Парафилия
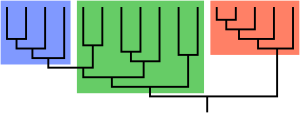
Парафилия — это таксономический группы термин, описывающий группу, состоящую из последнего общего предка и некоторых, но не всех, ее потомков. Группировка называется парафилетической по отношению к исключенным подгруппам. Напротив, монофилетическая группа ( клада ) включает общего предка и всех его потомков.
Эти термины обычно используются в филогенетике (раздел биологии ) и в древовидной модели исторической лингвистики . Парафилетические группы идентифицируются по сочетанию синапоморфий и симплезиоморфий . Если в названной группе отсутствуют многие подгруппы, ее называют полипарафилетической.
Этот термин получил распространение во время дебатов 1960-х и 1970-х годов, сопровождавших подъем кладистики , будучи придуманным зоологом Вилли Хеннигом для применения к таким известным таксонам, как Reptilia ( рептилии ), которые являются парафилетическими по отношению к птицам . Reptilia содержит последнего общего предка рептилий и всех потомков этого предка, кроме птиц. Другие общепризнанные парафилетические группы включают рыб , обезьян и ящериц . [1] [ нужна страница ]
Этимология
[ редактировать ]Термин парафилия , или парафилетика , происходит от двух древнегреческих слов παρά ( пара ), означающих «рядом, рядом», и φῦλον ( phûlon ), означающих «род, вид». [2] [3] и относится к ситуации, в которой одна или несколько монофилетических подгрупп организмов (например, родов, видов) остаются обособленными от всех других потомков уникального общего предка.
И наоборот, термин «монофилия », или «монофилетика », основан на древнегреческом префиксе μόνος ( монос ), означающем «единственный, единственный, уникальный». [2] [3] и относится к тому, что к монофилетической группе относятся организмы, состоящие из всех потомков единственного общего предка.
Для сравнения, термин полифилия или полифилетика использует древнегреческий префикс πολύς ( polús ), что означает «много, много». [2] [3] и относится к тому факту, что полифилетическая группа включает организмы, возникшие из нескольких предковых источников.
Филогенетика
[ редактировать ]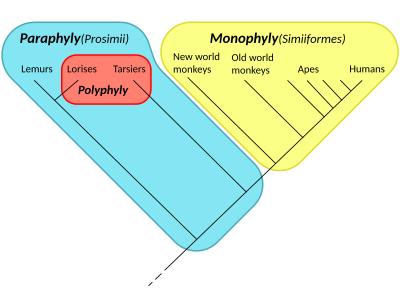
В кладистике
[ редактировать ]Группы, включающие всех потомков общего предка, называются монофилетическими . Парафилетическая группа — это монофилетическая группа, из которой исключены одна или несколько дочерних клад (монофилетические группы), образующие отдельную группу. Философ науки Марк Эрешефски утверждал, что парафилетические таксоны являются результатом анагенеза в исключенной группе или группах. [4] Кладистический подход обычно не предоставляет парафилетическим комплексам статус «групп» и не овеществляет их объяснениями, поскольку в кладистике они не рассматриваются как действительные продукты эволюционных событий. [5]
Группа, отличительные признаки которой развивались конвергентно в двух или более линиях, является полифилетической (греч. πολύς [ полис ], «многие»). В более широком смысле любой таксон, который не является парафилетическим или монофилетическим, можно назвать полифилетическим. Эмпирически различие между полифилетическими группами и парафилетическими группами довольно условно, поскольку состояния характера общих предков являются умозаключениями, а не наблюдениями. [ нужна ссылка ]
Эти термины были разработаны в ходе дебатов 1960-х и 1970-х годов, сопровождавших подъем кладистики .
Парафилетические группировки многие систематики считают проблематичными, поскольку невозможно точно говорить об их филогенетических связях, характерных чертах и буквальном вымирании. [6] [7] Родственные термины — это стволовая группа , хроновид , почковающийся кладогенез, анагенез или группы «сортов» . Парафилетические группы часто являются реликтами устаревших гипотез филогенетических взаимоотношений, существовавших еще до появления кладистики. [8]
Примеры
[ редактировать ]
Прокариоты , (одноклеточные формы жизни без клеточных ядер) представляют собой парафилетическую группу, поскольку они исключают эукариотов группу потомков. Бактерии и археи являются прокариотами, но археи и эукариоты имеют общего предка, который не является предком бактерий. Различие между прокариотами и эукариотами было предложено Эдуардом Шаттоном в 1937 году. [9] и был общепринятым после того, как был принят Роджером Станьером и CB ван Нилом в 1962 году. Ботанический кодекс (ICBN, ныне ICN ) отказался от рассмотрения бактериальной номенклатуры в 1975 году; в настоящее время номенклатура прокариот регулируется ICNB с начальной датой 1 января 1980 г. (в отличие от даты начала 1753 г. согласно ICBN/ICN). [10]
Среди растений двудольные (в традиционном понимании) являются парафилетическими, поскольку из группы исключаются однодольные . «Двудольные» десятилетиями не использовались в качестве ботанической классификации, но разрешены как синоним Magnoliopsida. [примечание 1] Филогенетический анализ показывает, что однодольные произошли от предка двудольных . Исключение однодольных из двудольных делает последние парафилетической группой. [11]
Среди животных несколько известных групп на самом деле не являются кладами. Отряд парнокопытные ( парнокопытные ) в традиционном определении является парафилетическим, поскольку исключает китообразных (китов, дельфинов и т. д.). В соответствии с Кодексом МКЗН эти два таксона представляют собой отдельные отряды. Однако молекулярные исследования показали, что китообразные происходят от предков парнокопытных, хотя точная филогения внутри отряда остается неопределенной. Без китообразных парнокопытные парафилетичны. [12] Класс Reptilia является парафилетическим, поскольку исключает птиц (класс Aves ). Согласно традиционной классификации, эти два таксона представляют собой отдельные классы. Однако птицы являются сестринским таксоном группы динозавров (часть Diapsida ), оба из которых являются «рептилиями». [13]
Osteichthyes , костистые рыбы, являются парафилетическими, если включить только Actinopterygii (рыба с лучеперыми плавниками) и Sarcopterygii (двоякодышащие рыбы и т. д.), а также исключить четвероногих ; в последнее время Osteichthyes рассматривают как кладу, включающую четвероногих. [14] [15]
« Осы » парафилетичны и состоят из узкоталийного Апокрита без муравьев и пчел . [16] Пилильщики ( Symphyta ) также парафилетичны, образуя все перепончатокрылые, за исключением Apocrita, клады глубоко внутри дерева пилильщиков. [14] Ракообразные не являются кладой, потому что Hexapoda (насекомые) исключены. Современная клада, охватывающая их все, — это Тетраконата . [17] [18]
Одной из целей современной систематики за последние пятьдесят лет было исключение парафилетических «групп», таких как приведенные здесь примеры, из формальных классификаций. [19] [20]
Парафилия у видов
[ редактировать ]Виды имеют в систематике особый статус как наблюдаемый признак самой природы и как основная классификационная единица. [21] Некоторые формулировки концепции филогенетических видов требуют, чтобы виды были монофилетическими, но парафилетические виды распространены в природе до такой степени, что у них нет единого общего предка. Действительно, среди таксонов, размножающихся половым путем, ни один вид не имеет организма «единого общего предка». Парафилия распространена в видообразовании , при которой материнский вид ( паравид ) дает начало дочернему виду, не вымирая при этом. [22] Исследования показывают, что до 20 процентов всех видов животных и от 20 до 50 процентов видов растений являются парафилетическими. [23] [24] Учитывая эти факты, некоторые систематики утверждают, что парафилия — это черта природы, которую следует признать на более высоких таксономических уровнях. [25] [26]
Кладисты отстаивают концепцию филогенетических видов. [27] это не предполагает, что виды проявляют свойства монофилии или парафилии, концепции с этой точки зрения применимы только к группам видов. [28] Они считают, что распространение Зандером аргумента о «парафилетических видах» на высшие таксоны представляет собой категориальную ошибку. [29]
Использование парафилетических групп
[ редактировать ]Когда появление существенных черт привело субклад на эволюционный путь, сильно отличающийся от пути более инклюзивной клады, часто имеет смысл изучать оставшуюся парафилетическую группу, не принимая во внимание более крупную кладу. Например, неогеновая эволюция парнокопытных , семейства , (парнокопытные, такие как олени, коровы, свиньи и бегемоты — Cervidae , Bovidae , Suidae и Hippopotamidae содержащие эти различные парнокопытные, представляют собой монофилетические группы) происходила в средах настолько отличаются от китообразных ( китов, дельфинов и морских свиней), что парнокопытных часто изучают изолированно, хотя китообразные являются потомком группы. Группа прокариот — еще один пример; он парафилетичен, поскольку состоит из двух доменов (эубактерий и архей) и исключает ( эукариоты ). Он очень полезен, поскольку имеет четко определенные и существенные отличия (отсутствие клеточного ядра, плезиоморфия ) от своих исключенных потомков. [ нужна ссылка ]
Кроме того, некоторые систематики признают, что парафилетические группы участвуют в эволюционных переходах, например, в развитии первых четвероногих от их предков. Любое имя, данное этим гипотетическим предкам, чтобы отличить их от четвероногих - например, «рыбы», - обязательно выделяет парафилетическую группу, потому что потомки четвероногих не включены. [30] Другие систематики считают, что реификация парафилетических групп скрывает предполагаемые закономерности эволюционной истории. [31]
Термин « эволюционная степень » иногда используется для парафилетических групп. [32] Более того, концепции монофилии , парафилии и полифилии использовались при выведении ключевых генов для штрих-кодирования различных групп видов. [33]
Независимо развитые черты
[ редактировать ]Современные филогенетические гипотезы взаимоотношений четвероногих предполагают, что живорождение , производство потомства без внешней откладки оплодотворенного яйца, развилось независимо в линиях, которые привели к человеку ( Homo sapiens ) и южным водным сцинкам ( Eulampus tympanum , разновидность ящерицы). Другими словами, живорождение — это синапоморфия Theria у млекопитающих и аутапоморфия Eulamprus tympanum (или, возможно, синапоморфия, если другие виды Eulamprus также являются живородящими). [ нужна ссылка ]
Группы, основанные на независимо развитых чертах, такие как эти примеры живорождения, представляют собой примеры полифилии , а не парафилии. [ нужна ссылка ]
Не парафилия
[ редактировать ]- Земноводные рыбы полифилетичны, а не парафилетичны. Хотя они кажутся похожими, несколько различных групп земноводных рыб, таких как илистые прыгуны и двоякодышащие рыбы, развивались независимо в процессе конвергентной эволюции у дальних родственников, столкнувшихся со схожими экологическими обстоятельствами. [34]
- Нелетающие птицы полифилетичны, поскольку самостоятельно (параллельно) утратили способность летать. [35]
- Животные со спинным плавником не являются парафилетическими, хотя их последний общий предок мог иметь такой плавник, поскольку у мезозойских предков морских свиней такого плавника не было, тогда как у домезозойских рыб он был.
- Четвероногие архозавры не являются парафилетической группой. Двуногие динозавры, такие как эораптор , предки четвероногих, были потомками последнего общего предка четвероногих динозавров и других четвероногих архозавров, таких как крокодилы.
Неисчерпывающий список парафилетических групп
[ редактировать ]В следующем списке резюмируется ряд парафилетических групп, предложенных в литературе, и представлены соответствующие монофилетические таксоны.
Лингвистика
[ редактировать ]Понятие парафилии также было применено к исторической лингвистике , где методы кладистики нашли некоторую полезность при сравнении языков. Например, формозские языки образуют парафилетическую группу австронезийских языков , поскольку они состоят из девяти ветвей австронезийской семьи, которые не являются малайско-полинезийскими и распространены только на острове Тайвань . [67]
См. также
[ редактировать ]Пояснительные примечания
[ редактировать ]- ^ Историю классификации цветковых растений можно найти в разделе « История классификации цветковых растений» .
Ссылки
[ редактировать ]- ^ Ромер, А.С. (1970) [1949]. Тело позвоночного (4-е изд.). УБ Сондерс.
- ^ Jump up to: Перейти обратно: а б с Байи, Анатоль (1 января 1981 г.). Сокращено из Французско-греческого словаря . Париж: Хашетт. ISBN 978-2-01-003528-9 . OCLC 461974285 .
- ^ Jump up to: Перейти обратно: а б с Байи, Анатоль. «Греко-французский словарь онлайн» . www.tabularium.be . Проверено 8 марта 2018 г.
- ^ Робертс, Кейт (10 декабря 2007 г.). Справочник по растениеводству . Джон Уайли и сыновья. ISBN 978-0-470-05723-0 .
- ^ Уильямс, Д.М. и Эбах. МК 2020. Кладистика: руководство по биологической классификации. Издательство Кембриджского университета.
- ^ Шилхаб, Тереза; Стьернфельт, Фредерик; Диакон, Терренс (2012). Эволюция символических видов . Спрингер. ISBN 978-94-007-2335-1 .
- ^ Виллмоаре, Брайан (2018). «Ранний человек и роль рода в палеоантропологии» . Американский журнал физической антропологии . 165 : 72–89. дои : 10.1002/ajpa.23387 . ПМИД 29380889 .
- ^ Домингес, Эдуардо; Уилер, Квентин Д. (1997). «Форум – Таксономическая стабильность – это невежество» . Кладистика . 13 (4): 367–372. дои : 10.1111/j.1096-0031.1997.tb00325.x . ПМИД 34911226 . S2CID 55540349 .
- ^ Сапп, Ян (июнь 2005 г.). «Дихотомия прокариот-эукариот: значения и мифология» . Обзоры микробиологии и молекулярной биологии . 69 (2): 292–305. дои : 10.1128/MMBR.69.2.292-305.2005 . ПМЦ 1197417 . ПМИД 15944457 .
- ^ Стакебрабдт, Э.; Тинделл, Б.; Людвиг, В.; Гудфеллоу, М. (1999). «Прокариотическое разнообразие и систематика». В Ленгелере, Джозеф В.; Дрюс, Герхарт; Шлегель, Ганс Гюнтер (ред.). Биология прокариот . Штутгарт: Георг Тиме Верлаг. п. 679.
- ^ Jump up to: Перейти обратно: а б Simpson 2006 , стр. 139–140. . «Сейчас считается, что наличие двух семядолей является наследственной особенностью таксонов цветковых растений, а не апоморфией какой-либо группы внутри. «Двудольные»… парафилетичны…».
- ^ Jump up to: Перейти обратно: а б О'Лири, Морин А. (2001). «Филогенетическое положение китообразных: дальнейший комбинированный анализ данных, сравнение со стратиграфическими данными и обсуждение оптимизации признаков» . Американский зоолог . 41 (3): 487–506. CiteSeerX 10.1.1.555.8631 . дои : 10.1093/icb/41.3.487 .
- ^ Ромер, А.С. и Парсонс, Т.С. (1985): Тело позвоночного. (6-е изд.) Сондерс, Филадельфия.
- ^ Jump up to: Перейти обратно: а б с Шарки, MJ (2007). «Филогения и классификация перепончатокрылых» (PDF) . Зоотакса . 1668 : 521–548. дои : 10.11646/zootaxa.1668.1.25 .
Symphyta и Apocrita долгое время считались подотрядами Hymenoptera, но с момента признания парафилетической природы Symphyta (Кёнингсманн, 1977, Расницын, 1988) и появления кладистических методов субпорядковой классификации следует избегать. Точно так же древесные осы считаются немонофилетическими, образуя класс, являющийся предком по отношению к Apocrita и Orussidae. Традиционная классификация перепончатокрылых ошибочна по кладистическим критериям, точно так же, как докладистические классификации позвоночных, в которых группы, имеющие плезиоморфные признаки, признавались естественными, например, рыбы когда-то были сгруппированы как «Рыбы», что исключало четвероногих.
- ^ Бетанкур-Р, Рикардо; и др. (2013). «Древо жизни и новая классификация костистых рыб» . ПЛОС Токи Древо Жизни . 5 (Издание 1). doi : 10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288 . hdl : 2027.42/150563 . ПМЦ 3644299 . ПМИД 23653398 . Архивировано из оригинала 13 октября 2013 года.
- ^ Jump up to: Перейти обратно: а б с Джонсон, Брайан Р.; Боровец, Марек Л.; Чиу, Джоанна С.; Ли, Эрнест К.; Аталлах, Джоэл; Уорд, Филип С. (2013). «Филогеномика решает эволюционные отношения между муравьями, пчелами и осами» (PDF) . Современная биология . 23 (20): 2058–2062. Бибкод : 2013CBio...23.2058J . дои : 10.1016/j.cub.2013.08.050 . ПМИД 24094856 . S2CID 230835 .
- ^ Jump up to: Перейти обратно: а б Дэвид Р. Эндрю (2011). «Новый взгляд на взаимоотношения насекомых и ракообразных II. Выводы на основе выраженных меток последовательностей и сравнения с нейронной кладистикой». Строение и развитие членистоногих . 40 (3): 289–302. Бибкод : 2011АртСД..40..289А . дои : 10.1016/j.asd.2011.02.001 . ПМИД 21315832 .
- ^ Jump up to: Перейти обратно: а б Бьерн, М.; фон Роймонт, Рональд А.; Дженнер, Мэтью А.; Уиллс, Эмилиано; Делл'Ампио, Гюнтер; Пасс, Инго; Эберсбергер, Бенджамин; Мейер, Стефан; Кенеманн, Томас М. Илифф (2012). «Филогения панкрустообразных в свете новых филогеномических данных: поддержка Remipedia как возможной сестринской группы Hexapoda» ( в формате PDF доказательства ) . Молекулярная биология и эволюция . 29 (3): 1031–1045. дои : 10.1093/molbev/msr270 . ПМИД 22049065 .
- ^ Шу, Рэндалл Т. «Система Линнея и ее 250-летнее существование». Ботаническое обозрение 69, вып. 1 (2003): 59.
- ^ Брауэр, Эндрю В.З. (2020). «Мертв по прибытии: посмертная оценка «филогенетической номенклатуры», более 20 лет спустя» . Кладистика . 36 (6): 627–637. дои : 10.1111/cla.12432 . S2CID 224927279 .
- ^ Кейрос, Кевин; Донохью, Майкл Дж. (декабрь 1988 г.). «Филогенетическая систематика и проблема видов». Кладистика . 4 (4): 317–338. дои : 10.1111/j.1096-0031.1988.tb00518.x . ПМИД 34949064 . S2CID 40799805 .
- ^ Альберт, Джеймс С.; Рейс, Роберто Э. (8 марта 2011 г.). Историческая биогеография неотропических пресноводных рыб . Издательство Калифорнийского университета. п. 308. ИСБН 978-0-520-26868-5 . Проверено 28 июня 2011 г. - через Google Книги.
- ^ Росс, Ховард А. (июль 2014 г.). «Заболеваемость парафилией на видовом уровне у животных: повторная оценка». Молекулярная филогенетика и эволюция . 76 : 10–17. дои : 10.1016/j.ympev.2014.02.021 . ПМИД 24583289 .
- ^ Крисп, доктор медицины; Чендлер, GT (1 июля 1996 г.). «Парафилетические виды» . Телопея . 6 (4): 813–844. дои : 10.7751/telopea19963037 . Проверено 22 января 2015 г.
- ^ Зандер, Ричард (2013). Основы постфилогенетической систематики . Сент-Луис: Zetetic Publications, Amazon CreateSpace.
- ^ Обер, Д. (2015). «Формальный анализ филогенетической терминологии: на пути к пересмотру современной парадигмы в систематике». Фитонейрон . 66 : 1–54.
- ^ Никсон, Кевин С.; Уилер, Квентин Д. (1990). «Расширение концепции филогенетического вида» . Кладистика . 6 (3): 211–23. дои : 10.1111/j.1096-0031.1990.tb00541.x . S2CID 84095773 .
- ^ Брауэр, Эндрю В.З.; Шу, Рэндалл Т. (2021). Биологическая систематика: принципы и приложения (3-е изд.). Итака, Нью-Йорк: Издательство Корнельского университета. ISBN 978-1-5017-5277-3 .
- ^ Шмидт-Лебун, Александр Н. (2012). «Заблуждения и ложные предпосылки — критическая оценка аргументов в пользу признания парафилетических таксонов в ботанике» . Кладистика . 28 (2): 174–87. дои : 10.1111/j.1096-0031.2011.00367.x . ПМИД 34861757 . S2CID 83900580 .
- ^ Казлев М.А. и Уайт Т. «Земноводные, систематика и кладистика» . Палеоса Сайт . Проверено 16 августа 2012 г.
- ^ Паттерсон, Колин (1982). «Морфология и взаимоотношения примитивных актиноптеригийных рыб» . Американский зоолог . 22 (2): 241–259. дои : 10.1093/icb/22.2.241 .
- ^ Докинз, Ричард (2004). «Млекопитающие рептилии». Рассказ предка, паломничество к заре жизни . Бостон: Хоутон Миффлин. ISBN 978-0-618-00583-3 .
- ^ Пархи, Дж.; Трипатия, ПС; Приядарши, Х.; Мандал СК; Пандей ПК (2019). «Диагностика митогенома для надежной филогении: случай группы рыб Cypriniformes». Джин . 713 : 143967. doi : 10.1016/j.gene.2019.143967 . ПМИД 31279710 . S2CID 195828782 .
- ^ Кучера, Ульрих; Эллиотт, Дж. Малкольм (26 марта 2013 г.). «Проясняют ли илистые прыгуны и двоякодышащие рыбы раннюю эволюцию четырехконечных позвоночных?» . Эволюция: образование и информационно-пропагандистская деятельность . 6 (8): 8. дои : 10.1186/1936-6434-6-8 .
- ^ Харшман, Джон; Браун, Эдвард Л.; и др. (2 сентября 2008 г.). «Филогеномные доказательства множественной потери полета у бескилевых птиц» . ПНАС . 105 (36): 13462–13467. Бибкод : 2008PNAS..10513462H . дои : 10.1073/pnas.0803242105 . ПМК 2533212 . ПМИД 18765814 .
- ^ Берг, Линда (2008). Вводная ботаника: растения, люди и окружающая среда (2-е изд.). Бельмонт, Калифорния: Thomson Corporation. п. 360. ИСБН 978-0-03-075453-1 .
- ^ Шлегель, Мартин; Хюльсманн, Норберт (2 августа 2007 г.). «Протисты - хрестоматийный пример парафилетического таксона». Разнообразие и эволюция организмов . 7 (2): 166–172. дои : 10.1016/j.ode.2006.11.001 . ISSN 1439-6092 .
- ^ Агассис, Луи (21 марта 2013 г.). Эссе по классификации . Курьер. стр. 115–. ISBN 978-0-486-15135-9 .
- ^ Федонкин, М.А.; Симонетта, А; Иванцов, А.Ю. (2007). «Новые данные о кимберелле , вендском моллюскообразном организме (Беломорье, Россия): палеоэкологическое и эволюционное значение». В Викерс-Рич, Патрисия; Комарауэр, Патрисия (ред.). Взлет и падение эдиакарской биоты . Специальные публикации. Том. 286. Лондон: Геологическое общество. стр. 157–179. дои : 10.1144/SP286.12 . ISBN 978-1-86239-233-5 . OCLC 156823511 .
- ^ Баттерфилд, Нью-Джерси (декабрь 2006 г.). «Зацепка некоторых «червей» стволовой группы: ископаемые лофотрохозойи в сланцах Берджесс». Биоэссе . 28 (12): 1161–6. doi : 10.1002/bies.20507 . ПМИД 17120226 . S2CID 29130876 .
- ^ Мартиндейл, Марк; Финнерти-младший; Генри, JQ (сентябрь 2002 г.). «Ладиаты и эволюционное происхождение двустороннего строения тела». Молекулярная филогенетика и эволюция . 24 (3): 358–365. дои : 10.1016/s1055-7903(02)00208-7 . ПМИД 12220977 .
- ^ «Gnathifera - Ричард К. Бруска» (PDF) .
- ↑ Веб-проект «Древо жизни» — Chordates . Архивировано 24 февраля 2007 года в Wayback Machine .
- ^ Тадж, Колин (2000). Разнообразие жизни . Издательство Оксфордского университета. ISBN 0-19-860426-2 .
- ^ Ридер, Тод В.; Таунсенд, Тед М.; Малкахи, Дэниел Г.; Нунан, Брайс П.; Вуд, Перри Л.; Сайты, Джек В.; Винс, Джон Дж. (2015). «Комплексный анализ разрешает конфликты по поводу филогении чешуйчатых рептилий и выявляет неожиданные места размещения ископаемых таксонов» . ПЛОС ОДИН . 10 (3): e0118199. Бибкод : 2015PLoSO..1018199R . дои : 10.1371/journal.pone.0118199 . ПМЦ 4372529 . ПМИД 25803280 .
- ^ Килан-Яворовска З. и Хурум Дж. (2001). «Филогения и систематика мультитуберкулезных животных» (PDF) . Палеонтология . 44 (3): 389–429. Бибкод : 2001Palgy..44..389K . дои : 10.1111/1475-4983.00185 . S2CID 83592270 .
- ^ Бентон, Майкл Дж. (2004). Палеонтология позвоночных (3-е изд.). Оксфорд: Блэквелл Наука. ISBN 978-0-632-05637-8 .
- ^ Сэвидж, Р.Дж.Г. и Лонг, М.Р. (1986). Эволюция млекопитающих: иллюстрированное руководство . Нью-Йорк: факты в архиве. стр. 208 . ISBN 0-8160-1194-Х .
- ^ Тьювиссен, JGM; Уильямс, Э.М. (2002). «Ранние проявления китообразных (млекопитающих): закономерности эволюции и корреляции развития». Ежегодный обзор экологии и систематики . 33 : 73–90. doi : 10.1146/annurev.ecolsys.33.020602.095426 . OCLC 4656321698 .
- ^ Гроувс, КП (1998). «Систематика долгопятов и лори». Приматы . 39 (1): 13–27. дои : 10.1007/BF02557740 . S2CID 10869981 .
- ^ Паразитические перепончатокрылые (Parasitica). Р. Л. Зупарко, Энциклопедия энтомологии, 2004 г.
- ^ Линдгрен, Арканзас; Гирибет, Г.; Нисигути, МК (2004). «Комбинированный подход к филогении головоногих (Mollusca)» . Кладистика . 20 (5): 454–486. дои : 10.1111/j.1096-0031.2004.00032.x . ПМИД 34892953 . S2CID 85975284 .
- ^ Беккер, Б.; Марин, Б. (2009). «Водоросли-стрептофиты и происхождение эмбриофитов» . Анналы ботаники . 103 (7): 999–1004. дои : 10.1093/aob/mcp044 . ПМК 2707909 . ПМИД 19273476 .
- ^ Скобл, MJ (1995). Чешуекрылые: форма, функции и разнообразие . Оксфорд: Издательство Оксфордского университета. п. 404.
- ^ Стампар, С.Н.; Маронна, ММ; Китахара, М.В.; Реймер, доктор медицинских наук; Морандини, AC (2014). «Быстро развивающаяся митохондриальная ДНК у цериантариев: отражение парафилии гексакораллий?» . ПЛОС ОДИН . 9 (1): е86612. Бибкод : 2014PLoSO...986612S . дои : 10.1371/journal.pone.0086612 . ПМЦ 3903554 . ПМИД 24475157 .
- ^ Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). «7 КНИДАРИЯ». Зоология беспозвоночных: функциональный эволюционный подход (7-е изд.). Бельмонт, Калифорния: Томсон-Брукс/Коул . стр. 132–148. ISBN 0-03-025982-7 . LCCN 2003107287 . OCLC 752875516 .
- ^ Цзоу, Х.; Чжан, Дж.; Ли, В.; Ву, С.; Ван, Г. (2012). «Митохондриальный геном пресноводной медузы Craspedacusta sowerbyi и филогенетика Medusozoa» . ПЛОС ОДИН . 7 (12): е51465. Бибкод : 2012PLoSO...751465Z . дои : 10.1371/journal.pone.0051465 . ПМК 3519871 . ПМИД 23240028 .
- ^ Маркес, Антонио К.; Аллен Г. Коллинз (март 2004 г.). «Кладистический анализ Medusozoa и эволюции книдарий». Биология беспозвоночных . 123 (1): 23–42. дои : 10.1111/j.1744-7410.2004.tb00139.x .
- ^ Сапата; и др. (2015). «Филогеномный анализ подтверждает традиционные отношения внутри Cnidaria» . ПЛОС ОДИН . 10 (10): e0139068. Бибкод : 2015PLoSO..1039068Z . дои : 10.1371/journal.pone.0139068 . ПМЦ 4605497 . ПМИД 26465609 .
- ^ Данн, CW; Хейнол, А; Матус, DQ; Панг, К; Браун, МЫ; Смит, ЮАР; Сивер, Э; Роуз, GW; и др. (2008). «Широкая филогеномная выборка улучшает разрешение древа жизни животных». Природа . 452 (7188): 745–749. Бибкод : 2008Natur.452..745D . дои : 10.1038/nature06614 . ПМИД 18322464 . S2CID 4397099 .
- ^ Вебстер, Бонни Л.; Копли, Ричард Р.; Дженнер, Рональд А.; Маккензи-Доддс, Жаклин А.; Бурла, Сара Дж.; Рота-Стабелли, Омар; Литтлвуд, DTJ; Телфорд, Максимилиан Дж. (ноябрь 2006 г.). «Митогеномика и филогомика выявили приапулидных червей как существующие модели предков Ecdysozoa». Эволюция и развитие . 8 (6): 502–510. дои : 10.1111/j.1525-142X.2006.00123.x . ПМИД 17073934 . S2CID 22823313 .
- ^ Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). «23 ГНАТИФЕРЫ». Зоология беспозвоночных: функциональный эволюционный подход (7-е изд.). Бельмонт, Калифорния: Томсон-Брукс/Коул . стр. 788 и далее. – см., в частности, стр. 804. ИСБН 0-03-025982-7 . LCCN 2003107287 . OCLC 752875516 .
- ^ Шимек, Рональд (январь 2006 г.). «Наноживотные, Часть I: Коловушки» . Reefkeeping.com . Проверено 27 июля 2008 г.
- ^ АронРа (16 января 2010 г.). Оказывается, мы произошли от обезьян! . Проверено 12 ноября 2018 г.
- ^ «Ранняя эволюция приматов: первые приматы» . anthro.palomar.edu . Архивировано из оригинала 10 января 2018 года . Проверено 12 августа 2017 г.
- ^ Уилсон, Дон Э. и Ридер, ДиЭнн М. (2005). Виды млекопитающих мира: Таксономический и географический справочник, Том 1 (3-е изд.). Бельмонт, Калифорния: Издательство Университета Джонса Хопкинса. ISBN 0-8018-8221-4 . , с. 699.
- ^ Гринхилл, Саймон Дж. и Рассел Д. Грей. (2009.) «Австронезийский язык и филогения: мифы и заблуждения о байесовских вычислительных методах», в книге «Австронезийская историческая лингвистика и история культуры: праздничный сборник для Роберта Бласта » под редакцией Александра Аделаара и Эндрю Поли. Канберра: Тихоокеанская лингвистика, Исследовательская школа тихоокеанских и азиатских исследований, Австралийский национальный университет .
Библиография
[ редактировать ]- Симпсон, Майкл Джордж (2006). Систематика растений . Берлингтон; Сан-Диего; Лондон: Академическая пресса . ISBN 978-0-12-644460-5 .
- Парафилетические группы как естественные единицы биологической классификации
Внешние ссылки
[ редактировать ] СМИ, связанные с парафилией, на Викискладе?
СМИ, связанные с парафилией, на Викискладе? - Фанк, диджей; Омланд, Кентукки (2003). «Парафилия и полифилия на видовом уровне: частота, причины и последствия, с учетом данных митохондриальной ДНК животных». Ежегодный обзор экологии, эволюции и систематики . 34 : 397–423. doi : 10.1146/annurev.ecolsys.34.011802.132421 . S2CID 33951905 .
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||