Эволюция глаза
Многие ученые нашли эволюцию глаза привлекательной для изучения, поскольку глаз является ярким примером аналогичного органа, обнаруженного у многих животных . Простое обнаружение света обнаружено у бактерий, одноклеточных организмов, растений и животных. Сложные глаза, формирующие изображение, несколько раз эволюционировали независимо. [1]
Разнообразные глаза известны из сланцев Бёрджесс среднего кембрия и из немного более древних сланцев Эму Бэй . [2] Глаза различаются по остроте зрения , диапазону длин волн, которые они могут обнаружить, чувствительности в темноте, способности обнаруживать движение или различать объекты, а также по способности различать цвета .
История исследований [ править ]
В 1802 году философ Уильям Пейли назвал это чудом « дизайна ». В 1859 году сам Чарльз Дарвин писал в своей книге «Происхождение видов» , что эволюция глаза путем естественного отбора кажется на первый взгляд «в высшей степени абсурдной». [3] Однако он продолжал, что, несмотря на трудность вообразить это, его эволюция вполне осуществима:
... если можно показать, что существуют многочисленные градации от простого и несовершенного глаза до одного сложного и совершенного, причем каждая степень полезна для своего владельца, что, безусловно, так и есть; если, кроме того, глаз когда-либо изменяется и эти вариации передаются по наследству, что, несомненно, имеет место, и если такие вариации должны быть полезны какому-либо животному в изменяющихся условиях жизни, тогда трудность поверить в то, что совершенный и сложный глаз может быть образован путем естественный отбор,хотя и непреодолимое для нашего воображения, его не следует рассматривать как подрыв теории. [3]
Он предложил ступенчатую эволюцию от «зрительного нерва, просто покрытого пигментом и без какого-либо другого механизма» к «умеренно высокой стадии совершенства» и привел примеры существующих промежуточных состояний. [3] Текущие исследования изучают генетические механизмы, лежащие в основе развития и эволюции глаз. [4]
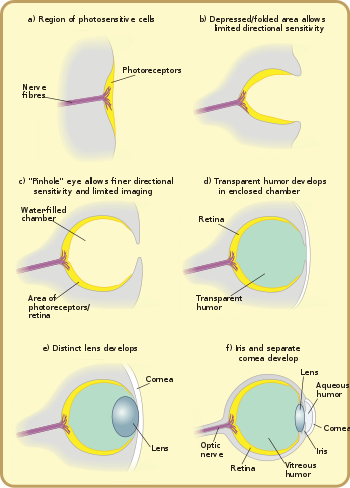
Биолог Д. Е. Нильссон независимо выдвинул теорию о четырех основных стадиях эволюции глаза позвоночного из участка фоторецепторов. [5] Нильссон и С. Пельгер в классической статье подсчитали, что для развития сложного глаза у позвоночных требуется всего несколько сотен тысяч поколений. [6] Другой исследователь, Дж. К. Янг, использовал летопись окаменелостей, чтобы сделать выводы об эволюции, основываясь на строении глазных орбит и отверстий в окаменелых черепах, через которые проходят кровеносные сосуды и нервы. [7] Все это увеличивает количество доказательств, подтверждающих теорию Дарвина.
Скорость эволюции [ править ]
Первые возможные окаменелости глаз, обнаруженные на сегодняшний день, относятся к эдиакарскому периоду (около 555 миллионов лет назад ). [8] в то время как самый старый окаменелый глаз принадлежит окаменелости Schmidtiellus reetae возрастом 530 млн лет назад, собранной в Савиранна на севере Эстонии. Строение похоже на сложные глаза современных стрекоз и пчел, но с (~ 100) омматидиями, расположенными дальше друг от друга, и без хрусталика . [9] [10] В нижнем кембрии произошел взрыв явно быстрой эволюции, названный « кембрийским взрывом ». Одной из многих гипотез «причин» кембрийского взрыва является теория «выключателя света» Эндрю Паркера : она утверждает, что эволюция развитых глаз положила начало гонке вооружений , которая ускорила эволюцию. [11] До кембрийского взрыва животные, возможно, чувствовали свет, но не использовали его для быстрого передвижения или навигации с помощью зрения.
Скорость эволюции глаз трудно оценить, поскольку летопись окаменелостей, особенно нижнего кембрия, скудна. Насколько быстро круглый участок фоторецепторных клеток может превратиться в полностью функциональный глаз позвоночного, оценивалось на основе скорости мутаций, относительного преимущества для организма и естественного отбора. Однако время, необходимое для каждого состояния, постоянно переоценивалось, а время генерации было установлено равным одному году, что часто встречается у мелких животных. Даже при таких пессимистических оценках глаз позвоночных все же мог эволюционировать из участка фоторецепторных клеток менее чем за 364 000 лет. [12] [примечание 1]
Происхождение глаза [ править ]
Эволюционировал ли глаз один или несколько раз, зависит от определения глаза. Все глазные животные имеют общий генетический механизм развития глаз. Это говорит о том, что предки глазных животных имели некую форму светочувствительного механизма, даже если это не был специальный оптический орган. Однако даже фоторецепторные клетки могли более одного раза эволюционировать из молекулярно сходных хеморецепторных клеток. Вероятно, фоторецепторные клетки существовали задолго до кембрийского взрыва. [13] Сходства более высокого уровня - такие как использование белка кристаллина в независимо полученных линзах головоногих и позвоночных. [14] – отражают возможность использования более фундаментального белка для выполнения новой функции в глазу. [15]
Общей чертой, общей для всех светочувствительных органов, являются опсины . Опсины принадлежат к семейству светочувствительных белков и делятся на девять групп, которые уже существовали у урбилатерий , последнего общего предка всех билатерально-симметричных животных . [16] Кроме того, генетический набор инструментов для позиционирования глаз является общим для всех животных: ген PAX6 контролирует развитие глаз у животных, начиная от осьминогов. [17] мышам и фруктовым мушкам . [18] [19] [20] Такие гены высокого уровня, как следствие, намного старше многих структур, которые они контролируют сегодня; изначально они, должно быть, служили другой цели, прежде чем их стали использовать для развития глаз. [15]
Глаза и другие органы чувств, вероятно, развились раньше мозга: нет необходимости в органе обработки информации (мозге), прежде чем появится информация для обработки. [21] Живым примером являются кубозойные медузы, у которых глаза сравнимы с позвоночных и головоногих глазами камеры , несмотря на отсутствие мозга. [22]
Этапы эволюции [ править ]

Самыми ранними предшественниками глаза были белки-фоторецепторы, воспринимающие свет, обнаруженные даже у одноклеточных организмов, называемые « глазными пятнами ». [23] Глазные пятна способны ощущать только окружающую яркость: они способны отличать свет от темноты, достаточной для фотопериодизма и суточной синхронизации циркадных ритмов . Их недостаточно для зрения, так как они не могут различать формы или определять направление света. Глазные пятна встречаются почти у всех основных групп животных и распространены среди одноклеточных организмов, включая эвглену . Глазное пятно эвглены, называемое рыльцем , расположено на ее переднем конце. Это небольшое пятно красного пигмента, которое оттеняет совокупность светочувствительных кристаллов. Вместе с ведущим жгутиком глазное пятно позволяет организму двигаться в ответ на свет, часто к свету, чтобы способствовать фотосинтезу . [24] и предсказывать день и ночь — основную функцию циркадных ритмов. Зрительные пигменты расположены в мозгу более сложных организмов и, как полагают, играют роль в синхронизации нереста с лунными циклами. Обнаружив тонкие изменения в ночном освещении, организмы могли синхронизировать высвобождение сперматозоидов и яйцеклеток, чтобы максимизировать вероятность оплодотворения. [25]
Само зрение зависит от базовой биохимии, общей для всех глаз. Однако то, как этот биохимический набор инструментов используется для интерпретации окружающей среды организма, широко варьируется: глаза имеют широкий спектр структур и форм, каждая из которых развилась довольно поздно по сравнению с лежащими в основе белками и молекулами. [24]
На клеточном уровне существует два основных типа глаз: один у протостомов ( моллюски , кольчатые черви и членистоногие ), другой у вторичноротых ( хордовые и иглокожие ). [24]
Функциональной единицей глаза является фоторецепторная клетка, которая содержит белки опсины и реагирует на свет, инициируя нервный импульс. Светочувствительные опсины располагаются на волосистом слое, чтобы максимально увеличить площадь поверхности. Природа этих «волосков» различается: в основе структуры фоторецепторов лежат две основные формы: микроворсинки и реснички . [26] В глазах протостом это микроворсинки: расширения или выпячивания клеточной мембраны. Но в глазах вторичноротых они происходят из ресничек, которые представляют собой отдельные структуры. [24] Однако вне глаз организм может использовать другой тип фоторецепторных клеток, например, моллюск Platynereis dumerilii использует микроворсинки в глазах, но дополнительно имеет ресничные фоторецепторные клетки глубокого мозга. [27] Фактическое происхождение может быть более сложным, поскольку некоторые микроворсинки содержат следы ресничек, но другие наблюдения, по-видимому, подтверждают фундаментальное различие между протостомами и вторичноротыми. [24] Эти соображения сосредоточены на реакции клеток на свет: некоторые используют натрий, чтобы вызвать электрический сигнал, который формирует нервный импульс, а другие используют калий; кроме того, протостомы в целом создают сигнал, пропуская больше натрия через свои клеточные стенки, тогда как вторичноротые пропускают меньше. [24]
Это говорит о том, что когда две линии разошлись в докембрии, у них были только очень примитивные световые рецепторы, которые независимо развились в более сложные глаза.
Ранние глаза [ править ]
Основной единицей обработки света в глазах является фоторецепторная клетка , специализированная клетка, содержащая два типа молекул, связанных друг с другом и расположенных в мембране: опсин , светочувствительный белок ; и хромофор , — пигмент поглощающий свет. Группы таких клеток называются «глазными пятнами» и развивались независимо где-то от 40 до 65 раз. Эти глазные пятна позволяют животным получить лишь базовое представление о направлении и интенсивности света, но недостаточно, чтобы отличить объект от его окружения. [24]
Разработка оптической системы, которая могла бы различать направление света с точностью до нескольких градусов, очевидно, гораздо сложнее, и только шесть из тридцати с лишним типов [примечание 2] иметь такую систему. Однако на эти типы приходится 96% ныне живущих видов. [24]

Эти сложные оптические системы возникли, когда многоклеточная глазная повязка постепенно превратилась в чашку, что сначала дало возможность различать яркость по направлениям, а затем по мере углубления ямы - во все более и более тонких направлениях. В то время как плоские глазные повязки были неэффективны для определения направления света, поскольку луч света активировал один и тот же участок светочувствительных клеток независимо от его направления, форма «чашечки» ямочных глаз допускала ограниченную направленную дифференциацию, изменяя, какие клетки свет будет падать в зависимости от угла падения света. Ямочные глаза, возникшие к кембрийскому периоду, были замечены у древних улиток . [ нужны разъяснения ] и обнаружены у некоторых улиток и других современных беспозвоночных, таких как планарии . Планарии могут слегка различать направление и интенсивность света благодаря своим чашеобразным, сильно пигментированным клеткам сетчатки , которые защищают светочувствительные клетки от воздействия во всех направлениях, за исключением единственного отверстия для света. Однако этот протоглаз все же гораздо полезнее для обнаружения отсутствия или присутствия света, чем его направления; это постепенно меняется по мере углубления глазной ямки и роста количества фоторецепторных клеток, что позволяет получать более точную визуальную информацию. [28]
Когда фотон поглощается хромофором, химическая реакция приводит к преобразованию энергии фотона в электрическую энергию и передаче у высших животных в нервную систему . Эти фоторецепторные клетки составляют часть сетчатки , тонкого слоя клеток, передающего зрительную информацию. [29] включая информацию об освещении и продолжительности светового дня, необходимую системе циркадных ритмов, в мозг. Однако у некоторых медуз , таких как Cladonema ( Cladonematidae ), есть сложные глаза, но нет мозга. Их глаза передают сообщение непосредственно мышцам без промежуточной обработки со стороны мозга. [21]
Во время кембрийского взрыва развитие глаза резко ускорилось благодаря радикальным улучшениям в обработке изображений и определении направления света. [30]

После того, как область светочувствительных клеток инвагинировалась , наступил момент, когда уменьшение ширины светового отверстия стало более эффективным для увеличения визуального разрешения, чем дальнейшее углубление чаши. [12] Уменьшив размер отверстия, организмы достигли истинного изображения, что позволило точно определять направление и даже в некоторой степени определять форму. Глаза такого типа в настоящее время встречаются у наутилуса . Не имея роговицы или хрусталика, они обеспечивают плохое разрешение и тусклое изображение, но по-прежнему с точки зрения зрения являются значительным улучшением по сравнению с ранними повязками на глазах. [31]
Разрастание прозрачных клеток предотвращало загрязнение и паразитарное заражение. Содержимое камеры, теперь разделенное, может постепенно превращаться в прозрачную жидкость для таких оптимизаций, как цветная фильтрация, более высокий показатель преломления , блокировка ультрафиолетового излучения или способность работать в воде и на суше. В некоторых классах этот слой может быть связан с линькой панциря или кожи организма. Пример этого можно наблюдать у Onychophorans , у которых кутикула панциря продолжается до роговицы. Роговица состоит из одного или двух кутикулярных слоев в зависимости от того, как давно животное линяло. [32] Наряду с хрусталиком и двумя юморами, роговица отвечает за сведение света и помощь в его фокусировке на задней части сетчатки. Роговица защищает глазное яблоко и в то же время обеспечивает примерно 2/3 общей преломляющей силы глаза. [33]
Вероятно, основная причина, по которой глаза специализируются на обнаружении определенного, узкого диапазона длин волн электромагнитного спектра ( видимого спектра ), заключается в том, что самые ранние виды, развившие светочувствительность, были водными, а вода фильтрует электромагнитное излучение, за исключением диапазона длин волн. , более короткие из которых мы называем синими, а более длинные волны мы идентифицируем как красные. Это же светофильтрационное свойство воды повлияло и на светочувствительность растений. [34] [35] [36]
Формирование и диверсификация линз [ править ]

глаза В безлинзовом глазу свет, исходящий из удаленной точки, попадает на заднюю часть глаза примерно того же размера, что и апертура . При добавлении линзы этот падающий свет концентрируется на меньшей площади поверхности, не уменьшая общую интенсивность раздражителя. [6] Фокусное расстояние раннего лобопода с простыми глазами, содержащими линзы, фокусировало изображение за сетчаткой, поэтому, хотя никакая часть изображения не могла быть сфокусирована, интенсивность света позволяла организму видеть в более глубоких (и, следовательно, более темных) водах. . [32] линзы Последующее увеличение показателя преломления , вероятно, привело к формированию сфокусированного изображения. [32]
Обратите внимание, что эта оптическая схема не была найдена и не ожидается. Окаменелость редко сохраняет мягкие ткани, и даже если бы это произошло, новая жидкость почти наверняка закрылась бы по мере высыхания останков или из-за того, что осадочные породы заставили слои соединиться, сделав окаменелый глаз похожим на предыдущую структуру.
Кристаллины [ править ]

позвоночных Хрусталики состоят из адаптированных эпителиальных клеток, которые имеют высокие концентрации белка кристаллина . Эти кристаллины принадлежат к двум основным семействам: α-кристаллинам и βγ-кристаллинам. Обе категории белков первоначально использовались для других функций в организмах, но со временем адаптировались для зрения глаз животных. [37] У эмбриона хрусталик представляет собой живую ткань, но клеточный механизм непрозрачен, поэтому его необходимо удалить, прежде чем организм сможет видеть. Удаление механизма означает, что хрусталик состоит из мертвых клеток, заполненных кристаллинами. Эти кристаллины особенные, поскольку они обладают уникальными характеристиками, необходимыми для прозрачности и функционирования хрусталика, такими как плотная упаковка, устойчивость к кристаллизации и чрезвычайная долговечность, поскольку они должны сохраняться на протяжении всей жизни организма. [37] Градиент показателя преломления , который делает линзу полезной, вызван радиальным сдвигом концентрации кристаллинов в разных частях хрусталика, а не конкретным типом белка: дело не в наличии кристаллина, а в его относительном распределении. делает линзу полезной. [38]
Биологически сложно поддерживать прозрачный слой клеток. [39] Отложение прозрачного неживого материала облегчило необходимость в поставке питательных веществ и удалении отходов. Распространено предположение, что трилобиты использовали кальцит , минерал, который, как сегодня известно, используется для зрения только у одного вида хрупких звезд . [40] Исследования глаз окаменелостей журавлей возрастом 55 миллионов лет из формации Фур показывают, что кальцит в глазах трилобитов является результатом тафономических и диагенетических процессов, а не оригинальной особенностью. [41] В других сложных глазах [ нужна проверка ] и глаза камеры, материал кристаллин . Зазор между слоями ткани естественным образом образует двояковыпуклую форму, оптически и механически идеальную для веществ нормального [ нужны разъяснения ] показатель преломления. Двояковыпуклая линза обеспечивает не только оптическое разрешение, но и светосилу и способность работать при слабом освещении, поскольку теперь разрешение не зависит от размера отверстия, которое снова медленно увеличивается, без ограничений кровообращения.
оболочка и влага, радужная роговица Водянистая
Независимо друг от друга прозрачный слой и непрозрачный слой могут отделяться вперед от хрусталика: отдельные роговица и радужная оболочка . (Это может произойти до или после отложения кристаллов, или не произойти вообще.) Отделение переднего слоя снова образует влага, водянистую влагу . Это увеличивает преломляющую силу и снова облегчает проблемы с кровообращением. Формирование непрозрачного кольца позволяет увеличить количество кровеносных сосудов, улучшить кровообращение и увеличить размер глаз. Этот клапан по периметру линзы также маскирует оптические недостатки, которые чаще встречаются по краям линзы. Необходимость маскировать недостатки линз постепенно возрастает с увеличением кривизны и силы линзы, общего размера линзы и глаза, а также потребностей организма в разрешении и диафрагме, что обусловлено требованиями охоты или выживания. Сейчас этот тип функционально идентичен глазу большинства позвоночных, включая человека. Действительно, «основной рисунок глаз всех позвоночных одинаков». [42]
Другие разработки [ править ]
Цветовое зрение [ править ]
Этот раздел нуждается в дополнительных цитатах для проверки . ( Октябрь 2016 г. ) |
пять классов зрительных опсинов У позвоночных обнаружено . Все, кроме одного, развились до расхождения круглоротых и рыб. [43] Пять классов опсинов по-разному адаптируются в зависимости от встречающегося светового спектра. Когда свет проходит через воду , более длинные волны, такие как красный и желтый, поглощаются быстрее, чем более короткие волны зеленого и синего. Это создает градиент спектральной плотности мощности , при этом средняя длина волны становится короче по мере увеличения глубины воды. [44] Зрительные опсины рыб более чувствительны к диапазону света в их среде обитания и на глубине. Однако наземная среда не различается по составу длин волн, поэтому чувствительность к опсину у наземных позвоночных не сильно различается. Это напрямую способствует значительному присутствию коммуникативных цветов. [43] Цветное зрение дает явные селективные преимущества, такие как лучшее распознавание хищников, пищи и партнеров. Действительно, вполне возможно, что простые сенсорно-нейронные механизмы могут избирательно контролировать общие модели поведения, такие как бегство, поиск пищи и сокрытие. Было выявлено множество примеров поведения, зависящего от длины волны, в двух основных группах: ниже 450 нм, связанное с прямым светом, и выше 450 нм, связанное с отраженным светом. [45] Поскольку молекулы опсина были настроены на обнаружение различных длин волн света, в какой-то момент появилось цветовое зрение , когда фоторецепторные клетки начали использовать опсины с разной настройкой. [29] Это могло произойти на любой из ранних стадий эволюции глаза и могло исчезнуть и возобновиться по мере изменения относительного селективного давления на линию.
Поляризационное зрение [ править ]
Поляризация — это организация неупорядоченного света в линейные структуры, которая происходит, когда свет проходит через щелевые фильтры, а также при переходе в новую среду. Чувствительность к поляризованному свету особенно полезна для организмов, среда обитания которых расположена на глубине более нескольких метров под водой. В этой среде цветовое зрение менее надежно и, следовательно, является более слабым селективным фактором. Хотя большинство фоторецепторов обладают способностью различать частично поляризованный свет, мембраны наземных позвоночных ориентированы перпендикулярно, поэтому они нечувствительны к поляризованному свету. [46] Однако некоторые рыбы могут различать поляризованный свет, что свидетельствует о наличии у них линейных фоторецепторов. Кроме того, каракатицы способны воспринимать поляризацию света с высокой визуальной точностью, хотя, по-видимому, у них отсутствует какая-либо значительная способность различать цвета. [47] Как и цветовое зрение, чувствительность к поляризации может помочь организму различать окружающие объекты и людей. Из-за незначительной отражательной интерференции поляризованного света его часто используют для ориентации и навигации, а также для различения скрытых объектов, таких как замаскированная добыча. [46]
Механизм фокусировки [ править ]
Используя мышцу сфинктера радужной оболочки и цилиарное тело , некоторые виды перемещают хрусталик вперед и назад, некоторые растягивают хрусталик более плоскими. Другой механизм регулирует фокусировку химически и независимо от этих двух, контролируя рост глаза и поддерживая фокусное расстояние. Кроме того, форму зрачка можно использовать для прогнозирования используемой фокальной системы. Щелевидный зрачок может указывать на обычную мультифокальную систему, тогда как круглый зрачок обычно указывает на монофокальную систему. При использовании круглой формы зрачок сужается при ярком свете, увеличивая число f, и расширяется в темноте, чтобы уменьшить глубину резкости. [48] Обратите внимание, что метод фокусировки не является обязательным. Как известно фотографам, ошибки фокусировки увеличиваются с увеличением диафрагмы . Таким образом, бесчисленное множество организмов с маленькими глазами активны под прямыми солнечными лучами и выживают вообще без механизма фокусировки. По мере того, как вид становится больше или переходит в более тусклую среду, средства фокусировки появляются постепенно.
Размещение [ править ]
У хищников обычно глаза расположены на передней части головы, чтобы лучше воспринимать глубину и фокусироваться на добыче. Глаза животных-жертв, как правило, расположены по бокам головы, что обеспечивает широкое поле зрения для обнаружения хищников с любого направления. [49] [50] Камбала — хищники, которые лежат на боку на дне, а глаза расположены асимметрично на одной стороне головы. Переходной окаменелостью от обычного симметричного положения к асимметричному положению является амфистиум .
См. также [ править ]
Пояснительные сноски [ править ]
- ↑ Дэвид Берлински , сторонник разумного замысла, поставил под сомнение обоснованность расчетов, а автор оригинальной статьи опроверг критику Берлински.
- Берлински, Дэвид (апрель 2001 г.). «Недовольный глаз» . Комментарий .
- Нильссон, Дэн-Э. «Остерегайтесь псевдонауки: ответ на нападки Дэвида Берлински на мои расчеты того, сколько времени требуется глазу, чтобы развиться» . Архивировано из оригинала 15 сентября 2003 года.
- «Эволюция глаза» на PBS
- ^ Точное число варьируется от автора к автору.
Ссылки [ править ]
- ^ ""Важным сигналом для понимания эволюции глаз является различие между различными типами фоторецепторных клеток. Сальвини-Плавен и Майр (1977) отметили замечательное разнообразие морфологии фоторецепторных клеток в животном мире и предположили, что фоторецепторы много раз развивались независимо » . ред.), Oxford University Press, Оксфорд (2012), стр. 7.
- ^ Ли, MSY; Джаго, Дж.Б.; Гарсия-Беллидо, округ Колумбия; Эджкомб, GE; Гелинг, Дж. Г.; Патерсон, младший (2011). «Современная оптика в исключительно сохранившихся глазах раннекембрийских членистоногих из Австралии». Природа . 474 (7353): 631–634. Бибкод : 2011Natur.474..631L . дои : 10.1038/nature10097 . ПМИД 21720369 . S2CID 4417647 .
- ^ Jump up to: Перейти обратно: а б с Дарвин, Чарльз (1859). О происхождении видов . Лондон, Великобритания: Джон Мюррей.
- ^ Геринг, WJ (2005). «Новые взгляды на развитие глаз и эволюцию глаз и фоторецепторов» (PDF) . Дж. Херед . 96 (3): 171–84. doi : 10.1093/jhered/esi027 . ПМИД 15653558 .
- ^ Нильссон, Д.-Э. (2013). «Эволюция глаза и его функциональная основа» . Визуальная нейронаука . 30 (1–2): 5–20. дои : 10.1017/s0952523813000035 . ПМЦ 3632888 . ПМИД 23578808 .
- ^ Jump up to: Перейти обратно: а б Нильссон, Д.-Э.; Пелгер, С. (1994). «Пессимистическая оценка времени, необходимого для развития глаза». Труды Королевского общества B: Биологические науки . 256 (1345): 53–58. Бибкод : 1994РСПСБ.256...53Н . дои : 10.1098/rspb.1994.0048 . ПМИД 8008757 . S2CID 13061351 .
- ^ Янг, GC (2008). «Ранняя эволюция глаза позвоночных - ископаемые свидетельства» . Эво Эду Пропагандистская деятельность . 1 (4): 427–438. дои : 10.1007/s12052-008-0087-y . hdl : 1885/28128 .
- ^ Макменамин, Марк А.С. (2016). Динамическая палеонтология: использование количественной оценки и других инструментов для расшифровки истории жизни . Спрингер. ISBN 978-3-319-22776-4 .
- ^ Дин, Сигне (8 декабря 2017 г.). «Это ископаемое возрастом 530 миллионов лет может быть самым старым глазом, когда-либо обнаруженным» . НаукаАлерт . Проверено 29 октября 2021 г.
- ^ Дэйли, Джейсон. «Взгляните в глаз возрастом 530 миллионов лет, самый старый из когда-либо обнаруженных» . Смитсоновский журнал . Проверено 29 октября 2021 г.
- ^ Паркер, Эндрю (2003). В мгновение ока: как видение спровоцировало большой взрыв эволюции . Кембридж, Массачусетс: Паб «Персей». ISBN 978-0-7382-0607-3 .
- ^ Jump up to: Перейти обратно: а б Нильссон, Делавэр; Пелгер С. (1994). «Пессимистическая оценка времени, необходимого для развития глаза». Труды Королевского общества Б. 256 (1345): 53–58. Бибкод : 1994РСПСБ.256...53Н . дои : 10.1098/rspb.1994.0048 . ПМИД 8008757 . S2CID 13061351 .
- ^ Нильссон, Делавэр (1996). «Происхождение глаз: старые гены для новых глаз» . Современная биология . 6 (1): 39–42. Бибкод : 1996CBio....6...39N . дои : 10.1016/S0960-9822(02)00417-7 . ПМИД 8805210 .
- ^ Зиновьева Р.; Пятигорский Ю.; Томарев, С.И. (1999). «О-Кристаллин, аргининкиназа и ферритин из хрусталика осьминога» . Biochimica et Biophysical Acta (BBA) - Структура белка и молекулярная энзимология . 1431 (2): 512–517. дои : 10.1016/S0167-4838(99)00066-7 . ПМИД 10350626 .
- ^ Jump up to: Перейти обратно: а б Шотландия, RW (2010). «Глубокая гомология: взгляд из систематики». Биоэссе . 32 (5): 438–449. дои : 10.1002/bies.200900175 . ПМИД 20394064 . S2CID 205469918 .
- ^ Рамирес, доктор медицины; Пайретт, АН; Панки, М.С.; Серб, Ю.М.; Спейзер, Д.И.; Сваффорд, Эй Джей; Окли, TH (26 октября 2016 г.). «Последний общий предок большинства двусторонних животных обладал по крайней мере 9 опсинами» . Геномная биология и эволюция . 8 (12): 3640–3652. дои : 10.1093/gbe/evw248 . ПМЦ 5521729 . ПМИД 28172965 .
- ^ Ёсида, Масааки; Юра, Кей; Огура, Ацуши (5 марта 2014 г.). «Эволюция глаз головоногих моллюсков была модулирована путем приобретения вариантов сплайсинга Pax-6» . Научные отчеты . 4 : 4256. Бибкод : 2014NatSR...4E4256Y . дои : 10.1038/srep04256 . ПМК 3942700 . ПМИД 24594543 .
- ^ Гальдер, Г.; Каллаертс, П.; Геринг, WJ (1995). «Новые взгляды на эволюцию глаз». Текущее мнение в области генетики и развития . 5 (5): 602–609. дои : 10.1016/0959-437X(95)80029-8 . ПМИД 8664548 .
- ^ Гальдер, Г.; Каллаертс, П.; Геринг, В. (1995). «Индукция эктопических глаз путем целевой экспрессии безглазого гена у дрозофилы». Наука . 267 (5205): 1788–92. Бибкод : 1995Sci...267.1788H . дои : 10.1126/science.7892602 . ПМИД 7892602 .
- ^ Томарев С.И.; Каллаертс, П.; Кос, Л.; Зиновьева Р.; Гальдер, Г.; Геринг, В.; Пятигорский, Дж. (1997). «Кальмар Пакс-6 и развитие глаз» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (6): 2421–2426. Бибкод : 1997PNAS...94.2421T . дои : 10.1073/pnas.94.6.2421 . ЧВК 20103 . ПМИД 9122210 .
- ^ Jump up to: Перейти обратно: а б Геринг, WJ (13 января 2005 г.). «Новые перспективы развития глаз и эволюции глаз и фоторецепторов» (PDF) . Журнал наследственности . 96 (3): 171–184. doi : 10.1093/jhered/esi027 . ПМИД 15653558 .
- ^ Коутс, Мелисса М. (2003). «Визуальная экология и функциональная морфология кубозоа (Cnidaria)» . Интегративная и сравнительная биология . 43 (4): 542–548. дои : 10.1093/icb/43.4.542 . ISSN 1540-7063 . ПМИД 21680462 .
- ^ Шваб, ИК (февраль 2018 г.). «Эволюция глаз: основные шаги. Лекция Килера 2017: столетие компании Keeler Ltd» . Глаз . 32 (2): 302–313. дои : 10.1038/eye.2017.226 . ISSN 0950-222X . ПМЦ 5811732 . ПМИД 29052606 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час МФ Ленд; РД Фернальд (1992). «Эволюция глаз». Ежегодный обзор неврологии . 15 : 1–29. дои : 10.1146/annurev.ne.15.030192.000245 . ПМИД 1575438 .
- ^ Кабальес, Сьемон Франк; Пратчетт, Морган С. (29 марта 2017 г.). «Экологические и биологические сигналы нереста морской звезды тернового венца» . ПЛОС ОДИН . 12 (3): e0173964. Бибкод : 2017PLoSO..1273964C . дои : 10.1371/journal.pone.0173964 . ISSN 1932-6203 . ПМЦ 5371309 . ПМИД 28355236 .
- ^ Отрум, Х (1979). "Введение". В Х. Отруме (ред.). Сравнительная физиология и эволюция зрения у беспозвоночных-А: Фоторецепторы беспозвоночных . Справочник по сенсорной физиологии. Том. VII/6А. Нью-Йорк: Springer-Verlag. стр. 6–9. ISBN 978-3-540-08837-0 .
- ^ Арендт, Д.; Тессмар-Райбл, К.; Снайман, Х.; Доррестейн, AW; Витбродт, Дж. (29 октября 2004 г.). «Цилиарные фоторецепторы с опсином позвоночного типа в мозге беспозвоночных». Наука . 306 (5697): 869–871. Бибкод : 2004Sci...306..869A . дои : 10.1126/science.1099955 . ПМИД 15514158 . S2CID 2583520 .
- ^ Эволюция глаз? Архивировано 15 сентября 2012 года в Wayback Machine.
- ^ Jump up to: Перейти обратно: а б Фернальд, Рассел Д. (2001). Эволюция глаз: как глаза улавливают фотоны? Архивировано 19 марта 2006 г. в Wayback Machine Karger Gazette 64: «Глаз в фокусе».
- ^ Конвей-Моррис, С. (1998). Горнило Творения . Оксфорд: Издательство Оксфордского университета.
- ^ Докинз, Ричард (1986). Слепой часовщик .
- ^ Jump up to: Перейти обратно: а б с Шенеманн, Б.; Лю, JN; Шу, Д.Г.; Хан, Дж.; Чжан, ZF (2008). «Миниатюрная оптимизированная зрительная система в нижнем кембрии». Летайя . 42 (3): 265–273. дои : 10.1111/j.1502-3931.2008.00138.x .
- ^ Али, Массачусетс и М.А. Клайн. 1985. Зрение у позвоночных. Нью-Йорк: Пленум Пресс
- ^ Фернальд, Рассел Д. (2001). Эволюция глаз: почему мы видим то, что видим? Архивировано 19 марта 2006 г. в Wayback Machine Karger Gazette 64: «Глаз в фокусе».
- ^ Фернальд, Рассел Д. (1998). Водные адаптации рыбьего глаза . Нью-Йорк, Спрингер.
- ^ Фернальд Р.Д. (1997). «Эволюция глаз». Мозговое поведение. Эвол . 50 (4): 253–9. дои : 10.1159/000113339 . ПМИД 9310200 .
- ^ Jump up to: Перейти обратно: а б Слингсби, К.; Вистоу, Дж.Дж.; Кларк, Арканзас (2013). «Эволюция кристаллинов, играющих роль в хрусталике глаза позвоночных» . Белковая наука . 22 (4): 367–380. дои : 10.1002/pro.2229 . ПМК 3610043 . ПМИД 23389822 .
- ^ Фернальд, Рассел Д. (2001). Эволюция глаз: откуда появились линзы? Karger Gazette 64: «Глаз в фокусе».
- ^ Инюшин Михаил; Мешалкина Дарья; Зуева, Лидия; Заяс-Сантьяго, Астрид (28 июня 2019 г.). «Прозрачность тканей in vivo» . Молекулы . 24 (13): 2388. doi : 10,3390/molecules24132388 . ISSN 1420-3049 . ПМК 6651221 . ПМИД 31261621 .
- ^ Берджесс, Дэниел С. (2001). Хрупкая звезда оснащена кальцитовыми линзами и спектрами фотоники
- ^ Линдгрен Дж., Нильссон Д.Э., Шёвалл П. и др. 2019. Глаза ископаемых насекомых проливают свет на оптику трилобитов и пигментный экран членистоногих. Природа 573, 122–125 (2019). https://doi.org/10.1038/s41586-019-1473-z
- ^ Али, Мохамед Атер; Клайн, Массачусетс (1985). Зрение у позвоночных . Нью-Йорк: Пленум Пресс. п. 1. ISBN 978-0-306-42065-8 .
- ^ Jump up to: Перейти обратно: а б Осорио, Д; Воробьев, М (2005). «Спектральная чувствительность фоторецепторов у наземных животных: адаптации к люминесценции и цветовому зрению» . Учеб. Р. Сок. Б. 272 (1574): 1745–52. дои : 10.1098/rspb.2005.3156 . ПМЦ 1559864 . ПМИД 16096084 .
- ^ Гуманн, Мартин; Цзя, Хён; Рэндел, Надин; Верасто, Чаба; Безарес-Кальдерон, Луи А.; Майклс, Нико К.; Ёкояма, Сёдзо; Жекели, Гаспар (август 2015 г.). «Спектральная настройка фототаксиса го-опсином в рабдомерных глазах Platynereis» . Современная биология . 25 (17): 2265–2271. Бибкод : 2015CBio...25.2265G . дои : 10.1016/j.cub.2015.07.017 . ПМИД 26255845 .
- ^ Мензель, Рэндольф (1979). «Спектральная чувствительность и цветовое зрение у беспозвоночных». В Х. Отруме (ред.). Сравнительная физиология и эволюция зрения у беспозвоночных - А: Фоторецепторы беспозвоночных . Справочник по сенсорной физиологии. Том. VII/6А. Нью-Йорк: Springer-Verlag. стр. 504–506, 551–558. ISBN 978-3-540-08837-0 .
- ^ Jump up to: Перейти обратно: а б Кронин, Т; Шашар, Н; Колдуэлл, Р.; Маршалл, Дж; Чероске, А; Чиу, Т (2003). «Поляризационное зрение и его роль в биологической передаче сигналов» . Интегр. Комп. Биол . 43 (4): 549–58. CiteSeerX 10.1.1.570.2694 . дои : 10.1093/icb/43.4.549 . ПМИД 21680463 .
- ^ Метгер, Лидия М.; Барбоза, Александра; Майнер, Саймон; Хэнлон, Роджер Т. (1 мая 2006 г.). «Цветовая слепота и восприятие контраста у каракатиц (Sepia officinalis), определяемые с помощью визуального сенсомоторного анализа». Исследование зрения . 46 (11): 1746–1753. дои : 10.1016/j.visres.2005.09.035 . ПМИД 16376404 . S2CID 16247757 .
- ^ Мальстрем, Т; Крогер, Р. (2006). «Форма зрачка и оптика хрусталика в глазах наземных позвоночных». Журнал экспериментальной биологии .
- ^ «Хищники» . Министерство внутренних дел США, Бюро землеустройства. 14 декабря 2009 г. Архивировано из оригинала 14 июня 2011 г. Проверено 28 марта 2011 г.
- ^ Бородицкий, Лера (24 июня 1999 г.). «Свет и глаза: конспект лекций» . Конспекты лекций . Стэнфорд . Архивировано из оригинала 5 июля 2010 года . Проверено 11 мая 2010 г.
Дальнейшее чтение [ править ]
- Лэмб Т.Д., Коллин С.П., Пью Э.Н. (декабрь 2007 г.). «Эволюция глаза позвоночных: опсины, фоторецепторы, сетчатка и глазная чашечка» . Обзоры природы Неврология . 8 (12): 960–76. дои : 10.1038/nrn2283 . ПМК 3143066 . ПМИД 18026166 . Иллюстрация. Обзор
- Лэмб, Т.Д. (2011). «Эволюция глаза» (PDF) . Научный американец . 305 (1): 64–69. Бибкод : 2011SciAm.305f..64L . doi : 10.1038/scientificamerican0711-64 . Архивировано из оригинала (PDF) 12 декабря 2013 года . Проверено 28 апреля 2013 г.
- Лэнд, Майкл Ф.; Нильссон, Дэн-Эрик (2012). «Происхождение зрения». Глаза животных (2-е изд.). Оксфорд: Издательство Оксфордского университета. стр. 1–22. ISBN 978-0-19-958114-6 .
- Специальный выпуск «Эволюция глаз» в журнале «Эволюция: образование и информационно-просветительская деятельность» , том 1, номер 4 / октябрь 2008 г. 26 статей, свободный доступ.
- Иван Р. Шваб (2012). Свидетель эволюции: как развивались глаза . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-536974-8 .
- Хаякава С., Такаку Ю., Хван Дж.С., Хоригучи Т., Суга Х., Геринг В. и др. (2015). «Функция и эволюционное происхождение одноклеточной структуры глаза камерного типа» . ПЛОС ОДИН . 10 (3): e0118415. Бибкод : 2015PLoSO..1018415H . дои : 10.1371/journal.pone.0118415 . ПМЦ 4348419 . ПМИД 25734540 .
- Греэ, К. (1968). «Ультраструктурная организация птиц двух Peridiniens Warnowiidae, Erythropsis pavillardi Kofoid и Swezy и Warnovia pulchra Schiller». Протистологическая . 4 : 209–230.
- Грегори С. Гавелис; Сихо Хаякава; Ричард А. Уайт III; Такаси Годобори; Кертис А. Саттл; Патрик Дж. Килинг; Брайан С. Леандер (2015). «Глазоподобные оцеллоиды построены из различных эндосимбиотически приобретенных компонентов» . Природа . 523 (7559): 204–7. Бибкод : 2015Natur.523..204G . дои : 10.1038/nature14593 . hdl : 10754/566109 . ПМИД 26131935 . S2CID 4462376 . Архивировано из оригинала 12 апреля 2018 года . Проверено 2 июля 2019 г.
- Окли, Тодд Х.; Спейзер, Дэниел И. (2015). «Как возникает сложность: эволюция глаз животных» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 46 : 237–260. doi : 10.1146/annurev-ecolsys-110512-135907 .
- Эд Янг; фотографии Дэвида Лютшвагера (февраль 2016 г.). «Внутри глаза: самое изысканное творение природы» . Нэшнл Географик . 229 (2): 30–57. Архивировано из оригинала 24 июня 2017 года.
Внешние ссылки [ править ]
- «Эволюция глаза» . Образовательный фонд WGBH и Clear Blue Sky Productions . ПБС. 2001.
- Креационизм опровергнут? Видео Национального центра научного образования об эволюции глаза
- Эволюция: образование и информационно-просветительская деятельность. Специальный выпуск: «Эволюция и глаза» , том 1, номер 4, октябрь 2008 г., страницы 351–559. ISSN 1936-6426 (печать) 1936–6434 (онлайн)
Зрение у животных |
|---|
