Инфракрасное зондирование у змей
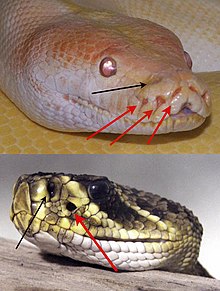
Способность воспринимать инфракрасное тепловое излучение независимо развилась у трех разных групп змей, состоящих из семейств Boidae (удавы), Pythonidae (питоны) и подсемейства Crotalinae (ямные гадюки). То, что обычно называют ямочным органом, позволяет этим животным, по сути, «видеть». [1] лучистое тепло на длинах волн от 5 до 30 мкм . Более развитое инфракрасное чутье гадюк позволяет этим животным точно поражать добычу даже в отсутствие света и обнаруживать теплые объекты на расстоянии нескольких метров. [2] [3] Ранее считалось, что эти органы развивались в первую очередь как детекторы добычи, но недавние данные свидетельствуют о том, что они также могут использоваться для терморегуляции и обнаружения хищников , что делает их сенсорным органом более общего назначения, чем предполагалось. [4] [5]
Филогения и эволюция
[ редактировать ]Лицевая ямка претерпела параллельную эволюцию у гадюк , некоторых удавов и питонов . Один раз он эволюционировал у гадюк и несколько раз у удавов и питонов. [6] Электрофизиология структурной структуры обеих линий схожа, но они различаются грубой анатомией . На первый взгляд, у гадюк имеется по одному большому ямочному органу по обе стороны головы, между глазом и ноздрей ( лореальные ямки ), в то время как удавы и питоны имеют три или более ямок сравнительно меньшего размера, выстилающих верхнюю, а иногда и нижнюю губу, внутри или между ними. чешуя (губные ямки). Змеиные змеи являются более продвинутыми, у них есть подвешенная сенсорная мембрана, а не простая структура ямок.
Анатомия
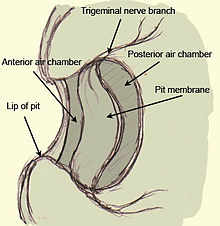
[ редактировать ]У гадюк тепловая яма представляет собой глубокий карман в роструме с перепонкой натянутой на него . За мембраной находится заполненная воздухом камера, обеспечивающая контакт с воздухом по обе стороны мембраны. Мембрана ямки хорошо васкуляризирована и сильно иннервирована многочисленными термочувствительными рецепторами, образованными терминальными образованиями тройничного нерва (терминальными нервными образованиями или TNM). Таким образом, рецепторы представляют собой не отдельные клетки, а часть самого тройничного нерва. Лабиальная ямка, обнаруженная у удавов и питонов, лишена подвешенной мембраны и состоит скорее из ямки, выстланной мембраной, которая имеет аналогичную иннервацию и сосуды, хотя морфология сосудистой сети различается у этих змей и кроталин . Целью сосудистой сети, помимо обеспечения кислородом терминалей рецепторов, является быстрое охлаждение рецепторов до их термонейтрального состояния после нагревания тепловым излучением раздражителя. Если бы не эта сосудистая сеть, рецептор оставался бы в теплом состоянии после воздействия теплого стимула и вызывал бы у животного остаточные изображения даже после того, как стимул был удален. [7]
Нейроанатомия
[ редактировать ]Во всех случаях лицевая ямка иннервируется тройничным нервом. У кроталинов информация из ямочного органа передается в ретикулярное ядро продолговатого мозга через латеральный нисходящий тройничный тракт. Оттуда он передается на контралатеральную покрышку зрительного нерва . У удавов и питонов информация из губной ямки передается непосредственно в контралатеральную покрышку зрительного нерва через латеральный нисходящий тройничный тракт, минуя ретикулярное ядро. [8]
Именно оптическая оболочка мозга в конечном итоге обрабатывает эти инфракрасные сигналы. Эта часть мозга также получает другую сенсорную информацию, в первую очередь оптическую стимуляцию, а также моторную, проприоцептивную и слуховую . Некоторые нейроны тектума реагируют только на визуальную или инфракрасную стимуляцию; другие сильнее реагируют на комбинированную визуальную и инфракрасную стимуляцию, а третьи реагируют только на комбинацию визуальной и инфракрасной стимуляции. Некоторые нейроны, по-видимому, настроены на обнаружение движения в одном направлении. Было обнаружено, что визуальные и инфракрасные карты мира змеи накладываются на зрительную покрышку. Эта объединенная информация передается через тектум в передний мозг. [9]
Нервные волокна в ямочном органе постоянно активируются с очень низкой скоростью. Объекты, находящиеся в нейтральном температурном диапазоне, не меняют темп стрельбы; нейтральный диапазон определяется средним тепловым излучением всех предметов, находящихся в рецептивном поле органа. Тепловое излучение выше заданного порога вызывает повышение температуры нервного волокна, что приводит к стимуляции нерва и последующему возбуждению, при этом повышение температуры приводит к увеличению скорости возбуждения. [10] Чувствительность нервных волокон оценивается на уровне <0,001 °C. [11]
Ямочный орган адаптируется к повторяющемуся раздражителю; если адаптированный стимул удалить, произойдет колебание в противоположном направлении. Например, если перед змеей поместить теплый предмет, то частота возбуждения органа сначала увеличится, но через некоторое время адаптируется к теплому предмету, и частота возбуждения нервов в ямочном органе вернется к норме. . Если затем этот теплый объект убрать, ямочный орган теперь зарегистрирует пространство, которое он занимал раньше, как более холодное, и поэтому скорость стрельбы будет снижаться до тех пор, пока он не адаптируется к удалению объекта. Латентный период адаптации составляет примерно от 50 до 150 мс. [10]
Лицевая ямка фактически визуализирует тепловое излучение, используя те же оптические принципы, что и камера-обскура , в которой местоположение источника теплового излучения определяется расположением излучения на мембране тепловой ямы. Однако исследования, в которых с помощью компьютерного анализа визуализировались тепловые изображения лицевой ямки, показали, что разрешение чрезвычайно низкое. Размер отверстия ямы приводит к плохому разрешению небольших теплых объектов, а в сочетании с небольшим размером ямы и последующей плохой теплопроводностью получаемое изображение имеет чрезвычайно низкое разрешение и контрастность. Известно, что некоторая фокусировка и резкость изображения происходит в латеральном нисходящем тройничном тракте, и возможно, что зрительная и инфракрасная интеграция, происходящая в покровной кишке, также может использоваться для повышения резкости изображения.
Молекулярный механизм
[ редактировать ]Несмотря на обнаружение инфракрасного света, механизм обнаружения инфракрасного излучения не похож на фоторецепторы - в то время как фоторецепторы обнаруживают свет посредством фотохимических реакций, белок в ямках змей представляет собой тип временного рецепторного потенциального канала , TRPA1 , который представляет собой термочувствительный ион. канал. Он воспринимает инфракрасные сигналы посредством механизма, включающего нагревание ямочного органа, а не химическую реакцию на свет. [12] По структуре и функциям он напоминает биологическую версию прибора для измерения тепла, называемого болометром . Это согласуется с очень тонкой мембраной ямки, которая позволяет входящему инфракрасному излучению быстро и точно нагревать данный ионный канал и запускать нервный импульс, а также васкуляризировать мембрану ямки, чтобы быстро охладить ионный канал обратно в исходное температурное состояние. Хотя молекулярные предшественники этого механизма обнаружены и у других змей, белок экспрессируется в гораздо меньшей степени и гораздо менее чувствителен к теплу. [12]
Поведенческие и экологические последствия
[ редактировать ]Змеи, чувствительные к инфракрасному излучению, широко используют ямочные органы для обнаружения и нацеливания на теплокровную добычу, такую как грызуны и птицы. Слепые или с завязанными глазами гремучие змеи могут точно поразить добычу при полном отсутствии видимого света. [13] [14] хотя не похоже, что они оценивают животных-жертв по температуре их тела. [15] Кроме того, змеи могут намеренно выбирать места для засад, которые облегчают обнаружение добычи с помощью инфракрасного излучения. [16] [17] Ранее предполагалось, что орган развился специально для захвата добычи. [11] Однако недавние данные показывают, что ямочный орган также может использоваться для терморегуляции. В эксперименте, в котором проверялась способность змей находить прохладное термальное убежище в неприятно жарком лабиринте, все ямные гадюки смогли найти убежище быстро и легко, в то время как настоящие гадюки не смогли этого сделать. Это говорит о том, что гадюки использовали свои ямочные органы для принятия решений по терморегуляции. [4] Также возможно, что орган мог даже развиться как защитная адаптация, а не хищническая, или что развитию органа могло способствовать множественное давление. [5] Использование тепловой ямы для управления терморегуляцией или другим поведением питонов и удавов еще не определено.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Ньюман, Э.А.; Хартлайн, штат Пенсильвания (1981). «Интеграция зрительной и инфракрасной информации в бимодальных нейронах зрительного покрова гремучей змеи» . Наука . 213 (4509): 789–91. Бибкод : 1981Sci...213..789N . дои : 10.1126/science.7256281 . ПМК 2693128 . ПМИД 7256281 .
- ^ Горис, РЦ; Терашима, С. (1973). «Центральный ответ на инфракрасную стимуляцию ямочных рецепторов у кроталиновой змеи Trimeresurus flavoviridis». Журнал экспериментальной биологии . 58 (1): 59–76. дои : 10.1242/jeb.58.1.59 . ПМИД 4350276 .
- ^ «Раскрыто инфракрасное обнаружение змей» . Архивировано из оригинала 28 декабря 2016 года . Проверено 20 января 2017 г.
- ^ Jump up to: а б Крохмаль, Аарон Р.; Джордж С. Баккен; Трэвис Дж. ЛаДюк (15 ноября 2004 г.). «Жара на кухне эволюции: эволюционные взгляды на функции и происхождение лицевой ямки гадюк (Viperidae: Crotalinae)» . Журнал экспериментальной биологии . 207 (Часть 24): 4231–4238. дои : 10.1242/jeb.01278 . ПМИД 15531644 .
- ^ Jump up to: а б Грин Х.В. 1992. Экологический и поведенческий контекст эволюции гадюки. Кэмпбелл Дж. А., Броди Э. Д. младший, 1992. Биология питвайперов. Техас: Сельва. 467 стр. 17 пластин. ISBN 0-9630537-0-1 .
- ^ Паф и др. 1992. Герпетология: третье издание. Пирсон Прентис Холл: Pearson Education, Inc., 2002.
- ^ Горис, ЧР; и др. (2003). «Микроциркуляция ямочных органов питона: морфология и кинетика кровотока». Микрососудистые исследования . 65 (3): 179–185. дои : 10.1016/s0026-2862(03)00003-7 . ПМИД 12711259 .
- ^ Ньюман, Э.А.; Груберд, ER; Хартлайн, штат Пенсильвания (1980). «Инфракрасный тригемино-тектальный путь у гремучей змеи и питона». Журнал сравнительной неврологии . 191 (3): 465–477. дои : 10.1002/cne.901910309 . ПМИД 7410602 . S2CID 10279222 .
- ^ Хартлайн, штат Пенсильвания; Л Касс; МС Луп (17 марта 1978 г.). «Слияние модальностей в зрительном тектуме: инфракрасная и визуальная интеграция у гремучих змей». Наука . 199 (4334): 1225–1229. Бибкод : 1978Sci...199.1225H . дои : 10.1126/science.628839 . ПМИД 628839 .
- ^ Jump up to: а б Буллок, TH; Коулз, РБ (1952). «Физиология инфракрасного рецептора: лицевая ямка гадюк». Наука . 115 (2994): 541–543. Бибкод : 1952Sci...115..541B . дои : 10.1126/science.115.2994.541-a . ПМИД 17731960 . S2CID 30122231 .
- ^ Jump up to: а б Баккен, Джордж С.; Крохмаль, Аарон Р. (2007), «Свойства визуализации и чувствительность лицевых ямок гадюк, определенные с помощью оптического анализа и анализа теплопередачи», Journal of Experimental Biology , 210 (16): 2801–2810, doi : 10.1242/ jeb.006965 , PMID 17690227
- ^ Jump up to: а б Грачева Елена О.; Николас Т. Инголия; Ивонн М. Келли; Хулио Ф. Кордеро-Моралес; Гюнтер Холлопетер; Александр Т. Чеслер; Эльда Э. Санчес; Джон К. Перес; Джонатан С. Вайсман; Дэвид Юлиус (15 апреля 2010 г.). «Молекулярные основы обнаружения инфракрасного излучения змеями» . Природа . 464 (7291): 1006–1011. Бибкод : 2010Natur.464.1006G . дои : 10.1038/nature08943 . ПМК 2855400 . ПМИД 20228791 .
- ^ Чен, Кью; Лю, Ю; Браут, SE; Фанг, Дж; Тан, Ю (2017). «Тепловой фон определяет, как взаимодействуют инфракрасная и зрительная системы у гадюк» . Журнал экспериментальной биологии . 220 (Часть 17): 3103–3109. дои : 10.1242/jeb.155382 . ПМИД 28855322 .
- ^ Кардонг, КВ; Макесси, СП (1991). «Ударное поведение врожденно слепой гремучей змеи». Журнал герпетологии . 25 (2): 208–211. дои : 10.2307/1564650 . JSTOR 1564650 .
- ^ Шрафт, штат Ха; Гудман, К; Кларк, RW (2017). «Используют ли гремучие змеи, живущие на свободе, тепловые сигналы для оценки добычи?». Журнал сравнительной физиологии А. 204 (3): 295–303. дои : 10.1007/s00359-017-1239-8 . ПМИД 29218413 . S2CID 3370317 .
- ^ Шрафт, штат Ха; Баккен, Г.С.; Кларк, RW (2019). «Змеи, чувствительные к инфракрасному излучению, выбирают ориентацию засады на основе теплового фона» . Научные отчеты . 9 (1): 3950. Бибкод : 2019НатСР...9.3950С . дои : 10.1038/s41598-019-40466-0 . ПМК 6408448 . PMID 30850649 – через проверку.
- ^ Шайн, Р; Солнце, Л; Кирни, М; Фицджеральд, М. (2002). «Термические корреляты выбора места кормления китайской гадюкой (Gloydius shedaoensis, Viperidae)». Журнал термической биологии . 27 (5): 405–412. дои : 10.1016/S0306-4565(02)00009-8 .
| Концепции | |
|---|---|
| Люди | |
| Методы | |
| Системы | |