Вкусовой рецептор
| Вкусовой рецептор | |
|---|---|
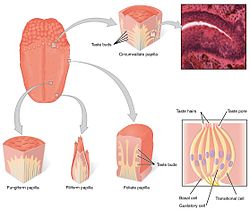 Вкусовые рецепторы языка присутствуют во вкусовых почках сосочков. | |
| Идентификаторы | |
| ФМА | 84662 |
| Анатомическая терминология | |
Рецептор вкуса или дегустатор — это тип клеточного рецептора , который облегчает ощущение вкуса . Когда пища или другие вещества попадают в рот, молекулы взаимодействуют со слюной и связываются с вкусовыми рецепторами в полости рта и других местах. Молекулы, дающие ощущение вкуса, считаются «сырыми». [1]
Вкусовые рецепторы позвоночных делятся на два семейства: [ нужна ссылка ]
- Тип 1, сладкий , впервые охарактеризован в 2001 году: [2] ТАС1Р2 – ТАС1Р3
- Тип 2, горький , впервые охарактеризован в 2000 году: [3] У человека известно 25 различных рецепторов горечи, у кошек — 12, у кур — три, а у мышей — 35 различных рецепторов горечи. [4]
Зрительный, обонятельный, «сапиктивный» (восприятие вкусов), тройничный (горячий, прохладный), механический — все они способствуют восприятию вкуса . Из них ваниллоидные рецепторы члена 1 подсемейства V катионного канала временного рецепторного потенциала ( TRPV1 ) ответственны за восприятие тепла от некоторых молекул, таких как капсаицин, а рецептор CMR1 отвечает за восприятие холода от таких молекул, как ментол , эвкалиптол , и ицилин . [1]
Распределение в тканях [ править ]
Вкусовая система состоит из вкусовых рецепторных клеток во вкусовых почках . Вкусовые рецепторы, в свою очередь, содержатся в структурах, называемых сосочками . Существует три типа сосочков, участвующих в формировании вкуса: грибовидные сосочки , листовидные сосочки и округлые сосочки . (Четвертый тип – нитевидные сосочки не содержат вкусовых почек). Помимо сосочков, вкусовые рецепторы также находятся на нёбе и в ранних частях пищеварительной системы, таких как гортань и верхний отдел пищевода . Есть три черепно-мозговых нерва , иннервирующих язык; блуждающий нерв , языкоглоточный нерв и лицевой нерв . Языкоглоточный нерв и барабанная струна лицевого нерва иннервируют вкусовые рецепторы TAS1R и TAS2R. Помимо вкусовых рецепторов на языке, эпителий кишечника также оснащен тонкой хемосенсорной системой, которая передает сенсорную информацию нескольким эффекторным системам, участвующим в регуляции аппетита, иммунных реакций и моторики желудочно-кишечного тракта. [5]
В 2010 году исследователи обнаружили в легочной ткани рецепторы горечи, которые заставляют дыхательные пути расслабляться при попадании горького вещества. Они полагают, что этот механизм является эволюционно адаптивным, поскольку помогает избавиться от легочных инфекций, но его также можно использовать для лечения астмы и хронической обструктивной болезни легких . [6]
Рецептор сладкого вкуса (T1R2/T1R3) можно найти в различных внеротовых органах человеческого тела, таких как мозг, сердце, почки, мочевой пузырь, респираторный эпителий носа и другие. В большинстве органов функция рецепторов неясна. [7] Было обнаружено, что рецептор сладкого вкуса, обнаруженный в кишечнике и поджелудочной железе, играет важную роль в метаболической регуляции процесса восприятия углеводов в кишечнике и секреции инсулина. [8] Этот рецептор также обнаружен в мочевом пузыре, что позволяет предположить, что потребление искусственных подсластителей, активирующих этот рецептор, может вызвать чрезмерное сокращение мочевого пузыря. [9]
Функция [ править ]
Вкус помогает идентифицировать токсины , поддерживать питание и регулировать аппетит, иммунные реакции и перистальтику желудочно-кишечного тракта. [5] Сегодня признаны пять основных вкусов: соленый, сладкий, горький, кислый и умами . Ощущения соленого и кислого вкуса обнаруживаются через ионные каналы . Однако сладкий, горький вкус и вкус умами обнаруживаются с помощью вкусовых рецепторов , связанных с G-белком . [10]
Кроме того, некоторые агенты могут действовать как модификаторы вкуса , например, миракулин или куркулин для сладкого или стерубин для маскировки горького вкуса .
Механизм действия [ править ]
Стандартный рецептор горького, сладкого вкуса или вкуса умами представляет собой связанный с G-белком рецептор с семью трансмембранными доменами . Связывание лиганда вкусовых рецепторов активирует каскады вторичных мессенджеров, деполяризуя вкусовую клетку. Густдуцин является наиболее распространенной вкусовой субъединицей Gα, играющей важную роль в восприятии горького вкуса TAS2R. Густдуцин является гомологом трансдуцина , G-белка, участвующего в зрительной трансдукции. [11] Кроме того, вкусовые рецепторы используют ионный канал TRPM5, а также фосфолипазу PLCβ2. [12]
Пикантные или глутаматы (умами) [ править ]
Гетеродимерный рецептор TAS1R1 аминокислот + TAS1R3 функционирует как рецептор умами, реагируя на связывание L- , особенно L- глутамата . [2] Вкус умами чаще всего связан с пищевой добавкой глутамата натрия (MSG) и может быть усилен за счет связывания молекул инозинмонофосфата (IMP) и гуанозинмонофосфата (GMP). [13] [14] Клетки, экспрессирующие TAS1R1+3, обнаруживаются в основном в грибовидных сосочках на кончике и краях языка, а также в клетках вкусовых рецепторов неба на нёбе. [2] Показано, что эти клетки образуют синапсы на нервах барабанной хорды, чтобы посылать сигналы в мозг, хотя некоторая активация языкоглоточного нерва . была обнаружена [13] [15]
Альтернативные кандидаты на вкусовые рецепторы умами включают сплайсинговые варианты метаботропных рецепторов глутамата, mGluR4 и mGluR1 , а также рецептор NMDA . [10] [16] [17] [18]
В ходе эволюции певчих птиц вкусовой рецептор умами претерпел структурные изменения в месте связывания лиганда, что позволило этим птицам ощущать сладкий вкус с помощью этого рецептора. [19]
Сладкий [ править ]
Гетеродимерный рецептор TAS1R2 , + TAS1R3 действует как рецептор сладкого связываясь с широким спектром сахаров и заменителей сахара . [2] [20] Клетки, экспрессирующие TAS1R2+3, обнаруживаются в околоваловидных и листовидных сосочках вблизи задней части языка , а также в клетках небных вкусовых рецепторов на нёбе. [2] Показано, что эти клетки образуют синапсы на барабанной хорде и языкоглоточных нервах, чтобы посылать сигналы в мозг. [10] [15] Годимер TAS1R3 также функционирует как рецептор сладкого, почти так же, как TAS1R2+3, но имеет пониженную чувствительность к сладким веществам. Природные сахара легче обнаруживаются рецептором TAS1R3, чем заменители сахара . Это может помочь объяснить, почему сахар и искусственные подсластители имеют разные вкусы. [21] Генетический полиморфизм TAS1R3 частично объясняет разницу в восприятии сладкого вкуса и потреблении сахара между людьми афроамериканского происхождения и людьми европейского и азиатского происхождения. [22] [23]
Ощущение сладкого вкуса менялось на протяжении эволюции разных животных. Млекопитающие ощущают сладкий вкус, передавая сигнал через гетеродимер T1R2/T1R3, рецептор сладкого вкуса. Однако у птиц мономер T1R2 не существует, и они ощущают сладкий вкус посредством гетеродимера T1R1/T1R3, вкусового рецептора умами, который претерпел модификации в ходе своей эволюции. Недавно проведенное исследование показало, что на этапах эволюции певчих птиц наблюдалось снижение способности ощущать вкус умами и увеличение способности ощущать сладкий вкус, тогда как изначальные родители певчих птиц могли ощущать только вкус умами. Исследователи обнаружили, что возможным объяснением этого явления могут быть структурные изменения в месте связывания лиганда рецептора умами у певчих птиц, чувствующих сладкий вкус, и не чувствующих их. Предполагается, что со временем произошла мутация в сайте связывания, что позволило им ощутить сладкий вкус через вкусовой рецептор умами. [19]
Горький [ править ]
Белки TAS2R ( InterPro : IPR007960 ) действуют как рецепторы горького вкуса. [24] Существует 43 гена TAS2R человека , каждый из которых (за исключением пяти псевдогенов ) не имеет интронов и кодирует белок GPCR . [10] Эти белки, в отличие от белков TAS1R, имеют короткие внеклеточные домены и расположены в околовалятчатых сосочках , нёбе , листовидных сосочках и вкусовых сосочках надгортанника , с пониженной экспрессией в грибовидных сосочках . [3] [10] Хотя несомненно, что в одной клетке вкусового рецептора экспрессируется несколько TAS2R, до сих пор ведутся споры о том, могут ли млекопитающие различать вкусы разных горьких лигандов . [3] [10] Однако некоторое совпадение должно иметь место, поскольку горьких соединений гораздо больше, чем генов TAS2R. Обычные горькие лиганды включают циклогексимид , денатоний , ПРОП ( 6- н- пропил-2-тиоурацил ), ПТК ( фенилтиокарбамид ) и β- глюкопиранозиды . [10]
Передача сигнала горьких стимулов осуществляется через α-субъединицу густдуцина . Эта субъединица G-белка активирует вкусовую фосфодиэстеразу и снижает уровень циклических нуклеотидов . Дальнейшие этапы пути трансдукции пока неизвестны. βγ-субъединица густдуцина также опосредует вкус, активируя IP 3 ( инозитолтрифосфат ) и DAG ( диглицерид ). Эти вторичные мессенджеры могут открывать закрытые ионные каналы или вызывать высвобождение внутреннего кальция . [25] Хотя все TAS2R расположены в клетках, содержащих густдуцин, нокаут густдуцина не устраняет полностью чувствительность к горьким соединениям, что указывает на избыточный механизм возникновения горького вкуса. [12] (неудивительно, учитывая, что горький вкус обычно сигнализирует о присутствии токсина ) . [12] Одним из предложенных механизмов независимого от густдуцина горького вкуса является взаимодействие ионных каналов со специфическими горькими лигандами, аналогично взаимодействию ионных каналов, которое происходит при ощущении вкуса кислых и соленых стимулов. [10]
Одним из наиболее изученных белков TAS2R является TAS2R38 , который способствует ощущению вкуса как PROP, так и PTC. Это первый вкусовой рецептор, полиморфизм которого , как показано, отвечает за различия в восприятии вкуса. Текущие исследования сосредоточены на определении других подобных полиморфизмов, определяющих вкусовой фенотип. [10] Более поздние исследования показывают, что генетический полиморфизм в других генах рецепторов горького вкуса влияет на восприятие горького вкуса кофеина, хинина и денатония бензоата. [26]
Этот раздел может потребовать очистки Википедии , чтобы соответствовать стандартам качества . Конкретная проблема: необходимо проверить тон и форматирование в следующем абзаце. ( сентябрь 2022 г. ) |
Показано, что рецепторы горечи (TAS2R) играют важную роль во врожденной иммунной системе дыхательных путей ( носа и пазух ) тканей мерцательного эпителия . [27] Эта врожденная иммунная система добавляет «активную крепость». [28] к физическому поверхностному барьеру иммунной системы .Эта фиксированная иммунная система активируется путем связывания лигандов со специфическими рецепторами.Эти природные лиганды являются бактериальными маркерами, например TAS2R38 : лактоны ацил-гомосерина. [29] или хинолоны [30] вырабатывается Pseudomonas aeruginosa . Чтобы защититься от хищников, некоторые растения производят вещества, имитирующие бактериальные маркеры. Эти растительные мимы интерпретируются языком и мозгом как горечь .Фиксированные рецепторы иммунной системы идентичны рецепторам горького вкуса TAS2R. Вещества горечи являются агонистами фиксированной иммунной системы TAS2R. [27] Врожденная иммунная система использует оксид азота и дефенсины , которые способны уничтожать бактерии, а также вирусы. [31] [32] Эти фиксированные врожденные иммунные системы (Активные крепости) известны и в других эпителиальных тканях, помимо верхних дыхательных путей ( нос , пазухи , трахея , бронхи ), например: грудь (эпителиальные клетки молочной железы), кишечник человека , а также кожа (кератиноциты). [33] Молекулы горького вкуса, связанные с ними рецепторы горького вкуса, а также последовательности и модели гомологии рецепторов горького вкуса доступны через BitterDB .
Кислый [ править ]
Исторически считалось, что кислый вкус возникает исключительно тогда, когда свободные ионы водорода ( H + ) непосредственно деполяризующие вкусовые рецепторы. Однако в настоящее время предлагаются специфические рецепторы кислого вкуса с другими способами действия. Каналы HCN выступили с таким предложением; поскольку они представляют собой каналы, управляемые циклическими нуклеотидами. Два ионных канала, которые, как предполагается, способствуют кислому вкусу, — это ASIC2 и TASK-1.
Соль [ править ]
Также были предложены различные рецепторы для соленого вкуса, а также возможное вкусовое обнаружение липидов, сложных углеводов и воды. Доказательства существования этих рецепторов в большинстве исследований на млекопитающих были неубедительными. Например, можно показать, что предлагаемый рецептор ENaC для обнаружения натрия способствует вкусу натрия только у дрозофилы . [10] Однако было показано, что протеолизированные формы ENaC действуют как рецепторы солевого вкуса человека. Протеолиз – это процесс расщепления белка. Считается, что зрелая форма ENaC протеолизирована, однако характеристика того, какие протеолизированные формы существуют в каких тканях, неполная. Протеолиз клеток, созданных для сверхэкспрессии гетермулитмерного ENaC, содержащего альфа-, бета- и гамма-субъединицы, использовали для идентификации соединений, которые избирательно усиливают активность протеолизированного ENaC по сравнению с непротеолизированным ENaC. Сенсорные исследования человека показали, что соединение, которое усиливает протеолизированный ENaC, усиливает соленый вкус поваренной соли или хлорида натрия, что подтверждает, что протеолизированный ENaC является первым рецептором солевого вкуса человека. [34]
Карбонизация [ править ]
Фермент, связанный с кислым рецептором, передает информацию о газированной воде. [35]
Жир [ править ]
возможный вкусовой рецептор жира, CD36 . Идентифицирован [36] CD36 локализован в околовалятных и листовидных сосочках , которые присутствуют во вкусовых сосочках. [37] и где производится лингвальная липаза , и исследования показали, что рецептор CD36 связывает длинноцепочечные жирные кислоты . [38] Различия в уровне экспрессии CD36 у людей были связаны с их способностью ощущать вкус жиров. [39] создавая аргументы в пользу связи рецептора с дегустацией жира. Дальнейшие исследования рецептора CD36 могут быть полезны для определения существования настоящего рецептора, определяющего вкус жира.
Рецептор свободных жирных кислот 4 (также называемый GPR120) и, в гораздо меньшей степени, рецептор свободных жирных кислот 1 (также называемый GPR40) [40] были вовлечены в реакцию на жир в полости рта, [41] а их отсутствие приводит к снижению предпочтения жиров и снижению реакции нейронов на перорально вводимые жирные кислоты. [42]
TRPM5 участвует в реакции жира в полости рта и идентифицирован как возможный рецептор орального жира, но недавние данные показывают, что он в первую очередь является вторичным действующим лицом. Было показано, что [43] [44]
Типы [ править ]
Гены рецепторов горького вкуса человека называются от TAS2R1 до TAS2R64, со многими пробелами из-за несуществующих генов, псевдогенов или предполагаемых генов, которые не были аннотированы в самой последней сборке генома человека. Многие гены рецепторов горького вкуса также имеют запутанные синонимические названия, поскольку несколько разных названий генов относятся к одному и тому же гену. Полный список генов рецепторов горького вкуса человека см. в таблице ниже:
| Сорт | Ген | Синонимы | Псевдонимы | Локус | Описание |
|---|---|---|---|---|---|
| тип 1 (сладкий) | ТАС1Р1 | GPR70 | 1п36.23 | ||
| ТАС1Р2 | ГПР71 | 1п36.23 | |||
| ТАС1Р3 | 1п36 | ||||
| тип 2 (горький) | ТАС2Р1 | 5п15 | |||
| ТАС2Р2 | 7p21.3 | псевдоген | |||
| ТАС2Р3 | 7q31.3-q32 | ||||
| ТАС2Р4 | 7q31.3-q32 | ||||
| ТАС2Р5 | 7q31.3-q32 | ||||
| ТАС2Р6 | 7 | не аннотирован в сборке генома человека | |||
| ТАС2Р7 | 12p13 | ||||
| ТАС2Р8 | 12p13 | ||||
| ТАС2Р9 | 12p13 | ||||
| ТАС2Р10 | 12p13 | ||||
| ТАС2Р11 | отсутствует у человека | ||||
| ТАС2Р12 | ТАС2Р26 | 12p13.2 | псевдоген | ||
| ТАС2Р13 | 12p13 | ||||
| ТАС2Р14 | 12p13 | ||||
| ТАС2Р15 | 12p13.2 | псевдоген | |||
| ТАС2Р16 | 7q31.1-q31.3 | ||||
| ТАС2Р17 | отсутствует у человека | ||||
| ТАС2Р18 | 12p13.2 | псевдоген | |||
| ТАС2Р19 | ТАС2Р23, ТАС2Р48 | 12p13.2 | |||
| ТАС2Р20 | ТАС2Р49 | 12p13.2 | |||
| ТАС2Р21 | отсутствует у человека | ||||
| ТАС2Р22 | 12 | не аннотирован в сборке генома человека | |||
| ТАС2Р24 | отсутствует у человека | ||||
| ТАС2Р25 | отсутствует у человека | ||||
| ТАС2Р27 | отсутствует у человека | ||||
| ТАС2Р28 | отсутствует у человека | ||||
| ТАС2Р29 | отсутствует у человека | ||||
| ТАС2Р30 | ТАС2Р47 | 12p13.2 | |||
| ТАС2Р31 | ТАС2Р44 | 12p13.2 | |||
| ТАС2Р32 | отсутствует у человека | ||||
| ТАС2Р33 | 12 | не аннотирован в сборке генома человека | |||
| ТАС2Р34 | отсутствует у человека | ||||
| ТАС2Р35 | отсутствует у человека | ||||
| ТАС2Р36 | 12 | не аннотирован в сборке генома человека | |||
| ТАС2Р37 | 12 | не аннотирован в сборке генома человека | |||
| ТАС2Р38 | 7q34 | ||||
| ТАС2Р39 | 7q34 | ||||
| ТАС2Р40 | ГПР60 | 7q34 | |||
| ТАС2Р41 | 7q34 | ||||
| ТАС2Р42 | 12p13 | ||||
| ТАС2Р43 | 12p13.2 | ||||
| ТАС2Р45 | ГПР59 | 12 | |||
| ТАС2Р46 | 12p13.2 | ||||
| ТАС2Р50 | ТАС2Р51 | 12p13.2 | |||
| ТАС2Р52 | отсутствует у человека | ||||
| ТАС2Р53 | отсутствует у человека | ||||
| ТАС2Р54 | отсутствует у человека | ||||
| ТАС2Р55 | отсутствует у человека | ||||
| ТАС2Р56 | отсутствует у человека | ||||
| ТАС2Р57 | отсутствует у человека | ||||
| ТАС2Р58 | отсутствует у человека | ||||
| ТАС2Р59 | отсутствует у человека | ||||
| ТАС2Р60 | 7 | ||||
| ТАС2Р62П | 7q34 | псевдоген | |||
| ТАС2Р63П | 12p13.2 | псевдоген | |||
| ТАС2Р64П | 12p13.2 | псевдоген |
Потеря функции [ править ]
У многих видов вкусовые рецепторы потеряли функции. Считается, что эволюционный процесс, в ходе которого вкусовые рецепторы утратили свою функцию, является адаптивной эволюцией, связанной с экологией питания, которая приводит к специализации и раздвоению вкусовых рецепторов. [45] Показано, что из всех вкусовых рецепторов горький, сладкий и умами имеют корреляцию между инактивацией вкусовых рецепторов и пищевым поведением. [45] Однако нет убедительных доказательств того, что у каких-либо позвоночных отсутствуют гены рецептора горького вкуса. [45]
Рецептор сладкого вкуса — один из вкусовых рецепторов, функция которого утрачена. У млекопитающих преобладающим рецептором сладкого вкуса является вкусовой рецептор 1-го типа Tas1r2/Tas1r3. [46] Некоторые виды млекопитающих, такие как кошки и летучие мыши-вампиры, продемонстрировали неспособность ощущать сладкий вкус. [46] У этих видов причина потери функции рецептора сладкого связана с псевдогенизацией Tas1r2. [46] Псевдогенизация Tas1r2 также наблюдается у видов, не относящихся к млекопитающим, таких как куры и безъязыкая западная шпорцевая лягушка, и эти виды также демонстрируют неспособность ощущать сладкий вкус. [46] Псевдогенизация Tas1r2 широко распространена и независима у отряда Carnivora. [46] Многие исследования показали, что псевдогенизация вкусовых рецепторов вызвана вредной мутацией в открытых рамках считывания (ORF). [47] В ходе исследования было обнаружено, что у плотоядных видов, не относящихся к кошачьим, эти виды обнаруживают мутации Tas1r2, нарушающие ORF, и они происходят независимо среди видов. [46] Они также показали высокую вариативность в своих родословных. [46] Предполагается, что псевдогенизация Tas1r2 произошла в результате конвергентной эволюции, когда хищные виды утратили способность ощущать сладкий вкус из-за пищевого поведения. [46]
Умами также является вкусовым рецептором, функция которого у многих видов утрачена. Преобладающими вкусовыми рецепторами умами являются Tas1r1/Tas1r3. [46] Было обнаружено, что у двух линий водных млекопитающих, включая дельфинов и морских львов, Tas1r1 псевдогенизирован. [46] Псевдогенизация Tas1r1 также была обнаружена у наземных хищных видов. [46] Хотя панда принадлежит к отряду хищных, она травоядна, 99% ее рациона составляет бамбук, и она не может попробовать умами. [48] Последовательность генома панды показывает, что ее ген Tas1r1 псевдогенизирован. [48] В ходе исследования было обнаружено, что у всех видов отряда Carnivora, кроме панды, открытая рамка считывания . сохранялась [48] Было обнаружено, что у панд соотношение несинонимических и синонимичных замен намного выше, чем у других видов отряда Carnivora. [48] Эти данные коррелируют с датой окаменелостей панды и показывают, когда панда перешла от плотоядной диеты к травоядной. [46] Таким образом, предполагается, что потеря функции умами у панды вызвана изменением диеты, когда панда стала меньше зависеть от мяса. [46] Однако эти исследования не объясняют существование травоядных животных, таких как лошади и коровы, которые сохранили рецептор Tas1r1. [48]
В целом, потеря функции вкусового рецептора — это эволюционный процесс, произошедший из-за изменения в питании видов. [47]
См. также [ править ]
Список различных типов клеток в организме взрослого человека
Ссылки [ править ]
- ^ Перейти обратно: а б Это, Эрве (2012). «Наука о духовке. Отрывок из главы 1» . Архивировано из оригинала 2 мая 2014 года . Проверено 30 апреля 2014 г.
- ^ Перейти обратно: а б с д и Нельсон Г., Хун М.А., Чандрашекар Дж. и др. (август 2001 г.). «Рецепторы сладкого вкуса млекопитающих» . Клетка . 106 (3): 381–90. дои : 10.1016/S0092-8674(01)00451-2 . ПМИД 11509186 . S2CID 11886074 .
- ^ Перейти обратно: а б с Адлер Э., Хун М.А., Мюллер К.Л. и др. (март 2000 г.). «Новое семейство вкусовых рецепторов млекопитающих» . Клетка . 100 (6): 693–702. дои : 10.1016/S0092-8674(00)80705-9 . ПМИД 10761934 . S2CID 14604586 .
- ^ «Обзор рецепторов» . Горькая БД .
- ^ Перейти обратно: а б Стинселс С., Депортере I (2018). «Хеморецепторы в кишечнике». Ежегодный обзор физиологии . 80 : 117–141. doi : 10.1146/annurev-физиология-021317-121332 . ПМИД 29029594 .
- ^ Дешпанде Д.А., Ван В.К., Макилмойл Э.Л. и др. (ноябрь 2010 г.). «Рецепторы горького вкуса на гладких мышцах дыхательных путей бронходилатируют за счет локализованной передачи сигналов кальция и обратной обструкции» . Природная медицина . 16 (11): 1299–304. дои : 10.1038/нм.2237 . ПМК 3066567 . ПМИД 20972434 .
- ^ Лаффит, Анни; Нейерс, Фабрис; Бриан, Лоик (июль 2014 г.). «Функциональная роль рецептора сладкого вкуса в тканях полости рта и вне полости рта» . Текущее мнение о клиническом питании и метаболической помощи . 17 (4): 379–385. doi : 10.1097/mco.0000000000000058 . ISSN 1363-1950 . ПМК 4059820 . ПМИД 24763065 .
- ^ Энкен, Ж.-К. (21 августа 2012 г.). «Вкусят ли клетки поджелудочной железы питательные вещества, секретирующие инсулин?» . Научная сигнализация . 5 (239): пе36. дои : 10.1126/scisignal.2003325 . ISSN 1945-0877 . ПМИД 22932700 . S2CID 206671643 .
- ^ Эллиотт, Рут А.; Капур, Шалини; Тинчелло, Дуглас Г. (декабрь 2011 г.). «Экспрессия и распределение изоформ рецепторов сладкого вкуса T1R2 и T1R3 в мочевом пузыре человека и крысы» . Журнал урологии . 186 (6): 2455–2462. дои : 10.1016/j.juro.2011.07.083 . HDL : 2381/10362 . ISSN 0022-5347 . ПМИД 22019168 .
- ^ Перейти обратно: а б с д и ж г час я дж Бахманов А.А., Бошан ГК (2007). «Гены вкусовых рецепторов» . Ежегодный обзор питания . 27 : 389–414. дои : 10.1146/annurev.nutr.26.061505.111329 . ПМЦ 2721271 . ПМИД 17444812 .
- ^ Сайнс Э., Кавена М.М., ЛопесХименес Н.Д. и др. (июнь 2007 г.). «Свойства G-белка, связывающие человеческие рецепторы сладкого и аминокислотного вкуса». Развивающая нейробиология . 67 (7): 948–59. дои : 10.1002/dneu.20403 . ПМИД 17506496 . S2CID 29736077 .
- ^ Перейти обратно: а б с Чжан Ю., Хун М.А., Чандрашекар Дж. и др. (февраль 2003 г.). «Кодирование сладкого, горького вкуса и вкуса умами: разные рецепторные клетки имеют схожие сигнальные пути» . Клетка . 112 (3): 293–301. дои : 10.1016/S0092-8674(03)00071-0 . ПМИД 12581520 . S2CID 718601 .
- ^ Перейти обратно: а б Нельсон Г., Чандрашекар Дж., Хун М.А. и др. (март 2002 г.). «Аминокислотный вкусовой рецептор». Природа . 416 (6877): 199–202. Бибкод : 2002Natur.416..199N . дои : 10.1038/nature726 . ПМИД 11894099 . S2CID 1730089 .
- ^ Задержка ER, Бивер А.Дж., Вагнер К.А. и др. (октябрь 2000 г.). «Синергия вкусовых предпочтений между агонистами глутаматных рецепторов и инозинмонофосфатом у крыс» . Химические чувства . 25 (5): 507–15. дои : 10.1093/chemse/25.5.507 . ПМИД 11015322 .
- ^ Перейти обратно: а б Данилова В, Хеллекант Г (март 2003 г.). «Сравнение ответов барабанной хорды и языкоглоточного нервов на вкусовые стимулы у мышей C57BL/6J» . BMC Нейронаука . 4 :5. дои : 10.1186/1471-2202-4-5 . ПМК 153500 . ПМИД 12617752 .
- ^ Бренд JG (апрель 2000 г.). «Рецепторы и процессы трансдукции вкуса умами» . Журнал питания . 130 (дополнение 4S): 942S–5S. дои : 10.1093/jn/130.4.942S . ПМИД 10736357 .
- ^ Чаудхари Н., Ландин А.М., Ропер С.Д. (февраль 2000 г.). «Вариант метаботропного рецептора глутамата действует как вкусовой рецептор». Природная неврология . 3 (2): 113–9. дои : 10.1038/72053 . ПМИД 10649565 . S2CID 16650588 .
- ^ Тойоно Т., Сета Ю., Катаока С. и др. (июль 2003 г.). «Экспрессия группы I метаботропных глутаматных рецепторов во вкусовых сосочках крыс». Исследования клеток и тканей . 313 (1): 29–35. дои : 10.1007/s00441-003-0740-2 . ПМИД 12898387 . S2CID 41546387 .
- ^ Перейти обратно: а б Тода, Ясука; Ко, Мэн-Чинг; Лян, Цяойи; и др. (08.07.2021). «Раннее возникновение сладкого восприятия в излучении певчих птиц» . Наука . 373 (6551): 226–231. Бибкод : 2021Sci...373..226T . дои : 10.1126/science.abf6505 . ISSN 0036-8075 . ПМИД 34244416 . S2CID 235769720 .
- ^ Ли Х, Сташевски Л, Сюй Х и др. (апрель 2002 г.). «Человеческие рецепторы вкуса сладкого и умами» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (7): 4692–6. Бибкод : 2002PNAS...99.4692L . дои : 10.1073/pnas.072090199 . ПМЦ 123709 . ПМИД 11917125 .
- ^ Чжао GQ, Чжан Ю, Хун М.А. и др. (октябрь 2003 г.). «Рецепторы сладкого вкуса и вкуса умами млекопитающих» . Клетка . 115 (3): 255–66. дои : 10.1016/S0092-8674(03)00844-4 . ПМИД 14636554 . S2CID 11773362 .
- ^ Хван Л.Д., Лин С., Гараххани П. и др. (апрель 2019 г.). «Новое понимание сладкого вкуса человека: полногеномное исследование взаимосвязей восприятия и потребления сладких веществ» . Американский журнал клинического питания . 109 (6): 1724–1737. дои : 10.1093/ajcn/nqz043 . ПМК 6537940 . ПМИД 31005972 .
- ^ Юсиф Р. (март 2020 г.). «Изучение молекулярных взаимодействий между неокулином и рецепторами сладкого вкуса человека с помощью вычислительных подходов» (PDF) . Сайнс Малайзия . 49 (3): 517–525. дои : 10.17576/jsm-2020-4903-06 .
- ^ Чандрашекар Дж., Мюллер К.Л., Хун М.А. и др. (март 2000 г.). «T2R функционируют как рецепторы горького вкуса» . Клетка . 100 (6): 703–11. дои : 10.1016/S0092-8674(00)80706-0 . ПМИД 10761935 . S2CID 7293493 .
- ^ Маргольский РФ (январь 2002 г.). «Молекулярные механизмы преобразования горького и сладкого вкуса» . Журнал биологической химии . 277 (1): 1–4. дои : 10.1074/jbc.R100054200 . ПМИД 11696554 .
- ^ Хван Л.Д., Гараххани П., Бреслин П.А. и др. (сентябрь 2018 г.). «Двумерный общегеномный анализ ассоциаций усиливает роль кластеров рецепторов горького на хромосомах 7 и 12 в формировании горького вкуса человека» . БМК Геномика . 19 (1): 678. doi : 10.1186/s12864-018-5058-2 . ПМК 6142396 . ПМИД 30223776 .
- ^ Перейти обратно: а б Ли Р.Дж., Коэн, Северная Каролина (2015). «Вкусовые рецепторы при врожденном иммунитете» . Клетка. Мол. Наука о жизни . 72 (2): 217–236. дои : 10.1007/s00018-014-1736-7 . ПМЦ 4286424 . ПМИД 25323130 .
- ^ «Фиксированная иммунная система, TAS2R» (на французском языке). 4 сентября 2020 г.
- ^ Ли Р.Дж., Коэн, Северная Каролина (2015). «Роль рецептора горького вкуса T2R38 при инфекциях верхних дыхательных путей и хроническом риносинусите» . Современное мнение в области аллергии и клинической иммунологии . 15 (1): 14–20. doi : 10.1097/ACI.0000000000000120 . ПМК 5902169 . ПМИД 25304231 .
- ^ Фрейнд Дж., Мэнсфилд СиДжей, Дограмджи ЭлДжей и Ли Р.Дж. (2018). «Активация эпителиальных рецепторов горького вкуса дыхательных путей хинолонами Pseudomonas aeruginosa модулирует передачу сигналов кальция, циклического АМФ и оксида азота» . Журнал биологической химии . 293 (25): 9824–9840. дои : 10.1074/jbc.RA117.001005 . ПМК 6016477 . ПМИД 29748385 .
- ^ Клотман М.Э., Чанг Т.Л. (июнь 2006 г.). «Дефенсины во врожденном противовирусном иммунитете». Обзоры природы. Иммунология . 6 (6): 447–56. дои : 10.1038/nri1860 . ПМИД 16724099 . S2CID 8603587 .
- ^ Кроен К. (1993). «Доказательства противовирусного действия оксида азота. Ингибирование репликации вируса простого герпеса 1 типа» . Журнал клинических исследований . 91 (6): 2446–2452. дои : 10.1172/JCI116479 . ПМК 443304 . ПМИД 8390481 .
- ^ Вулфле Ю, Эльсхольц Ф (2015). «Экспрессия и функциональная активность рецепторов горького вкуса TAS2R1 и TAS2R38 в кератиноцитах человека». Кожная фармакология и физиология . 28 (3): 137–146. дои : 10.1159/000367631 . ПМИД 25573083 . S2CID 19912094 .
- ^ Шекдар К., Лангер Дж., Венкатачалан С. и др. (март 2021 г.). «Метод клеточной инженерии с использованием сигнальных зондов флюорогенных олигонуклеотидов и проточной цитометрии» . Письма о биотехнологиях . 43 (5): 949–958. дои : 10.1007/s10529-021-03101-5 . ISSN 0141-5492 . ПМЦ 7937778 . ПМИД 33683511 .
- ^ «Ученые обнаружили белковый рецептор карбонизированного вкуса, пресс-релиз от 15 октября 2009 г. – Национальные институты здравоохранения (NIH)» . Архивировано из оригинала 3 июля 2015 г. Проверено 6 октября 2014 г.
- ^ Логеретт Ф., Пассильи-Деграс П., Патрис Б. и др. (ноябрь 2005 г.). «Участие CD36 в оросенсорном обнаружении пищевых липидов, спонтанном предпочтении жиров и пищеварительных секрециях» . Журнал клинических исследований . 115 (11): 3177–84. дои : 10.1172/JCI25299 . ПМЦ 1265871 . ПМИД 16276419 .
- ^ Саймонс П.Дж., Куммер Дж.А., Люйкен Дж.Дж., Бун Л. (декабрь 2011 г.). «Апикальная иммунолокализация CD36 во вкусовых сосочках человека и свиньи из окружных и листовидных сосочков». Акта гистохимика . 113 (8): 839–43. дои : 10.1016/j.acthis.2010.08.006 . ПМИД 20950842 .
- ^ Бэйли А.Г., Коберн, Коннектикут, Абумрад Н.А. (сентябрь 1996 г.). «Обратимое связывание длинноцепочечных жирных кислот с очищенным жиром, гомологом CD36 жировой ткани». Журнал мембранной биологии . 153 (1): 75–81. дои : 10.1007/s002329900111 . ПМИД 8694909 . S2CID 5911289 .
- ^ Пепино М.Ю., Лав-Грегори Л., Кляйн С., Абумрад Н.А. (март 2012 г.). «Ген транслоказы жирных кислот CD36 и лингвальная липаза влияют на пероральную чувствительность к жиру у людей с ожирением» . Журнал исследований липидов . 53 (3): 561–6. дои : 10.1194/jlr.M021873 . ПМК 3276480 . ПМИД 22210925 .
- ^ Кимура И., Ичимура А., Оуэ-Китано Р., Игараси М. (январь 2020 г.). «Рецепторы свободных жирных кислот в здоровье и болезнях» . Физиологические обзоры . 100 (1): 171–210. doi : 10.1152/physrev.00041.2018 . ПМИД 31487233 .
- ^ ДиПатрицио Н.В. (сентябрь 2014 г.). «Готов ли жирный вкус к прайм-тайму?» . Физиология и поведение . 136 : 145–54. дои : 10.1016/j.physbeh.2014.03.002 . ПМК 4162865 . ПМИД 24631296 .
- ^ Картони С., Ясумацу К., Окури Т. и др. (июнь 2010 г.). «Вкусовые предпочтения жирных кислот опосредуются GPR40 и GPR120» . Журнал неврологии . 30 (25): 8376–82. doi : 10.1523/JNEUROSCI.0496-10.2010 . ПМК 6634626 . ПМИД 20573884 .
- ^ Мэттес Р.Д. (сентябрь 2011 г.). «Накопление данных подтверждает наличие вкусового компонента свободных жирных кислот у людей» . Физиология и поведение . 104 (4): 624–31. дои : 10.1016/j.physbeh.2011.05.002 . ПМК 3139746 . ПМИД 21557960 .
- ^ Лю П., Шах Б.П., Кроасделл С., Гилбертсон Т.А. (июнь 2011 г.). «Переходный потенциальный рецепторный канал типа М5 необходим для определения вкуса жира» . Журнал неврологии . 31 (23): 8634–42. doi : 10.1523/JNEUROSCI.6273-10.2011 . ПМК 3125678 . ПМИД 21653867 .
- ^ Перейти обратно: а б с Фэн П., Чжао Х. (июнь 2013 г.). «Сложная эволюционная история генов рецепторов вкуса сладкого/умами у позвоночных» . Китайский научный бюллетень . 58 (18): 2198–2204. Бибкод : 2013ЧСБу..58.2198Ф . дои : 10.1007/s11434-013-5811-5 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Цзян П., Хосуэ Дж., Ли Икс и др. (март 2012 г.). «Сильная потеря вкуса у плотоядных млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (13): 4956–61. дои : 10.1073/pnas.1118360109 . ПМК 3324019 . ПМИД 22411809 .
- ^ Перейти обратно: а б Антинуччи М., Риссо Д. (28 ноября 2017 г.). «Вопрос вкуса: специфичная для линии потеря функции генов вкусовых рецепторов у позвоночных» . Границы молекулярной биологии . 4:81 . doi : 10.3389/fmolb.2017.00081 . ПМЦ 5712339 . ПМИД 29234667 .
- ^ Перейти обратно: а б с д и Чжао Х., Ян-младший, Сюй Х., Чжан Дж. (декабрь 2010 г.). «Псевдогенизация гена вкусового рецептора умами Tas1r1 у гигантской панды совпала с ее переходом на бамбук» . Молекулярная биология и эволюция . 27 (12): 2669–73. дои : 10.1093/molbev/msq153 . ПМК 3108379 . ПМИД 20573776 .
Внешние ссылки [ править ]
- Адлер Э., Хун М.А., Мюллер К.Л. и др. (2000). «Новое семейство вкусовых рецепторов млекопитающих - обзор исследования» . Факультет биологии колледжа Дэвидсона . Проверено 11 августа 2008 г.
- вкус+рецепторы,+тип+1 в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- вкус + рецепторы, + тип + 2 в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)