Видообразование
| Часть серии о |
| Эволюционная биология |
|---|
 |
Видообразование – это эволюционный процесс, в ходе которого популяции превращаются в отдельные виды . Биолог Оратор Ф. Кук ввел термин в 1906 году для кладогенеза , разделения линий, в отличие от анагенеза , филетической эволюции внутри линий. [1] [2] [3] Чарльз Дарвин был первым, кто описал роль естественного отбора в видообразовании в своей книге 1859 года «Происхождение видов» . [4] Он также назвал половой отбор вероятным механизмом, но счел его проблематичным.
В природе существует четыре географических способа видообразования, основанные на степени видообразующих популяций изоляции друг от друга: аллопатрический , перипатрический , парапатрический и симпатрический . Является ли генетический дрейф второстепенным или основным фактором видообразования, является предметом многочисленных продолжающихся дискуссий. [5]
Быстрое симпатрическое видообразование может происходить посредством полиплоидии , например, за счет удвоения числа хромосом; в результате получается потомство, которое немедленно репродуктивно изолируется от родительской популяции. Новые виды также могут быть созданы путем гибридизации с последующей репродуктивной изоляцией, если естественный отбор благоприятствует гибриду. [ нужна ссылка ]
Историческая справка [ править ]
При рассмотрении происхождения видов есть два ключевых вопроса:
- эволюционные механизмы видообразования
- как сохраняется обособленность и индивидуальность видов
Со времен Чарльза Дарвина усилия по пониманию природы видов в основном были сосредоточены на первом аспекте, и сейчас широко распространено мнение, что решающим фактором возникновения новых видов является репродуктивная изоляция. [6]
Дилемма Дарвина: почему существуют виды? [ редактировать ]
В книге «Происхождение видов» (1859) Дарвин интерпретировал биологическую эволюцию с точки зрения естественного отбора, но был озадачен группировкой организмов в виды. [7] Глава 6 книги Дарвина называется «Трудности теории». Обсуждая эти «трудности», он отметил
Во-первых, почему, если виды произошли от других видов посредством незаметно тонких ступеней, то мы не видим повсюду бесчисленных переходных форм? Почему не вся природа находится в замешательстве, а виды, как мы их видим, четко определены?
- О происхождении видов (1859 г.), глава 6. [7]
Эту дилемму можно охарактеризовать как отсутствие или редкость переходных разновидностей в пространстве обитания. [8]
Еще одна дилемма, [9] С первым связано отсутствие или редкость переходных разновидностей во времени. Дарвин указал, что согласно теории естественного отбора «должно было существовать бесчисленное количество переходных форм», и задался вопросом, «почему мы не находим их в бесчисленных количествах внедренными в земную кору». Тот факт, что четко определенные виды действительно существуют в природе как в пространстве, так и во времени, означает, что некая фундаментальная особенность естественного отбора действует для создания и поддержания видов. [7]
Влияние полового видообразование на размножения
Утверждалось, что решение первой дилеммы Дарвина заключается в том факте, что ауткроссинговое половое размножение имеет внутреннюю цену редкости. [10] [11] [12] [13] [14] Стоимость редкости возникает следующим образом. Если по градиенту ресурсов развивается большое количество отдельных видов, каждый из которых прекрасно приспособлен к очень узкой полосе этого градиента, каждый вид по необходимости будет состоять из очень небольшого числа представителей. Поиск партнера в таких обстоятельствах может представлять трудности, если многие особи по соседству принадлежат к другим видам. В этих условиях, если численность популяции какого-либо вида случайно увеличится (за счет того или иного соседнего вида, если среда насыщена), это немедленно облегчит его представителям поиск половых партнеров. . Представители соседних видов, численность популяций которых уменьшилась, испытывают большие трудности с поиском партнеров и поэтому реже образуют пары, чем более крупные виды. Это имеет эффект снежного кома: крупные виды растут за счет более мелких и редких видов, что в конечном итоге приводит к их исчезновению . В конце концов, остается лишь несколько видов, каждый из которых резко отличается от другого. [10] [11] [13] Цена редкости включает не только издержки невозможности найти партнера, но и косвенные затраты, такие как стоимость общения при поиске партнера при низкой плотности населения.

Редкость влечет за собой и другие затраты. Редкие и необычные особенности очень редко приносят пользу. В большинстве случаев они указывают на ( не молчащую ) мутацию , которая почти наверняка будет вредной. Поэтому половым существам следует избегать партнеров, обладающих редкими или необычными особенностями ( койнофилия ). [16] [17] Таким образом, половые популяции быстро теряют редкие или периферические фенотипические особенности, таким образом канализируя весь внешний вид, как показано на сопроводительном изображении африканского карликового зимородка Ispidina picta . Это единообразие всех взрослых представителей полового вида стимулировало распространение полевых справочников по птицам, млекопитающим, рептилиям, насекомым и многим другим таксонам , в которых вид может быть описан одной иллюстрацией (или двумя, в случае полового диморфизма ). Как только популяция станет однородной по внешнему виду, что типично для большинства видов (и это показано на фотографии африканского карликового зимородка), ее члены будут избегать спаривания с представителями других популяций, которые внешне отличаются от них самих. [18] Таким образом, избегание партнеров, демонстрирующих редкие и необычные фенотипические особенности, неизбежно приводит к репродуктивной изоляции, одной из отличительных черт видообразования. [19] [20] [21] [22]
В противоположном случае с организмами, размножающимися бесполым путем , редкость не требует платы; следовательно, мелкомасштабная адаптация приносит только пользу. Таким образом, бесполые организмы очень часто демонстрируют непрерывные изменения формы (часто во многих различных направлениях), которые Дарвин ожидал от эволюции, что делает их классификацию на «виды» (точнее, морфовиды ) очень трудной. [10] [16] [17] [23] [24] [25]
Режимы [ править ]

Все формы естественного видообразования имели место в ходе эволюции ; однако продолжаются споры относительно относительной важности каждого механизма в сохранении биоразнообразия . [26]
Одним из примеров естественного видообразования является разнообразие трехиглой колюшки , морской рыбы, которая после последнего ледникового периода претерпела видообразование в новые пресноводные колонии в изолированных озерах и ручьях. По оценкам, за 10 000 поколений колюшки демонстрируют структурные различия, которые больше, чем те, которые наблюдаются между различными родами рыб, включая различия в плавниках, изменения в количестве или размере их костных пластин, различное строение челюстей и различия в цвете. [27]
Аллопатрический [ править ]
Во время аллопатрического (от древнегреческого allos , «другой» + patrā , «отечество») видообразования популяция разделяется на две географически изолированные популяции (например, из-за фрагментации среды обитания из-за географических изменений, таких как горообразование ). Изолированные популяции затем подвергаются генотипической или фенотипической дивергенции, поскольку: (а) они подвергаются разнородному селективному давлению; (б) разные мутации в двух популяциях возникают . Когда популяции снова вступают в контакт, они эволюционировали так, что репродуктивно изолированы и больше не способны обмениваться генами . Островная генетика — это термин, связанный с тенденцией небольших изолированных генетических пулов к образованию необычных признаков. Примеры включают островную карликовость и радикальные изменения среди некоторых известных цепочек островов, например, на Комодо . Галапагосские острова особенно известны своим влиянием на Чарльза Дарвина. За пять недель пребывания там он услышал, что галапагосских черепах можно идентифицировать по островам, и заметил, что зяблики различались от одного острова к другому, но только девять месяцев спустя он подумал, что такие факты могут показать, что виды изменчивы. Когда он вернулся в Англию , его размышления об эволюции углубились после того, как эксперты сообщили ему, что это отдельные виды, а не просто разновидности, и, как известно, все другие различающиеся птицы Галапагосских островов были видами вьюрков. Хотя зяблики не имели для Дарвина особого значения, более поздние исследования показали, что птицы, ныне известные как зяблики Дарвина, представляют собой классический случай адаптивной эволюционной радиации. [28]
Перипатрический [ править ]
При перипатрическом видообразовании, подформе аллопатрического видообразования, новые виды образуются в изолированных, более мелких периферийных популяциях, которые не могут обмениваться генами с основной популяцией. Это связано с концепцией эффекта основателя , поскольку небольшие группы населения часто сталкиваются с узкими местами . Часто предполагается, что генетический дрейф играет значительную роль в перипатрическом видообразовании. [29] [30]
Тематические исследования включают исследование Майра фауны птиц; [31] австралийская птица Petroica multicolor ; [32] и репродуктивная изоляция в популяциях дрозофилы, подверженных ограничению популяции. [ нужна ссылка ]
Парапатрик [ править ]
При парапатрическом видообразовании существует лишь частичное разделение зон двух расходящихся популяций, обеспечиваемое географическими условиями; особи каждого вида могут время от времени вступать в контакт или пересекать места обитания, но снижение приспособленности гетерозигот приводит к отбору поведения или механизмов, которые предотвращают их скрещивание . Парапатрическое видообразование моделируется на основе непрерывных изменений внутри «единой», связанной среды обитания, действующей как источник естественного отбора, а не на эффектах изоляции сред обитания, возникающих в результате перипатрического и аллопатрического видообразования. [33]
, зависящим от ландшафта Парапатрическое видообразование может быть связано с дифференциальным отбором . Даже если между двумя популяциями существует поток генов , сильный дифференциальный отбор может препятствовать ассимиляции, и в конечном итоге могут развиться разные виды. [34] Различия в среде обитания могут иметь более важное значение для развития репродуктивной изоляции, чем время изоляции. Кавказские скальные ящерицы Darevskia rudis , D. валентини и D. portschinsky друг гибридизируются с другом в своей гибридной зоне ; однако гибридизация сильнее между D. portschinsky и D. rudis , которые отделились раньше, но обитают в сходных местообитаниях, чем между D. валентини и двумя другими видами, которые отделились позже, но обитают в разных климатических местообитаниях. [35]
Экологи обращаются к [ нужны разъяснения ] парапатрическое и перипатрическое видообразование по экологическим нишам . Для успеха нового вида должна быть доступна ниша. кольцевые виды, такие как чайки Larus, иллюстрируют происходящее видообразование, хотя ситуация может быть более сложной. Утверждается, что [36] Трава Anthoxanthum odoratum может начать парапатрическое видообразование в районах минного загрязнения. [37]
Симпатрический [ править ]

Симпатрическое видообразование — это образование двух или более видов-потомков от одного предкового вида, занимающих одно и то же географическое положение.
Часто цитируемые примеры симпатрического видообразования встречаются у насекомых, которые становятся зависимыми от разных растений- хозяев на одной и той же территории. [38] [39]
Самый известный пример симпатрического видообразования - это цихлиды Восточной Африки , населяющие озера Рифт-Валли , особенно озеро Виктория , озеро Малави и озеро Танганьика . Описано более 800 видов, а по оценкам, в регионе может существовать более 1600 видов. Их эволюция приводится как пример естественного и полового отбора . [40] [41] Исследование 2008 года предполагает, что симпатрическое видообразование произошло у пещерных саламандр Теннесси . [42] Симпатрическое видообразование, обусловленное экологическими факторами, может также объяснить необычайное разнообразие ракообразных, обитающих в глубинах сибирского озера Байкал . [43]
Почковающееся видообразование было предложено как особая форма симпатрического видообразования, при которой небольшие группы особей становятся все более изолированными от предков, преимущественно размножаясь друг с другом. Этот тип видообразования будет обусловлен сочетанием различных преимуществ инбридинга, таких как выражение выгодных рецессивных фенотипов, снижение рекомбинационной нагрузки и снижение стоимости пола. [44]

Боярышниковая муха ( Ragoletis pomonella ), также известная как яблочная личинка, по-видимому, подвергается симпатрическому видообразованию. [45] Разные популяции боярышников питаются разными плодами. Отдельная популяция возникла в Северной Америке в 19 веке, спустя некоторое время после того, как были завезены яблоки , неместный вид. Эта популяция, питающаяся яблоками, обычно питается только яблоками, а не исторически предпочитаемыми плодами боярышника . Нынешняя популяция, питающаяся боярышником, обычно не питается яблоками. Некоторые данные, например, что шесть из тринадцати аллозимных локусов различны, что боярышники созревают позже в течение сезона и для созревания требуется больше времени, чем яблонные мухи; и то, что доказательств скрещивания мало (исследователи зафиксировали уровень гибридизации 4–6%), позволяет предположить, что происходит симпатрическое видообразование. [46]
Методы отбора [ править ]
Армирование [ править ]

Подкрепление, также называемое эффектом Уоллеса , — это процесс, посредством которого естественный отбор увеличивает репродуктивную изоляцию. [19] Это может произойти после разделения двух популяций одного и того же вида, а затем повторного контакта. Если их репродуктивная изоляция была полной, то они уже развились в два отдельных несовместимых вида. Если их репродуктивная изоляция неполная, то дальнейшее спаривание между популяциями приведет к образованию гибридов, которые могут быть или не быть плодовитыми. Если гибриды бесплодны или плодовиты, но менее приспособлены, чем их предки, тогда произойдет дальнейшая репродуктивная изоляция и, по сути, произойдет видообразование, как у лошадей и ослов . [47]
Одна из причин этого заключается в том, что если каждый из родителей гибридного потомства естественным образом выбрал черты для своей определенной среды, гибридное потомство будет нести черты обоих, поэтому не будет соответствовать ни экологической нише, ни одному из родителей (экологическое видообразование). Низкая приспособленность гибридов приведет к тому, что отбор будет отдавать предпочтение ассортативному спариванию , которое будет контролировать гибридизацию. Иногда это называют эффектом Уоллеса в честь биолога-эволюциониста Альфреда Рассела Уоллеса , который в конце 19 века предположил, что это может быть важным фактором видообразования. [48] И наоборот, если гибридное потомство более приспособлено, чем их предки, тогда популяции снова сольются в один и тот же вид в пределах области, в которой они находятся в контакте. [ нужна ссылка ]
Другим важным теоретическим механизмом является возникновение внутренней генетической несовместимости, рассматриваемой в модели Бейтсона-Добжанского-Мюллера . [49] Гены из аллопатрических популяций будут иметь различное эволюционное происхождение и никогда не тестируются вместе до гибридизации при вторичном контакте, когда будут выявлены негативные эпистатические взаимодействия. Другими словами, новые аллели появятся в популяции и пройдут отбор только в том случае, если они хорошо работают вместе с другими генами в той же популяции, но они могут быть несовместимы с генами в аллопатрической популяции, будь то другие вновь полученные аллели или сохраненные. предковые аллели. Это обнаруживается только посредством новой гибридизации. [49] [50] Такая несовместимость приводит к снижению приспособленности гибридов независимо от экологической среды и, таким образом, является внутренней, хотя может возникнуть в результате адаптации к разным средам. [51] Накопление таких несовместимостей со временем увеличивается все быстрее и быстрее, создавая эффект «снежного кома». [52] Существует большое количество доказательств, подтверждающих эту теорию, в первую очередь из лабораторных популяций, таких как Drosophila и Mus , и были идентифицированы некоторые гены, участвующие в несовместимости. [50]
Подкрепление, благоприятствующее репродуктивной изоляции, необходимо как для парапатрического, так и для симпатрического видообразования. Без усиления географическая область контакта между различными формами одного и того же вида, называемая их «гибридной зоной», не превратится в границу между разными видами. Гибридные зоны — это регионы, где встречаются и скрещиваются различные популяции. В этих регионах распространено гибридное потомство, которое обычно создается разошедшимися видами, вступающими в вторичный контакт . Без подкрепления у этих двух видов возникло бы неконтролируемое инбридинг . [ нужна ссылка ] Подкрепление можно вызвать в экспериментах по искусственному отбору, как описано ниже.
Экологический [ править ]
Экологический отбор — это «взаимодействие особей с окружающей средой во время добычи ресурсов». [53] Естественный отбор по своей сути вовлечен в процесс видообразования, при котором «при экологическом видообразовании популяции в разных средах или популяции, эксплуатирующие разные ресурсы, испытывают контрастное давление естественного отбора на признаки, которые прямо или косвенно вызывают эволюцию репродуктивной изоляции». [54] Существуют доказательства роли экологии в процессе видообразования. Исследования популяций колюшки подтверждают экологически связанное видообразование, возникающее как побочный продукт. [55] наряду с многочисленными исследованиями параллельного видообразования, где изоляция развивается между независимыми популяциями видов, адаптирующимися к контрастным средам, а не между независимыми популяциями, адаптирующимися к сходным средам. [56] Экологическое видообразование происходит на основании большей части доказательств, «...накопленных в результате нисходящих исследований адаптации и репродуктивной изоляции». [56]
Половой отбор [ править ]
Половой отбор может стимулировать видообразование в кладе независимо от естественного отбора . [57] Однако термин «видообразование» в этом контексте имеет тенденцию использоваться в двух разных, но не взаимоисключающих смыслах. Первое и наиболее часто используемое значение относится к «рождению» новых видов. То есть разделение существующего вида на два отдельных вида или возникновение нового вида от родительского вида, и то, и другое обусловлено биологическим «модным увлечением» (предпочтением признака или признаков одного или обоих видов). пола, которые не обязательно обладают какими-либо адаптивными качествами). [57] [58] [59] [60] Во втором смысле «видообразование» относится к широко распространенной тенденции половых существ группироваться в четко определенные виды. [61] [20] вместо того, чтобы формировать континуум фенотипов как во времени, так и в пространстве, что было бы более очевидным или логичным следствием естественного отбора. Это действительно было признано Дарвином проблематичным и включено в его книгу «Происхождение видов » (1859 г.) под заголовком «Трудности с теорией». [7] Есть несколько предположений относительно того, как выбор партнера может сыграть значительную роль в решении дилеммы Дарвина . [20] [10] [16] [17] [18] [62] Если видообразование происходит в отсутствие естественного отбора, его можно назвать неэкологическим видообразованием . [63] [64]
Искусственное видообразование [ править ]


созданы новые виды Животноводством , но даты и способы зарождения таких видов не ясны. Часто домашний аналог все же может скрещиваться и производить плодовитое потомство со своим диким предком. Так обстоит дело с домашним скотом , которого можно считать тем же видом, что и некоторые разновидности дикого быка , гаура и яка ; и с домашними овцами , которые могут скрещиваться с муфлоном . [65] [66]
Наиболее документально подтвержденное создание новых видов в лаборатории было осуществлено в конце 1980-х годов. Уильям Р. Райс и Джордж У. Солт разводили Drosophila melanogaster, плодовых мух используя лабиринт с тремя различными вариантами среды обитания, такими как светлая/темная и влажная/сухая. Каждое поколение помещалось в лабиринт, и группы мух, вышедшие из двух из восьми выходов, были разделены для размножения друг с другом в соответствующих группах. Через тридцать пять поколений эти две группы и их потомство были репродуктивно изолированы из-за их сильных предпочтений в среде обитания: они спаривались только в пределах предпочитаемых ими территорий и поэтому не спаривались с мухами, предпочитавшими другие территории. [67] История таких попыток описана Райс и Элен Э. Хостерт (1993). [68] [69] Дайан Додд использовала лабораторный эксперимент, чтобы показать, как репродуктивная изоляция может развиваться у плодовых мух Drosophila pseudoobscura после нескольких поколений, помещая их в разные среды, среды на основе крахмала и мальтозы. [70]

Эксперимент Додда повторялся много раз, в том числе с другими видами плодовых мушек и пищевыми продуктами. [71] Такая быстрая эволюция репродуктивной изоляции иногда может быть следствием заражения бактериями Wolbachia . [72]
Альтернативное объяснение состоит в том, что эти наблюдения согласуются с тем, что животные, размножающиеся половым путем, по своей природе неохотно спариваются с особями, чья внешность или поведение отличаются от нормы. Риск того, что такие отклонения обусловлены наследственными дезадаптациями, высок. Таким образом, если животное, неспособное предсказать будущее направление естественного отбора, приучено производить наиболее приспособленное потомство, оно будет избегать партнеров с необычными привычками или особенностями. [73] [74] [16] [17] [18] Сексуальные существа неизбежно группируются в репродуктивно изолированные виды. [17]
Генетика [ править ]
Генов видообразования обнаружено немного. Обычно они включают процесс подкрепления на поздних стадиях видообразования. В 2008 году было сообщено о гене видообразования, вызывающем репродуктивную изоляцию. [75] Это вызывает гибридную стерильность между родственными подвидами. Порядок видообразования трех групп от общего предка может быть неясным или неизвестным; коллекция из трех таких видов называется «трихотомией». [ нужна ссылка ]
посредством полиплоидии Видообразование

Полиплоидия - это механизм, который вызвал множество быстрых событий видообразования при симпатрии , поскольку потомство, например, от спаривания тетраплоида с диплоидом часто приводит к триплоидному стерильному потомству. [76] Однако среди растений не все полиплоиды репродуктивно изолированы от своих родителей, и поток генов все же может происходить, например, посредством спариваний триплоидного гибрида и диплоида, которые производят тетраплоиды, или спаривания между мейотически нередуцированными гаметами диплоидов и гаметами тетраплоидов (см. также гибридные видообразование ). [ нужна ссылка ]
Было высказано предположение, что многие из существующих видов растений и большинства животных в своей эволюционной истории претерпели полиплоидизацию. [77] [78] Размножение успешных полиплоидных видов иногда бывает бесполым, путем партеногенеза или апомиксиса , поскольку по неизвестным причинам многие бесполые организмы являются полиплоидными. Известны редкие случаи полиплоидных млекопитающих, но чаще всего они приводят к внутриутробной смерти. [ нужна ссылка ]
Гибридный вид [ править ]
Гибридизация двух разных видов иногда приводит к различному фенотипу . Этот фенотип также может быть более приспособленным, чем родительская линия, и поэтому естественный отбор может тогда благоприятствовать этим особям. В конце концов, если будет достигнута репродуктивная изоляция, это может привести к появлению отдельного вида. Однако репродуктивную изоляцию между гибридами и их родителями достичь особенно трудно, и поэтому гибридное видообразование считается крайне редким явлением. произошла Считается, что Марианская кряква в результате гибридного видообразования. [ нужна ссылка ]
Гибридизация — важный способ видообразования у растений, поскольку полиплоидия (наличие более двух копий каждой хромосомы ) переносится растениями легче, чем животными. [79] [80] Полиплоидия важна для гибридов, поскольку она обеспечивает размножение, при этом каждый из двух разных наборов хромосом может образовывать пары с идентичным партнером во время мейоза. [78] Полиплоиды также обладают большим генетическим разнообразием, что позволяет им избегать инбридинговой депрессии в небольших популяциях. [81]
Гибридизация без изменения числа хромосом называется гомоплоидным гибридным видообразованием. Считается очень редким, но был обнаружен у Heliconius. бабочек [82] и подсолнухи . Полиплоидное видообразование, связанное с изменением числа хромосом, является более распространенным явлением, особенно у видов растений. [ нужна ссылка ]
Транспозиция генов [ править ]
Теодосий Добжанский , изучавший плодовых мух на заре генетических исследований в 1930-х годах, предположил, что части хромосом, которые переключаются из одного места в другое, могут привести к разделению вида на два разных вида. Он наметил, как участки хромосом могут перемещаться в геноме. Эти подвижные секции могут вызывать бесплодие у межвидовых гибридов, что может оказывать давление на видообразование. Теоретически его идея была здравой, но ученые долго спорили, действительно ли это произошло в природе. В конце концов было показано, что конкурирующая теория, предполагающая постепенное накопление мутаций, встречается в природе так часто, что генетики в значительной степени отвергли гипотезу движущегося гена. [83] Однако исследования 2006 года показывают, что переход гена с одной хромосомы на другую может способствовать рождению новых видов. [84] Это подтверждает механизм репродуктивной изоляции, ключевой компонент видообразования. [85]
Тарифы [ править ]
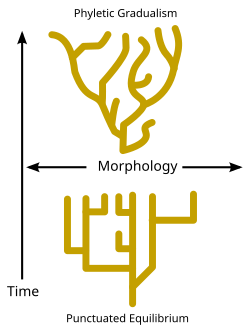
Ведутся споры относительно скорости, с которой события видообразования происходят в течение геологического времени. В то время как некоторые биологи-эволюционисты утверждают, что события видообразования оставались относительно постоянными и постепенными с течением времени (известные как «филетический градуализм» - см. Диаграмму), некоторые палеонтологи, такие как Найлс Элдридж и Стивен Джей Гулд, [86] утверждали, что виды обычно остаются неизменными в течение длительных периодов времени и что видообразование происходит только в течение относительно коротких интервалов времени. Эта точка зрения известна как прерывистое равновесие . (См. диаграмму и дилемму Дарвина .) [ нужна ссылка ]
эволюция Прерывистая
Эволюция может быть чрезвычайно быстрой, как показывает создание домашних животных и растений за очень короткий геологический промежуток времени, охватывающий всего несколько десятков тысяч лет. кукуруза ( Zea mays Например, ) возникла в Мексике всего за несколько тысяч лет, примерно 7000–12 000 лет назад. [87] Это поднимает вопрос о том, почему долгосрочная скорость эволюции намного медленнее, чем теоретически возможно. [88] [89] [90] [91]







Эволюция навязывается видам или группам. Это не планируется и не стремится к чему-то ламаркистскому . [92] Мутации, от которых зависит этот процесс, являются случайными событиями и, за исключением « тихих мутаций », не влияющих на функциональность или внешний вид носителя, таким образом, обычно невыгодны, и шансы на то, что они окажутся полезными в будущем, исчезают. маленький. Таким образом, хотя вид или группа могут выиграть от возможности адаптироваться к новой среде путем накопления широкого спектра генетических вариаций, это наносит ущерб особям, которые должны нести эти мутации до тех пор, пока в конечном итоге не появится небольшое, непредсказуемое меньшинство из них. способствует такой адаптации. Таким образом, способность к развитию потребует группового отбора — концепции, дискредитированной, например, Джорджем К. Уильямсом . [93] Джон Мейнард Смит [94] и Ричард Докинз [95] [96] [97] [98] как избирательно невыгодное для человека.
Таким образом, решение второй дилеммы Дарвина может быть следующим:
Если половые особи оказываются в невыгодном положении из-за передачи мутаций своему потомству, они будут избегать партнеров-мутантов со странными или необычными характеристиками. [74] [16] [17] [62] Мутации, влияющие на внешний вид их носителей, тогда редко будут передаваться следующему и последующим поколениям. Поэтому они редко подвергались проверке естественным отбором. Таким образом, эволюция фактически остановлена или значительно замедлена. С точки зрения прерывистого равновесия, единственные мутации, которые могут накапливаться в популяции, — это те, которые не оказывают заметного влияния на внешний вид и функциональность их носителей (т. е. это «тихие» или « нейтральные мутации », которые могут быть и используются для отслеживания родства и возраста популяций и видов . [16] [99] )
Этот аргумент подразумевает, что эволюция может произойти только в том случае, если нельзя избежать партнеров-мутантов из-за серьезной нехватки потенциальных партнеров. Чаще всего это происходит в небольших изолированных сообществах . Чаще всего они встречаются на небольших островах, в отдаленных долинах, озерах, речных системах или пещерах. [100] или во время последствий массового вымирания . [99] В этих обстоятельствах не только выбор партнеров строго ограничен, но и узкие места в популяции, эффекты основателя, генетический дрейф и инбридинг вызывают быстрые, случайные изменения в генетическом составе изолированной популяции. [100] Более того, гибридизация с родственными видами, попавшими в один и тот же изолят, может привести к дополнительным генетическим изменениям. Если изолированная популяция, такая как эта, переживет свои генетические потрясения и впоследствии расширится в незанятую нишу или в нишу, в которой она имеет преимущество перед своими конкурентами, возникнет новый вид или подвид. С геологической точки зрения это будет внезапное событие. Возобновление избегания партнеров-мутантов впоследствии снова приведет к эволюционному застою. [86] [89]
В явном подтверждении этого прерывистого равновесного взгляда на эволюцию летопись окаменелостей эволюционного прогресса обычно состоит из видов, которые внезапно появляются и в конечном итоге исчезают сотни тысяч или миллионы лет спустя без каких-либо изменений во внешнем виде. [86] [99] [101] Графически эти ископаемые виды представлены линиями, параллельными оси времени, длина которых показывает, как долго существовал каждый из них. Тот факт, что линии остаются параллельными оси времени, иллюстрирует неизменный внешний вид каждого из ископаемых видов, изображенных на графике. В ходе существования каждого вида новые виды появляются через случайные промежутки времени, каждый из которых также существует в течение многих сотен тысяч лет, прежде чем исчезнуть без изменения внешнего вида. Точное родство этих одновременных видов обычно невозможно определить. Это иллюстрируется диаграммой, изображающей распределение видов гомининов во времени с тех пор, как гоминиды отделились от линии, которая привела к эволюции их ближайших ныне живущих родственников-приматов, шимпанзе. [101]
Похожие временные линии эволюции см., например, в палеонтологическом списке африканских динозавров , азиатских динозавров , Lampriformes и Amiiformes . [ нужна ссылка ]
См. также [ править ]
Ссылки [ править ]
- ^ Берлохер 1998 , стр. 3.
- ^ Кук, Оратор Ф. (30 марта 1906 г.). «Факторы видообразования» . Наука . 23 (587): 506–507. Бибкод : 1906Sci....23..506C . дои : 10.1126/science.23.587.506 . ПМИД 17789700 .
- ^ Кук, оратор Ф. (ноябрь 1908 г.). «Эволюция без изоляции». Американский натуралист . 42 (503): 727–731. дои : 10.1086/279001 . S2CID 84565616 .
- ^ Виа, Сара (16 июня 2009 г.). «Естественный отбор в действии при видообразовании» . ПНАС . 106 (Приложение 1): 9939–9946. Бибкод : 2009PNAS..106.9939V . дои : 10.1073/pnas.0901397106 . ПМК 2702801 . ПМИД 19528641 .
- ^ Шнайдер, Кристофер Дж. (31 октября 2000 г.). «Естественный отбор и видообразование» . Труды Национальной академии наук . 97 (23): 12398–12399. Бибкод : 2000PNAS...9712398S . дои : 10.1073/pnas.240463297 . ПМК 34057 . ПМИД 11058173 .
- ^ Майр 1982 , с. 273
- ↑ Перейти обратно: Перейти обратно: а б с д Дарвин 1859 г.
- ^ Сепкоски, Дэвид (2012). «1. Дилемма Дарвина: палеонтология, летопись окаменелостей и эволюционная теория» . Перечитывая летопись окаменелостей: развитие палеобиологии как эволюционной дисциплины . Издательство Чикагского университета. стр. 9–50. ISBN 978-0-226-74858-0 .
Одно из его самых больших опасений заключалось в том, что «неполнота» летописи окаменелостей будет использована для критики его теории: что очевидные «пробелы» в последовательности окаменелостей могут быть названы, по крайней мере, отрицательным свидетельством его предположения о том, что все организмы произошли от общего предка путем мелких и постепенных изменений.
- ^ Стоуэр, Ханна (2013). «Разрешение дилеммы Дарвина» . Обзоры природы Генетика . 14 (747): 747. дои : 10.1038/nrg3614 . S2CID 45302603 .
Почти одновременное появление строения тела большинства современных животных во время кембрийского взрыва предполагает короткий период быстрой фенотипической и генетической эволюции, которая, по мнению Дарвина, была слишком быстрой, чтобы ее можно было объяснить естественным отбором.
- ↑ Перейти обратно: Перейти обратно: а б с д Бернштейн, Харрис; Байерли, Генри К.; Хопф, Фредерик А.; и др. (21 декабря 1985 г.). «Секс и возникновение видов». Журнал теоретической биологии . 117 (4): 665–690. Бибкод : 1985JThBi.117..665B . дои : 10.1016/S0022-5193(85)80246-0 . ПМИД 4094459 .
- ↑ Перейти обратно: Перейти обратно: а б Хопф, Фредерик А.; Хопф, Ф.В. (февраль 1985 г.). «Роль эффекта Алли в упаковке видов». Теоретическая популяционная биология . 27 (1): 27–50. дои : 10.1016/0040-5809(85)90014-0 .
- ^ Бернштейн и Бернштейн 1991
- ↑ Перейти обратно: Перейти обратно: а б Мишо 1995 г.
- ^ Мишо 1999
- ^ Хоккей, Дин и Райан 2005 , стр. 176, 193.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Коэслаг, Йохан Х. (10 мая 1990 г.). «Койнофилия группирует сексуальные существа в виды, способствует застою и стабилизирует социальное поведение». Журнал теоретической биологии . 144 (1): 15–35. Бибкод : 1990JThBi.144...15K . дои : 10.1016/s0022-5193(05)80297-8 . ISSN 0022-5193 . ПМИД 2200930 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Коэслаг, Йохан Х. (21 декабря 1995 г.). «О машине видообразования». Журнал теоретической биологии . 177 (4): 401–409. Бибкод : 1995JThBi.177..401K . дои : 10.1006/jtbi.1995.0256 . ISSN 0022-5193 .
- ↑ Перейти обратно: Перейти обратно: а б с Поэльстра, Джелмер В.; Виджай, Нагарджун; Боссу, Кристен М.; и др. (20 июня 2014 г.). «Геномный ландшафт, лежащий в основе фенотипической целостности перед лицом потока генов у ворон» . Наука . 344 (6190): 1410–1414. Бибкод : 2014Sci...344.1410P . дои : 10.1126/science.1253226 . ПМИД 24948738 . S2CID 14431499 .
Фенотипические различия между воронами и воронами в зоне гибридизации в Европе вряд ли обусловлены ассортативным спариванием.
- Комментарий Мажуванчерри К. Унникришнана и Х.С. Ахилы. - ↑ Перейти обратно: Перейти обратно: а б Ридли, Марк . «Видообразование. Какова роль подкрепления в видообразовании?» . Проверено 7 сентября 2015 г. Адаптировано из книги «Эволюция» (2004 г.), 3-е издание (Малден, Массачусетс: Blackwell Publishing ), ISBN 978-1-4051-0345-9 .
- ↑ Перейти обратно: Перейти обратно: а б с Мейнард Смит 1989 , стр. 275–280.
- ^ Майр 1988 г.
- ^ Уильямс 1992 , с. 118
- ^ Мейнард Смит, Джон (декабрь 1983 г.). «Генетика стазиса и пунктуации» (PDF) . Ежегодный обзор генетики . 17 :11–25. дои : 10.1146/annurev.ge.17.120183.000303 . ПМИД 6364957 . S2CID 3901837 . Архивировано из оригинала (PDF) 5 марта 2019 г.
- ^ Клэпхэм, Тутин и Варбург, 1952 г.
- ^ Грант 1971
- ^ Бейкер, Джейсон М. (июнь 2005 г.). «Адаптивное видообразование: роль естественного отбора в механизмах географического и негеографического видообразования» (PDF) . Исследования по истории и философии науки. Часть C: Исследования по истории и философии биологических и биомедицинских наук . 36 (2): 303–326. дои : 10.1016/j.shpsc.2005.03.005 . ПМИД 19260194 . S2CID 3555049 .
- ^ Кингсли, Дэвид М. (январь 2009 г.). «Раскрытое разнообразие: от атомов к чертам». Научный американец . 300 (1): 52–59. doi : 10.1038/scientificamerican0109-52 . ПМИД 19186749 .
- ^ Саллоуэй, Фрэнк Дж. (30 сентября 1982 г.). « Коллекции дарвиновских вьюрков (Geospizinae)» в составе гончей . Бюллетень Британского музея (Естественная история), Зоология . 43 (2): 49–58.
- ^ Койн и Орр 2004 , с. 105.
- ^ Лоусон, Люсинда П.; Бейтс, Джон М.; Менегон, Мишель; Погрузчик, Саймон П. (2015). «Расхождение по краям: перипатрическая изоляция в горном комплексе колючих тростниковых лягушек» . Эволюционная биология BMC . 15 (128): 128. Бибкод : 2015BMCEE..15..128L . дои : 10.1186/s12862-015-0384-3 . ПМЦ 4487588 . ПМИД 26126573 .
- ^ Майр 1992 , стр. 21–53.
- ^ Токеши, М. (1999). Сосуществование видов: экологические и эволюционные перспективы . Оксфорд: Блэквелл Сайенс. ISBN 0632061464 . OCLC 47011551 .
- ^ «Видообразование: происхождение новых видов | Изучайте науку в Scitable» . Природа . Проверено 16 февраля 2020 г.
- ^ Эндлер 1977
- ^ Тархнишвили, Давид; Мурцхваладзе, морской пехотинец; Гавашелишвили, Александр (август 2013 г.). «Видообразование у кавказских ящериц: климатическое несходство местообитаний важнее времени изоляции» . Биологический журнал Линнеевского общества . 109 (4): 876–892. дои : 10.1111/bij.12092 .
- ^ Либерс, Дорит; Книфф, Питер де; Хельбиг, Андреас Дж. (2004). «Комплекс серебристых чаек не является кольцевым видом» . Учебник по биологическим наукам . 271 (1542): 893–901. дои : 10.1098/rspb.2004.2679 . ПМК 1691675 . ПМИД 15255043 .
- ^ «Парапатрическое видообразование» . Калифорнийский университет в Беркли . Проверено 3 апреля 2017 г.
- ^ Федер, Джеффри Л.; Сяньфа Се; Рулль, Хуан; и др. (3 мая 2005 г.). «Майр, Добжанский и Буш и сложности симпатрического видообразования у Rhagoletis » . ПНАС . 102 (Приложение 1): 6573–6580. Бибкод : 2005PNAS..102.6573F . дои : 10.1073/pnas.0502099102 . ПМЦ 1131876 . ПМИД 15851672 .
- ^ Берлохер, Стюарт Х.; Федер, Джеффри Л. (январь 2002 г.). «Симпатрическое видообразование у насекомых-фитофагов: выход за рамки противоречий?». Ежегодный обзор энтомологии . 47 : 773–815. дои : 10.1146/annurev.ento.47.091201.145312 . ПМИД 11729091 . S2CID 9677456 .
- ^ Мачадо, Хизер Э.; Пыльца, Александр А.; Хофманн, Ганс А.; и др. (декабрь 2009 г.). «Межвидовое профилирование экспрессии генов на основе сравнительной геномной гибридизации: обзор и новый подход к африканским цихлидам» . Интегративная и сравнительная биология . 49 (6): 644–659. дои : 10.1093/icb/icp080 . ПМИД 21665847 .
- ^ Фань, Шаохуа; Элмер, Кэтрин Р.; Мейер, Аксель (5 февраля 2012 г.). «Геномика адаптации и видообразования цихлид: последние достижения и анализ африканских и неотропических линий» . Философские труды Королевского общества Б. 367 (1587): 385–394. дои : 10.1098/rstb.2011.0247 . ПМЦ 3233715 . ПМИД 22201168 .
- ^ Нимиллер, Мэтью Л.; Фитцпатрик, Бенджамин М.; Миллер, Брайан Т. (май 2008 г.). «Недавнее расхождение с потоком генов у пещерных саламандр Теннесси (Plethodontidae: Gyrinophilus ), выведенное из генеалогии» . Молекулярная экология . 17 (9): 2258–2275. Бибкод : 2008MolEc..17.2258N . дои : 10.1111/j.1365-294X.2008.03750.x . ПМИД 18410292 . S2CID 20761880 .
- ^ Мартенс, Коэн (май 1997 г.). «Видообразование в древних озерах». Тенденции в экологии и эволюции . 12 (5): 177–182. дои : 10.1016/S0169-5347(97)01039-2 . ПМИД 21238028 .
- ^ Жоли, Э. (9 декабря 2011 г.). «Существование видов опирается на метастабильное равновесие между инбридингом и аутбридингом. Эссе о тесной связи между видообразованием, инбридингом и рецессивными мутациями» . Биология Директ . 6:62 . дои : 10.1186/1745-6150-6-62 . ПМК 3275546 . ПМИД 22152499 .
- ^ Федер, Джеффри Л.; Рётеле, Джозеф Б.; Филчак, Кеннет; и др. (март 2003 г.). «Доказательства инверсионного полиморфизма, связанные с формированием симпатрической расы хозяина у яблонной личинки Rhagoletis pomonella » . Генетика . 163 (3): 939–953. дои : 10.1093/генетика/163.3.939 . ПМЦ 1462491 . ПМИД 12663534 . Проверено 7 сентября 2015 г.
- ^ Берлохер, Стюарт Х.; Буш, Гай Л. (июнь 1982 г.). «Электрофоретический анализ филогении Rhagoletis (Diptera: Tephritidae)». Систематическая зоология . 31 (2): 136–155. дои : 10.2307/2413033 . JSTOR 2413033 .
- ^ Сэтре, Гленн-Питер (2012). «Укрепление». ЭЛС . дои : 10.1002/9780470015902.a0001754.pub3 . ISBN 978-0470016176 .
- ^ Оллертон, Джефф (сентябрь 2005 г.). «Видообразование: время цветения и эффект Уоллеса» (PDF) . Наследственность . 95 (3): 181–182. дои : 10.1038/sj.hdy.6800718 . ПМИД 16077739 . S2CID 13300641 . Архивировано из оригинала (PDF) 5 июня 2007 г. Проверено 7 сентября 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б Орр, HA (декабрь 1996 г.). «Добжанский, Бейтсон и генетика видообразования» . Генетика . 144 (4): 1331–1335. дои : 10.1093/генетика/144.4.1331 . ISSN 0016-6731 . ПМК 1207686 . ПМИД 8978022 .
- ↑ Перейти обратно: Перейти обратно: а б Прегрейвс, Дэйвен К. (декабрь 2010 г.). «Дарвин и происхождение межвидовой генетической несовместимости» . Американский натуралист . 176 (С1): С45–С60. дои : 10.1086/657058 . ISSN 0003-0147 . ПМИД 21043780 . S2CID 5592958 .
- ^ Кулмуни, Дж.; Вестрам, AM (2017). «Внутренняя несовместимость, развивающаяся как побочный продукт дивергентного экологического отбора: рассмотрение их в эмпирических исследованиях дивергенции с потоком генов» . Молекулярная экология . 26 (12): 3093–3103. Бибкод : 2017MolEc..26.3093K . дои : 10.1111/mec.14147 . ISSN 1365-294X . ПМИД 28423210 . S2CID 41904934 .
- ^ Орр, ХА (1 апреля 1995 г.). «Популяционная генетика видообразования: эволюция гибридной несовместимости» . Генетика . 139 (4): 1805–1813. дои : 10.1093/генетика/139.4.1805 . ISSN 1943-2631 . ПМК 1206504 . ПМИД 7789779 .
- ^ Ховард Д. Рандл и Патрик Носил (2005), «Экологическое видообразование», Ecology Letters , 8 (3): 336–352, Бибкод : 2005EcolL...8..336R , doi : 10.1111/j.1461-0248.2004.00715 .x
- ^ Дольф Шлютер (2001), «Экология и происхождение видов», Тенденции в экологии и эволюции , 16 (7): 372–380, doi : 10.1016/S0169-5347(01)02198-X , PMID 11403870 , S2CID 9845298
- ^ Джеффри С. Маккиннон; и др. (2004), «Доказательства роли экологии в видообразовании», Nature , 429 (6989): 294–298, Bibcode : 2004Natur.429..294M , doi : 10.1038/nature02556 , PMID 15152252 , S2CID 2744267
- ↑ Перейти обратно: Перейти обратно: а б Дольф Шлютер (2009), «Свидетельства экологического видообразования и его альтернативы», Science , 326 (5915): 737–740, Бибкод : 2009Sci...323..737S , doi : 10.1126/science.1160006 , PMID 19197053 , S2CID 307207
- ↑ Перейти обратно: Перейти обратно: а б Панхейс, Тами М.; Батлин, Роджер; Зук, Марлен ; и др. (июль 2001 г.). «Половой отбор и видообразование» (PDF) . Тенденции в экологии и эволюции . 16 (7): 364–371. дои : 10.1016/s0169-5347(01)02160-7 . ПМИД 11403869 .
- ^ Дарвин, Чарльз; А. Р. Уоллес (1858 г.). «О склонности видов к образованию разновидностей; и о сохранении разновидностей и видов естественными средствами отбора» (PDF) . Журнал трудов Лондонского Линнеевского общества. Зоология . 3 (9): 46–50. дои : 10.1111/j.1096-3642.1858.tb02500.x .
- ^ Дарвин 1859 , с. 89, «IV. Естественный отбор» .
- ^ Эберхард, WG (1985). Половой отбор и гениталии животных. Издательство Гарвардского университета, Кембридж, Массачусетс
- ^ Гулд 1980 , стр. 204–213 , «Куахог - это Куахог».
- ↑ Перейти обратно: Перейти обратно: а б Миллер 2013 , стр. 177, 395–396.
- ^ Рунделл, Ребекка Дж.; Прайс, Тревор Д. (1 июля 2009 г.). «Адаптивная радиация, неадаптивная радиация, экологическое видообразование и неэкологическое видообразование» . Тенденции в экологии и эволюции . 24 (7): 394–399. дои : 10.1016/j.tree.2009.02.007 . ISSN 0169-5347 . ПМИД 19409647 .
- ^ Чекански-Мойр, Джесси Э.; Рунделл, Ребекка Дж. (01 мая 2019 г.). «Экология неэкологического видообразования и неадаптивных излучений» . Тенденции в экологии и эволюции . 34 (5): 400–415. дои : 10.1016/j.tree.2019.01.012 . ISSN 0169-5347 . ПМИД 30824193 . S2CID 73494468 .
- ^ Новак 1999 г.
- ^ Хиндледер, Стефан; Каупе, Бернхард; Васмут, Рудольф; и др. (7 мая 2002 г.). «Молекулярный анализ диких и домашних овец ставит под сомнение текущую номенклатуру и предоставляет доказательства одомашнивания двух разных подвидов» . Труды Королевского общества Б. 269 (1494): 893–904. дои : 10.1098/рспб.2002.1975 . ПМК 1690972 . ПМИД 12028771 .
- ^ Райс, Уильям Р.; Солт, Джордж У. (июнь 1988 г.). «Видообразование посредством разрушительного отбора в зависимости от предпочтений среды обитания: экспериментальные данные». Американский натуралист . 131 (6): 911–917. дои : 10.1086/284831 . S2CID 84876223 .
- ^ Райс, Уильям Р.; Хостерт, Эллен Э. (декабрь 1993 г.). «Лабораторные эксперименты по видообразованию: чему мы научились за 40 лет?». Эволюция . 47 (6): 1637–1653. дои : 10.2307/2410209 . JSTOR 2410209 . ПМИД 28568007 .
- ^ Гаврилец, Сергей (октябрь 2003 г.). «Перспектива: модели видообразования: чему мы научились за 40 лет?». Эволюция . 57 (10): 2197–2215. дои : 10.1554/02-727 . ПМИД 14628909 . S2CID 198158082 .
- ^ Додд, Дайан М.Б. (сентябрь 1989 г.). «Репродуктивная изоляция как следствие адаптивной дивергенции у Drosophila pseudoobscura ». Эволюция . 43 (6): 1308–1311. дои : 10.2307/2409365 . JSTOR 2409365 . ПМИД 28564510 .
- ^ Киркпатрик, Марк; Равинье, Вирджиния (март 2002 г.). «Видообразование путем естественного и полового отбора: модели и эксперименты». Американский натуралист . 159 (С3): С22–С35. дои : 10.1086/338370 . ISSN 0003-0147 . ПМИД 18707367 . S2CID 16516804 .
- ^ Куку, Катерина; Павликаки, Харис; Килиас, Джордж; и др. (январь 2006 г.). «Влияние лечения антибиотиками и лечения Wolbachia на сексуальную изоляцию среди клеточных популяций Drosophila melanogaster ». Эволюция . 60 (1): 87–96. дои : 10.1554/05-374.1 . ПМИД 16568634 . S2CID 198153238 .
- ^ Саймонс 1979
- ↑ Перейти обратно: Перейти обратно: а б Ланглуа, Джудит Х.; Роггман, Лори А. (март 1990 г.). «Привлекательные лица бывают только средними». Психологическая наука . 1 (2): 115–121. дои : 10.1111/j.1467-9280.1990.tb00079.x . S2CID 18557871 .
- ^ Фаднис, Нитин; Орр, Х. Аллен (16 января 2009 г.). «Один ген вызывает как мужскую стерильность, так и искажение сегрегации у дрозофилы гибридов » . Наука . 323 (5912): 376–379. Бибкод : 2009Sci...323..376P . дои : 10.1126/science.1163934 . ПМК 2628965 . ПМИД 19074311 .
- ^ Рэмси, Джастин; Шемске, Дуглас В. (ноябрь 1998 г.). «Пути, механизмы и скорость образования полиплоидов у цветковых растений» (PDF) . Ежегодный обзор экологии и систематики . 29 : 467–501. дои : 10.1146/annurev.ecolsys.29.1.467 . S2CID 31637733 . Архивировано из оригинала (PDF) 8 июня 2020 г.
- ^ Отто, Сара П .; Уиттон, Жаннетт (декабрь 2000 г.). «Распространенность и эволюция полиплоидов» (PDF) . Ежегодный обзор генетики . 34 : 401–437. CiteSeerX 10.1.1.323.1059 . дои : 10.1146/annurev.genet.34.1.401 . ПМИД 11092833 .
- ↑ Перейти обратно: Перейти обратно: а б Комай, Лука (ноябрь 2005 г.). «Преимущества и недостатки полиплоида». Обзоры природы Генетика . 6 (11): 836–846. дои : 10.1038/nrg1711 . ПМИД 16304599 . S2CID 3329282 .
- ^ Вендел, Джонатан Ф. (январь 2000 г.). «Эволюция генома полиплоидов». Молекулярная биология растений . 42 (1): 225–249. дои : 10.1023/A:1006392424384 . ПМИД 10688139 . S2CID 14856314 .
- ^ Семон, Мари; Вулф, Кеннет Х. (декабрь 2007 г.). «Последствия дупликации генома». Текущее мнение в области генетики и развития . 17 (6): 505–512. дои : 10.1016/j.gde.2007.09.007 . ПМИД 18006297 .
- ^ Солтис, Памела С .; Солтис, Дуглас Э. (20 июня 2000 г.). «Роль генетических и геномных признаков в успехе полиплоидов» . ПНАС . 97 (13): 7051–7057. Бибкод : 2000PNAS...97.7051S . дои : 10.1073/pnas.97.13.7051 . ПМК 34383 . ПМИД 10860970 .
- ^ Маварес, Хесус; Салазар, Камило А.; Бермингем, Элдридж; и др. (15 июня 2006 г.). «Видообразование путем гибридизации бабочек Heliconius ». Природа . 441 (7095): 868–871. Бибкод : 2006Natur.441..868M . дои : 10.1038/nature04738 . ПМИД 16778888 . S2CID 2457445 .
- ^ Шервуд, Джонатан (8 сентября 2006 г.). «Генетический сюрприз подтверждает забытую 70-летнюю эволюционную теорию» (пресс-релиз). Университет Рочестера . Проверено 10 сентября 2015 г.
- ^ Масли, Джон П.; Джонс, Корбин Д.; Мохамед, А. Ф. Нур ; и др. (8 сентября 2006 г.). «Транспозиция генов как причина гибридной стерильности у дрозофилы ». Наука . 313 (5792): 1448–1450. Бибкод : 2006Sci...313.1448M . дои : 10.1126/science.1128721 . ПМИД 16960009 . S2CID 23462115 .
- ^ Минкель-младший (8 сентября 2006 г.). «Ген странствующей мухи поддерживает новую модель видообразования» . Научный американец . Проверено 11 сентября 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б с Гулд, Стивен Джей; Элдридж, Найлз (весна 1977 г.). «Периодическое равновесие: пересмотренный темп и способ эволюции» (PDF) . Палеобиология . 3 (2): 115–151. Бибкод : 1977Pbio....3..115G . дои : 10.1017/s0094837300005224 . JSTOR 2400177 . S2CID 83492071 . Архивировано из оригинала (PDF) 24 июня 2014 г. Проверено 15 сентября 2015 г.
- ^ Законы 2010 г. , стр. 210–215.
- ^ Уильямс 1992 , гл. 9
- ↑ Перейти обратно: Перейти обратно: а б Элдридж и Гулд, 1972 , гл. 5
- ^ Майр 1954 , стр. 157–180.
- ^ Мейнард Смит 1989 , с. 281
- ^ Гулд 1980 , стр. 4, гл. 18
- ^ Уильямс 1974
- ^ Мейнард Смит, Джон (14 марта 1964 г.). «Групповой отбор и родственный отбор». Природа . 201 (4924): 1145–1147. Бибкод : 1964Natur.201.1145S . дои : 10.1038/2011145a0 . S2CID 4177102 .
- ^ Докинз 1995 , гл. 4
- ^ Докинз, Ричард (декабрь 1994 г.). «Захоронение автомобиля» . Поведенческие и мозговые науки . 17 (4): 616–617. дои : 10.1017/S0140525X00036207 . ISSN 0140-525X . S2CID 143378724 . Архивировано из оригинала 15 сентября 2006 г. Проверено 15 сентября 2015 г. «Замечания к более ранней статье [Эллиота] Собера [так в оригинале] и Дэвида Слоана Уилсона , которые представили более развернутые аргументы в своей недавней книге «Другим: эволюция и психология бескорыстного поведения ».
- ^ Деннетт, Дэниел К. (декабрь 1994 г.). «Е Pluribus Unum?» . Поведенческие и мозговые науки . 17 (4): 617–618. дои : 10.1017/S0140525X00036219 . S2CID 146359497 . Архивировано из оригинала 27 декабря 2007 г. «Комментарий к книге Wilson & Sober: выбор группы».
- ^ Пинкер, Стивен (18 июня 2012 г.). «Ложная привлекательность группового отбора» . Edge.org . Edge Foundation, Inc. Проверено 15 сентября 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б с Кэмпбелл 1990 , стр. 450–451, 487–490, 499–501.
- ↑ Перейти обратно: Перейти обратно: а б Аяла 1982 , стр. 73–83, 182–190, 198–215.
- ↑ Перейти обратно: Перейти обратно: а б Маккарти и Рубидж, 2005 г.
Библиография [ править ]
- Аяла, Франсиско Дж . (1982). Популяционная и эволюционная генетика . Серия Бенджамина/Каммингса по наукам о жизни. Менло-Парк, Калифорния: Паб Benjamin/Cummings. компании ISBN 978-0-8053-0315-5 . LCCN 81021623 . OCLC 8034790 .
- Берлохер, Стюарт Х. (1998). «Происхождение: краткая история исследований видообразования» . В Ховарде, Дэниел Дж.; Берлохер, Стюарт Х. (ред.). Бесконечные формы: виды и видообразование . Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-510901-6 . LCCN 97031461 . OCLC 37545522 .
- Бернштейн, Кэрол; Бернштейн, Харрис (1991). Старение, пол и восстановление ДНК . Сан-Диего, Калифорния: Academic Press . ISBN 978-0-12-092860-6 . LCCN 90014467 . OCLC 22542921 .
- Кэмпбелл, Нил А. (1990). Биология (2-е изд.). Редвуд-Сити, Калифорния: Паб Benjamin/Cummings. компании ISBN 978-0-8053-1800-5 . LCCN 89017952 . OCLC 20352649 .
- Клэпхэм, Артур Рой ; Тутин, Томас Г .; Варбург, Эдмунд Ф. (1952). Флора Британских островов . Кембридж, Великобритания: Издательство Кембриджского университета . LCCN 52008880 . ОСЛК 1084058 .
- Койн, Джерри А .; Орр, Х. Аллен (2004). Видообразование . Сандерлендс, Массачусетс: Sinauer Associates . ISBN 978-0-87893-089-0 . LCCN 2004009505 . OCLC 55078441 .
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей . LCCN 06017473 . OCLC 741260650 . Книга доступна по адресу Джон ван Вай, изд. (2002), Полное собрание сочинений Чарльза Дарвина в Интернете , получено 12 сентября 2015 г.
- Докинз, Ричард (1995). Река из Эдема: дарвиновский взгляд на жизнь . Серия «Мастера естественных наук». Нью-Йорк: Основные книги . ISBN 978-0-465-01606-8 . LCCN 94037146 . OCLC 31376584 .
- Элдридж, Найлс ; Гулд, Стивен Джей (1972). «Периодическое равновесие: альтернатива филетическому градуализму». В Шопфе, Томас Дж. М. (ред.). Модели в палеобиологии . Сан-Франциско, Калифорния: ISBN Freeman Cooper & Co. 978-0-87735-325-6 . LCCN 72078387 . OCLC 572084 . Перепечатано в Элдридже, 1985 г. , стр. 193–223.
- Элдридж, Найлз (1985). Временные рамки: переосмысление дарвиновской эволюции и теории прерывистого равновесия . Нью-Йорк: Саймон и Шустер . ISBN 978-0-671-49555-8 . LCCN 84023632 . OCLC 11443805 .
- Эндлер, Джон А. (1977). Географическая изменчивость, видообразование и клины . Монографии по популяционной биологии. Том. 10. Принстон, Нью-Джерси: Издательство Принстонского университета . стр. 1–246. ISBN 978-0-691-08187-8 . LCCN 76045896 . ОСЛК 2645720 . ПМИД 409931 .
- Гулд, Стивен Джей (1980). Большой палец панды: больше размышлений о естествознании (1-е изд.). Нью-Йорк: WW Norton & Company . ISBN 978-0-393-01380-1 . LCCN 80015952 . OCLC 6331415 . Издание 1982 года через Archive.org.
- Грант, Верн (1971). Вид растения . Нью-Йорк: Издательство Колумбийского университета . ISBN 978-0-231-03208-7 . LCCN 75125620 . OCLC 139834 .
- Хоккей, Фил А.Р.; Дин, В. Ричард Дж.; Райан, Питер Г., ред. (2005). Робертс Бердс из Южной Африки (7-е изд.). Кейптаун, Южная Африка: Попечители Книжного фонда Дж. Фелькера. ISBN 978-0-620-34053-3 . LCCN 2006376728 . OCLC 65978899 .
- Законы, Билл (2010). Пятьдесят растений, изменивших ход истории . Буффало, Нью-Йорк: Firefly Books. ISBN 978-1-55407-798-4 . LCCN 2011414731 . OCLC 711609823 .
- Мейнард Смит, Джон (1989). Эволюционная генетика . Оксфорд; Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-854215-5 . LCCN 88017041 . OCLC 18069049 .
- Майр, Эрнст (1954). «Изменение генетической среды и эволюция». В Хаксли, Джулиан ; Харди, Алистер С .; Форд, Эдмунд Б. (ред.). Эволюция как процесс . Лондон: Аллен и Анвин . LCCN 54001781 . OCLC 974739 .
- Майр, Эрнст (1982). Рост биологической мысли: разнообразие, эволюция и наследование . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета . ISBN 978-0-674-36445-5 . LCCN 81013204 . OCLC 7875904 .
- Майр, Эрнст (1988). К новой философии биологии: наблюдения эволюциониста . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета. ISBN 978-0-674-89665-9 . LCCN 87031892 . ОСЛК 17108004 .
- Майр, Эрнст (1992). «Специальная эволюция или прерывистое равновесие». В Сомите, Альберт; Петерсон, Стивен А. (ред.). Динамика эволюции: споры о прерывистом равновесии в естественных и социальных науках . Итака, Нью-Йорк: Издательство Корнельского университета . ISBN 978-0-8014-9763-6 . LCCN 91055569 . ОСЛК 24374091 .
- Маккарти, Теренс; Рубидж, Брюс (2005). История Земли и жизни: взгляд на путешествие длиной в 4,6 миллиарда лет из Южной Африки . Кейптаун, Южная Африка: Struik Publishers. ISBN 978-1-77007-148-3 . LCCN 2006376206 . OCLC 62098231 .
- Мишод, Ричард Э. (1995). Эрос и эволюция: естественная философия секса . Книги Хеликс. Ридинг, Массачусетс: Аддисон-Уэсли . ISBN 978-0-201-40754-9 . LCCN 94013158 . ОСЛК 30625193 .
- Мишод, Ричард Э. (1999). Дарвиновская динамика: эволюционные переходы в приспособленности и индивидуальности . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02699-2 . LCCN 98004166 . OCLC 38948118 .
- Миллер, Уильям Б. младший (2013). Микрокосм внутри: эволюция и вымирание в гологеноме . Бока-Ратон, Флорида: Universal-Publishers . ISBN 978-1-61233-277-2 . LCCN 2013033832 . OCLC 859168474 .
- Новак, Рональд М. (1999). Млекопитающие мира Уокера (6-е изд.). Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса . ISBN 978-0-8018-5789-8 . LCCN 98023686 . ОСЛК 39045218 .
- Саймонс, Дональд (1979). Эволюция человеческой сексуальности . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-502535-4 . LCCN 78023361 . OCLC 4494283 .
- Уильямс, Джордж К. (1974) [первоначально опубликовано в 1966 году]. Адаптация и естественный отбор: критика некоторых современных эволюционных идей . Принстонская научная библиотека. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02357-1 . LCCN 65017164 . ОСЛК 8500898 .
- Уильямс, Джордж К. (1992). Естественный отбор: области, уровни и проблемы . Оксфордская серия по экологии и эволюции. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-506933-4 . LCCN 91038938 . OCLC 228136567 .
Дальнейшее чтение [ править ]
- Гаврилец, С. (2004). Фитнес-ландшафты и происхождение видов . Издательство Принстонского университета. ISBN 978-0691119830 .
- Грант, Верн (1981). Видообразование растений (2-е изд.). Нью-Йорк: Издательство Колумбийского университета . ISBN 978-0-231-05112-5 . LCCN 81006159 . OCLC 7552165 .
- Марко, Питер Б. (2008). «Аллопатрия». В Йоргенсене, Свен Эрик ; Фат, Брайан (ред.). Энциклопедия экологии . Том. 1, AC (1-е изд.). Оксфорд, Великобритания: Elsevier . стр. 131–138. ISBN 978-0-444-52033-3 . LCCN 2008923435 . OCLC 173240026 .
- Майр, Эрнст (1963). Виды животных и эволюция . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета . ISBN 978-0-674-03750-2 . LCCN 63009552 . OCLC 899044868 .
- Шильтуизен, Менно (2001). Лягушки, мухи и одуванчики: создание видов . Оксфорд; Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-850393-4 . LCCN 2001270180 . OCLC 46729094 .
- Шапиро, Дж. Б.; Ледюк, JB.; Маллет, Дж. (2016). «Что такое видообразование?» . ПЛОС Генетика . 12 (3): e1005860. дои : 10.1371/journal.pgen.1005860 . ПМЦ 4816541 . ПМИД 27030977 .
- Уайт, Майкл Джей Ди (1978). Способы видообразования . Серия книг по биологии. Сан-Франциско, Калифорния: WH Freeman and Company . ISBN 978-0-7167-0284-9 . LCCN 77010955 . ОСЛК 3203453 .
Внешние ссылки [ править ]
- Боксхорн, Джозеф (1 сентября 1995 г.). «Наблюдаемые случаи видообразования» . Архив TalkOrigins . Хьюстон, Техас: Фонд TalkOrigins, Inc.
- Хоукс, Джон Д. (9 февраля 2005 г.). «Видообразование» . Блог Джона Хоукса .
- «Видообразование» . Калифорнийский университет, Беркли. 13 марта 2021 г.
