HR
| HR | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | RRH , гомолог родопсина, происходящий из пигментного эпителия сетчатки. | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 605224 ; МГИ : 1097709 ; Гомологен : 55977 ; GeneCards : RRH ; ОМА : RRH – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Перопсин , зрительный пигментоподобный рецептор, представляет собой белок , который у человека кодируется RRH геном . [5] [6] Как и другие опсины животных, он принадлежит к рецепторам, связанным с G-белком . [6] Несмотря на это, первые перопсины были обнаружены у мышей и людей уже в 1997 году. [5] о них мало что известно. [7]
Фотохимия
[ редактировать ]Как и большинство опсинов, перопсины имеют в своем седьмом трансмембранном домене лизин , соответствующий положению аминокислоты 296 в крупного рогатого скота родопсине . [5] [7] что важно для связывания сетчатки и светочувствительности. [8]
У амфиоксуса , головохордового , перопсин связывается в темном состоянии полностью с транс- ретиналем вместо 11-цис-ретиналя, [9] как и в родопсине крупного рогатого скота. [10] [11] [12] [13] [14] Поэтому было предложено считать перопсины фотоизомеразами. [9]
Тканевая локализация
[ редактировать ]У мышей перопсин локализуется в апикальных микроворсинках ( пигментного эпителия сетчатки ПЭС). [5] Там он регулирует хранение или перемещение витамина А из сетчатки в РПЭ. [15] Перопсин также экспрессируется в кератиноцитах человека кожи . В культуре клеток кератиноцитов он реагирует на УФ-свет, если в него поступает ретиналь. [16] У кур перопсин экспрессируется вместе с RGR-опсином в шишковидной железе и сетчатке. [17]
Локализация и структура гена
[ редактировать ]Ген перопсина человека расположен на участке 4q25 хромосомы 4 и имеет шесть интронов. [6] [18] как RGR-опсины. Однако только два из этих интронов встраиваются в одно и то же место, что все же указывает на то, что перопсины и RGR-опсины более тесно связаны друг с другом, чем с реснитчатыми и рабдомерными опсинами. [18] Эта общая структура генов также отражена в филогении опсинов, где перопсины и RGR-опсины находятся в одной группе: хромопсины. [18] [7] [19] [20]
Филогения
[ редактировать ]Перопсины встречаются только в краниатах и головохордовых. [7] Краниаты — это таксон , в который входят млекопитающие , а вместе с ними и люди. Перопсины — одна из семи подгрупп хромопсинов. Другие группы — это RGR-опсины , ретинохромы , немопсины, астропсины, варропсины и глюопсины. [7] Хромопсины являются одной из трех подгрупп тетраопсинов (также известных как RGR/Go или опсины группы 4). Другие группы — это нейропсины и гоопсины. Тетраопсины — одна из пяти основных групп животных опсинов , также известных как опсины типа 2). Другие группы — это цилиарные опсины (c-опсины, цилопсины), рабдомерные опсины (r-опсины, рабопсины), ксенопсины и нессопсины. Четыре из этих субкладов встречаются в Bilateria (все, кроме нессопсинов). [7] [19] Однако двулатеральные клады представляют собой парафилетический таксон без опсинов книдарий . [7] [19] [20] [21]
 Филогенетическая реконструкция опсинов. Внешняя группа содержит другие рецепторы, связанные с G-белком . В рамке выделены тетраопсины, которые расширены на следующем изображении.
Филогенетическая реконструкция опсинов. Внешняя группа содержит другие рецепторы, связанные с G-белком . В рамке выделены тетраопсины, которые расширены на следующем изображении. Филогенетическая реконструкция тетраопсинов. Внешняя группа содержит другие рецепторы, связанные с G-белком, включая другие опсины. В рамке выделены хромопсины, которые на следующем изображении расширены.
Филогенетическая реконструкция тетраопсинов. Внешняя группа содержит другие рецепторы, связанные с G-белком, включая другие опсины. В рамке выделены хромопсины, которые на следующем изображении расширены.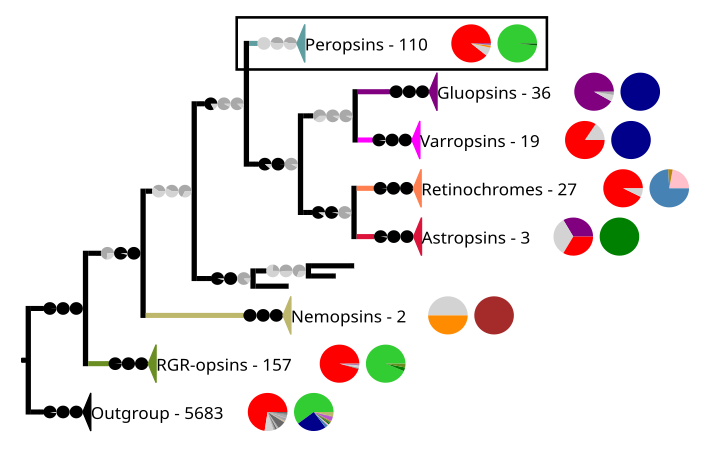 Филогенетическая реконструкция хромопсинов. Внешняя группа содержит другие рецепторы, связанные с G-белком, включая другие опсины. В рамке выделены перопсины.
Филогенетическая реконструкция хромопсинов. Внешняя группа содержит другие рецепторы, связанные с G-белком, включая другие опсины. В рамке выделены перопсины.



В приведенной выше филогении каждая клада содержит последовательности опсинов и других рецепторов, связанных с G-белком. Рядом с кладой показано количество последовательностей и две круговые диаграммы. Первая круговая диаграмма показывает процентное содержание определенной аминокислоты в положении последовательности, соответствующем положению 296 в родопсине крупного рогатого скота. Аминокислоты имеют цветовую маркировку. Цвета: красный для лизина (K), фиолетовый для глутаминовой кислоты (E), оранжевый для аргинина (R), тёмный и средне-серый для других аминокислот и светло-серый для последовательностей, по которым нет данных в этом положении. На второй круговой диаграмме показан таксонный состав каждой клады: зеленый цвет обозначает краниаты , темно-зеленый цвет — головохордовых , средне-зеленый — иглокожих , коричневый — нематод , бледно-розовый — кольчатых червей , темно-синий — членистоногих , светло-синий — моллюсков и фиолетовый — книдарий. . Ветви клад имеют круговые диаграммы, на которых указаны значения поддержки для ветвей. Значения указаны справа налево SH-aLRT/aBayes/UFBoot. Ветви считаются поддерживаемыми, если SH-aLRT ≥ 80%, aBayes ≥ 0,95 и UFBoot ≥ 95%. Если значение поддержки превышает пороговое значение, круговая диаграмма отображается черным цветом, в противном случае — серым. [7]
Клиническое значение
[ редактировать ]Поскольку RGR-опсин может быть связан с пигментным ретинитом , [22] который, как и перопсин, также экспрессируется в пигментном эпителии сетчатки, перопсин был проверен на связь с пигментным ретинитом. [23] Однако связь установить не удалось. [23] [24]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000180245 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000028012 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Перейти обратно: а б с д Сан Х., Гилберт Дж., Коупленд Н.Г., Дженкинс Н.А., Натанс Дж. (сентябрь 1997 г.). «Перопсин, новый зрительный пигментоподобный белок, расположенный в апикальных микроворсинках пигментного эпителия сетчатки» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (18): 9893–9898. Бибкод : 1997PNAS...94.9893S . дои : 10.1073/pnas.94.18.9893 . ПМК 23288 . ПМИД 9275222 .
- ^ Перейти обратно: а б с «Ген Энтрез: гомолог родопсина, полученный из пигментного эпителия сетчатки RRH» .
- ^ Перейти обратно: а б с д и ж г час Гюманн М., Портер М.Л., Бок М.Дж. (август 2022 г.). «Глюопсины: опсины без лизина, связывающего сетчатку» . Клетки . 11 (15): 2441. doi : 10.3390/cells11152441 . ПМЦ 9368030 . ПМИД 35954284 .
 Материал был скопирован и адаптирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован и адаптирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Люнг Нью-Йорк, Тхакур Д.П., Гурав А.С., Ким Ш., Ди Пизио А., Нив М.Ю. и др. (апрель 2020 г.). «Функции опсинов во вкусе дрозофилы» . Современная биология . 30 (8): 1367–1379.е6. Бибкод : 2020CBio...30E1367L . дои : 10.1016/j.cub.2020.01.068 . ПМЦ 7252503 . ПМИД 32243853 .
- ^ Перейти обратно: а б Коянаги М., Теракита А., Кубокава К., Шичида Ю. (ноябрь 2002 г.). «Амфиоксусные гомологи Go-связанного родопсина и перопсина, имеющие 11-цис- и полностью транс-ретинали в качестве хромофоров» . Письма ФЭБС . 531 (3): 525–528. дои : 10.1016/s0014-5793(02)03616-5 . ПМИД 12435605 . S2CID 11669142 .
- ^ Вальд Г. (июль 1934 г.). «Каротиноиды и цикл витамина А в зрении» . Природа . 134 (3376): 65. Бибкод : 1934Natur.134...65W . дои : 10.1038/134065a0 . S2CID 4022911 .
- ^ Уолд Г., Браун П.К., Хаббард Р., Орошник В. (июль 1955 г.). «Затрудненные цис-изомеры витамина А и ретинена: структура нео-B-изомера» . Труды Национальной академии наук Соединенных Штатов Америки . 41 (7): 438–451. Бибкод : 1955PNAS...41..438W . дои : 10.1073/pnas.41.7.438 . ПМК 528115 . ПМИД 16589696 .
- ^ Браун П.К., Уолд Г. (октябрь 1956 г.). «Нео-b-изомер витамина А и ретинена» . Журнал биологической химии . 222 (2): 865–877. дои : 10.1016/S0021-9258(20)89944-X . ПМИД 13367054 .
- ^ Орошник В. (июнь 1956 г.). «Синтез и конфигурация нео-витамина А и неоретинина b». Журнал Американского химического общества . 78 (11): 2651–2652. дои : 10.1021/ja01592a095 .
- ^ Орошник В., Браун П.К., Хаббард Р., Уолд Г. (сентябрь 1956 г.). «Затрудненные цис-изомеры витамина А и ретинена: структура нео-B-изомера» . Труды Национальной академии наук Соединенных Штатов Америки . 42 (9): 578–580. Бибкод : 1956PNAS...42..578O . дои : 10.1073/pnas.42.9.578 . ПМК 534254 . ПМИД 16589909 .
- ^ Кук Дж.Д., Нг С.И., Ллойд М., Эддингтон С., Сан Х., Натанс Дж. и др. (декабрь 2017 г.). «Перопсин модулирует транзит витамина А из сетчатки в пигментный эпителий сетчатки» . Журнал биологической химии . 292 (52): 21407–21416. дои : 10.1074/jbc.M117.812701 . ПМК 5766940 . ПМИД 29109151 .
- ^ Тох П.П., Бильярди-Ци М., Яп А.М., Шрирам Г., Стельмашенко О., Бильярди П. (декабрь 2016 г.). «Экспрессия перопсина в коже человека связана с фототрансдукцией фиолетового света в кератиноцитах» . Экспериментальная дерматология . 25 (12): 1002–1005. дои : 10.1111/exd.13226 . ПМИД 27676658 . S2CID 1373924 .
- ^ Бейли М.Дж., Кассоне В.М. (март 2004 г.). «Фотоизомеразы опсина в сетчатке и шишковидной железе кур: характеристика, локализация и циркадная регуляция». Исследовательская офтальмология и визуальные науки . 45 (3): 769–775. дои : 10.1167/iovs.03-1125 . ПМИД 14985289 .
- ^ Перейти обратно: а б с Беллингем Дж., Уэллс DJ, Фостер Р.Г. (январь 2003 г.). «Характеристика in silico и хромосомная локализация RRH человека (перопсина) - последствия для эволюции опсина» . БМК Геномика . 4 (1): 3. дои : 10.1186/1471-2164-4-3 . ПМК 149353 . ПМИД 12542842 .
- ^ Перейти обратно: а б с Рамирес М.Д., Пайретт А.Н., Панки М.С., Серб Дж.М., Спейзер Д.И., Сваффорд А.Дж. и др. (26 октября 2016 г.). «Последний общий предок большинства двусторонних животных обладал по крайней мере 9 опсинами» . Геномная биология и эволюция : evw248. дои : 10.1093/gbe/evw248 . ПМЦ 5521729 . ПМИД 27797948 .
- ^ Перейти обратно: а б Портер М.Л., Бласик Дж.Р., Бок М.Дж., Кэмерон Э.Г., Прингл Т., Кронин Т.В. и др. (январь 2012 г.). «Проливая новый свет на эволюцию опсинов» . Слушания. Биологические науки . 279 (1726): 3–14. дои : 10.1098/rspb.2011.1819 . ПМЦ 3223661 . ПМИД 22012981 .
- ^ Лигертова М., Пергнер Дж., Козьмикова И., Фабиан П., Помбиньо А.Р., Стрнад Х. и др. (июль 2015 г.). «Геном кубозоев проливает свет на функциональное разнообразие опсинов и эволюцию фоторецепторов» . Научные отчеты . 5 : 11885. Бибкод : 2015NatSR...511885L . дои : 10.1038/srep11885 . ПМК 5155618 . ПМИД 26154478 .
- ^ Моримура Х., Сенделль-Рибодо Ф., Берсон Э.Л., Дрия Т.П. (декабрь 1999 г.). «Мутации в RGR, кодирующем светочувствительный гомолог опсина, у пациентов с пигментным ретинитом». Природная генетика . 23 (4): 393–394. дои : 10.1038/70496 . ПМИД 10581022 . S2CID 35176366 .
- ^ Перейти обратно: а б Ксантини М., Сенешаль А., Умберт Г., Арно Б., Амель К.П. (март 2007 г.). «RRH, кодирующий опсиноподобный перопсин, экспрессируемый RPE, не мутирует при пигментном ретините и родственных заболеваниях» (PDF) . Офтальмологическая генетика . 28 (1): 31–37. дои : 10.1080/13816810701202052 . ПМИД 17454745 . S2CID 225451 .
- ^ Риволта С., Берсон Э.Л., Дрия Т.П. (декабрь 2006 г.). «Скрининг мутаций гена перопсина, специфического гомолога родопсина пигментного эпителия сетчатки, у пациентов с пигментным ретинитом и родственными заболеваниями». Молекулярное видение . 12 : 1511–1515. ПМИД 17167409 .
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .