Эволюционная таксономия
Эта статья может быть слишком технической для понимания большинства читателей . ( Июль 2018 г. ) |
Эволюционная таксономия , эволюционная систематика или дарвиновская классификация — это раздел биологической классификации , который стремится классифицировать организмы , используя комбинацию филогенетических отношений (общее происхождение), отношений прародитель-потомок (серийное происхождение) и степени эволюционных изменений. Этот тип таксономии может рассматривать целые таксоны, а не отдельные виды , так что можно сделать вывод, что группы видов дают начало новым группам. [1] Эта концепция нашла свою наиболее известную форму в современном эволюционном синтезе начала 1940-х годов.
Эволюционная таксономия отличается от строгой додарвиновской таксономии Линнея (составляющей только упорядоченные списки) тем, что она строит эволюционные деревья. В то время как в филогенетической номенклатуре каждый таксон должен состоять из одного предкового узла и всех его потомков, эволюционная систематика позволяет исключать группы из их родительских таксонов (например, динозавры не считаются птицами , но дали начало им ), таким образом разрешающие парафилетические таксоны. [2] [3]
Происхождение эволюционной таксономии

Эволюционная систематика возникла в результате влияния теории эволюции на систематику Линнея . Идея перевода систематики Линнея в своего рода дендрограмму царств животных и растений . была сформулирована в конце XVIII века, задолго до Чарльза Дарвина книги «Происхождение видов» публикации [4] Первым, кто предположил, что организмы имеют общее происхождение, был Пьер-Луи Моро де Мопертюи в своем «Опыте космологии» 1751 года . [5] [6] Трансмутация видов вошла в более широкие научные круги с Эразма Дарвина 1796 года «Зоономией» и Жана-Батиста Ламарка 1809 года «Зоологической философией» . [7] Идея была популяризирована в англоязычном мире благодаря спекулятивной, но широко читаемой книге «Остатки естественной истории творения» , анонимно опубликованной Робертом Чемберсом в 1844 году. [8]
После появления «Происхождения видов » изображения Древа жизни стали популярны в научных трудах. В «Происхождении видов » предок оставался в основном гипотетическим видом; Дарвин был в первую очередь занят демонстрацией этого принципа, тщательно воздерживаясь от спекуляций о взаимоотношениях между живыми или ископаемыми организмами и используя только теоретические примеры. [7] Напротив, Чемберс предложил конкретные гипотезы, например, об эволюции плацентарных млекопитающих от сумчатых. [9]
После публикации Дарвина Томас Генри Хаксли использовал окаменелости археоптерикса и гесперорниса, чтобы доказать, что птицы являются потомками динозавров. [10] Таким образом, группа современных животных могла быть связана с группой ископаемых. Полученное в результате описание динозавров, «давших начало» или «предков» птиц, демонстрирует важнейший признак эволюционного таксономического мышления.
За последние три десятилетия произошло резкое увеличение использования последовательностей ДНК для реконструкции филогении и параллельный сдвиг акцента с эволюционной таксономии на «филогенетическую систематику» Хеннига. [7]
Сегодня, с появлением современной геномики , ученые во всех областях биологии используют молекулярную филогению в качестве руководства для своих исследований. Одним из распространенных методов является множественное выравнивание последовательностей . [ нужна ссылка ]
Томас Кавальер-Смит , [7] Г.Г. Симпсон и Эрнст Майр [11] являются некоторыми представительными эволюционными систематиками.
Новые методы в эволюционной современной систематике
Этот раздел может чрезмерно полагаться на источники, слишком тесно связанные с предметом , что может помешать статье стать проверяемой и нейтральной . ( Июль 2018 г. ) |
В последнее время появились попытки объединить современные методы кладистики, филогенетики и анализа ДНК с классическими взглядами на систематику. Некоторые авторы обнаружили, что филогенетический анализ приемлем с научной точки зрения до тех пор, пока допустима парафилия, по крайней мере, для определенных групп. Такую позицию продвигает в статьях Тод Ф. Стюсси. [12] и другие. Особенно строгая форма эволюционной систематики была представлена Ричардом Х. Зандером в ряде статей, но обобщена в его «Рамках постфилогенетической систематики». [13]
Коротко говоря, плюралистическая систематика Зандера основана на неполноте каждой из теорий: метод, который не может фальсифицировать гипотезу, так же ненаучен, как и гипотеза, которую невозможно фальсифицировать. Кладистика генерирует только деревья общего, а не серийного происхождения. С таксонами, развивающимися последовательно, невозможно справиться путем анализа общего происхождения кладистическими методами. Такие гипотезы, как адаптивная радиация одного предкового таксона, не могут быть опровергнуты кладистикой. Кладистика предлагает способ группировки по трансформациям признаков, но ни одно эволюционное дерево не может быть полностью дихотомическим. Филогенетика полагает, что общие предковые таксоны являются причинами дихотомии, но доказательств существования таких таксонов нет. Молекулярная систематика использует данные о последовательностях ДНК для отслеживания эволюционных изменений, таким образом, парафилия, а иногда и филогенетическая полифилия сигнализируют о трансформациях предков-потомков на уровне таксона, но в остальном молекулярная филогенетика не учитывает вымершую парафилию. Чтобы сделать вывод о серийном происхождении, необходим дополнительный трансформационный анализ.
Бессейский кактус или коммаграмма — лучшее эволюционное древо, показывающее как общее, так и серийное происхождение. Сначала генерируется кладограмма или естественный ключ. Выявляются обобщенные предковые таксоны, а специализированные таксоны-потомки отмечаются как вышедшие из линии с линией одного цвета, обозначающей прародителя во времени. Затем разрабатывается бессеянский кактус или коммаграмма, отражающая как общее, так и серийное происхождение. Таксоны-прародители могут иметь один или несколько таксонов-потомков. Меры поддержки могут быть предоставлены с точки зрения факторов Байеса, следуя методу трансформационного анализа Зандера с использованием децибанов.
Кладистический анализ группирует таксоны по общим признакам, но включает модель дихотомического ветвления, заимствованную из фенетики. По сути, это упрощенный дихотомический естественный ключ, хотя допускаются обращения. Проблема, конечно, в том, что эволюция не обязательно дихотомична. Предковому таксону, дающему двух или более потомков, требуется более длинное и менее экономное дерево. Узел кладограммы суммирует все признаки, дистальные от него, а не какого-либо одного таксона, и непрерывность в кладограмме происходит от узла к узлу, а не от таксона к таксону. Это не модель эволюции, а вариант иерархического кластерного анализа (изменения признаков и неультраметрические ветви). Вот почему дерево, основанное исключительно на общих признаках, называется не эволюционным деревом, а просто кладистическим деревом. Это дерево отражает в значительной степени эволюционные отношения посредством трансформации признаков, но игнорируют отношения, возникающие в результате трансформации существующих таксонов на уровне вида.
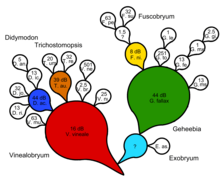
Филогенетика пытается внедрить серийный элемент, постулируя специальные, не поддающиеся доказыванию общие предки в каждом узле кладистического дерева. Для полностью дихотомической кладограммы на одного невидимого общего предка меньше, чем терминальных таксонов. Таким образом, мы фактически получаем дихотомический естественный ключ с невидимым общим предком, генерирующим каждый куплет. Это не может подразумевать объяснение, основанное на процессах, без обоснования дихотомии и предположения об общих предках как причинах. По мнению Зандера, кладистическая форма анализа эволюционных взаимоотношений не может фальсифицировать какой-либо подлинный эволюционный сценарий, включающий серийную трансформацию. [14]
У Зандера есть подробные методы создания мер поддержки молекулярного серийного происхождения. [15] и для морфологического серийного спуска с использованием факторов Байеса и последовательного анализа Байеса посредством добавления децибана Тьюринга или добавления информационных битов Шеннона. [16] [17] [18] [19]
Древо Жизни [ править ]

Поскольку в конце 19 и начале 20 века было обнаружено и признано все больше и больше групп ископаемых, палеонтологи работали над пониманием истории животных на протяжении веков, связывая вместе известные группы. [20] Древо жизни медленно составлялось, и группы ископаемых занимали свои места на дереве по мере того, как понимание увеличивалось. [ нужна ссылка ]
Эти группы все еще сохраняли свои формальные линнеевские таксономические ранги . Некоторые из них парафилетичны в том смысле, что, хотя каждый организм в группе связан с общим предком непрерывной цепью промежуточных предков внутри группы, некоторые другие потомки этого предка лежат вне группы. Эволюцию и распространение различных таксонов во времени обычно изображают в виде диаграммы веретена (часто называемой ромерограммой в честь американского палеонтолога Альфреда Ромера ), где различные веретена ответвляются друг от друга, причем каждое веретено представляет собой таксон. Ширина веретен означает зависимость численности (часто количества семей) от времени. [21]
Палеонтология позвоночных наметила эволюционную последовательность позвоночных, которая в настоящее время довольно хорошо понимается, к концу 19-го века, после чего последовало разумное понимание эволюционной последовательности царства растений к началу 20-го века . Объединение различных деревьев в великое Древо Жизни по-настоящему стало возможным только благодаря достижениям микробиологии и биохимии в период между мировыми войнами. [ нужна ссылка ]
Терминологическая разница [ править ]
Два подхода, эволюционная таксономия и филогенетическая систематика, заимствованная у Вилли Хеннига , различаются использованием слова « монофилетический ». Для эволюционных систематиков «монофилетика» означает лишь то, что группа произошла от одного общего предка. В филогенетической номенклатуре есть дополнительная оговорка: в группу следует включать предковые виды и всех потомков. [2] термин « голофилетический Для последнего значения был предложен ». Например, согласно эволюционной систематике амфибии являются монофилетическими, поскольку они произошли от рыб только один раз. Согласно филогенетической таксономии, амфибии не составляют монофилетическую группу, поскольку амниоты ( рептилии , птицы и млекопитающие ) произошли от предка земноводных и все же не считаются амфибиями. Такие парафилетические группы отвергаются в филогенетической номенклатуре, но эволюционные систематики считают их признаком серийного происхождения. [22]
Ссылки [ править ]
- ^ Майр, Эрнст и Бок, WJ (2002), «Классификации и другие системы упорядочения» (PDF) , Дж. Зоол. Сист. Эвол. Исследования , 40 (4): 169–94, doi : 10.1046/j.1439-0469.2002.00211.x
- ^ Jump up to: Перейти обратно: а б Грант, В. (2003), «Несоответствие между кладистическими и таксономическими системами», American Journal of Botany , 90 (9): 1263–70, doi : 10.3732/ajb.90.9.1263 , PMID 21659226
- ^ Обер, Д. (2015). «Формальный анализ филогенетической терминологии: на пути к пересмотру современной парадигмы в систематике». Фитонейрон . 2015–66: 1–54.
- ^ Арчибальд, Дж. Дэвид (2009). «Додарвиновское (1840 г.) «Древо жизни» Эдварда Хичкока » (PDF) . Журнал истории биологии . 42 (3): 561–592. CiteSeerX 10.1.1.688.7842 . дои : 10.1007/s10739-008-9163-y . ПМИД 20027787 . S2CID 16634677 .
- ^ Дж. С. Бромли, Новая современная история Кембриджа: подъем Великобритании и России, 1688-1715/25 , Архив CUP, 1970, ISBN 978-0-521-07524-4 , стр. 62-63.
- ^ Сокровище Джеффри Рассела Ричардса, Создание современной Европы, 1648-1780 , Тейлор и Фрэнсис, 1985, ISBN 978-0-416-72370-0 , стр. 142
- ^ Jump up to: Перейти обратно: а б с д Кавальер-Смит, Томас (2010). «Глубокая филогения, предковые группы и четыре возраста жизни» (PDF) . Философские труды Королевского общества Б. 365 (1537): 111–132. дои : 10.1098/rstb.2009.0161 . ПМЦ 2842702 . ПМИД 20008390 .
- ^ Секорд, Джеймс А. (2000), Викторианская сенсация: выдающаяся публикация, прием и тайное авторство остатков естественной истории творения , Чикаго: University of Chicago Press, ISBN 978-0-226-74410-0 , архивировано из оригинала 16 мая 2008 г. , получено 17 января 2011 г.
- ^ Чемберс, Роберт (1844). Остатки естественной истории творения . Лондон: Джон Черчилль. п. 122 . Проверено 10 мая 2013 г.
- ^ Хаксли, TH (1876): Лекции по эволюции. Нью-Йорк Трибьюн . Дополнительный. № 36. В сборнике сочинений IV: стр. 46–138, оригинальный текст с рисунками.
- ^ Манер, Мартин; Бунге, Марио. Основы биофилософии . Спрингер, с. 251.
- ^ Стюсси, Тод Ф. (2010). «Парафилия, происхождение и классификация покрытосеменных». Таксон . 59 (3): 689–693. дои : 10.1002/tax.593001 .
- ^ Зандер, Ричард (2013). Основы постфилогенетической систематики . Сент-Луис: Zetetic Publications, Amazon CreateSpace.
- ^ Зандер, Ричард (2014). «Ответ на особенно неприятную рецензию в журнале Cladistics». Фитонейрон . 2014 (110): 1–4.
- ^ Зандер, Ричард (2014). «Меры поддержки каулистических макроэволюционных преобразований в эволюционных деревьях». Анналы ботанического сада Миссури . 100 (1–2): 100–107. дои : 10.3417/2012090 . S2CID 86754093 .
- ^ Зандер, Ричард (2014). «Классическое определение монофилии на примере Didymodon s. lat. (Bryophyta). Часть 1 из 3, краткий обзор и упрощенные концепции» (PDF) . Фитонейрон . 2014 (78): 1–7 . Проверено 4 января 2015 г.
- ^ Зандер, Ричард (2014). «Классическое определение монофилии на примере Didymodon s. lat. (Bryophyta). Часть 2 из 3, концепции» (PDF) . Фитонейрон . 2014 (79): 1–23 . Проверено 4 января 2015 г.
- ^ Зандер, Ричард (2014). «Классическое определение монофилии на примере Didymodon s. lat. (Bryophyta). Часть 3 из 3, анализ» (PDF) . Фитонейрон . 2014 (80): 1–19 . Проверено 4 января 2015 г.
- ^ Зандер Р.Х. 2018. Макроэволюционная систематика Streptotrichaceae мохообразных и ее применение к термодинамической стабильности экосистемы. Издание 2. Zetetic Publications, независимое издательство CreateSpace, Amazon, Сент-Луис.
- ^ Рудвик, MJS (1985), Значение окаменелостей: эпизоды в истории палеонтологии , University of Chicago Press , стр. 24, ISBN 978-0-226-73103-2
- ^ Уайт, Т.; Казлев М.А. "Филогения и систематика : Глоссарий - Ромерограмма" . palaeos.com . Проверено 21 января 2013 г.
- ^ Зандер, Р.Х. (2008). «Эволюционные выводы из немонофилии молекулярных деревьев». Таксон . 57 (4): 1182–1188. дои : 10.1002/tax.574011 .
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||