морской свинки , была впервые опубликована в абстрактной форме в 1960 году, а затем более подробно в 1963 и 1964 годах. [36] [37] а статья группы де Робертиса, демонстрирующая обогащение связанного ацетилхолина во фракциях синаптических пузырьков мозга крыс, появилась в 1963 году. [38] Обе группы высвободили синаптические везикулы из изолированных синаптосом посредством осмотического шока . Первоначально предполагалось, что содержание ацетилхолина в везикуле составляет 1000–2000 молекул. [39] Последующие работы выявили везикулярную локализацию других нейротрансмиттеров, таких как аминокислоты , катехоламины , серотонин и АТФ . Позже синаптические везикулы также удалось выделить из других тканей, таких как верхний шейный ганглий , [40] или мозг осьминога . [41] Выделение высокоочищенной фракции холинергических синаптических везикул из лучевой торпеды . электрического органа [42] [43] стало важным шагом вперед в изучении биохимии и функции везикул.
См. также [ править ]
Ссылки [ править ]
- ^ Икеда, К; Беккерс, Дж. М. (2009). «Подсчет количества высвобождаемых синаптических пузырьков в пресинаптическом терминале» . Proc Natl Acad Sci США . 106 (8): 2945–50. Бибкод : 2009PNAS..106.2945I . дои : 10.1073/pnas.0811017106 . ПМЦ 2650301 . ПМИД 19202060 .
- ^ Цюй, Лей; Акбергенова Юлия; Ху, Юньмин; Шикорски, Томас (март 2009 г.). «Межсинаптические изменения среднего размера синаптических пузырьков и его связь с синаптической морфологией и функцией» . Журнал сравнительной неврологии . 514 (4): 343–352. дои : 10.1002/cne.22007 . ПМИД 19330815 . S2CID 23965024 . Архивировано из оригинала 5 января 2013 г.
- ^ Тонна, Ноэми; Бьянко, Фабио; Маттеоли, Микела; Каньоли, Чинция; Антонуччи, Флавия; Манфреди, Амедея; Мауро, Николо; Рануччи, Элизабетта; Феррути, Паоло (2014). «Растворимый биосовместимый гуанидин-содержащий полиамидоамин в качестве промотора первичной адгезии клеток головного мозга и культивирования клеток in vitro» . Наука и технология перспективных материалов . 15 (4): 045007. Бибкод : 2014STAdM..15d5007T . дои : 10.1088/1468-6996/15/4/045007 . ПМК 5090696 . ПМИД 27877708 .
- ^ Jump up to: Перейти обратно: а б Бенфенати, Ф.; Грингард, П.; Бруннер, Дж.; Бэлер, М. (1989). «Электростатические и гидрофобные взаимодействия синапсин I и фрагментов синапсин I с фосфолипидными бислоями» . Журнал клеточной биологии . 108 (5): 1851–1862. дои : 10.1083/jcb.108.5.1851 . ПМК 2115549 . ПМИД 2497105 .
- ^ Ли, Хуэйнань; Ву, Ченг; Арамайо, Родольфо; Сакс, Мэтью С.; Харлоу, Марк Л. (08 октября 2015 г.). «Синаптические везикулы содержат небольшие рибонуклеиновые кислоты (мРНК), включая фрагменты транспортной РНК (trfRNA) и микроРНК (миРНК)» . Научные отчеты . 5 : 14918. Бибкод : 2015NatSR...514918L . дои : 10.1038/srep14918 . ПМЦ 4597359 . ПМИД 26446566 .
- ^ Кандель Э.Р., Шварц Дж.Х., Джесселл Т.М., ред. (2000). «Выпуск передатчика». Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. ISBN 978-0-8385-7701-1 .
- ^ Риццоли, Сильвио О; Бетц, Уильям Дж. (январь 2005 г.). «Синаптические пулы пузырьков». Обзоры природы Неврология . 6 (1): 57–69. дои : 10.1038/nrn1583 . ПМИД 15611727 . S2CID 7473893 .
- ^ Роуз, Тобиас; Шёненбергер, Филипп; Езек, Карел; Ортнер, Томас Г. (2013). «Уточнение развития цикла везикул в коллатеральных синапсах Шаффера» . Нейрон . 77 (6): 1109–1121. дои : 10.1016/j.neuron.2013.01.021 . ПМИД 23522046 .
- ^ Сюэ, Лэй; Шэн, Цзяньсун; У, Синь-Шэн; Ву, Вэй; Ло, Фуцзюнь; Шин, Вончуль; Чан, Сюэ-Чэн; Ву, Лин-Ганг (15 мая 2013 г.). «Большинство везикул в центральных нервных окончаниях участвуют в переработке» . Журнал неврологии . 33 (20): 8820–8826. doi : 10.1523/jneurosci.4029-12.2013 . ПМЦ 3710729 . ПМИД 23678124 .
- ^ Jump up to: Перейти обратно: а б Зюдхоф, ТК (2004). «Цикл синаптических пузырьков». Ежегодный обзор неврологии . 27 : 509–547. дои : 10.1146/annurev.neuro.26.041002.131412 . ПМИД 15217342 . S2CID 917924 .
- ^ Тьен, северо-запад; Ву, Г.Х.; Сюй, CC; Чанг, CY; Вагнер, О.И. (2011). «Tau/PTL-1 связывается с кинезином-3 KIF1A/UNC-104 и влияет на двигательные характеристики нейронов C. Elegans». Нейробиология болезней . 43 (2): 495–506. дои : 10.1016/j.nbd.2011.04.023 . ПМИД 21569846 . S2CID 9712304 .
- ^ Аримото, М.; Кошика, ИП; Чоудхари, Британская Колумбия; Ли, К.; Мацумото, К.; Хисамото, Н. (2011). «Белок JIP3 Caenorhabditis elegans UNC-16 действует как адаптер, связывающий кинезин-1 с цитоплазматическим динеином» . Журнал неврологии . 31 (6): 2216–2224. doi : 10.1523/JNEUROSCI.2653-10.2011 . ПМК 6633058 . ПМИД 21307258 .
- ^ Сандовал, генеральный менеджер; Дюрр, Дж. С.; Ходжкин, Дж.; Рэнд, Дж.Б.; Рувкун, Г. (2006). «Генетическое взаимодействие между везикулярным переносчиком ацетилхолина VAChT/UNC-17 и синаптобревином/SNB-1 у C. Elegans». Природная неврология . 9 (5): 599–601. дои : 10.1038/nn1685 . ПМИД 16604067 . S2CID 11812089 .
- ^ Авраам, К.; Залог.; Леубе, Р.Э. (2011). «Синаптогирин-зависимая модуляция синаптической нейротрансмиссии у Caenorhabditis elegans». Нейронаука . 190 : 75–88. doi : 10.1016/j.neuroscience.2011.05.069 . ПМИД 21689733 . S2CID 14547322 .
- ^ Хаммарлунд, Марк; Палфриман, Марк Т; Ватанабэ, Сигеки; Олсен, Шон; Йоргенсен, Эрик М. (август 2007 г.). «Открытые синтаксиновые доки синаптических везикул» . ПЛОС Биология . 5 (8): е198. doi : 10.1371/journal.pbio.0050198 . ISSN 1544-9173 . ПМК 1914072 . ПМИД 17645391 .
- ^ Кезер, Паскаль С.; Дэн, Лунбинь; Ван, Юн; Дулубова Ирина; Лю, Синьрань; Ризо, Хосеп; Зюдхоф, Томас К. (2011). «Белки RIM связывают каналы Ca2+ с пресинаптическими активными зонами посредством прямого взаимодействия PDZ-домена» . Клетка . 144 (2): 282–295. дои : 10.1016/j.cell.2010.12.029 . ПМК 3063406 . ПМИД 21241895 .
- ^ Лин, XG; Мин, М.; Чен, MR; Ню, В.П.; Чжан, Ю.Д.; Лю, Б.; Цзю, Ю.М.; Ю, Дж.В.; Сюй, Т.; Ву, ZX (2010). «UNC-31/CAPS стыкуется и праймирует везикулы с плотным ядром в нейронах C. Elegans». Связь с биохимическими и биофизическими исследованиями . 397 (3): 526–531. дои : 10.1016/j.bbrc.2010.05.148 . ПМИД 20515653 .
- ^ Jump up to: Перейти обратно: а б Брекенридж, LJ; Алмерс, В. (1987). «Ток через пору слияния, которая образуется во время экзоцитоза секреторного пузырька». Природа . 328 (6133): 814–817. Бибкод : 1987Natur.328..814B . дои : 10.1038/328814a0 . ПМИД 2442614 . S2CID 4255296 .
- ^ Хойзер, Дж. Э.; Риз, Т.С. (1973). «Доказательства повторного использования мембраны синаптических пузырьков во время высвобождения передатчика в нервно-мышечном соединении лягушки» . Журнал клеточной биологии . 57 (2): 315–344. дои : 10.1083/jcb.57.2.315 . ПМК 2108984 . ПМИД 4348786 .
- ^ Миллер, ТМ; Хойзер, Дж. Э. (1984). «Эндоцитоз мембраны синаптического пузырька в нервно-мышечном соединении лягушки» . Журнал клеточной биологии . 98 (2): 685–698. дои : 10.1083/jcb.98.2.685 . ПМК 2113115 . ПМИД 6607255 .
- ^ Райан, штат Калифорния; Смит, С.Дж.; Рейтер, Х. (1996). «Время эндоцитоза синаптических пузырьков» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (11): 5567–5571. Бибкод : 1996PNAS...93.5567R . дои : 10.1073/pnas.93.11.5567 . ПМК 39287 . ПМИД 8643616 .
- ^ Сюй, Х.; Зик, М.; Викнер, WT; Джун, Ю. (2011). «Закрепленный на липидах SNARE поддерживает слияние мембран» . Труды Национальной академии наук . 108 (42): 17325–17330. Бибкод : 2011PNAS..10817325X . дои : 10.1073/pnas.1113888108 . ПМК 3198343 . ПМИД 21987819 .
- ^ Форан, П.Г.; Мохаммед, Н.; Лиск, ГО; Нагвани, С.; Лоуренс, GW; Джонсон, Э.; Смит, Л.; Аоки, КР; Долли, Джо (2002). «Оценка терапевтической полезности ботулинического нейротоксина B, C1, E и F по сравнению с длительно действующим типом A. ОСНОВА ДЛЯ РАЗЛИЧНОЙ ПРОДОЛЖИТЕЛЬНОСТИ ИНГИБИРОВАНИЯ ЭКЗОЦИТОЗА В ЦЕНТРАЛЬНЫХ НЕЙРОНАХ» . Журнал биологической химии . 278 (2): 1363–1371. дои : 10.1074/jbc.M209821200 . ПМИД 12381720 .
- ^ Jump up to: Перейти обратно: а б Харата, Северная Каролина; Араванис, AM; Цянь, RW (2006). «Поцелуй и беги и полный коллапс слияния как способы экзоэндоцитоза при нейросекреции». Журнал нейрохимии . 97 (6): 1546–1570. дои : 10.1111/j.1471-4159.2006.03987.x . ПМИД 16805768 . S2CID 36749378 .
- ^ Альварес Де Толедо, Г.; Алес, Э.; Табарес, Луизиана; Пойато, Дж. М.; Валеро, В.; Линдау, М. (1999). «Высокие концентрации кальция переводят режим экзоцитоза на механизм «поцелуй и беги». Природная клеточная биология . 1 (1): 40–44. дои : 10.1038/9012 . ПМИД 10559862 . S2CID 17624473 .
- ^ Чжан, К.; Ли, Ю.; Цянь, RW (2009). «Динамический контроль процессов «поцелуй и беги» и повторного использования везикул, исследованных с помощью одиночных наночастиц» . Наука . 323 (5920): 1448–1453. Бибкод : 2009Sci...323.1448Z . дои : 10.1126/science.1167373 . ПМК 2696197 . ПМИД 19213879 .
- ^ Палай, Сэнфорд Л.; Паладе, Джордж Э. (1954). «Электронно-микроскопическое исследование цитоплазмы нейронов». Анатомическая запись (устное изложение). 118 : 336. дои : 10.1002/ar.1091180211 .
- ^ Эдуардо ДП, Де Робертис; Стэнли, Беннетт, Х. (25 января 1955 г.). «Некоторые особенности субмикроскопической морфологии синапсов лягушки и дождевого червя» . Журнал биофизической и биохимической цитологии . 1 (1): 47–58. дои : 10.1083/jcb.1.1.47 . JSTOR 1602913 . ПМЦ 2223594 . ПМИД 14381427 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Де Робертис EDP, Беннетт Х.С. (1954). «Субмикроскопический везикулярный компонент в синапсе». Протокол ФРС . 13:35 .
- ^ Фатт, П.; Кац, Б. (7 октября 1950 г.). «Некоторые наблюдения о биологическом шуме». Природа . 166 (4223): 597–598. Бибкод : 1950Natur.166..597F . дои : 10.1038/166597a0 . ПМИД 14780165 . S2CID 9117892 .
- ^ Фатт, П.; Кац, Б. (28 мая 1952 г.). «Спонтанная подпороговая активность двигательных нервных окончаний» (PDF) . Журнал физиологии . 117 (1): 109–128. дои : 10.1113/jphysicalol.1952.sp004735 . ПМЦ 1392564 . ПМИД 14946732 . Архивировано из оригинала (PDF) 2 февраля 2014 года . Проверено 1 февраля 2014 г.
- ^ Дель Кастильо Дж. Б., Кац Б. (1954). «Квантовые компоненты потенциала концевой пластинки» . Дж. Физиол . 124 (3): 560–573. дои : 10.1113/jphysicalol.1954.sp005129 . ПМЦ 1366292 . ПМИД 13175199 .
- ^ Дель Кастильо Дж. Б., Кац Б. (1954). «Биофизические аспекты нервно-мышечной передачи». Прога Биофиз Биофиз Хим . 6 : 121–170. ПМИД 13420190 .
- ^ Грей Э.Г., вице-президент Уиттакера (1962). «Выделение нервных окончаний из головного мозга: электронно-микроскопическое исследование фрагментов клеток, полученных в результате гомогенизации и центрифугирования» . Дж Анат . 96 (Часть 1): 79–88. ПМЦ 1244174 . ПМИД 13901297 .
- ^ Циммерманн, Герберт (2018). «Открытие синаптосомы и ее последствия». Нейрометоды . 141 : 9–26. дои : 10.1007/978-1-4939-8739-9_2 .
- ^ Уиттакер В.П., Майклсон И.А., Киркланд Р.Дж. (1963). «Отделение синаптических везикул от разрушенных частиц нервных окончаний». Биохим Фармакол . 12 (2): 300–302. дои : 10.1016/0006-2952(63)90156-4 . ПМИД 14000416 .
- ^ Уиттакер В.П., Майклсон И.А., Киркланд Р.Дж. (1964). «Отделение синаптических везикул от частиц нервных окончаний («синаптосом»)» . Биохим Дж . 90 (2): 293–303. дои : 10.1042/bj0900293 . ПМК 1202615 . ПМИД 5834239 .
- ^ Де Робертис Э., Родригес де Лорес Арнаис Г., Салгаников Г.Л., Пеллегрино де Иральди А., Зихер Л.М. (1963). «Выделение синаптических везикул и структурная организация ацетилхолиновой системы в нервных окончаниях головного мозга». Дж. Нейрохем . 10 (4): 225–235. дои : 10.1111/j.1471-4159.1963.tb05038.x . ПМИД 14026026 . S2CID 33266876 .
- ^ Уиттакер, вице-президент, Шеридан, Миннесота (1965). «Морфология и содержание ацетилхолина в изолированных синаптических пузырьках коры головного мозга». Дж. Нейрохем . 12 (5): 363–372. дои : 10.1111/j.1471-4159.1965.tb04237.x . ПМИД 14333293 . S2CID 5746357 .
- ^ Уилсон В.С., Шульц Р.А., Купер-младший (1973). «Выделение холинергических синаптических пузырьков из верхнего шейного ганглия крупного рогатого скота и оценка содержания в них ацетилхолина». Дж. Нейрохем . 20 (3): 659–667. дои : 10.1111/j.1471-4159.1973.tb00026.x . ПМИД 4574192 . S2CID 6157415 .
- ^ Джонс Д.Г. (1970). «Выделение синаптических пузырьков из мозга осьминога». Мозговой Рес . 17 (2): 181–193. дои : 10.1016/0006-8993(70)90077-6 . ПМИД 5412681 .
- ^ Исраэль М., Гаутрон Дж., Лесбатс Б. (1970). «Субклеточное фракционирование электрического органа Torpedo marmorata ». Дж. Нейрохем . 17 (10): 1441–1450. дои : 10.1111/j.1471-4159.1970.tb00511.x . ПМИД 5471906 . S2CID 8087195 .
- ^ Уиттакер, вице-президент, Эссман В.Б., Доу Г.Х. (1972). «Выделение чистых холинергических синаптических везикул из электрических органов пластиножаберных рыб семейства Torpidinidae» . Биохим Дж . 128 (4): 833–846. дои : 10.1042/bj1280833 . ПМЦ 1173903 . ПМИД 4638794 .