Аллопатрическое видообразование
| Часть серии о |
| Эволюционная биология |
|---|
 |
Аллопатрическое видообразование (от древнегреческого ἄλλος (állos) «другой» и πατρίς (patrís) «отечество») — также называемое географическим видообразованием , викариантным видообразованием или его более ранним названием « модель гантели». [1] : 86 – это способ видообразования , который происходит, когда биологические популяции становятся географически изолированными друг от друга до такой степени, что предотвращает или мешает потоку генов .
Могут возникнуть различные географические изменения, такие как движение континентов и образование гор, островов, водоемов или ледников. Человеческая деятельность, такая как сельское хозяйство или развитие, также может изменить распределение популяций видов. Эти факторы могут существенно изменить географию региона, что приведет к разделению популяции вида на изолированные субпопуляции. Затем викариантные популяции претерпевают генетические изменения, поскольку они подвергаются различному давлению отбора , испытывают генетический дрейф и накапливают различные мутации отдельных популяций в генофондах . Барьеры препятствуют обмену генетической информацией между двумя популяциями, что приводит к репродуктивной изоляции . Если две популяции вступят в контакт, они не смогут воспроизводиться, то есть фактически видообразовываться. Другие изолирующие факторы, такие как рассредоточение популяции, ведущее к эмиграции, могут вызвать видообразование (например, расселение и изоляция вида на океаническом острове) и считаются особым случаем аллопатрического видообразования, называемого перипатрическое видообразование .
Аллопатрическое видообразование обычно подразделяется на две основные модели: викариантное и перипатрическое. Эти модели отличаются друг от друга размером населения и механизмами географической изоляции. Термины аллопатрия и викариантность часто используются в биогеографии для описания взаимоотношений между организмами которых , ареалы существенно не перекрываются, но непосредственно соседствуют друг с другом — они не встречаются вместе или встречаются только в узкой зоне контакта. Исторически язык, используемый для обозначения способов видообразования, напрямую отражал биогеографическое распространение. [2] Таким образом, аллопатрия представляет собой географическое распространение, противоположное симпатрии (видообразованию в пределах одной и той же территории). Более того, термины аллопатрическое, викариантное и географическое видообразование часто используются в научной литературе как синонимы. [2] Эта статья будет посвящена аналогичной теме, за исключением особых случаев, таких как перипатрические, центробежные и другие.
Наблюдение за природой создает трудности в наблюдении за аллопатрическим видообразованием от «начала до конца», поскольку оно действует как динамический процесс. [3] Из-за этого возникает множество различных проблем, связанных с определением видов , определением изолирующих барьеров, измерением репродуктивной изоляции и т. д. Тем не менее, вербальные и математические модели, лабораторные эксперименты и эмпирические данные в подавляющем большинстве подтверждают существование аллопатрического видообразования в природе. [4] [1] : 87–105 Математическое моделирование генетической основы репродуктивной изоляции подтверждает вероятность аллопатрического видообразования; тогда как лабораторные эксперименты с дрозофилой и другими видами животных и растений подтвердили, что репродуктивная изоляция развивается как побочный продукт естественного отбора . [1] : 87
Модель викарианства [ править ]

Идея викариантной эволюции была впервые разработана Леоном Круаза в середине двадцатого века. [5] [6] Теория викарианства, которая показала согласованность с признанием тектоники плит в 1960-х годах, была разработана в начале 1950-х годов этим венесуэльским ботаником, который нашел объяснение сходству растений и животных, обитающих в Южной Америке и Африке, сделав вывод, что Первоначально они представляли собой единую популяцию, прежде чем два континента разошлись.
В настоящее время видообразование путем викариантности широко рассматривается как наиболее распространенная форма видообразования; [4] и является основной моделью аллопатрического видообразования. Викариантность — это процесс, при котором географический ареал отдельного таксона или всей биоты разделяется на прерывистые популяции ( дизъюнктивные распределения ) за счет образования внешнего барьера для обмена генами: то есть барьера, возникающего извне по отношению к разновидность. Эти внешние барьеры часто возникают в результате различных геологических изменений , топографических таких как: образование гор ( орогения ); образование рек или водоемов; оледенение ; образование или ликвидация сухопутных мостов ; движение континентов во времени ( тектоническими плитами ); или островное образование, включая небесные острова . Вивариантные барьеры могут изменить распределение популяций видов. Подходящая или неподходящая среда обитания может возникнуть, расшириться, сузиться или исчезнуть в результате глобального изменения климата или даже крупномасштабной деятельности человека (например, сельскохозяйственного, гражданского строительства и фрагментации среды обитания ). Такие факторы могут существенно изменить географию региона, что приведет к разделению популяции вида на изолированные субпопуляции. Затем викариантные популяции могут подвергнуться генотипическая или фенотипическая дивергенция, поскольку: (а) популяций возникают различные мутации в генофондах , (б) они подвергаются различному селективному давлению и/или (в) они независимо подвергаются генетическому дрейфу . Внешние барьеры препятствуют обмену генетической информацией между двумя популяциями, что потенциально может привести к дифференциации из-за экологически разных мест обитания, в которых они живут; тогда избирательное давление неизменно приводит к полной репродуктивной изоляции . [1] : 86 Более того, склонность вида оставаться в своей экологической нише (см. консерватизм филогенетической ниши ) из-за изменения условий окружающей среды также может играть роль в изоляции популяций друг от друга, стимулируя эволюцию новых линий. [7] [8]
Аллопатрическое видообразование можно представить как крайнюю точку континуума потока генов . Таким образом, уровень потока генов между популяциями в аллопатрии будет равен , где соответствует скорости генного обмена. В сочувствии (панмиксис), находясь в парапатрическом видообразовании , представляет собой весь континуум, [9] хотя некоторые учёные утверждают [2] [10] что схема классификации, основанная исключительно на географическом режиме, не обязательно отражает сложность видообразования. [11] Аллопатрия часто рассматривается как стандартная или «нулевая» модель видообразования. [2] [12] но это тоже обсуждается. [13]




изоляция Репродуктивная
Репродуктивная изоляция действует как основной механизм, способствующий генетической дивергенции в аллопатрии. [14] и может быть усилен дивергентным отбором . [15] Презиготическая и постзиготическая изоляция часто являются наиболее цитируемыми механизмами аллопатрического видообразования, и поэтому трудно определить, какая форма развилась первой в событии аллопатрического видообразования. [14] Презиготический просто подразумевает наличие барьера перед любым актом оплодотворения (например, экологический барьер, разделяющий две популяции), тогда как постзиготический подразумевает предотвращение успешного межпопуляционного скрещивания после оплодотворения (например, образование бесплодной популяции). гибрид ). Поскольку пары видов, которые разошлись в аллопатрии, часто демонстрируют механизмы пре- и постзиготической изоляции, исследование самых ранних стадий жизненного цикла вида может указать, произошла ли дивергенция из-за презиготического или постзиготического фактора. Однако установление конкретного механизма может быть неточным, поскольку пары видов со временем постоянно расходятся. Например, если растение испытывает событие дупликации хромосом , произойдет размножение, но в результате появятся стерильные гибриды, функционирующие как форма постзиготической изоляции. Впоследствии вновь образовавшаяся пара видов может столкнуться с презиготическими барьерами для размножения, поскольку отбор, действующий на каждый вид независимо, в конечном итоге приведет к генетическим изменениям, делающим гибриды невозможными. С точки зрения исследователя, нынешний изолирующий механизм может не отражать прежний изолирующий механизм. [14]
Армирование [ править ]

Подкрепление было спорным фактором в видообразовании. [16] Его чаще используют в исследованиях симпатрического видообразования , поскольку он требует потока генов между двумя популяциями. Однако подкрепление может также играть роль в аллопатрическом видообразовании, при котором репродуктивный барьер удаляется, воссоединяя две ранее изолированные популяции. При вторичном контакте особи размножаются, создавая низкоприспособленные гибриды . [17] Черты гибридов заставляют людей различать выбор партнера , в результате чего презиготическая изоляция между популяциями увеличивается. [11] Были выдвинуты некоторые аргументы, которые предполагают, что гибриды сами по себе могут стать самостоятельными видами: [18] известное как гибридное видообразование . Подкрепление может играть роль во всех географических (и других негеографических) способах видообразования, пока присутствует поток генов и могут образовываться жизнеспособные гибриды. Производство нежизнеспособных гибридов — это форма смещения репродуктивного признака , согласно которой большинство определений подразумевает завершение события видообразования. [11]
Исследования четко установили тот факт, что межвидовая дискриминация партнеров происходит в большей степени между симпатрическими популяциями, чем в чисто аллопатрических популяциях; однако были предложены другие факторы, объясняющие наблюдаемые закономерности. [19] Было показано, что подкрепление при аллопатрии происходит в природе ( доказательство видообразования путем подкрепления ), хотя и с меньшей частотой, чем классическое событие аллопатрического видообразования. [14] Основная трудность возникает при интерпретации роли подкрепления в аллопатрическом видообразовании, поскольку текущие филогенетические закономерности могут указывать на прошлый поток генов. Это маскирует возможную первоначальную дивергенцию в аллопатрии и может указывать на событие видообразования «смешанного типа», демонстрирующее как аллопатрические, так и симпатрические процессы видообразования. [13]
Математические модели [ править ]
Математические сценарии, разработанные в контексте генетической основы репродуктивной изоляции, моделируют как презиготическую , так и постзиготическую изоляцию с учетом эффектов генетического дрейфа, отбора, полового отбора или различных комбинаций этих трех факторов. Масатоши Ней и его коллеги были первыми, кто разработал модель видообразования нейтральную стохастическую только за счет генетического дрейфа. И отбор, и дрейф могут привести к постзиготической изоляции, что подтверждает тот факт, что две географически разделенные популяции могут развить репродуктивную изоляцию. [1] : 87 — иногда происходит быстро. [20] Рыболовный половой отбор также может привести к репродуктивной изоляции, если между каждой популяцией существуют незначительные различия в селективном давлении (например, риск нападения хищников или различия в среде обитания). [21] (См. раздел «Дополнительная литература» ниже). Математические модели, касающиеся репродуктивной изоляции на расстоянии, показали, что население может испытывать возрастающую репродуктивную изоляцию, которая напрямую коррелирует с физической и географической удаленностью. [22] [23] Это было проиллюстрировано на моделях кольцевых видов ; [11] однако утверждалось, что кольцевые виды представляют собой особый случай, представляющий репродуктивную изоляцию на расстоянии и вместо этого демонстрируют парапатрическое видообразование. [1] : 102 — поскольку парапатрическое видообразование представляет собой видообразование, происходящее вдоль клина .
Другие модели [ править ]
Были разработаны различные альтернативные модели аллопатрического видообразования. Особые случаи викариантного видообразования были изучены очень подробно, одним из которых является перипатрическое видообразование, при котором небольшая часть популяции вида становится географически изолированной; и центробежное видообразование, альтернативная модель перипатрического видообразования, касающаяся расширения и сокращения ареала вида. [4] Были разработаны и другие второстепенные аллопатрические модели, которые обсуждаются ниже.
Перипатрический [ править ]

Перипатрическое видообразование — это способ видообразования, при котором новый вид образуется из изолированной периферической популяции. [1] : 105 Если небольшая популяция вида становится изолированной ( например, популяция птиц на океаническом острове), отбор может действовать на популяцию независимо от родительской популяции. Учитывая как географическое разделение, так и достаточное количество времени, видообразование может стать побочным продуктом. [14] Его можно отличить от аллопатрического видообразования по трем важным характеристикам: 1) размеру изолированной популяции, 2) сильному отбору, вызванному расселением и колонизацией в новых средах, и 3) потенциальным последствиям генетического дрейфа для небольших популяций. [1] : 105 Однако исследователям часто бывает трудно определить, имело ли место перипатрическое видообразование, поскольку можно использовать викариантные объяснения из-за того, что обе модели постулируют отсутствие потока генов между популяциями. [24] Размер изолированной популяции важен, поскольку особи, колонизирующие новую среду обитания, вероятно, содержат лишь небольшой образец генетических вариаций исходной популяции. приводит к быстрой фиксации аллеля Это способствует дивергенции из-за сильного давления отбора, что в популяции потомков. Это приводит к потенциальному развитию генетической несовместимости. Эта несовместимость вызывает репродуктивную изоляцию, вызывая быстрое видообразование. [1] : 105–106 Модели перипатрии в основном подтверждаются закономерностями распределения видов в природе. Океанические острова и архипелаги представляют собой убедительные эмпирические доказательства существования перипатрического видообразования. [1] : 106–110
Центробежный [ править ]
Центробежное видообразование - это вариант, альтернативная модель перипатрического видообразования. Эта модель контрастирует с перипатрическим видообразованием в силу происхождения генетической новизны, которая приводит к репродуктивной изоляции. [25] Когда популяция вида переживает период расширения и сокращения географического ареала, она может оставить после себя небольшие, фрагментированные, изолированные на периферии популяции. Эти изолированные популяции будут содержать образцы генетических вариаций более крупной родительской популяции. Это изменение приводит к более высокой вероятности специализации экологической ниши и эволюции репродуктивной изоляции. [4] [26] Центробежное видообразование в значительной степени игнорировалось в научной литературе. [27] [25] [28] Тем не менее, исследователи представили множество доказательств в поддержку этой модели, большая часть которых до сих пор не опровергнута. [4] Одним из примеров является возможный центр происхождения в Индо-Западной части Тихого океана . [27]
Микроаллопатрический [ править ]

Микроаллопатрия относится к аллопатрическому видообразованию, происходящему в небольшом географическом масштабе. [29] Описаны примеры микроаллопатрического видообразования в природе. Рико и Тернер обнаружили внутриозёрное аллопатрическое расхождение Pseudotropeus callainos ( Maylandia callainos ) в озере Малави, разделенное всего лишь 35 метрами. [30] Гюстав Поле обнаружил доказательства того, что виды подсемейства Cryptorhynchinae микроаллопатрически видообразовались на Рапе и окружающих ее островках . [31] Симпатрически распределенная тройка видов жуков-плавунчиков ( Paroster ) , обитающих в водоносных горизонтах австралийского региона Йилгарн , вероятно, видоизменилась микроаллопатрически в пределах 3,5 км. 2 область. [32] Этот термин был первоначально предложен Хобартом М. Смитом для описания уровня географического разрешения. Симпатрическая популяция может существовать в низком разрешении, тогда как при более высоком разрешении ( т.е. в небольшом, локализованном масштабе внутри популяции) она является «микроаллопатрической». [33] Бен Фитцпатрик и его коллеги утверждают, что это первоначальное определение «вводит в заблуждение, поскольку путает географические и экологические концепции». [29]
Режимы со вторичным контактом [ править ]
Экологическое видообразование может происходить аллопатрически, симпатрически или парапатрически; единственное требование состоит в том, чтобы это происходило в результате адаптации к различным экологическим или микроэкологическим условиям. [34] Экологическая аллопатрия — это обратная форма аллопатрического видообразования в сочетании с подкреплением . [13] Во-первых, дивергентный отбор отделяет неаллопатрическую популяцию, возникающую из презиготических барьеров, из которых развиваются генетические различия из-за препятствия полному потоку генов. [35] Термины алло-парапатрический и алло-симпатрический использовались для описания сценариев видообразования, когда в аллопатрии происходит расхождение, но видообразование происходит только при вторичном контакте. [1] : 112 По сути, это модели армирования. [36] или события видообразования «смешанного режима». [13]
Наблюдательные данные [ править ]

Поскольку аллопатрическое видообразование широко признано распространенным способом видообразования, в научной литературе имеется множество исследований, подтверждающих его существование. Биолог Эрнст Майр был первым, кто обобщил современную литературу того времени в 1942 и 1963 годах. [1] : 91 Многие из приведенных им примеров остаются убедительными; однако современные исследования подтверждают географическое видообразование с помощью молекулярной филогенетики. [37] — добавление уровня надежности, недоступного ранним исследователям. [1] : 91 Самым последним тщательным исследованием аллопатрического видообразования (и исследований видообразования в целом) является публикация Джерри Койна и Х. Аллена Орра » 2004 года «Видообразование . Они перечисляют шесть основных аргументов, подтверждающих концепцию викариантного видообразования:
- Близкородственные пары видов чаще всего обитают в прилегающих друг к другу географических ареалах, разделенных географическим или климатическим барьером.
- Пары молодых видов (или сестринские виды) часто встречаются в аллопатрии даже без известного барьера.
- В тех случаях, когда несколько пар родственных видов имеют общий ареал, они распределяются граничащими друг с другом, с границами, демонстрирующими зоны гибридизации .
- В регионах, где географическая изоляция сомнительна, виды не имеют сестринских пар.
- Корреляция генетических различий между множеством отдаленно родственных видов, которые соответствуют известным текущим или историческим географическим барьерам.
- Меры репродуктивной изоляции возрастают с увеличением географического расстояния между двумя парами видов. (Это часто называют репродуктивной изоляцией на расстоянии . [11] )
Эндемизм [ править ]
Аллопатрическое видообразование привело к появлению многих закономерностей биогеографии и биоразнообразия, обнаруженных на Земле: на островах, [38] континенты, [39] и даже среди гор. [40]
Острова часто являются домом для видов -эндемиков , существующих только на острове и больше нигде в мире, причем почти все таксоны, обитающие на изолированных островах, имеют общее происхождение с видами на ближайшем континенте. [41] Не без проблем, обычно существует корреляция между островными эндемиками и разнообразием ; [42] то есть чем больше разнообразие (видовое богатство) острова, тем больше рост эндемизма. [43] Увеличение разнообразия эффективно стимулирует видообразование. [44] Кроме того, количество эндемиков на острове напрямую коррелирует с относительной изоляцией острова и его территории. [45] В некоторых случаях видообразование на островах происходило быстро. [46]
Распространение и видообразование in situ — это факторы, объясняющие происхождение организмов на Гавайях. [47] Различные географические способы видообразования были тщательно изучены в биоте Гавайев, и, в частности, покрытосеменные , по- видимому, видоизменились преимущественно аллопатрическим и парапатрическим способами. [47]
Острова — не единственные географические места, где обитают эндемичные виды. Южная Америка широко изучена, ее районы эндемизма представляют собой совокупности аллопатрически распределенных групп видов. Чарис Бабочки являются основным примером, ограниченным определенными регионами, соответствующими филогении других видов бабочек, земноводных , птиц, сумчатых , приматов , рептилий и грызунов . [48] Эта закономерность указывает на повторяющиеся события викариантного видообразования среди этих групп. [48] Считается, что реки могут играть роль географических барьеров для Чариса . [1] : 97 мало чем отличается от гипотезы речных барьеров, используемой для объяснения высокого уровня разнообразия в бассейне Амазонки , хотя эта гипотеза оспаривается. [49] [50] Аллопатрическое видообразование, опосредованное расселением, также считается важным фактором диверсификации в Неотропах . [51]

Модели роста эндемизма на возвышенностях как на островах, так и на континентах были задокументированы на глобальном уровне. [40] По мере увеличения топографической высоты виды изолируются друг от друга; [52] часто ограничены градуированными зонами. [40] Эта изоляция на «горных островах» создает барьеры для потока генов, поощряя аллопатрическое видообразование и вызывая образование эндемичных видов. [40] Горообразование ( орогения ) напрямую связано с биоразнообразием и напрямую влияет на него. [53] [54] Например, образование Гималаев и Цинхай-Тибетского нагорья способствовало видообразованию и разнообразию многочисленных растений и животных. [55] такие как Lepisorus ; папоротники [56] глиптостерноидные рыбы ( Sisoridae ); [57] и комплекс видов Rana chensinensis . [58] Аплифт также привел к викариантному видообразованию маргариток Macowania горах Южной Африки в Драконовых . [59] вместе с древолазами Dendrocincla в южноамериканских Андах . [60] Ларамидная складчатость в позднем мелу даже вызвала викариантное видообразование и распространение динозавров в Северной Америке. [61]
Адаптивная радиация , как и у галапагосских вьюрков, наблюдаемых Чарльзом Дарвином , часто является следствием быстрого аллопатрического видообразования среди популяций. Однако в случае с вьюрками Галапагосских островов, среди других островных излучений, таких как медоносы на Гавайях, представляют собой случаи ограниченного географического разделения и, вероятно, были обусловлены экологическим видообразованием .
Панамский перешеек [ править ]

Геологические данные подтверждают окончательное закрытие Панамского перешейка примерно 2,7–3,5 млн лет назад. [62] с некоторыми данными, предполагающими более ранний временный мост, существовавший между 13 и 15 млн лет назад. [63] Недавние данные все больше указывают на более древнее и сложное возникновение перешейка, при котором распространение ископаемых и существующих видов (часть американского биотического обмена ) происходит тремя основными импульсами: в Северную и Южную Америку и обратно. [64] Кроме того, изменения в наземном биотическом распространении на обоих континентах, такие как армейские муравьи Eciton, подтверждают более ранний мост или серию мостов. [65] [66] Независимо от точного времени приближения перешейка, биологи смогут изучать виды на Тихоокеанской и Карибской сторонах в рамках того, что было названо «одним из величайших естественных экспериментов в эволюции». [62] Кроме того, как и в случае с большинством геологических событий, закрытие вряд ли произошло быстро, а скорее динамично — постепенное обмеление морской воды в течение миллионов лет. [1] : 93
Исследования щелкающих креветок рода Alpheus предоставили прямые доказательства аллопатрического видообразования. [67] поскольку филогенетические реконструкции подтверждают взаимоотношения 15 пар сестринских видов Alpheus , каждая пара разделена перешейком. [62] а датировка по молекулярным часам подтверждает их разделение между 3 и 15 миллионами лет назад. [68] Недавно выделившиеся виды обитают в мелких мангровых зарослей. водах [68] в то время как более старые дивергентные виды обитают на большей глубине, что коррелирует с постепенным закрытием перешейка. [1] : 93 В пользу аллопатрической дивергенции также свидетельствуют лабораторные эксперименты над парами видов, показавшие почти полную репродуктивную изоляцию. [1] : 93
Подобные закономерности родства и распространения по Тихоокеанской и Атлантической сторонам были обнаружены у других пар видов, таких как: [69]
- Диадема Вест-Индии и диадема Мексики.
- Эхинометра лукантерская и Эхинометра ванбрунти.
- Echinometra viridis и E. vanbrunti
- Bathygobius soporator и Bathygobius ramosus
- B. soporator и Bathygobius andrei
- Excirolana braziliensis и варианты морф
Refugia[editУбежище
Ледниковые периоды сыграли важную роль в облегчении видообразования среди видов позвоночных. [70] Эта концепция рефугиумов применялась к многочисленным группам видов и их биогеографическому распространению. [1] : 97
Оледенение и последующее отступление вызвали видообразование у многих бореальных лесных птиц. [70] например, с североамериканскими сапсосками ( желтобрюхими , красношерстными и красногрудыми ); славки рода Setophaga ( S. Townsendii , S. occidentalis и S. virens ), Oreothlypis ( O. Virginiae , O. Ridgwayi и O. ruficapilla ) и Oporornis ( O. tolmiei и O. philadelphia , теперь отнесенные к род Geothlypis ); лисицы-воробьи (подвиды P. (i.) unalaschensis , P. (i.) megarhyncha и P. (i.) schistacea ); Виреон ( V. Plumbeus , V. cassinii и V. solitarius ); мухоловки-тираны ( E. occidentalis и E. difficilis ); синицы ( P. rufescens и P. hudsonicus ); и дрозды ( C. bicknelli и C. minimus ). [70]
Как особый случай аллопатрического видообразования, перипатрическое видообразование часто используется в случаях изоляции в рефугиумах оледенения, поскольку небольшие популяции становятся изолированными из-за фрагментации среды обитания, например, с североамериканской красной ( Picea Rubens ) и черной ( Picea mariana ) елью. [71] или луговые собачки Cynomys mexicanus и C. ludovicianus . [72]
Надвиды [ править ]

Многочисленные пары видов или группы видов демонстрируют смежные модели распространения, то есть проживают в географически различных регионах рядом друг с другом. Они часто имеют общие границы, многие из которых содержат гибридные зоны. Некоторые примеры смежных видов и надвидов (неформальный ранг, относящийся к комплексу близкородственных аллопатрически распространенных видов, также называемых алловидами). [73] ) включать:
- Западные и восточные луговые жаворонки в Северной Америке обитают в засушливых западных и влажных восточных географических регионах с редкими случаями гибридизации, большая часть которых приводит к бесплодию потомства. [41]
- мухоловки-монархи, эндемичные для Соломоновых островов ; комплекс нескольких видов и подвидов ( бугенвильские , белошапочные , каштановобрюхие монархи и родственные им подвиды). [41]
- Североамериканские сапсоски и представители рода Setophaga ( певчая птица-отшельник , чернозобая зеленая славка и славка Таунсенда ). [41] [70]
- Шестьдесят шесть подвидов рода Pachycephala, обитающего на Меланезийских островах. [41] [74]
- Бонобо и шимпанзе .
- Climacteris в Австралии. Птицы-ползуны [75]
- Райские птицы гор Новой Гвинеи (род Astrapia ). [75]
- красные и желтые стержни Мерцают ; черноголовые гробоклювы и розогрудые гробоклювы ; Балтиморские иволги и иволги Буллока ; и овсянки лазури и индиго . [76] Все эти пары видов соединяются в зонах гибридизации, которые соответствуют основным географическим барьерам. [1] : 97–99
- Плоские черви Dugesia в Европе, Азии и Средиземноморье. [75]
- Дихроматические туканеты рода Selenidera могут быть надвидом, возникшим по гипотезе рефугиумов в бассейне Амазонки . [77]
Что касается птиц, то в некоторых районах наблюдаются высокие темпы формирования надвидов, например, 105 надвидов в Меланезии , составляющие 66 процентов всех видов птиц в регионе. [78] Патагония является домом для 17 надвидов лесных птиц. [79] в то время как в Северной Америке насчитывается 127 надвидов как наземных, так и пресноводных птиц. [80] В Африке к югу от Сахары насчитывается 486 воробьиных птиц, сгруппированных в 169 надвидов. [81] В Австралии также есть множество надвидов птиц: 34 процента всех видов птиц сгруппированы в надвиды. [41]
Лабораторные данные [ править ]
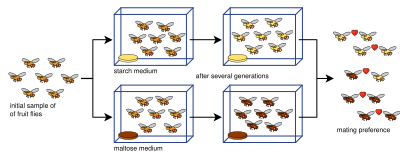
Эксперименты по аллопатрическому видообразованию часто сложны и не позволяют просто разделить популяцию вида на две части. Это связано с множеством определяющих параметров: измерение репродуктивной изоляции, размеры выборки (количество спариваний, проведенных в тестах на репродуктивную изоляцию), узкие места, продолжительность экспериментов, количество разрешенных поколений, [84] или недостаточное генетическое разнообразие. [85] Для измерения репродуктивной изоляции были разработаны различные индексы изоляции (и часто используются в лабораторных исследованиях видообразования), такие как здесь (индекс [86] и индекс [87] ):




Здесь, и представляют количество спариваний в гетерогаметности , где и представляют собой гомогаметные спаривания . и это одна популяция и и это вторая популяция. Отрицательное значение обозначает отрицательное ассортативное спаривание, положительное значение означает положительное ассортативное спаривание (т.е. выражение репродуктивной изоляции), а нулевое значение (нулевое) означает, что в популяциях происходит случайное спаривание . [84]




Экспериментальные данные твердо установили тот факт, что репродуктивная изоляция развивается как побочный продукт отбора. [15] [1] : 90 Было показано, что репродуктивная изоляция возникает в результате плейотропии ( т. е. непрямого отбора, действующего на гены, кодирующие более одного признака) – того, что называется генетическим автостопом . [15] Существуют ограничения и противоречия относительно того, могут ли лабораторные эксперименты точно отразить долгомасштабный процесс аллопатрического видообразования, происходящий в природе. Эксперименты часто не достигают 100 поколений, что намного меньше, чем ожидалось, поскольку скорость видообразования в природе намного выше. считается, что [1] : 87 Более того, показатели, конкретно касающиеся эволюции репродуктивной изоляции у дрозофилы, значительно выше, чем те, которые практикуются в лабораторных условиях. [88] Используя индекс Y, представленный ранее, обзор 25 экспериментов по аллопатрическому видообразованию (включенных в таблицу ниже) показал, что репродуктивная изоляция не была такой сильной, как обычно поддерживается, и что лабораторная среда не очень подходит для моделирования аллопатрического видообразования. [84] Тем не менее, многочисленные эксперименты показали презиготическую и постзиготическую изоляцию при викариантности, некоторые менее чем за 100 поколений. [1] : 87
Ниже представлена неполная таблица лабораторных экспериментов, проведенных по аллопатрическому видообразованию. В первом столбце указаны виды, использованные в указанном исследовании, где столбец «Признак» относится к конкретной характеристике, выбранной за или против этого вида. В столбце «Поколения» указано количество поколений в каждом проведенном эксперименте. Если было сформировано более одного эксперимента, поколения разделяются точкой с запятой или тире (задаются в виде диапазона). В некоторых исследованиях указывается продолжительность эксперимента. В столбце «Тип выборки» указывается, моделировалось ли в исследовании викариантное или перипатрическое видообразование (это может быть неявно). Прямой отбор относится к отбору, направленному на содействие репродуктивной изоляции, тогда как непрямой отбор подразумевает изоляцию, происходящую как плейотропный побочный продукт естественного отбора; тогда как дивергентный отбор подразумевает преднамеренный отбор каждой аллопатрической популяции в противоположных направлениях ( например, одна линия с большим количеством щетинок, а другая линия с меньшим количеством щетинок). В некоторых исследованиях проводились эксперименты по моделированию или контролю генетического дрейфа. Репродуктивная изоляция происходила презиготно, постзиготически и то и другое или не происходила вообще. Важно отметить, что многие из проведенных исследований включают в себя несколько экспериментов, разрешение которых не отражено в этой таблице.
| Разновидность | Черта | ~Поколения (продолжительность) | Тип выбора | Изучал дрифт | Репродуктивная изоляция | Год и ссылка |
|---|---|---|---|---|---|---|
| Дрозофила меланогастр | Ответ на побег | 18 | Косвенный; расходящийся | Да | Презиготический | 1969 [89] |
| Передвижение | 112 | Косвенный; расходящийся | Нет | Презиготический | 1974 [90] | |
| Температура, влажность | 70–130 | Косвенный; расходящийся | Да | Презиготический | 1980 [91] | |
| ДДТ Адаптация | 600 (25 лет, +15 лет) | Прямой | Нет | Презиготический | 2003 [92] | |
| 17, 9, 9, 1, 1, 7, 7, 7, 7 | Прямой, расходящийся | Презиготический | 1974 [93] | |||
| 40; 50 | Прямой; расходящийся | Презиготический | 1974 [94] | |||
| Передвижение | 45 | Прямой; расходящийся | Нет | Никто | 1979 [95] [96] | |
| Прямой; расходящийся | Презиготический | 1953 [97] | ||||
| 36; 31 | Прямой; расходящийся | Презиготический | 1956 [98] | |||
| ЭДТА Адаптация | 3 эксперимента по 25 каждый | Косвенный | Нет | Постзиготический | 1966 [99] [100] | |
| 8 экспериментов по 25 каждый | Прямой | 1997 [101] | ||||
| Брюшная чета число | 21–31 | Прямой | Да | Никто | 1958 [102] | |
| стерноплевральных Число хет | 32 | Прямой | Нет | Никто | 1969 [103] | |
| Фототакси , геотаксис | 20 | Нет | Никто | 1975 [104] 1981 [105] | ||
| Да | 1998 [106] | |||||
| Да | 1999 [107] | |||||
| Прямой; расходящийся | Презиготический | 1971 [108] 1973 [109] 1979 [110] 1983 [111] | ||||
| Д. притворяется | Щетинки щитка , скорость развития, ширина крыла; устойчивость к высыханию , плодовитость , устойчивость к этанолу ; проявление ухаживания , скорость повторного спаривания, поведение лек ; высота окукливания , комковатая яйцекладка, общая активность | 3 года | Да | Постзиготический | 1985 [112] | |
| Д. Паулисторум | 131; 131 | Прямой | Презиготический | 1976 [113] | ||
| 5 лет | 1966 [114] | |||||
| Д. Уиллистони | pH адаптация | 34–122 | Косвенный; расходящийся | Нет | Презиготический | 1980 [115] |
| Д. псевдообскура | углеводов Источник | 12 | Косвенный | Да | Презиготический | 1989 [83] |
| Температурная адаптация | 25–60 | Прямой | 1964 [116] 1969 [117] | |||
| Фототакси, геотаксис | 5–11 | Косвенный | Нет | Презиготический | 1966 [118] | |
| Презиготический | 1978 [119] 1985 [120] | |||||
| Да | 1993 [121] | |||||
| Температурный фотопериод ; еда | 37 | Дивергент | Да | Никто | 2003 [122] | |
| Д.псевдообскура и | 22; 16; 9 | Прямой; расходящийся | Презиготический | 1950 [123] | ||
| 4 эксперимента по 18 каждый | Прямой | Презиготический | 1966 [124] | |||
| Д. мохавенсис | 12 | Прямой | Презиготический | 1987 [125] | ||
| Время разработки | 13 | Дивергент | Да | Никто | 1998 [126] | |
| Д. адиастола | Да | Презиготический | 1974 [127] | |||
| Д. сильвестрис | Да | 1980 [128] | ||||
| Дом летать | Геотаксис | 38 | Косвенный | Нет | Презиготический | 1974 [129] |
| Геотаксис | 16 | Прямой; расходящийся | Нет | Презиготический | 1975 [130] | |
| Да | 1991 [131] | |||||
| Бактроцера тыквенная | Время разработки | 40–51 | Дивергент | Да | Презиготический | 1999 [132] |
| Зеа Мэйс | 6; 6 | Прямой; расходящийся | Презиготический | 1969 [133] | ||
| Д. Гримшави | [134] |
и методы исследования История
Ранние исследования видообразования обычно отражали географическое распространение и поэтому назывались географическими, полугеографическими и негеографическими. [2] Географическое видообразование соответствует сегодняшнему использованию термина аллопатрическое видообразование, и в 1868 году Мориц Вагнер был первым, кто предложил эту концепцию. [135] из которых он использовал термин Separationstheorie . [136] Его идея была позже интерпретирована Эрнстом Майром как форма видообразования по эффекту основателя , поскольку она была сосредоточена в первую очередь на небольших географически изолированных популяциях. [136]
Эдвард Бэгналл Поултон , биолог-эволюционист и ярый сторонник важности естественного отбора, подчеркнул роль географической изоляции в содействии видообразованию: [11] в процессе создания термина «симпатрическое видообразование» в 1903 году. [137]
Существуют разногласия относительно того, признал ли Чарльз Дарвин истинную географическую модель видообразования в своей публикации « Происхождение видов» . [136] В главе 11 «Географическое распределение» Дарвин обсуждает географические барьеры на пути миграции, заявляя, например, что «барьеры любого рода или препятствия на пути свободной миграции тесно и важно связаны с различиями между производством в различных регионах [ мира]». [138] Ф. Дж. Саллоуэй утверждает, что позиция Дарвина по вопросу видообразования была, по меньшей мере, «вводящей в заблуждение». [139] и, возможно, позже дезинформировал Вагнера и Дэвида Старра Джордана, заставив их поверить в то, что Дарвин считал симпатрическое видообразование наиболее важным способом видообразования. [1] : 83 Тем не менее, Дарвин так и не принял полностью концепцию географического видообразования Вагнера. [136]

Дэвид Старр Джордан сыграл значительную роль в продвижении аллопатрического видообразования в начале 20 века, предоставив множество природных доказательств в поддержку этой теории. [1] : 86 [135] [140] Намного позже биолог Эрнст Майр был первым, кто изложил современную тогда литературу в своей публикации 1942 года «Систематика и происхождение видов с точки зрения зоолога» и в своей последующей публикации 1963 года «Виды животных и эволюция» . Как и работы Джордана, они основывались на прямых наблюдениях за природой, документируя возникновение аллопатрического видообразования, которое сегодня широко признано. [1] : 83–84 До этого исследования Феодосий Добжанский в 1937 году опубликовал книгу «Генетика и происхождение видов» , в которой сформулировал генетическую основу того, как может происходить видообразование. [1] : 2
Другие ученые отметили существование в природе аллопатрически распределенных пар видов, например, Джоэл Асаф Аллен (который придумал термин «Закон Джордана», согласно которому близкородственные, географически изолированные виды часто оказываются разделенными физическим барьером). [1] : 91 ) и Роберт Гринлиф Ливитт ; [141] однако считается, что Вагнер, Карл Джордан и Дэвид Старр Джордан сыграли большую роль в формировании аллопатрического видообразования как эволюционной концепции; [142] где Майр и Добжанский внесли свой вклад в формирование современного эволюционного синтеза .
В конце 20-го века были разработаны математические модели аллопатрического видообразования, что привело к явной теоретической правдоподобности того, что географическая изоляция может привести к репродуктивной изоляции двух популяций. [1] : 87
С 1940-х годов принято аллопатрическое видообразование. [143] Сегодня это широко считается наиболее распространенной формой видообразования, имеющей место в природе. [1] : 84 Однако это не лишено разногласий, поскольку как парапатрическое, так и симпатрическое видообразование считаются приемлемыми способами видообразования, встречающимися в природе. [143] Некоторые исследователи даже считают, что существует предвзятость в сообщениях о положительных событиях аллопатрического видообразования, и в одном исследовании, в котором рассматривались 73 статьи о видообразовании, опубликованные в 2009 году, только 30 процентов, которые предлагали аллопатрическое видообразование в качестве основного объяснения наблюдаемых закономерностей, считали другие способы видообразования возможный. [13]
Современные исследования во многом опираются на множество доказательств, чтобы определить способ видообразования; то есть определение закономерностей географического распределения в сочетании с филогенетическим родством на основе молекулярных методов. [1] : 123–124 Этот метод был эффективно представлен Джоном Д. Линчем в 1986 году, и многие исследователи использовали его и подобные методы, получив поучительные результаты. [144] Корреляция географического распределения с филогенетическими данными также породила подраздел биогеографии, называемый викариантной биогеографией. [1] : 92 разработан Джоэлом Кракрафтом , Джеймсом Брауном , Марком В. Ломолино и другими биологами, специализирующимися в области экологии и биогеографии. Аналогичным образом были предложены и применены полные аналитические подходы для определения того, какой способ видообразования прошел вид в прошлом, с использованием различных подходов или их комбинаций: филогения на уровне вида, перекрытие ареалов, симметрия размеров ареалов между парами сестринских видов и перемещение видов в пределах географических территорий. диапазоны. [37] Методы датирования по молекулярным часам также часто используются для точного измерения времени расхождения, которое отражает ископаемые или геологические летописи. [1] : 93 (например, с щелкающей креветкой, разделенной закрытием Панамского перешейка [68] или события видообразования внутри рода Cyclamen [145] ). Другие методы, используемые сегодня, используют измерение потока генов между популяциями. [13] моделирование экологической ниши (например, в случае славок Мирта и Одюбона) [146] или видообразование, опосредованное окружающей средой, происходящее среди дендробатидных лягушек в Эквадоре [144] ) и статистическое тестирование монофилетических групп. [147] Биотехнологические достижения позволили провести крупномасштабные многолокусные сравнения геномов (например, возможное событие аллопатрического видообразования, которое произошло между предками человека и шимпанзе). [148] ), связывая эволюционную историю видов с экологией и уточняя филогенетические закономерности. [149]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в Койн, Джерри А .; Орр, Х. Аллен (2004). Видообразование . Синауэр Ассошиэйтс. стр. 1–545. ISBN 978-0-87893-091-3 .
- ^ Jump up to: Перейти обратно: а б с д и Ричард Г. Харрисон (2012), «Язык видообразования», Evolution , 66 (12): 3643–3657, doi : 10.1111/j.1558-5646.2012.01785.x , PMID 23206125 , S2CID 31893065
- ^ Эрнст Майр (1970), Популяции, виды и эволюция: сокращение видов животных и эволюции , издательство Гарвардского университета, стр. 279, ISBN 978-0674690134
- ^ Jump up to: Перейти обратно: а б с д и Ховард, Дэниел Дж. (2003). «Видение: Аллопатрическое». Энциклопедия наук о жизни . дои : 10.1038/npg.els.0001748 . ISBN 978-0470016176 .
- ^ Круаза Л (1958). Панбиогеография или вводный синтез зоогеографии, фитогеографии, геологии; с заметками по эволюции, систематике, экологии, антропологии и т. д . . Каракас: Издано автором, 2755 стр.
- ^ Круаза Л (1964). Пространство, время, форма: биологический синтез . Каракас: Опубликовано автором. п. 676.
- ^ Джон Дж. Винс (2004), «Возвращение к видообразованию и экологии: консерватизм филогенетической ниши и происхождение видов», Evolution , 58 (1): 193–197, doi : 10.1554/03-447 , PMID 15058732 , S2CID 198159058
- ^ Джон Дж. Винс; Кэтрин Х. Грэм (2005), «Нишевый консерватизм: интеграция эволюции, экологии и биологии сохранения», Annual Review of Ecology, Evolution and Systematics , 36 : 519–539, doi : 10.1146/annurev.ecolsys.36.102803.095431 , S2CID 3895737
- ^ Сергей Гаврилец (2004), Фитнес-ландшафты и происхождение видов , Princeton University Press, с. 13
- ^ Сара Виа (2001), «Симпатрическое видообразование у животных: гадкий утенок растет», Trends in Ecology & Evolution , 16 (1): 381–390, doi : 10.1016/S0169-5347(01)02188-7 , PMID 11403871
- ^ Jump up to: Перейти обратно: а б с д и ж Ханнес Шулер; Глен Р. Худ; Скотт П. Иган; Джеффри Л. Федер (2016), «Способы и механизмы видообразования», Обзоры по клеточной биологии и молекулярной медицине , 2 (3): 60–93, doi : 10.1002/3527600906
- ^ Керстин Йоханнессон (2009), «Инвертирование нулевой гипотезы видообразования: взгляд на морскую улитку», Evolutionary Ecology , 23 (1): 5–16, Бибкод : 2009EvEco..23....5J , doi : 10.1007/s10682 -007-9225-1 , S2CID 23644576
- ^ Jump up to: Перейти обратно: а б с д и ж Керстин Йоханнессон (2010), «Анализируем ли мы видообразование без предубеждений?», Annals of the New York Academy of Sciences , 1206 (1): 143–149, Бибкод : 2010NYASA1206..143J , doi : 10.1111/j.1749-6632.2010 .05701.x , PMID 20860687 , S2CID 41791817
- ^ Jump up to: Перейти обратно: а б с д и Майкл Турелли; Николас Х. Бартон; Джерри А. Койн (2001), «Теория и видообразование», Тенденции в экологии и эволюции , 16 (7): 330–343, doi : 10.1016/s0169-5347(01)02177-2 , PMID 11403865
- ^ Jump up to: Перейти обратно: а б с д Уильям Р. Райс; Эллен Э. Хостерт (1993), «Лабораторные эксперименты по видообразованию: чему мы научились за 40 лет?», Evolution , 47 (6): 1637–1653, doi : 10.1111/j.1558-5646.1993.tb01257.x , JSTOR 2410209 , ПМИД 28568007 , С2КИД 42100751
- ^ Хвала, Джон А.; Вуд, Трой Э. (2012). Специализация: Введение . дои : 10.1002/9780470015902.a0001709.pub3 . ISBN 978-0470016176 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Конрад Дж. Хоскин; Меган Хигги; Кейт Р. Макдональд; Крейг Мориц (2005), «Подкрепление способствует быстрому аллопатрическому видообразованию», Nature , 437 (7063): 1353–1356, Бибкод : 2005Natur.437.1353H , doi : 10.1038/nature04004 , PMID 16251964 , S2CID 4417281
- ^ Арнольд, МЛ (1996). Естественная гибридизация и эволюция . Нью-Йорк: Издательство Оксфордского университета. п. 232. ИСБН 978-0-19-509975-1 .
- ^ Мохамед А. Ф. Нур (1999), «Подкрепление и другие последствия симпатии», Наследственность , 83 (5): 503–508, doi : 10.1038/sj.hdy.6886320 , PMID 10620021
- ^ Кристофер Дж. Уиллс (1977), «Механизм быстрого аллопатрического видообразования», The American Naturalist , 111 (979): 603–605, doi : 10.1086/283191 , S2CID 84293637
- ^ Эндрю Помианковски и Йо Иваса (1998), «Безудержное разнообразие орнаментов, вызванное фишеровским половым отбором», PNAS , 95 (9): 5106–5111, Бибкод : 1998PNAS...95.5106P , doi : 10.1073/pnas.95.9.5106 , ПМК 20221 , ПМИД 9560236
- ^ Сьюэлл Райт (1943), «Изоляция на расстоянии», Genetics , 28 (2): 114–138, doi : 10.1093/genetics/28.2.114 , PMC 1209196 , PMID 17247074
- ^ Монтгомери Слаткин (1993), «Изоляция расстоянием в равновесных и неравновесных популяциях», Evolution , 47 (1): 264–279, doi : 10.2307/2410134 , JSTOR 2410134 , PMID 28568097
- ^ Люсинда П. Лоусон; и др. (2015), «Дивергенция по краям: перипатрическая изоляция в комплексе горной колючей тростниковой лягушки», BMC Evolutionary Biology , 15 (128): 128, Bibcode : 2015BMCEE..15..128L , doi : 10.1186/s12862-015 -0384-3 , ПМК 4487588 , ПМИД 26126573
- ^ Jump up to: Перейти обратно: а б Сергей Гаврилец; и др. (2000), «Паттерны парапатрического видообразования», Evolution , 54 (4): 1126–1134, CiteSeerX 10.1.1.42.6514 , doi : 10.1554/0014-3820(2000)054[1126:pops]2.0.co;2 , PMID 11005282 , S2CID 198153997
- ^ У.Л. Браун младший (1957), «Центробежное видообразование», Ежеквартальный обзор биологии , 32 (3): 247–277, doi : 10.1086/401875 , S2CID 225071133
- ^ Jump up to: Перейти обратно: а б Джон К. Бриггс (2000), «Центробежное видообразование и центры происхождения», Journal of Biogeography , 27 (5): 1183–1188, Бибкод : 2000JBiog..27.1183B , doi : 10.1046/j.1365-2699.2000.00459. х , S2CID 86734208
- ^ Дженнифер К. Фрей (1993), «Способы формирования и видообразования периферических изолятов», Systematic Biology , 42 (3): 373–381, doi : 10.1093/sysbio/42.3.373 , S2CID 32546573
- ^ Jump up to: Перейти обратно: а б Б.М. Фицпатрик; А.А. Фордайс; С. Гаврилец (2008), «Что такое симпатрическое видообразование?», Journal of Evolutionary Biology , 21 (6): 1452–1459, doi : 10.1111/j.1420-9101.2008.01611.x , PMID 18823452 , S2CID 8721116
- ^ К. Рико; Г. Ф. Тернер (2002), «Чрезвычайное микроаллопатрическое расхождение у видов цихлид из озера Малави», Molecular Ecology , 11 (8): 1585–1590, Бибкод : 2002MolEc..11.1585R , doi : 10.1046/j.1365-294X.2002.01537 .x , hdl : 10261/59425 , PMID 12144678 , S2CID 16543963
- ^ Густав Паулай (1985), «Адаптивная радиация на изолированном океаническом острове: новый взгляд на Cryptorhynchinae (Curculionidae) Рапы», Биологический журнал Линнеевского общества , 26 (2): 95–187, doi : 10.1111/j.1095-8312.1985 .tb01554.x
- ^ М.Т. Гузик; С.Дж.Б. Купер; У. Ф. Хамфрис; А. Д. Остин (2009), «Мелкомасштабная сравнительная филогеография тройки симпатрических сестринских видов подземных жуков-плавунчиков из одного калькретового водоносного горизонта в Западной Австралии», Molecular Ecology , 18 (17): 3683–3698, Bibcode : 2009MolEc..18.3683 G , doi : 10.1111/j.1365-294X.2009.04296.x , PMID 19674311 , S2CID 25821896
- ^ Хобарт М. Смит (1965), «Больше эволюционных терминов», Systematic Biology , 14 (1): 57–58, doi : 10.2307/2411904 , JSTOR 2411904.
- ^ Носил, П. (2012). Экологический вид . Оксфорд: Издательство Оксфордского университета. п. 280. ИСБН 978-0199587117 .
- ^ Сара Виа (2009), «Естественный отбор в действии во время видообразования», PNAS , 106 (Приложение 1): 9939–9946, Bibcode : 2009PNAS..106.9939V , doi : 10.1073/pnas.0901397106 , PMC 2702801 , PMID 1952 8641
- ^ Гай Л. Буш (1994), «Симпатрическое видообразование у животных: новое вино в старых бутылках», Trends in Ecology & Evolution , 9 (8): 285–288, doi : 10.1016/0169-5347(94)90031-0 , ПМИД 21236856
- ^ Jump up to: Перейти обратно: а б Тимоти Дж. Барраклаф; Альфред П. Фоглер (2000), «Обнаружение географической структуры видообразования на основе филогении на уровне вида», American Naturalist , 155 (4): 419–434, doi : 10.2307/3078926 , JSTOR 3078926 , PMID 10753072
- ^ Роберт Дж. Уиттакер; Хосе Мария Фернандес-Паласиос (2007), Биогеография острова: экология, эволюция и сохранение (2-е изд.), Oxford University Press
- ^ Хун Цянь; Роберт Э. Риклефс (2000), «Крупномасштабные процессы и азиатский уклон в видовом разнообразии растений умеренного пояса», Nature , 407 (6801): 180–182, Бибкод : 2000Natur.407..180Q , doi : 10.1038/35025052 , PMID 11001054 , S2CID 4416820
- ^ Jump up to: Перейти обратно: а б с д Мануэль Дж. Штайнбауэр; Ричард Филд; Джон-Арвид Гритнес; Панайотис Тригас; Клодин А-Пэн; Фабио Атторре; Х. Джон Б. Биркс; Пауло А.В. Борхес; Педро Кардозо; Чанг-Хун Чжоу; Мишель Де Санктис; Мигель М. де Секейра; Мария К. Дуарте; Руи Б. Элиас; Хосе Мария Фернандес-Паласиос; Розалина Габриэль; Рой Э. Джеро; Розмари Дж. Гиллеспи; Йозеф Греймлер; Дэвид Э.В. Хартер; Цурнг-Джун Хуанг; Северин Д.Х. Ирландия; Даниэль Жанмоно; Анке Йентш; Алистер С. Джамп; Кристоф Кюффер; Сандра Ноге; Рюдигер Отто; Джонатан Прайс; Мария М. Ромейрас; Доминик Страсберг; Тод Стюсси; Йенс-Кристиан Свеннинг; Оле Р. Ветаас; Карл Байеркунляйн (2016), «Топографическая изоляция, видообразование и глобальное увеличение эндемизма с высотой» (PDF) , Глобальная экология и биогеография , 25 (9): 1097–1107, Бибкод : 2016GloEB..25.1097S , doi : 10.1111/geb.12469 , hdl : 1893/23221
- ^ Jump up to: Перейти обратно: а б с д и ж Тревор Прайс (2008), Видообразование у птиц , издательство Roberts and Company, стр. 1–64, ISBN 978-0-9747077-8-5
- ^ Сяо-Юн Чен; Фанлян Хэ (2009), «Видообразование и эндемизм в рамках модели островной биогеографии», Ecology , 90 (1): 39–45, Bibcode : 2009Ecol...90...39C , doi : 10.1890/08-1520.1 , PMID 19294911 , S2CID 24127933
- ^ Карлос Даниэль Кадена; Роберт Э. Риклефс; Иван Хименес; Элдридж Бермингем (2005), «Экология: видообразование обусловлено видовым разнообразием?», Nature , 438 (7064): E1–E2, Bibcode : 2005Natur.438E...1C , doi : 10.1038/nature04308 , PMID 16267504 , S2CID 4418564
- ^ Брент С. Эмерсон; Никлас Колм (2005), «Видовое разнообразие может стимулировать видообразование», Nature , 434 (7036): 1015–1017, Bibcode : 2005Natur.434.1015E , doi : 10.1038/nature03450 , PMID 15846345 , S2CID 3195603
- ^ Тревор Прайс (2008), Видообразование у птиц , издательство Roberts and Company, стр. 141–155, ISBN 978-0-9747077-8-5
- ^ Джонатан Б. Лосос; Дольф Шлютер (2000), «Анализ эволюционных взаимоотношений вида и территории», Nature , 408 (6814): 847–850, Бибкод : 2000Natur.408..847L , doi : 10.1038/35048558 , PMID 11130721 , S2CID 4400514
- ^ Jump up to: Перейти обратно: а б Джонатан П. Прайс; Уоррен Л. Вагнер (2004), «Видообразование в гавайских линиях покрытосеменных: причина, следствие и режим», Evolution , 58 (10): 2185–2200, doi : 10.1554/03-498 , PMID 15562684 , S2CID 198157925
- ^ Jump up to: Перейти обратно: а б Джейсон П.В. Холл; Дональд Дж. Харви (2002), «Возвращение к филогеографии Амазонии: новые данные по бабочкам-риодинидам», Evolution , 56 (7): 1489–1497, doi : 10.1554/0014-3820(2002)056[1489:tpoarn]2.0 .co;2 , PMID 12206248 , S2CID 198157398
- ^ Серджио Санторелли-младший, Уильям Э. Магнуссон и Клаудия П. Деус (2018), «Большинство видов не ограничиваются рекой Амазонки, которая считается границей между эндемическими областями», Scientific Reports , 8 (2294): 2294, Bibcode : 2018NatSR...8.2294S , doi : 10.1038/s41598-018-20596-7 , PMC 5797105 , PMID 29396491
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Лучано Н. Нака и Мария В. Пил (2020), «Выход за пределы викариантной парадигмы речного барьера», Molecular Ecology , 29 (12): 2129–2132, Бибкод : 2020MolEc..29.2129N , doi : 10.1111/mec.15465 , PMID 32392379
- ^ Брайан Тилстон Смит; Джон Э. МакКормак; Андрес М. Куэрво; Майкл. Дж. Хикерсон; Александр Алейшо; Карлос Даниэль Кадена; Хорхе Перес-Эман; Кертис В. Берни; Сяоу Се; Майкл Дж. Харви; Брант К. Фэрклот; Трэвис С. Гленн; Элизабет П. Дерриберри; Джесси Преджан; Саманта Филдс; Робб Т. Брумфилд (2014), «Движущие силы тропического видообразования» , Nature , 515 (7527): 406–409, Bibcode : 2014Natur.515..406S , doi : 10.1038/nature13687 , PMID 25209666 , S2CID 1415798
- ^ С. К. Галамбор; РБ Хьюи; PR Мартин; Дж. Т. Тьюксбери; Г. Ван (2014), «Горные перевалы в тропиках выше? Новый взгляд на гипотезу Янзена», Integrative and Comparative Biology , 46 (1): 5–7, doi : 10.1093/icb/icj003 , PMID 21672718
- ^ Карина Хорн; Фолькер Мосбругер; Андреас Мульч; Александр Антонелли (2013), «Биоразнообразие в результате горообразования» (PDF) , Nature Geoscience , 6 (3): 154, Бибкод : 2013NatGe...6..154H , doi : 10.1038/ngeo1742
- ^ Джон Фьелдсо; Раури С. К. Боуи; Карстен Рахбек (2012), «Роль горных хребтов в диверсификации птиц», Ежегодный обзор экологии, эволюции и систематики , 43 : 249–265, doi : 10.1146/annurev-ecolsys-102710-145113 , S2CID 85868089
- ^ Яоу Син; Ричард Х. Ри (2017), «Диверсификация в горах Хэндуань, горячая точка умеренного биоразнообразия, обусловленная подъемом», PNAS , 114 (17): 3444–3451, Бибкод : 2017PNAS..114E3444X , doi : 10.1073/pnas.1616063114 , ПМК 5410793 , ПМИД 28373546
- ^ Ли Ван; Харальд Шнайдер; Сянь-Чунь Чжан; Цяо-Пин Сян (2012), «Подъем Гималаев способствовал диверсификации папоротников Юго-Восточной Азии за счет изменения режима муссонов», BMC Plant Biology , 12 (210): 1–9, doi : 10.1186/1471-2229-12. -210 , ПМК 3508991 , ПМИД 23140168
- ^ Шуньпинг Хэ; Вэньсюань Цао; Юй Чен (2001), «Поднятие плато Цинхай-Сизан (Тибет) и викариантное видообразование глиптостерноидных рыб (Siluriformes: Sisoridae)» , Наука в Китае, серия C: Науки о жизни , 44 (6): 644–651, doi : 10.1007/bf02879359 , PMID 18763106 , S2CID 22432209
- ^ Вэй-Вэй Чжоу; Ян Вэнь; Цзиньчжун Фу; Юн-Бяо Сюй; Цзе-Цюн Цзинь; Ли Дин; Ми-Сук Мин; Цзин Че; Я-Пин Чжан (2012), «Видообразование в комплексе видов Rana chensinensis и его связь с поднятием Цинхай-Тибетского нагорья», Molecular Ecology , 21 (4): 960–973, Bibcode : 2012MolEc..21.. 960Z , doi : 10.1111/j.1365-294X.2011.05411.x , PMID 22221323 , S2CID 37992915
- ^ Джоан Бентли; Дж. Энтони Вербум; Никола Дж. Берг (2014), «Эрозивные процессы после тектонического поднятия стимулируют викариантное и адаптивное видообразование: эволюция эндемичного для Африки умеренного пояса рода бумажных маргариток», BMC Evolutionary Biology , 14 (27): 1–16, Bibcode : 2014BMCEE..14. ..27B , doi : 10.1186/1471-2148-14-27 , PMC 3927823 , PMID 24524661
- ^ Джейсон Т. Вейр; Момоко Прайс (2011), «Андское поднятие способствует видообразованию в низинах посредством викариантности и расселения древолазов Dendrocincla», Molecular Ecology , 20 (21): 4550–4563, Bibcode : 2011MolEc..20.4550W , doi : 10.1111/j.1365-294X .2011.05294.x , PMID 21981112 , S2CID 33626056
- ^ Терри А. Гейтс; Альберт Прието-Маркес; Линдси Э. Занно (2012), «Горостроение вызвало радиацию позднемеловых североамериканских мегатравоядных динозавров», PLOS ONE , 7 (8): e42135, Bibcode : 2012PLoSO...742135G , doi : 10.1371/journal.pone.0042135 , PMC 3410882 , ПМИД 22876302
- ^ Jump up to: Перейти обратно: а б с Карла Хёрт; Артур Анкер; Нэнси Ноултон (2008), «Многолокусный тест одновременной дивергенции на Панамском перешейке с использованием щелкающих креветок рода Alpheus», Evolution , 63 (2): 514–530, doi : 10.1111/j.1558-5646.2008.00566. х , PMID 19154357 , S2CID 11820649
- ^ К. Монтес; А. Кардона; К. Харамильо; А. Пардо; Дж. К. Сильва; В. Валенсия; К. Айала; Л.С. Перес-Анхель; Л.А. Родригес-Парра; В. Рамирес; Х. Ниньо; и др. (2015), «Закрытие Центральноамериканского морского пути в среднем миоцене», Science , 348 (6231): 226–229, Бибкод : 2015Sci...348..226M , doi : 10.1126/science.aaa2815 , PMID 25859042
- ^ Кристин Д. Бэкон; Даниэле Сильвестро; Карлос Харамильо; Брайан Тилстон Смит; Просанта Чакрабарти; Александр Антонелли (2015), «Биологические данные подтверждают раннее и сложное появление Панамского перешейка», PNAS , 112 (9): 6110–6115, Бибкод : 2015PNAS..112.6110B , doi : 10.1073/pnas.1423853112 , PMC 4434730 , ПМИД 25918375
- ^ Шон Брэди (2017), «Нашествие армейских муравьев раскрывает филогеографические процессы на Панамском перешейке», Molecular Ecology , 26 (3): 703–705, Бибкод : 2017MolEc..26..703B , doi : 10.1111/mec.13981 , ПМИД 28177197
- ^ Макс Э. Уинстон; Дэниел Дж. Кронауэр; Корри С. Моро (2017), «Ранняя и динамичная колонизация Центральной Америки стимулирует видообразование неотропических армейских муравьев», Molecular Ecology , 26 (3): 859–870, Bibcode : 2017MolEc..26..859W , doi : 10.1111/ mec.13846 , PMID 27778409
- ^ Нэнси Ноултон (1993), «Дивергенция в белках, митохондриальной ДНК и репродуктивной совместимости на Панамском перешейке», Science , 260 (5114): 1629–1632, Bibcode : 1993Sci...260.1629K , doi : 10.1126/science. 8503007 , ПМИД 8503007 , С2КИД 31875676
- ^ Jump up to: Перейти обратно: а б с Нэнси Ноултон; Ли А. Вейгт (1998), «Новые даты и новые темпы расхождения через Панамский перешеек», Proc. Р. Сок. Лонд. Б , 265 (1412): 2257–2263, doi : 10.1098/рспб.1998.0568 , PMC 1689526
- ^ ХА Лессиос. (1998). Первая стадия видообразования у организмов, разделенных Панамским перешейком. В «Бесконечных формах: виды и видообразование» (под ред. Д. Ховарда и С. Берлохера). Издательство Оксфордского университета
- ^ Jump up to: Перейти обратно: а б с д Джейсон Т. Вейр; Дольф Шлютер (2004), «Ледяные щиты способствуют видообразованию у бореальных птиц», Proceedings: Biological Sciences , 271 (1551): 1881–1887, doi : 10.1098/rspb.2004.2803 , PMC 1691815 , PMID 15347509
- ^ Хуан П. Харамильо-Корреа; Жан Буске (2003), «Новые данные на основе митохондриальной ДНК о видовых отношениях предшественника и производного между черной и красной елью (Pinaceae)», American Journal of Botany , 90 (12): 1801–1806, doi : 10.3732/ajb.90.12 .1801 , PMID 21653356
- ^ Габриэла Кастельянос-Моралес; Низа Гамес; Рейна А. Кастильо-Гамес; Луис Э. Эгиарте (2016), «Перипатрическое видообразование эндемичного вида, вызванное изменением климата плейстоцена: случай мексиканской луговой собачки ( Cynomys mexicanus )», Molecular Phylogenetics and Evolution , 94 (Pt A): 171–181, doi : 10.1016/j.ympev.2015.08.027 , PMID 26343460
- ^ Амадон Д. (1966). «Концепция надвида». Систематическая биология . 15 (3): 245–249. дои : 10.2307/sysbio/15.3.245 .
- ^ Эрнст Майр; Джаред Даймонд (2001), Птицы Северной Меланезии , Oxford University Press, стр. 143, ISBN 978-0-19-514170-2
- ^ Jump up to: Перейти обратно: а б с Эрнст Майр (1963), Виды животных и эволюция , издательство Гарвардского университета, стр. 488–515, ISBN 978-0674037502
- ^ Remington CL (1968) Шовные зоны гибридного взаимодействия между недавно присоединившимися биотами. В: Добжанский Т., Хехт М.К., Стир В.К. (ред.) Эволюционная биология. Спрингер, Бостон, Массачусетс
- ^ Юрген Хаффер (1969), «Видообразование лесных птиц Амазонки», Science , 165 (3889): 131–137, Бибкод : 1969Sci...165..131H , doi : 10.1126/science.165.3889.131 , PMID 17834730
- ^ Эрнст Майр; Джаред Даймонд (2001), Птицы Северной Меланезии , Oxford University Press, стр. 127, ISBN 978-0-19-514170-2
- ^ Франсуа Вийемье (1985), «Лесные птицы Патагонии: экологическая география, видообразование, эндемизм и история фауны», Орнитологические монографии (36): 255–304, doi : 10.2307/40168287 , JSTOR 40168287
- ^ Майр, Э., и Шорт, LL (1970). Видовые таксоны птиц Северной Америки: вклад в сравнительную систематику .
- ^ Холл, BP, и Моро, RE (1970). Атлас видообразования африканских воробьиных птиц . Попечители Британского музея (Естественная история).
- ^ Дж. Р. Пауэлл; М. Анджелкович (1983), «Популяционная генетика амилазы дрозофилы . IV. Селекция в лабораторных популяциях, поддерживаемых на различных углеводах», Genetics , 103 (4): 675–689, doi : 10.1093/genetics/103.4.675 , PMC 1202048 , ПМИД 6189764
- ^ Jump up to: Перейти обратно: а б Дайан М.Б. Додд (1989), «Репродуктивная изоляция как следствие адаптивной дивергенции у Drosophila pseudoobscura », Evolution , 43 (6): 1308–1311, doi : 10.2307/2409365 , JSTOR 2409365 , PMID 28564510
- ^ Jump up to: Перейти обратно: а б с д Анн-Бритт Флорин; Андерс Один (2002), «Лабораторная среда не способствует аллопатрическому видообразованию», Журнал эволюционной биологии , 15 : 10–19, doi : 10.1046/j.1420-9101.2002.00356.x , S2CID 85410953
- ^ Jump up to: Перейти обратно: а б Марк Киркпатрик; Виржини Равинье (2002), «Видение посредством естественного и полового отбора: модели и эксперименты», The American Naturalist , 159 (3): S22, doi : 10.2307/3078919 , JSTOR 3078919
- ^ Бишоп, Ю.М .; Финберг, SE; Холланд, PW (1975), Дискретный многомерный анализ: теория и практика , MIT Press: Кембридж, Массачусетс.
- ^ HD Stalker (1942), «Исследования половой изоляции в видовом комплексе Drosophila virilis », Genetics , 27 (2): 238–257, doi : 10.1093/genetics/27.2.238 , PMC 1209156 , PMID 17247038
- ^ Джерри А. Койн ; Х. Аллен Орр (1997), « Возвращение к закономерностям видообразования у дрозофилы», Evolution , 51 (1): 295–303, doi : 10.1111/j.1558-5646.1997.tb02412.x , PMID 28568795 , S2CID 40390753
- ^ Грант бакалавра наук; LE Mettler (1969), «Разрушительный и стабилизирующий отбор на «побеговое» поведение Drosophila melanogaster », Genetics , 62 (3): 625–637, doi : 10.1093/genetics/62.3.625 , PMC 1212303 , PMID 17248452
- ^ Б. Бернет; К. Коннолли (1974), «Активность и сексуальное поведение Drosophila melanogaster », «Генетика поведения» : 201–258.
- ^ Г. Килиас; С.Н. Алахиотис; М. Пелеканос (1980), «Многофакторное генетическое исследование теории видообразования с использованием Drosophila melanogaster », Evolution , 34 (4): 730–737, doi : 10.2307/2408027 , JSTOR 2408027 , PMID 28563991
- ^ КРБ Боаке; К. Макдональд; С. Майтра; Р. Гангули (2003), «Сорок лет одиночества: расхождение в истории жизни и поведенческая изоляция между лабораторными линиями Drosophila melanogaster », Journal of Evolutionary Biology , 16 (1): 83–90, doi : 10.1046/j.1420- 9101.2003.00505.x , PMID 14635883 , S2CID 24040182
- ^ Дж. С. Ф. Баркер; LJE Karlsson (1974), «Влияние размера популяции и интенсивности отбора на реакцию на разрушительный отбор у Drosophila melanogaster », Genetics , 78 (2): 715–735, doi : 10.2307/2407287 , JSTOR 2407287 , PMC 1213230 , PMID 4217303
- ^ Стелла А. Кроссли (1974), «Изменения в брачном поведении, вызванные отбором для этологической изоляции между черным деревом и рудиментарными мутантами Drosophila melanogaster », Evolution , 28 (4): 631–647, doi : 10.1111/j.1558-5646.1974. tb00795.x , PMID 28564833 , S2CID 35867118
- ^ Ф.Р. ван Дейкен; В. Шарлоо (1979), «Дивергентный отбор по двигательной активности Drosophila melanogaster . I. Реакция отбора», Behavior Genetics , 9 (6): 543–553, doi : 10.1007/BF01067350 , PMID 122270 , S2CID 39352792
- ^ Ф.Р. ван Дейкен; В. Шарлоо (1979), «Дивергентный отбор по двигательной активности Drosophila melanogaster . II. Тест на репродуктивную изоляцию между выбранными линиями», Behavior Genetics , 9 (6): 555–561, doi : 10.1007/BF01067351 , PMID 122271 , S2CID 40169222
- ^ Б. Уоллес (1953), «Генетическая дивергенция изолированных популяций Drosophila melanogaster », Труды Девятого Международного конгресса генетиков , 9 : 761–764.
- ^ ГР Найт; и др. (1956), «Отбор на сексуальную изоляцию внутри вида», Evolution , 10 : 14–22, doi : 10.1111/j.1558-5646.1956.tb02825.x , S2CID 87729275 .
- ^ Форбс В. Робертсон (1966), «Тест сексуальной изоляции у дрозофилы », Genetical Research , 8 (2): 181–187, doi : 10.1017/s001667230001003x , PMID 5922518
- ^ Форбс В. Робертсон (1966), «Экологическая генетика роста дрозофилы 8. Адаптация к новой диете», Genetical Research , 8 (2): 165–179, doi : 10.1017/s0016672300010028 , PMID 5922517
- ^ Эллен Э. Хостерт (1997), «Подкрепление: новый взгляд на старое противоречие», Evolution , 51 (3): 697–702, doi : 10.1111/j.1558-5646.1997.tb03653.x , PMID 28568598 , S2CID 21054233
- ^ Кореф Сантибаньес, С.; Уоддингтон, CH (1958), «Происхождение половой изоляции между разными линиями внутри вида», Evolution , 12 (4): 485–493, doi : 10.2307/2405959 , JSTOR 2405959.
- ^ Баркер, JSF; Камминс, Л.Дж. (1969), «Эффект отбора по числу грудиноплевральных щетинок на брачное поведение Drosophila melanogaster », Genetics , 61 (3): 713–719, doi : 10.1093/genetics/61.3.713 , PMC 1212235 , PMID 17248436
- ^ Марков, Т.А. (1975), «Генетический анализ фототаксического поведения Drosophila melanogaster », Genetics , 79 (3): 527–534, doi : 10.1093/genetics/79.3.527 , PMC 1213291 , PMID 805084
- ^ Марков, Т.А. (1981), «Предпочтения спаривания не предсказывают направление эволюции экспериментальных популяций дрозофилы », Science , 213 (4514): 1405–1407, Бибкод : 1981Sci...213.1405M , doi : 10.1126/science .213.4514.1405 , PMID 17732575 , S2CID 15497733
- ^ Рандл, HD; Моерс, А. О.; Уитлок, MC (1998), «События массового притока одиночных основателей и эволюция репродуктивной изоляции», Evolution , 52 (6): 1850–1855, doi : 10.1111/j.1558-5646.1998.tb02263.x , JSTOR 2411356 , PMID 28565304 , S2CID 24502821
- ^ Моерс, А. О.; Рандл, HD; Уитлок, MC (1999), «Влияние отбора и узких мест на успех спаривания самцов у периферических изолятов», American Naturalist , 153 (4): 437–444, doi : 10.1086/303186 , PMID 29586617 , S2CID 4411105
- ^ Ли Эрман (1971), «Естественный отбор и происхождение репродуктивной изоляции», American Naturalist , 105 (945): 479–483, doi : 10.1086/282739 , S2CID 85401244
- ^ Ли Эрман (1973), «Подробнее о естественном отборе и происхождении репродуктивной изоляции», American Naturalist , 107 (954): 318–319, doi : 10.1086/282835 , S2CID 83780632
- ^ Ли Эрман (1979), «Еще больше о естественном отборе и происхождении репродуктивной изоляции», American Naturalist , 113 : 148–150, doi : 10.1086/283371 , S2CID 85237458
- ^ Ли Эрман (1983), «Четвертый отчет о естественном отборе в целях возникновения репродуктивной изоляции», American Naturalist , 121 (3): 290–293, doi : 10.1086/284059 , S2CID 83654887
- ^ Джон Ринго; Дэвид Вуд; Роберт Роквелл; Гарольд Доуз (1985), «Эксперимент по проверке двух гипотез видообразования», The American Naturalist , 126 (5): 642–661, doi : 10.1086/284445 , S2CID 84819968
- ^ Т. Добжанский; О. Павловский; Дж. Р. Пауэлл (1976), «Частично успешная попытка усилить репродуктивную изоляцию между полувидами Drosophila paulistorum », Evolution , 30 (2): 201–212, doi : 10.2307/2407696 , JSTOR 2407696 , PMID 28563045
- ^ Т. Добжанский; О. Павловский (1966), «Спонтанное возникновение зарождающегося вида в комплексе Drosophila paulistorum », PNAS , 55 (4): 723–733, Bibcode : 1966PNAS...55..727D , doi : 10.1073/pnas.55.4 .727 , PMC 224220 , PMID 5219677
- ^ Алиса Калиш де Оливейра; Антонио Родригес Кордейро (1980), «Адаптация экспериментальных популяций Drosophila willistoni к среде с экстремальным pH», Heredity , 44 : 123–130, doi : 10.1038/hdy.1980.11
- ^ Л. Эрман (1964), «Генетическая дивергенция в экспериментальных популяциях М. Ветухова Drosophila pseudoobscura », Genetical Research , 5 : 150–157, doi : 10.1017/s0016672300001099
- ^ М. Ветухова Л. Эрман (1969), «Генетическая дивергенция в экспериментальных популяциях Drosophila pseudoobscura . 5. Дальнейшее изучение зачатков половой изоляции», American Midland Naturalist , 82 (1): 272–276, doi : 10.2307/2423835 , АЭСТОР 2423835
- ^ Эдуардо дель Солар (1966), «Половая изоляция, вызванная отбором на положительный и отрицательный фототаксис и геотаксис у Drosophila pseudoobscura », Труды Национальной академии наук , 56 (2): 484–487, Бибкод : 1966PNAS...56. .484D , doi : 10.1073/pnas.56.2.484 , PMC 224398 , PMID 5229969
- ^ Джеффри Р. Пауэлл (1978), «Теория видообразования Основателя-Flush: экспериментальный подход», Evolution , 32 (3): 465–474, doi : 10.1111/j.1558-5646.1978.tb04589.x , JSTOR 2407714 , PMID 28567948 , S2CID 30943286
- ^ Дайан М.Б. Додд; Джеффри Р. Пауэлл (1985), «Видообразование-основатель: обновленные результаты экспериментов с дрозофилой », Evolution , 39 (6): 1388–1392, doi : 10.1111/j.1558-5646.1985.tb05704.x , JSTOR 2408795 , PMID 28564258 , S2CID 34137489
- ^ Галиана А.; Мойя, А.; со смывом-основателем Аяла, Ф.Дж. (1993), «Видообразование Drosophila pseudoobscura : крупномасштабный эксперимент», Evolution , 47 (2): 432–444, doi : 10.1111/j.1558-5646.1993.tb02104.x , JSTOR 2410062 , ПМИД 28568735 , С2КИД 42232235
- ^ Рандл, HD (2003), «Дивергентная среда и узкие места популяций не могут привести к изоляции перед сматыванием у Drosophila pseudoobscura », Evolution , 57 (11): 2557–2565, doi : 10.1554/02-717 , PMID 14686531 , S2CID 6162106
- ^ Карл Ф. Купман (1950), «Естественный отбор для репродуктивной изоляции между Drosophila pseudoobscura и Drosophila persimilis », Evolution , 4 (2): 135–148, doi : 10.2307/2405390 , JSTOR 2405390
- ^ Сеймур Кесслер (1966), «Отбор за и против этологической изоляции между Drosophila pseudoobscura и Drosophila persimilis », Evolution , 20 (4): 634–645, doi : 10.2307/2406597 , JSTOR 2406597 , PMID 28562900
- ^ Х. Роберта Кепфер (1987), «Отбор для сексуальной изоляции между географическими формами Drosophila mojavensis . I Взаимодействия между выбранными формами», Evolution , 41 (1): 37–48, doi : 10.2307/2408971 , JSTOR 2408971 , PMID 28563762
- ^ Этгес, В.Дж. (1998), «Изоляция перед спариванием определяется субстратами для выращивания личинок у кактофилы Drosophila mojavensis . IV. Коррелирующие реакции при поведенческой изоляции на искусственный отбор по признаку жизненного цикла», American Naturalist , 152 (1): 129–144. , doi : 10.1086/286154 , PMID 18811406 , S2CID 17689372
- ^ Лорна Х. Арита; Кеннет Ю. Канеширо (1979), «Этологическая изоляция между двумя группами дрозофилы Adiastola Hardy», Proc. Гавайи. Энтомол. Соц. , 13 : 31–34
- ^ Дж. Н. Ахерн (1980), «Эволюция поведенческой репродуктивной изоляции в лабораторных популяциях Drosophila silvestris », Experientia , 36 (1): 63–64, doi : 10.1007/BF02003975 , S2CID 43809774
- ^ А. Бенедикт Соанс; Дэвид Пиментел; Джойс С. Соанс (1974), «Эволюция репродуктивной изоляции в аллопатрических и симпатрических популяциях», The American Naturalist , 108 (959): 117–124, doi : 10.1086/282889 , S2CID 84913547
- ^ Л. Е. Херд; Роберт М. Айзенберг (1975), «Дивергентный отбор для геотаксической реакции и эволюция репродуктивной изоляции в симпатрических и аллопатрических популяциях комнатных мух», The American Naturalist , 109 (967): 353–358, doi : 10.1086/283002 , S2CID 85084378
- ^ Мефферт, Л.М.; Брайант, Э.Х. (1991), «Склонность к спариванию и ухаживающее поведение у линий комнатной мухи с серийными узкими местами», Evolution , 45 (2): 293–306, doi : 10.1111/j.1558-5646.1991.tb04404.x , JSTOR 2409664 , ПМИД 28567864 , S2CID 13379387
- ^ Такахиса Миятаке; Тору Симидзу (1999), «Генетические корреляции между историей жизни и поведенческими чертами могут вызвать репродуктивную изоляцию», Evolution , 53 (1): 201–208, doi : 10.2307/2640932 , JSTOR 2640932 , PMID 28565193
- ^ Патерниани, Э. (1969), «Отбор для репродуктивной изоляции между двумя популяциями кукурузы Zea mays L», Evolution , 23 (4): 534–547, doi : 10.1111/j.1558-5646.1969.tb03539.x , PMID 28562870 , S2CID 38650254
- ^ Андерс Один; Энн-Бритт Флорин (2002), «Половой отбор и перипатрическое видообразование: новый взгляд на модель Канеширо», Journal of Evolutionary Biology , 15 (2): 301–306, doi : 10.1046/j.1420-9101.2002.00378.x , S2CID 82095639
- ^ Jump up to: Перейти обратно: а б Дэвид Старр Джордан (1905), «Происхождение видов через изоляцию» , Science , 22 (566): 545–562, Бибкод : 1905Sci....22..545S , doi : 10.1126/science.22.566.545 , PMID 17832412
- ^ Jump up to: Перейти обратно: а б с д Джеймс Маллет (2010): «Почему биологи двадцатого века отвергли взгляд Дарвина на виды?» , Biology & Philosophy , 25 (4): 497–527, doi : 10.1007/s10539-010-9213-7 , S2CID 38621736.
- ^ Майр, Эрнст 1942. Систематика и происхождение видов . Издательство Колумбийского университета, Нью-Йорк. стр. 148
- ^ Дарвин, Чарльз (1859). О происхождении видов . Мюррей. п. 347.
- ^ Саллоуэй Ф.Дж. (1979). «Географическая изоляция в мышлении Дарвина: превратности решающей идеи». Исследования по истории биологии . 3 : 23–65. ПМИД 11610987 .
- ^ Дэвид Старр Джордан (1908), «Закон близнецовых видов», американский натуралист , 42 (494): 73–80, doi : 10.1086/278905
- ^ Джоэл Асаф Аллен (1907), «Мутации и географическое распространение близкородственных видов растений и животных», American Naturalist , 41 (490): 653–655, doi : 10.1086/278852
- ^ Эрнст Майр (1982), Рост биологической мысли , издательство Гарвардского университета, стр. 561–566, ISBN 978-0674364462
- ^ Jump up to: Перейти обратно: а б Джеймс Маллет (2001), «Революция видообразования», Журнал эволюционной биологии , 14 (6): 887–888, doi : 10.1046/j.1420-9101.2001.00342.x , S2CID 36627140
- ^ Jump up to: Перейти обратно: а б Кэтрин Х. Грэм; Сантьяго Р. Рон Хуан К. Сантос; Кристофер Дж. Шнайдер; Крейг Мориц (2004), «Интеграция моделей филогенетики и экологической ниши для изучения механизмов видообразования у дендробатидных лягушек», Evolution , 58 (8): 1781–1793, doi : 10.1554/03-274 , PMID 15446430 , S2CID 198157565
- ^ К. Йессон; Н. Х. Туми; А. Калхэм (2009), «Цикламен: биогеография времени, моря и видообразования с использованием филогении, калиброванной по времени», Journal of Biogeography , 36 (7): 1234–1252, Бибкод : 2009JBiog..36.1234Y , doi : 10.1111/j. 1365-2699.2008.01971.x , S2CID 85573501
- ^ Роберт М. Зинк (2012), «География видообразования: тематические исследования птиц», «Эволюция: образование и информационно-пропагандистская деятельность » , 5 (4): 541–546, doi : 10.1007/s12052-012-0411-4
- ^ РТ Чессер; RM Zink (1994), «Способы видообразования у птиц: проверка метода Линча», Evolution , 48 (2): 490–497, doi : 10.2307/2410107 , JSTOR 2410107 , PMID 28568302
- ^ Мэтью Т. Вебстер (2009), «Закономерности аутосомных расхождений между геномами человека и шимпанзе подтверждают аллопатрическую модель видообразования», Gene , 443 (1–2): 70–75, doi : 10.1016/j.gene.2009.05. 006 , ПМИД 19463924
- ^ Тейлор Эдвардс; Марк Толлис; Пин Сун Се; Райан Н. Гутенкунст; Чжэнь Лю; Кенро Кусуми ; Мелани Калвер; Роберт В. Мерфи (2016), «Оценка моделей видообразования в различных биогеографических сценариях; эмпирическое исследование с использованием мультилокусного анализа и анализа секвенирования РНК», Ecology and Evolution , 6 (2): 379–396, doi : 10.1002/ece3 .1865 , PMC 4729248 , PMID 26843925
Дальнейшее чтение [ править ]
Математические модели репродуктивной изоляции
- Х. Аллен Орр; Майкл Турелли (2001), «Эволюция постзиготической изоляции: накопление несовместимости Добжанского-Мюллера», Evolution , 55 (6): 1085–1094, arXiv : 0904.3308 , doi : 10.1554/0014-3820(2001)055[1085:teopia ]2.0.co;2 , PMID 11475044 , S2CID 198153495
- Х. Аллен Орр; Линн Х. Орр (1996), «Ожидание видообразования: влияние подразделения популяции на время видообразования», Evolution , 50 (5): 1742–1749, doi : 10.2307/2410732 , JSTOR 2410732 , PMID 28565607
- Х. Аллен Орр (1995), «Популяционная генетика видообразования: эволюция гибридной несовместимости», Genetics , 139 (4): 1805–1813, doi : 10.1093/genetics/139.4.1805 , PMC 1206504 , PMID 7789779
- Масатоши Нэй; Такео Маруяма; Чунг-и Ву (1983), «Модели эволюции репродуктивной изоляции», Genetics , 103 (3): 557–579, doi : 10.1093/genetics/103.3.557 , PMC 1202040 , PMID 6840540
- Масатоши Ней (1976), «Математические модели видообразования и генетической дистанции», Популяционная генетика и экология : 723–766.