млекопитающее
| Млекопитающие Временной диапазон: | |
|---|---|
 | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Amniota |
| Clade: | Synapsida |
| Clade: | Mammaliaformes |
| Class: | Mammalia Linnaeus, 1758 |
| Living subgroups | |
Млекопитающее mamma (от латинского « грудь») [1] — позвоночное животное класса млекопитающих ( / m ə ˈ m eɪ l i . ə / ) . Млекопитающие характеризуются наличием молоко вырабатывающих молочных желез, для кормления детенышей, неокортексной области мозга, шерсти или волос и трех косточек среднего уха . Эти характеристики отличают их от рептилий и птиц , от которых их предки отделились в каменноугольном периоде более 300 миллионов лет назад. около 6400 современных видов Описано млекопитающих, разделенных на 29 отрядов .
Крупнейшими отрядами млекопитающих по числу видов являются грызуны , летучие мыши и эвлипотифлы (в том числе ежи , кроты и землеройки ). Следующие три — приматы (включая людей , обезьян и лемуров ), парнокопытные (включая свиней , верблюдов и китов ) и хищные животные (включая кошек , собак и тюленей ).
Млекопитающие — единственные живые члены Synapsida ; эта клада вместе с Sauropsida (рептилиями и птицами) составляет более крупную кладу Amniota . Ранние синапсиды называются « пеликозаврами ». Более продвинутые терапсиды стали доминировать в средней перми . Млекопитающие произошли от цинодонтов , развитой группы терапсидов, в период от позднего триаса до ранней юры . Млекопитающие достигли своего современного разнообразия в палеогеновом и неогеновом периодах кайнозойской эры, после вымирания нептичьих динозавров , и были доминирующей группой наземных животных с 66 миллионов лет назад до настоящего времени.
Основной тип тела млекопитающих — четвероногий , и большинство млекопитающих используют свои четыре конечности для передвижения по земле ; но у некоторых конечности приспособлены к жизни на море , в воздухе , на деревьях , под землей или на двух ногах . Размер млекопитающих варьируется от шмелиной летучей мыши длиной 30 м (98 футов) размером 30–40 мм (1,2–1,6 дюйма) до синего кита — возможно, самого крупного животного, когда-либо жившего на свете. Максимальная продолжительность жизни варьируется от двух лет у землеройки до 211 лет у гренландского кита . Все современные млекопитающие рождают живых детенышей, за исключением пяти видов однопроходных , которые являются яйцекладущими млекопитающими. Самая богатая видами группа млекопитающих, инфракласс , называемый плацентами , имеет плаценту , которая обеспечивает питание плода во время беременности .
Most mammals are intelligent, with some possessing large brains, self-awareness, and tool use. Mammals can communicate and vocalize in several ways, including the production of ultrasound, scent-marking, alarm signals, singing, echolocation; and, in the case of humans, complex language. Mammals can organize themselves into fission–fusion societies, harems, and hierarchies—but can also be solitary and territorial. Most mammals are polygynous, but some can be monogamous or polyandrous.
Domestication of many types of mammals by humans played a major role in the Neolithic Revolution, and resulted in farming replacing hunting and gathering as the primary source of food for humans. This led to a major restructuring of human societies from nomadic to sedentary, with more co-operation among larger and larger groups, and ultimately the development of the first civilizations. Domesticated mammals provided, and continue to provide, power for transport and agriculture, as well as food (meat and dairy products), fur, and leather. Mammals are also hunted and raced for sport, kept as pets and working animals of various types, and are used as model organisms in science. Mammals have been depicted in art since Paleolithic times, and appear in literature, film, mythology, and religion. Decline in numbers and extinction of many mammals is primarily driven by human poaching and habitat destruction, primarily deforestation.
Classification

Mammal classification has been through several revisions since Carl Linnaeus initially defined the class, and at present, no classification system is universally accepted. McKenna & Bell (1997) and Wilson & Reeder (2005) provide useful recent compendiums.[2] Simpson (1945)[3] provides systematics of mammal origins and relationships that had been taught universally until the end of the 20th century.However, since 1945, a large amount of new and more detailed information has gradually been found: The paleontological record has been recalibrated, and the intervening years have seen much debate and progress concerning the theoretical underpinnings of systematization itself, partly through the new concept of cladistics. Though fieldwork and lab work progressively outdated Simpson's classification, it remains the closest thing to an official classification of mammals, despite its known issues.[4]
Most mammals, including the six most species-rich orders, belong to the placental group. The three largest orders in numbers of species are Rodentia: mice, rats, porcupines, beavers, capybaras, and other gnawing mammals; Chiroptera: bats; and Soricomorpha: shrews, moles, and solenodons. The next three biggest orders, depending on the biological classification scheme used, are the primates: apes, monkeys, and lemurs; the Cetartiodactyla: whales and even-toed ungulates; and the Carnivora which includes cats, dogs, weasels, bears, seals, and allies.[5] According to Mammal Species of the World, 5,416 species were identified in 2006. These were grouped into 1,229 genera, 153 families and 29 orders.[5] In 2008, the International Union for Conservation of Nature (IUCN) completed a five-year Global Mammal Assessment for its IUCN Red List, which counted 5,488 species.[6] According to research published in the Journal of Mammalogy in 2018, the number of recognized mammal species is 6,495, including 96 recently extinct.[7]
Definitions
The word "mammal" is modern, from the scientific name Mammalia coined by Carl Linnaeus in 1758, derived from the Latin mamma ("teat, pap"). In an influential 1988 paper, Timothy Rowe defined Mammalia phylogenetically as the crown group of mammals, the clade consisting of the most recent common ancestor of living monotremes (echidnas and platypuses) and Therian mammals (marsupials and placentals) and all descendants of that ancestor.[8] Since this ancestor lived in the Jurassic period, Rowe's definition excludes all animals from the earlier Triassic, despite the fact that Triassic fossils in the Haramiyida have been referred to the Mammalia since the mid-19th century.[9] If Mammalia is considered as the crown group, its origin can be roughly dated as the first known appearance of animals more closely related to some extant mammals than to others. Ambondro is more closely related to monotremes than to therian mammals while Amphilestes and Amphitherium are more closely related to the therians; as fossils of all three genera are dated about 167 million years ago in the Middle Jurassic, this is a reasonable estimate for the appearance of the crown group.[10]
T. S. Kemp has provided a more traditional definition: "Synapsids that possess a dentary–squamosal jaw articulation and occlusion between upper and lower molars with a transverse component to the movement" or, equivalently in Kemp's view, the clade originating with the last common ancestor of Sinoconodon and living mammals.[11] The earliest-known synapsid satisfying Kemp's definitions is Tikitherium, dated 225 Ma, so the appearance of mammals in this broader sense can be given this Late Triassic date.[12][13]However, this animal may have actually evolved during the Neogene.[14]
McKenna/Bell classification
In 1997, the mammals were comprehensively revised by Malcolm C. McKenna and Susan K. Bell, which has resulted in the McKenna/Bell classification. The authors worked together as paleontologists at the American Museum of Natural History. McKenna inherited the project from Simpson and, with Bell, constructed a completely updated hierarchical system, covering living and extinct taxa, that reflects the historical genealogy of Mammalia.[4] Their 1997 book, Classification of Mammals above the Species Level,[15] is a comprehensive work on the systematics, relationships and occurrences of all mammal taxa, living and extinct, down through the rank of genus, though molecular genetic data challenge several of the groupings.
In the following list, extinct groups are labelled with a dagger (†).
Class Mammalia
- Subclass Prototheria: monotremes: echidnas and the platypus
- Subclass Theriiformes: live-bearing mammals and their prehistoric relatives
- Infraclass †Allotheria: multituberculates
- Infraclass †Eutriconodonta: eutriconodonts
- Infraclass Holotheria: modern live-bearing mammals and their prehistoric relatives
- Superlegion †Kuehneotheria
- Supercohort Theria: live-bearing mammals
- Cohort Marsupialia: marsupials
- Magnorder Australidelphia: Australian marsupials and the monito del monte
- Magnorder Ameridelphia: New World marsupials. Now considered paraphyletic, with shrew opossums being closer to australidelphians.[16]
- Cohort Placentalia: placentals
- Magnorder Xenarthra: xenarthrans
- Magnorder Epitheria: epitheres
- Superorder †Leptictida
- Superorder Preptotheria
- Grandorder Anagalida: lagomorphs, rodents and elephant shrews
- Grandorder Ferae: carnivorans, pangolins, †creodonts and relatives
- Grandorder Lipotyphla: insectivorans
- Grandorder Archonta: bats, primates, colugos and treeshrews (now considered paraphyletic, with bats being closer to other groups)
- Grandorder Euungulata: ungulates
- Order Tubulidentata incertae sedis: aardvark
- Mirorder Eparctocyona: †condylarths, whales and artiodactyls (even-toed ungulates)
- Mirorder †Meridiungulata: South American ungulates
- Mirorder Altungulata: perissodactyls (odd-toed ungulates), elephants, manatees and hyraxes
- Cohort Marsupialia: marsupials
Molecular classification of placentals

As of the early 21st century, molecular studies based on DNA analysis have suggested new relationships among mammal families. Most of these findings have been independently validated by retrotransposon presence/absence data.[18] Classification systems based on molecular studies reveal three major groups or lineages of placental mammals—Afrotheria, Xenarthra and Boreoeutheria—which diverged in the Cretaceous. The relationships between these three lineages is contentious, and all three possible hypotheses have been proposed with respect to which group is basal. These hypotheses are Atlantogenata (basal Boreoeutheria), Epitheria (basal Xenarthra) and Exafroplacentalia (basal Afrotheria).[19] Boreoeutheria in turn contains two major lineages—Euarchontoglires and Laurasiatheria.
Estimates for the divergence times between these three placental groups range from 105 to 120 million years ago, depending on the type of DNA used (such as nuclear or mitochondrial)[20] and varying interpretations of paleogeographic data.[19]
| Tarver et al. 2016[21] | Sandra Álvarez-Carretero et al. 2022[22][23] |
|---|---|

Evolution
Origins
Synapsida, a clade that contains mammals and their extinct relatives, originated during the Pennsylvanian subperiod (~323 million to ~300 million years ago), when they split from the reptile lineage. Crown group mammals evolved from earlier mammaliaforms during the Early Jurassic. The cladogram takes Mammalia to be the crown group.[24]
| Mammaliaformes |
| ||||||||||||||||||||||||
Evolution from older amniotes

The first fully terrestrial vertebrates were amniotes. Like their amphibious early tetrapod predecessors, they had lungs and limbs. Amniotic eggs, however, have internal membranes that allow the developing embryo to breathe but keep water in. Hence, amniotes can lay eggs on dry land, while amphibians generally need to lay their eggs in water.
The first amniotes apparently arose in the Pennsylvanian subperiod of the Carboniferous. They descended from earlier reptiliomorph amphibious tetrapods,[25] which lived on land that was already inhabited by insects and other invertebrates as well as ferns, mosses and other plants. Within a few million years, two important amniote lineages became distinct: the synapsids, which would later include the common ancestor of the mammals; and the sauropsids, which now include turtles, lizards, snakes, crocodilians and dinosaurs (including birds).[26] Synapsids have a single hole (temporal fenestra) low on each side of the skull. Primitive synapsids included the largest and fiercest animals of the early Permian such as Dimetrodon.[27] Nonmammalian synapsids were traditionally—and incorrectly—called "mammal-like reptiles" or pelycosaurs; we now know they were neither reptiles nor part of reptile lineage.[28][29]
Therapsids, a group of synapsids, evolved in the Middle Permian, about 265 million years ago, and became the dominant land vertebrates.[28] They differ from basal eupelycosaurs in several features of the skull and jaws, including: larger skulls and incisors which are equal in size in therapsids, but not for eupelycosaurs.[28] The therapsid lineage leading to mammals went through a series of stages, beginning with animals that were very similar to their early synapsid ancestors and ending with probainognathian cynodonts, some of which could easily be mistaken for mammals. Those stages were characterized by:[30]
- The gradual development of a bony secondary palate.
- Abrupt acquisition of endothermy among Mammaliamorpha, thus prior to the origin of mammals by 30–50 millions of years [31].
- Progression towards an erect limb posture, which would increase the animals' stamina by avoiding Carrier's constraint. But this process was slow and erratic: for example, all herbivorous nonmammaliaform therapsids retained sprawling limbs (some late forms may have had semierect hind limbs); Permian carnivorous therapsids had sprawling forelimbs, and some late Permian ones also had semisprawling hindlimbs. In fact, modern monotremes still have semisprawling limbs.
- The dentary gradually became the main bone of the lower jaw which, by the Triassic, progressed towards the fully mammalian jaw (the lower consisting only of the dentary) and middle ear (which is constructed by the bones that were previously used to construct the jaws of reptiles).
First mammals
The Permian–Triassic extinction event about 252 million years ago, which was a prolonged event due to the accumulation of several extinction pulses, ended the dominance of carnivorous therapsids.[32] In the early Triassic, most medium to large land carnivore niches were taken over by archosaurs[33] which, over an extended period (35 million years), came to include the crocodylomorphs,[34] the pterosaurs and the dinosaurs;[35] however, large cynodonts like Trucidocynodon and traversodontids still occupied large sized carnivorous and herbivorous niches respectively. By the Jurassic, the dinosaurs had come to dominate the large terrestrial herbivore niches as well.[36]
The first mammals (in Kemp's sense) appeared in the Late Triassic epoch (about 225 million years ago), 40 million years after the first therapsids. They expanded out of their nocturnal insectivore niche from the mid-Jurassic onwards;[37] the Jurassic Castorocauda, for example, was a close relative of true mammals that had adaptations for swimming, digging and catching fish.[38] Most, if not all, are thought to have remained nocturnal (the nocturnal bottleneck), accounting for much of the typical mammalian traits.[39] The majority of the mammal species that existed in the Mesozoic Era were multituberculates, eutriconodonts and spalacotheriids.[40] The earliest-known metatherian is Sinodelphys, found in 125-million-year-old Early Cretaceous shale in China's northeastern Liaoning Province. The fossil is nearly complete and includes tufts of fur and imprints of soft tissues.[41]

The oldest-known fossil among the Eutheria ("true beasts") is the small shrewlike Juramaia sinensis, or "Jurassic mother from China", dated to 160 million years ago in the late Jurassic.[42] A later eutherian relative, Eomaia, dated to 125 million years ago in the early Cretaceous, possessed some features in common with the marsupials but not with the placentals, evidence that these features were present in the last common ancestor of the two groups but were later lost in the placental lineage.[43] In particular, the epipubic bones extend forwards from the pelvis. These are not found in any modern placental, but they are found in marsupials, monotremes, other nontherian mammals and Ukhaatherium, an early Cretaceous animal in the eutherian order Asioryctitheria. This also applies to the multituberculates.[44] They are apparently an ancestral feature, which subsequently disappeared in the placental lineage. These epipubic bones seem to function by stiffening the muscles during locomotion, reducing the amount of space being presented, which placentals require to contain their fetus during gestation periods. A narrow pelvic outlet indicates that the young were very small at birth and therefore pregnancy was short, as in modern marsupials. This suggests that the placenta was a later development.[45]
One of the earliest-known monotremes was Teinolophos, which lived about 120 million years ago in Australia.[46] Monotremes have some features which may be inherited from the original amniotes such as the same orifice to urinate, defecate and reproduce (cloaca)—as lizards and birds also do—[47] and they lay eggs which are leathery and uncalcified.[48]
Earliest appearances of features
Hadrocodium, whose fossils date from approximately 195 million years ago, in the early Jurassic, provides the first clear evidence of a jaw joint formed solely by the squamosal and dentary bones; there is no space in the jaw for the articular, a bone involved in the jaws of all early synapsids.[49]

The earliest clear evidence of hair or fur is in fossils of Castorocauda and Megaconus, from 164 million years ago in the mid-Jurassic. In the 1950s, it was suggested that the foramina (passages) in the maxillae and premaxillae (bones in the front of the upper jaw) of cynodonts were channels which supplied blood vessels and nerves to vibrissae (whiskers) and so were evidence of hair or fur;[50][51] it was soon pointed out, however, that foramina do not necessarily show that an animal had vibrissae, as the modern lizard Tupinambis has foramina that are almost identical to those found in the nonmammalian cynodont Thrinaxodon.[29][52] Popular sources, nevertheless, continue to attribute whiskers to Thrinaxodon.[53] Studies on Permian coprolites suggest that non-mammalian synapsids of the epoch already had fur, setting the evolution of hairs possibly as far back as dicynodonts.[54]
When endothermy first appeared in the evolution of mammals is uncertain, though it is generally agreed to have first evolved in non-mammalian therapsids.[54][55] Modern monotremes have lower body temperatures and more variable metabolic rates than marsupials and placentals,[56] but there is evidence that some of their ancestors, perhaps including ancestors of the therians, may have had body temperatures like those of modern therians.[57] Likewise, some modern therians like afrotheres and xenarthrans have secondarily developed lower body temperatures.[58]
The evolution of erect limbs in mammals is incomplete—living and fossil monotremes have sprawling limbs. The parasagittal (nonsprawling) limb posture appeared sometime in the late Jurassic or early Cretaceous; it is found in the eutherian Eomaia and the metatherian Sinodelphys, both dated to 125 million years ago.[59] Epipubic bones, a feature that strongly influenced the reproduction of most mammal clades, are first found in Tritylodontidae, suggesting that it is a synapomorphy between them and mammaliaformes. They are omnipresent in non-placental mammaliaformes, though Megazostrodon and Erythrotherium appear to have lacked them.[60]
It has been suggested that the original function of lactation (milk production) was to keep eggs moist. Much of the argument is based on monotremes, the egg-laying mammals.[61][62] In human females, mammary glands become fully developed during puberty, regardless of pregnancy.[63]
Rise of the mammals

Therian mammals took over the medium- to large-sized ecological niches in the Cenozoic, after the Cretaceous–Paleogene extinction event approximately 66 million years ago emptied ecological space once filled by non-avian dinosaurs and other groups of reptiles, as well as various other mammal groups,[65] and underwent an exponential increase in body size (megafauna).[66] Then mammals diversified very quickly; both birds and mammals show an exponential rise in diversity.[65] For example, the earliest-known bat dates from about 50 million years ago, only 16 million years after the extinction of the non-avian dinosaurs.[67]
Molecular phylogenetic studies initially suggested that most placental orders diverged about 100 to 85 million years ago and that modern families appeared in the period from the late Eocene through the Miocene.[68] However, no placental fossils have been found from before the end of the Cretaceous.[69] The earliest undisputed fossils of placentals come from the early Paleocene, after the extinction of the non-avian dinosaurs.[69] (Scientists identified an early Paleocene animal named Protungulatum donnae as one of the first placental mammals,[70] but it has since been reclassified as a non-placental eutherian.)[71] Recalibrations of genetic and morphological diversity rates have suggested a Late Cretaceous origin for placentals, and a Paleocene origin for most modern clades.[72]
The earliest-known ancestor of primates is Archicebus achilles[73] from around 55 million years ago.[73] This tiny primate weighed 20–30 grams (0.7–1.1 ounce) and could fit within a human palm.[73]
Anatomy
Distinguishing features
Living mammal species can be identified by the presence of sweat glands, including those that are specialized to produce milk to nourish their young.[74] In classifying fossils, however, other features must be used, since soft tissue glands and many other features are not visible in fossils.[75]
Many traits shared by all living mammals appeared among the earliest members of the group:
- Jaw joint – The dentary (the lower jaw bone, which carries the teeth) and the squamosal (a small cranial bone) meet to form the joint. In most gnathostomes, including early therapsids, the joint consists of the articular (a small bone at the back of the lower jaw) and quadrate (a small bone at the back of the upper jaw).[49]
- Middle ear – In crown-group mammals, sound is carried from the eardrum by a chain of three bones, the malleus, the incus and the stapes. Ancestrally, the malleus and the incus are derived from the articular and the quadrate bones that constituted the jaw joint of early therapsids.[76]
- Tooth replacement – Teeth can be replaced once (diphyodonty) or (as in toothed whales and murid rodents) not at all (monophyodonty).[77] Elephants, manatees, and kangaroos continually grow new teeth throughout their life (polyphyodonty).[78]
- Prismatic enamel – The enamel coating on the surface of a tooth consists of prisms, solid, rod-like structures extending from the dentin to the tooth's surface.[79]
- Occipital condyles – Two knobs at the base of the skull fit into the topmost neck vertebra; most other tetrapods, in contrast, have only one such knob.[80]
For the most part, these characteristics were not present in the Triassic ancestors of the mammals.[81] Nearly all mammaliaforms possess an epipubic bone, the exception being modern placentals.[82]
Sexual dimorphism

On average, male mammals are larger than females, with males being at least 10% larger than females in over 45% of investigated species. Most mammalian orders also exhibit male-biased sexual dimorphism, although some orders do not show any bias or are significantly female-biased (Lagomorpha). Sexual size dimorphism increases with body size across mammals (Rensch's rule), suggesting that there are parallel selection pressures on both male and female size. Male-biased dimorphism relates to sexual selection on males through male–male competition for females, as there is a positive correlation between the degree of sexual selection, as indicated by mating systems, and the degree of male-biased size dimorphism. The degree of sexual selection is also positively correlated with male and female size across mammals. Further, parallel selection pressure on female mass is identified in that age at weaning is significantly higher in more polygynous species, even when correcting for body mass. Also, the reproductive rate is lower for larger females, indicating that fecundity selection selects for smaller females in mammals. Although these patterns hold across mammals as a whole, there is considerable variation across orders.[83]
Biological systems
The majority of mammals have seven cervical vertebrae (bones in the neck). The exceptions are the manatee and the two-toed sloth, which have six, and the three-toed sloth which has nine.[84] All mammalian brains possess a neocortex, a brain region unique to mammals.[85] Placental brains have a corpus callosum, unlike monotremes and marsupials.[86]




Circulatory systems
The mammalian heart has four chambers, two upper atria, the receiving chambers, and two lower ventricles, the discharging chambers.[87] The heart has four valves, which separate its chambers and ensures blood flows in the correct direction through the heart (preventing backflow). After gas exchange in the pulmonary capillaries (blood vessels in the lungs), oxygen-rich blood returns to the left atrium via one of the four pulmonary veins. Blood flows nearly continuously back into the atrium, which acts as the receiving chamber, and from here through an opening into the left ventricle. Most blood flows passively into the heart while both the atria and ventricles are relaxed, but toward the end of the ventricular relaxation period, the left atrium will contract, pumping blood into the ventricle. The heart also requires nutrients and oxygen found in blood like other muscles, and is supplied via coronary arteries.[88]
Respiratory systems
The lungs of mammals are spongy and honeycombed. Breathing is mainly achieved with the diaphragm, which divides the thorax from the abdominal cavity, forming a dome convex to the thorax. Contraction of the diaphragm flattens the dome, increasing the volume of the lung cavity. Air enters through the oral and nasal cavities, and travels through the larynx, trachea and bronchi, and expands the alveoli. Relaxing the diaphragm has the opposite effect, decreasing the volume of the lung cavity, causing air to be pushed out of the lungs. During exercise, the abdominal wall contracts, increasing pressure on the diaphragm, which forces air out quicker and more forcefully. The rib cage is able to expand and contract the chest cavity through the action of other respiratory muscles. Consequently, air is sucked into or expelled out of the lungs, always moving down its pressure gradient.[89][90] This type of lung is known as a bellows lung due to its resemblance to blacksmith bellows.[90]
Integumentary systems
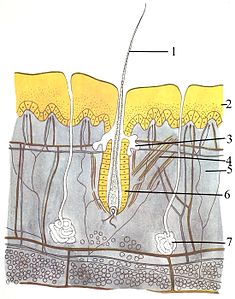
The integumentary system (skin) is made up of three layers: the outermost epidermis, the dermis and the hypodermis. The epidermis is typically 10 to 30 cells thick; its main function is to provide a waterproof layer. Its outermost cells are constantly lost; its bottommost cells are constantly dividing and pushing upward. The middle layer, the dermis, is 15 to 40 times thicker than the epidermis. The dermis is made up of many components, such as bony structures and blood vessels. The hypodermis is made up of adipose tissue, which stores lipids and provides cushioning and insulation. The thickness of this layer varies widely from species to species;[91]: 97 marine mammals require a thick hypodermis (blubber) for insulation, and right whales have the thickest blubber at 20 inches (51 cm).[92] Although other animals have features such as whiskers, feathers, setae, or cilia that superficially resemble it, no animals other than mammals have hair. It is a definitive characteristic of the class, though some mammals have very little.[91]: 61
Digestive systems


Herbivores have developed a diverse range of physical structures to facilitate the consumption of plant material. To break up intact plant tissues, mammals have developed teeth structures that reflect their feeding preferences. For instance, frugivores (animals that feed primarily on fruit) and herbivores that feed on soft foliage have low-crowned teeth specialized for grinding foliage and seeds. Grazing animals that tend to eat hard, silica-rich grasses, have high-crowned teeth, which are capable of grinding tough plant tissues and do not wear down as quickly as low-crowned teeth.[93] Most carnivorous mammals have carnassialiforme teeth (of varying length depending on diet), long canines and similar tooth replacement patterns.[94]
The stomach of even-toed ungulates (Artiodactyla) is divided into four sections: the rumen, the reticulum, the omasum and the abomasum (only ruminants have a rumen). After the plant material is consumed, it is mixed with saliva in the rumen and reticulum and separates into solid and liquid material. The solids lump together to form a bolus (or cud), and is regurgitated. When the bolus enters the mouth, the fluid is squeezed out with the tongue and swallowed again. Ingested food passes to the rumen and reticulum where cellulolytic microbes (bacteria, protozoa and fungi) produce cellulase, which is needed to break down the cellulose in plants.[95] Perissodactyls, in contrast to the ruminants, store digested food that has left the stomach in an enlarged cecum, where it is fermented by bacteria.[96] Carnivora have a simple stomach adapted to digest primarily meat, as compared to the elaborate digestive systems of herbivorous animals, which are necessary to break down tough, complex plant fibers. The caecum is either absent or short and simple, and the large intestine is not sacculated or much wider than the small intestine.[97]
Excretory and genitourinary systems


The mammalian excretory system involves many components. Like most other land animals, mammals are ureotelic, and convert ammonia into urea, which is done by the liver as part of the urea cycle.[98] Bilirubin, a waste product derived from blood cells, is passed through bile and urine with the help of enzymes excreted by the liver.[99] The passing of bilirubin via bile through the intestinal tract gives mammalian feces a distinctive brown coloration.[100] Distinctive features of the mammalian kidney include the presence of the renal pelvis and renal pyramids, and of a clearly distinguishable cortex and medulla, which is due to the presence of elongated loops of Henle. Only the mammalian kidney has a bean shape, although there are some exceptions, such as the multilobed reniculate kidneys of pinnipeds, cetaceans and bears.[101][102] Most adult placental mammals have no remaining trace of the cloaca. In the embryo, the embryonic cloaca divides into a posterior region that becomes part of the anus, and an anterior region that has different fates depending on the sex of the individual: in females, it develops into the vestibule or urogenital sinus that receives the urethra and vagina, while in males it forms the entirety of the penile urethra.[102][103] However, the tenrecs, golden moles, and some shrews retain a cloaca as adults.[104] In marsupials, the genital tract is separate from the anus, but a trace of the original cloaca does remain externally.[102] Monotremes, which translates from Greek into "single hole", have a true cloaca.[105] Urine flows from the ureters into the cloaca in monotremes and into the bladder in placental mammals.[102]
Sound production

As in all other tetrapods, mammals have a larynx that can quickly open and close to produce sounds, and a supralaryngeal vocal tract which filters this sound. The lungs and surrounding musculature provide the air stream and pressure required to phonate. The larynx controls the pitch and volume of sound, but the strength the lungs exert to exhale also contributes to volume. More primitive mammals, such as the echidna, can only hiss, as sound is achieved solely through exhaling through a partially closed larynx. Other mammals phonate using vocal folds. The movement or tenseness of the vocal folds can result in many sounds such as purring and screaming. Mammals can change the position of the larynx, allowing them to breathe through the nose while swallowing through the mouth, and to form both oral and nasal sounds; nasal sounds, such as a dog whine, are generally soft sounds, and oral sounds, such as a dog bark, are generally loud.[106]
Some mammals have a large larynx and thus a low-pitched voice, namely the hammer-headed bat (Hypsignathus monstrosus) where the larynx can take up the entirety of the thoracic cavity while pushing the lungs, heart, and trachea into the abdomen.[107] Large vocal pads can also lower the pitch, as in the low-pitched roars of big cats.[108] The production of infrasound is possible in some mammals such as the African elephant (Loxodonta spp.) and baleen whales.[109][110] Small mammals with small larynxes have the ability to produce ultrasound, which can be detected by modifications to the middle ear and cochlea. Ultrasound is inaudible to birds and reptiles, which might have been important during the Mesozoic, when birds and reptiles were the dominant predators. This private channel is used by some rodents in, for example, mother-to-pup communication, and by bats when echolocating. Toothed whales also use echolocation, but, as opposed to the vocal membrane that extends upward from the vocal folds, they have a melon to manipulate sounds. Some mammals, namely the primates, have air sacs attached to the larynx, which may function to lower the resonances or increase the volume of sound.[106]
The vocal production system is controlled by the cranial nerve nuclei in the brain, and supplied by the recurrent laryngeal nerve and the superior laryngeal nerve, branches of the vagus nerve. The vocal tract is supplied by the hypoglossal nerve and facial nerves. Electrical stimulation of the periaqueductal gray (PEG) region of the mammalian midbrain elicit vocalizations. The ability to learn new vocalizations is only exemplified in humans, seals, cetaceans, elephants and possibly bats; in humans, this is the result of a direct connection between the motor cortex, which controls movement, and the motor neurons in the spinal cord.[106]
Fur

The primary function of the fur of mammals is thermoregulation. Others include protection, sensory purposes, waterproofing, and camouflage.[111] Different types of fur serve different purposes:[91]: 99
- Definitive – which may be shed after reaching a certain length
- Vibrissae – sensory hairs, most commonly whiskers
- Pelage – guard hairs, under-fur, and awn hair
- Spines – stiff guard hair used for defense (such as in porcupines)
- Bristles – long hairs usually used in visual signals. (such as a lion's mane)
- Velli – often called "down fur" which insulates newborn mammals
- Wool – long, soft and often curly
Thermoregulation
Hair length is not a factor in thermoregulation: for example, some tropical mammals such as sloths have the same length of fur length as some arctic mammals but with less insulation; and, conversely, other tropical mammals with short hair have the same insulating value as arctic mammals. The denseness of fur can increase an animal's insulation value, and arctic mammals especially have dense fur; for example, the musk ox has guard hairs measuring 30 cm (12 in) as well as a dense underfur, which forms an airtight coat, allowing them to survive in temperatures of −40 °C (−40 °F).[91]: 162–163 Some desert mammals, such as camels, use dense fur to prevent solar heat from reaching their skin, allowing the animal to stay cool; a camel's fur may reach 70 °C (158 °F) in the summer, but the skin stays at 40 °C (104 °F).[91]: 188 Aquatic mammals, conversely, trap air in their fur to conserve heat by keeping the skin dry.[91]: 162–163

Coloration
Mammalian coats are colored for a variety of reasons, the major selective pressures including camouflage, sexual selection, communication, and thermoregulation. Coloration in both the hair and skin of mammals is mainly determined by the type and amount of melanin; eumelanins for brown and black colors and pheomelanin for a range of yellowish to reddish colors, giving mammals an earth tone.[112][113] Some mammals have more vibrant colors; certain monkeys such mandrills and vervet monkeys, and opossums such as the Mexican mouse opossums and Derby's woolly opossums, have blue skin due to light diffraction in collagen fibers.[114] Many sloths appear green because their fur hosts green algae; this may be a symbiotic relation that affords camouflage to the sloths.[115]
Camouflage is a powerful influence in a large number of mammals, as it helps to conceal individuals from predators or prey.[116] In arctic and subarctic mammals such as the arctic fox (Alopex lagopus), collared lemming (Dicrostonyx groenlandicus), stoat (Mustela erminea), and snowshoe hare (Lepus americanus), seasonal color change between brown in summer and white in winter is driven largely by camouflage.[117] Some arboreal mammals, notably primates and marsupials, have shades of violet, green, or blue skin on parts of their bodies, indicating some distinct advantage in their largely arboreal habitat due to convergent evolution.[114]
Aposematism, warning off possible predators, is the most likely explanation of the black-and-white pelage of many mammals which are able to defend themselves, such as in the foul-smelling skunk and the powerful and aggressive honey badger.[118] Coat color is sometimes sexually dimorphic, as in many primate species.[119] Differences in female and male coat color may indicate nutrition and hormone levels, important in mate selection.[120] Coat color may influence the ability to retain heat, depending on how much light is reflected. Mammals with a darker colored coat can absorb more heat from solar radiation, and stay warmer, and some smaller mammals, such as voles, have darker fur in the winter. The white, pigmentless fur of arctic mammals, such as the polar bear, may reflect more solar radiation directly onto the skin.[91]: 166–167 [111] The dazzling black-and-white striping of zebras appear to provide some protection from biting flies.[121]
Reproductive system

Mammals are solely gonochoric (an animal is born with either male or female genitalia, as opposed to hermaphrodites where there is no such schism).[122] The penis is used both for urination and copulation in male placentals[123] and marsupials.[124] Depending on the species, an erection may be fueled by blood flow into vascular, spongy tissue or by muscular action. A penis may be contained in a prepuce when not erect, and some placentals also have a penis bone (baculum).[125] Marsupials typically have forked penises,[126] while the echidna penis generally has four heads with only two functioning.[127] The testicles of most mammals descend into the scrotum which is typically posterior to the penis but is often anterior in marsupials. Female mammals generally have a vulva (clitoris and labia) on the outside, while the internal system contains paired oviducts, 1–2 uteri, 1–2 cervices and a vagina.[128][129] Marsupials have two lateral vaginas and a medial vagina. The "vagina" of monotremes is better understood as a "urogenital sinus". The uterine systems of placental mammals can vary between a duplex, where there are two uteri and cervices which open into the vagina, a bipartite, where two uterine horns have a single cervix that connects to the vagina, a bicornuate, which consists where two uterine horns that are connected distally but separate medially creating a Y-shape, and a simplex, which has a single uterus.[130][131][91]: 220–221, 247

The ancestral condition for mammal reproduction is the birthing of relatively undeveloped young, either through direct vivipary or a short period as soft-shelled eggs. This is likely due to the fact that the torso could not expand due to the presence of epipubic bones. The oldest demonstration of this reproductive style is with Kayentatherium, which produced undeveloped perinates, but at much higher litter sizes than any modern mammal, 38 specimens.[132] Most modern mammals are viviparous, giving birth to live young. However, the five species of monotreme, the platypus and the four species of echidna, lay eggs. The monotremes have a sex-determination system different from that of most other mammals.[133] In particular, the sex chromosomes of a platypus are more like those of a chicken than those of a therian mammal.[134]
Viviparous mammals are in the subclass Theria; those living today are in the marsupial and placental infraclasses. Marsupials have a short gestation period, typically shorter than its estrous cycle and generally giving birth to a number of undeveloped newborns that then undergo further development; in many species, this takes place within a pouch-like sac, the marsupium, located in the front of the mother's abdomen. This is the plesiomorphic condition among viviparous mammals; the presence of epipubic bones in all non-placental mammals prevents the expansion of the torso needed for full pregnancy.[82] Even non-placental eutherians probably reproduced this way.[44] The placentals give birth to relatively complete and developed young, usually after long gestation periods.[135] They get their name from the placenta, which connects the developing fetus to the uterine wall to allow nutrient uptake.[136] In placental mammals, the epipubic is either completely lost or converted into the baculum; allowing the torso to be able to expand and thus birth developed offspring.[132]
The mammary glands of mammals are specialized to produce milk, the primary source of nutrition for newborns. The monotremes branched early from other mammals and do not have the teats seen in most mammals, but they do have mammary glands. The young lick the milk from a mammary patch on the mother's belly.[137] Compared to placental mammals, the milk of marsupials changes greatly in both production rate and in nutrient composition, due to the underdeveloped young. In addition, the mammary glands have more autonomy allowing them to supply separate milks to young at different development stages.[138] Lactose is the main sugar in placental mammal milk while monotreme and marsupial milk is dominated by oligosaccharides.[139] Weaning is the process in which a mammal becomes less dependent on their mother's milk and more on solid food.[140]
Endothermy
Nearly all mammals are endothermic ("warm-blooded"). Most mammals also have hair to help keep them warm. Like birds, mammals can forage or hunt in weather and climates too cold for ectothermic ("cold-blooded") reptiles and insects. Endothermy requires plenty of food energy, so mammals eat more food per unit of body weight than most reptiles.[141] Small insectivorous mammals eat prodigious amounts for their size. A rare exception, the naked mole-rat produces little metabolic heat, so it is considered an operational poikilotherm.[142] Birds are also endothermic, so endothermy is not unique to mammals.[143]
Species lifespan
Among mammals, species maximum lifespan varies significantly (for example the shrew has a lifespan of two years, whereas the oldest bowhead whale is recorded to be 211 years).[144] Although the underlying basis for these lifespan differences is still uncertain, numerous studies indicate that the ability to repair DNA damage is an important determinant of mammalian lifespan. In a 1974 study by Hart and Setlow,[145] it was found that DNA excision repair capability increased systematically with species lifespan among seven mammalian species. Species lifespan was observed to be robustly correlated with the capacity to recognize DNA double-strand breaks as well as the level of the DNA repair protein Ku80.[144] In a study of the cells from sixteen mammalian species, genes employed in DNA repair were found to be up-regulated in the longer-lived species.[146] The cellular level of the DNA repair enzyme poly ADP ribose polymerase was found to correlate with species lifespan in a study of 13 mammalian species.[147] Three additional studies of a variety of mammalian species also reported a correlation between species lifespan and DNA repair capability.[148][149][150]
Locomotion
Terrestrial

Most vertebrates—the amphibians, the reptiles and some mammals such as humans and bears—are plantigrade, walking on the whole of the underside of the foot. Many mammals, such as cats and dogs, are digitigrade, walking on their toes, the greater stride length allowing more speed. Some animals such as horses are unguligrade, walking on the tips of their toes. This even further increases their stride length and thus their speed.[151] A few mammals, namely the great apes, are also known to walk on their knuckles, at least for their front legs. Giant anteaters[152] and platypuses[153] are also knuckle-walkers. Some mammals are bipeds, using only two limbs for locomotion, which can be seen in, for example, humans and the great apes. Bipedal species have a larger field of vision than quadrupeds, conserve more energy and have the ability to manipulate objects with their hands, which aids in foraging. Instead of walking, some bipeds hop, such as kangaroos and kangaroo rats.[154][155]
Animals will use different gaits for different speeds, terrain and situations. For example, horses show four natural gaits, the slowest horse gait is the walk, then there are three faster gaits which, from slowest to fastest, are the trot, the canter and the gallop. Animals may also have unusual gaits that are used occasionally, such as for moving sideways or backwards. For example, the main human gaits are bipedal walking and running, but they employ many other gaits occasionally, including a four-legged crawl in tight spaces.[156] Mammals show a vast range of gaits, the order that they place and lift their appendages in locomotion. Gaits can be grouped into categories according to their patterns of support sequence. For quadrupeds, there are three main categories: walking gaits, running gaits and leaping gaits.[157] Walking is the most common gait, where some feet are on the ground at any given time, and found in almost all legged animals. Running is considered to occur when at some points in the stride all feet are off the ground in a moment of suspension.[156]
Arboreal

Arboreal animals frequently have elongated limbs that help them cross gaps, reach fruit or other resources, test the firmness of support ahead and, in some cases, to brachiate (swing between trees).[158] Many arboreal species, such as tree porcupines, silky anteaters, spider monkeys, and possums, use prehensile tails to grasp branches. In the spider monkey, the tip of the tail has either a bare patch or adhesive pad, which provides increased friction. Claws can be used to interact with rough substrates and reorient the direction of forces the animal applies. This is what allows squirrels to climb tree trunks that are so large to be essentially flat from the perspective of such a small animal. However, claws can interfere with an animal's ability to grasp very small branches, as they may wrap too far around and prick the animal's own paw. Frictional gripping is used by primates, relying upon hairless fingertips. Squeezing the branch between the fingertips generates frictional force that holds the animal's hand to the branch. However, this type of grip depends upon the angle of the frictional force, thus upon the diameter of the branch, with larger branches resulting in reduced gripping ability. To control descent, especially down large diameter branches, some arboreal animals such as squirrels have evolved highly mobile ankle joints that permit rotating the foot into a 'reversed' posture. This allows the claws to hook into the rough surface of the bark, opposing the force of gravity. Small size provides many advantages to arboreal species: such as increasing the relative size of branches to the animal, lower center of mass, increased stability, lower mass (allowing movement on smaller branches) and the ability to move through more cluttered habitat.[158] Size relating to weight affects gliding animals such as the sugar glider.[159] Some species of primate, bat and all species of sloth achieve passive stability by hanging beneath the branch. Both pitching and tipping become irrelevant, as the only method of failure would be losing their grip.[158]
Aerial
Bats are the only mammals that can truly fly. They fly through the air at a constant speed by moving their wings up and down (usually with some fore-aft movement as well). Because the animal is in motion, there is some airflow relative to its body which, combined with the velocity of the wings, generates a faster airflow moving over the wing. This generates a lift force vector pointing forwards and upwards, and a drag force vector pointing rearwards and upwards. The upwards components of these counteract gravity, keeping the body in the air, while the forward component provides thrust to counteract both the drag from the wing and from the body as a whole.[160]
The wings of bats are much thinner and consist of more bones than those of birds, allowing bats to maneuver more accurately and fly with more lift and less drag.[161][162] By folding the wings inwards towards their body on the upstroke, they use 35% less energy during flight than birds.[163] The membranes are delicate, ripping easily; however, the tissue of the bat's membrane is able to regrow, such that small tears can heal quickly.[164] The surface of their wings is equipped with touch-sensitive receptors on small bumps called Merkel cells, also found on human fingertips. These sensitive areas are different in bats, as each bump has a tiny hair in the center, making it even more sensitive and allowing the bat to detect and collect information about the air flowing over its wings, and to fly more efficiently by changing the shape of its wings in response.[165]
Fossorial and subterranean


A fossorial (from Latin fossor, meaning "digger") is an animal adapted to digging which lives primarily, but not solely, underground. Some examples are badgers, and naked mole-rats. Many rodent species are also considered fossorial because they live in burrows for most but not all of the day. Species that live exclusively underground are subterranean, and those with limited adaptations to a fossorial lifestyle sub-fossorial. Some organisms are fossorial to aid in temperature regulation while others use the underground habitat for protection from predators or for food storage.[166]
Fossorial mammals have a fusiform body, thickest at the shoulders and tapering off at the tail and nose. Unable to see in the dark burrows, most have degenerated eyes, but degeneration varies between species; pocket gophers, for example, are only semi-fossorial and have very small yet functional eyes, in the fully fossorial marsupial mole the eyes are degenerated and useless, talpa moles have vestigial eyes and the cape golden mole has a layer of skin covering the eyes. External ears flaps are also very small or absent. Truly fossorial mammals have short, stout legs as strength is more important than speed to a burrowing mammal, but semi-fossorial mammals have cursorial legs. The front paws are broad and have strong claws to help in loosening dirt while excavating burrows, and the back paws have webbing, as well as claws, which aids in throwing loosened dirt backwards. Most have large incisors to prevent dirt from flying into their mouth.[167]
Many fossorial mammals such as shrews, hedgehogs, and moles were classified under the now obsolete order Insectivora.[168]
Aquatic
Fully aquatic mammals, the cetaceans and sirenians, have lost their legs and have a tail fin to propel themselves through the water. Flipper movement is continuous. Whales swim by moving their tail fin and lower body up and down, propelling themselves through vertical movement, while their flippers are mainly used for steering. Their skeletal anatomy allows them to be fast swimmers. Most species have a dorsal fin to prevent themselves from turning upside-down in the water.[169][170] The flukes of sirenians are raised up and down in long strokes to move the animal forward, and can be twisted to turn. The forelimbs are paddle-like flippers which aid in turning and slowing.[171]
Semi-aquatic mammals, like pinnipeds, have two pairs of flippers on the front and back, the fore-flippers and hind-flippers. The elbows and ankles are enclosed within the body.[172][173] Pinnipeds have several adaptions for reducing drag. In addition to their streamlined bodies, they have smooth networks of muscle bundles in their skin that may increase laminar flow and make it easier for them to slip through water. They also lack arrector pili, so their fur can be streamlined as they swim.[174] They rely on their fore-flippers for locomotion in a wing-like manner similar to penguins and sea turtles.[175] Fore-flipper movement is not continuous, and the animal glides between each stroke.[173] Compared to terrestrial carnivorans, the fore-limbs are reduced in length, which gives the locomotor muscles at the shoulder and elbow joints greater mechanical advantage;[172] the hind-flippers serve as stabilizers.[174] Other semi-aquatic mammals include beavers, hippopotamuses, otters and platypuses.[176] Hippos are very large semi-aquatic mammals, and their barrel-shaped bodies have graviportal skeletal structures,[177] adapted to carrying their enormous weight, and their specific gravity allows them to sink and move along the bottom of a river.[178]
Behavior
Communication and vocalization

Many mammals communicate by vocalizing. Vocal communication serves many purposes, including in mating rituals, as warning calls,[180] to indicate food sources, and for social purposes. Males often call during mating rituals to ward off other males and to attract females, as in the roaring of lions and red deer.[181] The songs of the humpback whale may be signals to females;[182] they have different dialects in different regions of the ocean.[183] Social vocalizations include the territorial calls of gibbons, and the use of frequency in greater spear-nosed bats to distinguish between groups.[184] The vervet monkey gives a distinct alarm call for each of at least four different predators, and the reactions of other monkeys vary according to the call. For example, if an alarm call signals a python, the monkeys climb into the trees, whereas the eagle alarm causes monkeys to seek a hiding place on the ground.[179] Prairie dogs similarly have complex calls that signal the type, size, and speed of an approaching predator.[185] Elephants communicate socially with a variety of sounds including snorting, screaming, trumpeting, roaring and rumbling. Some of the rumbling calls are infrasonic, below the hearing range of humans, and can be heard by other elephants up to 6 miles (9.7 km) away at still times near sunrise and sunset.[186]
Mammals signal by a variety of means. Many give visual anti-predator signals, as when deer and gazelle stot, honestly indicating their fit condition and their ability to escape,[187][188] or when white-tailed deer and other prey mammals flag with conspicuous tail markings when alarmed, informing the predator that it has been detected.[189] Many mammals make use of scent-marking, sometimes possibly to help defend territory, but probably with a range of functions both within and between species.[190][191][192] Microbats and toothed whales including oceanic dolphins vocalize both socially and in echolocation.[193][194][195]
Feeding
To maintain a high constant body temperature is energy expensive—mammals therefore need a nutritious and plentiful diet. While the earliest mammals were probably predators, different species have since adapted to meet their dietary requirements in a variety of ways. Some eat other animals—this is a carnivorous diet (and includes insectivorous diets). Other mammals, called herbivores, eat plants, which contain complex carbohydrates such as cellulose. An herbivorous diet includes subtypes such as granivory (seed eating), folivory (leaf eating), frugivory (fruit eating), nectarivory (nectar eating), gummivory (gum eating) and mycophagy (fungus eating). The digestive tract of an herbivore is host to bacteria that ferment these complex substances, and make them available for digestion, which are either housed in the multichambered stomach or in a large cecum.[95] Some mammals are coprophagous, consuming feces to absorb the nutrients not digested when the food was first ingested.[91]: 131–137 An omnivore eats both prey and plants. Carnivorous mammals have a simple digestive tract because the proteins, lipids and minerals found in meat require little in the way of specialized digestion. Exceptions to this include baleen whales who also house gut flora in a multi-chambered stomach, like terrestrial herbivores.[196]
The size of an animal is also a factor in determining diet type (Allen's rule). Since small mammals have a high ratio of heat-losing surface area to heat-generating volume, they tend to have high energy requirements and a high metabolic rate. Mammals that weigh less than about 18 ounces (510 g; 1.1 lb) are mostly insectivorous because they cannot tolerate the slow, complex digestive process of an herbivore. Larger animals, on the other hand, generate more heat and less of this heat is lost. They can therefore tolerate either a slower collection process (carnivores that feed on larger vertebrates) or a slower digestive process (herbivores).[197] Furthermore, mammals that weigh more than 18 ounces (510 g; 1.1 lb) usually cannot collect enough insects during their waking hours to sustain themselves. The only large insectivorous mammals are those that feed on huge colonies of insects (ants or termites).[198]


Some mammals are omnivores and display varying degrees of carnivory and herbivory, generally leaning in favor of one more than the other. Since plants and meat are digested differently, there is a preference for one over the other, as in bears where some species may be mostly carnivorous and others mostly herbivorous.[200] They are grouped into three categories: mesocarnivory (50–70% meat), hypercarnivory (70% and greater of meat), and hypocarnivory (50% or less of meat). The dentition of hypocarnivores consists of dull, triangular carnassial teeth meant for grinding food. Hypercarnivores, however, have conical teeth and sharp carnassials meant for slashing, and in some cases strong jaws for bone-crushing, as in the case of hyenas, allowing them to consume bones; some extinct groups, notably the Machairodontinae, had saber-shaped canines.[199]
Some physiological carnivores consume plant matter and some physiological herbivores consume meat. From a behavioral aspect, this would make them omnivores, but from the physiological standpoint, this may be due to zoopharmacognosy. Physiologically, animals must be able to obtain both energy and nutrients from plant and animal materials to be considered omnivorous. Thus, such animals are still able to be classified as carnivores and herbivores when they are just obtaining nutrients from materials originating from sources that do not seemingly complement their classification.[201] For example, it is well documented that some ungulates such as giraffes, camels, and cattle, will gnaw on bones to consume particular minerals and nutrients.[202] Also, cats, which are generally regarded as obligate carnivores, occasionally eat grass to regurgitate indigestible material (such as hairballs), aid with hemoglobin production, and as a laxative.[203]
Many mammals, in the absence of sufficient food requirements in an environment, suppress their metabolism and conserve energy in a process known as hibernation.[204] In the period preceding hibernation, larger mammals, such as bears, become polyphagic to increase fat stores, whereas smaller mammals prefer to collect and stash food.[205] The slowing of the metabolism is accompanied by a decreased heart and respiratory rate, as well as a drop in internal temperatures, which can be around ambient temperature in some cases. For example, the internal temperatures of hibernating arctic ground squirrels can drop to −2.9 °C (26.8 °F); however, the head and neck always stay above 0 °C (32 °F).[206] A few mammals in hot environments aestivate in times of drought or extreme heat, for example the fat-tailed dwarf lemur (Cheirogaleus medius).[207]
Drinking

By necessity, terrestrial animals in captivity become accustomed to drinking water, but most free-roaming animals stay hydrated through the fluids and moisture in fresh food,[208] and learn to actively seek foods with high fluid content.[209] When conditions impel them to drink from bodies of water, the methods and motions differ greatly among species.[210]
Cats, canines, and ruminants all lower the neck and lap in water with their powerful tongues.[210] Cats and canines lap up water with the tongue in a spoon-like shape.[211] Canines lap water by scooping it into their mouth with a tongue which has taken the shape of a ladle. However, with cats, only the tip of their tongue (which is smooth) touches the water, and then the cat quickly pulls its tongue back into its mouth which soon closes; this results in a column of liquid being pulled into the cat's mouth, which is then secured by its mouth closing.[212] Ruminants and most other herbivores partially submerge the tip of the mouth in order to draw in water by means of a plunging action with the tongue held straight.[213] Cats drink at a significantly slower pace than ruminants, who face greater natural predation hazards.[210]
Many desert animals do not drink even if water becomes available, but rely on eating succulent plants.[210] In cold and frozen environments, some animals like hares, tree squirrels, and bighorn sheep resort to consuming snow and icicles.[214] In savannas, the drinking method of giraffes has been a source of speculation for its apparent defiance of gravity; the most recent theory contemplates the animal's long neck functions like a plunger pump.[215] Uniquely, elephants draw water into their trunks and squirt it into their mouths.[210]Intelligence
In intelligent mammals, such as primates, the cerebrum is larger relative to the rest of the brain. Intelligence itself is not easy to define, but indications of intelligence include the ability to learn, matched with behavioral flexibility. Rats, for example, are considered to be highly intelligent, as they can learn and perform new tasks, an ability that may be important when they first colonize a fresh habitat. In some mammals, food gathering appears to be related to intelligence: a deer feeding on plants has a brain smaller than a cat, which must think to outwit its prey.[198]

Tool use by animals may indicate different levels of learning and cognition. The sea otter uses rocks as essential and regular parts of its foraging behaviour (smashing abalone from rocks or breaking open shells), with some populations spending 21% of their time making tools.[216] Other tool use, such as chimpanzees using twigs to "fish" for termites, may be developed by watching others use tools and may even be a true example of animal teaching.[217] Tools may even be used in solving puzzles in which the animal appears to experience a "Eureka moment".[218] Other mammals that do not use tools, such as dogs, can also experience a Eureka moment.[219]
Brain size was previously considered a major indicator of the intelligence of an animal. Since most of the brain is used for maintaining bodily functions, greater ratios of brain to body mass may increase the amount of brain mass available for more complex cognitive tasks. Allometric analysis indicates that mammalian brain size scales at approximately the 2⁄3 or 3⁄4 exponent of the body mass. Comparison of a particular animal's brain size with the expected brain size based on such allometric analysis provides an encephalisation quotient that can be used as another indication of animal intelligence.[220] Sperm whales have the largest brain mass of any animal on earth, averaging 8,000 cubic centimetres (490 cu in) and 7.8 kilograms (17 lb) in mature males.[221]
Self-awareness appears to be a sign of abstract thinking. Self-awareness, although not well-defined, is believed to be a precursor to more advanced processes such as metacognitive reasoning. The traditional method for measuring this is the mirror test, which determines if an animal possesses the ability of self-recognition.[222] Mammals that have passed the mirror test include Asian elephants (some pass, some do not);[223] chimpanzees;[224] bonobos;[225] orangutans;[226] humans, from 18 months (mirror stage);[227] common bottlenose dolphins;[a][228] orcas;[229] and false killer whales.[229]
Social structure

Eusociality is the highest level of social organization. These societies have an overlap of adult generations, the division of reproductive labor and cooperative caring of young. Usually insects, such as bees, ants and termites, have eusocial behavior, but it is demonstrated in two rodent species: the naked mole-rat[230] and the Damaraland mole-rat.[231]
Presociality is when animals exhibit more than just sexual interactions with members of the same species, but fall short of qualifying as eusocial. That is, presocial animals can display communal living, cooperative care of young, or primitive division of reproductive labor, but they do not display all of the three essential traits of eusocial animals. Humans and some species of Callitrichidae (marmosets and tamarins) are unique among primates in their degree of cooperative care of young.[232] Harry Harlow set up an experiment with rhesus monkeys, presocial primates, in 1958; the results from this study showed that social encounters are necessary in order for the young monkeys to develop both mentally and sexually.[233]
A fission–fusion society is a society that changes frequently in its size and composition, making up a permanent social group called the "parent group". Permanent social networks consist of all individual members of a community and often varies to track changes in their environment. In a fission–fusion society, the main parent group can fracture (fission) into smaller stable subgroups or individuals to adapt to environmental or social circumstances. For example, a number of males may break off from the main group in order to hunt or forage for food during the day, but at night they may return to join (fusion) the primary group to share food and partake in other activities. Many mammals exhibit this, such as primates (for example orangutans and spider monkeys),[234] elephants,[235] spotted hyenas,[236] lions,[237] and dolphins.[238]
Solitary animals defend a territory and avoid social interactions with the members of its species, except during breeding season. This is to avoid resource competition, as two individuals of the same species would occupy the same niche, and to prevent depletion of food.[239] A solitary animal, while foraging, can also be less conspicuous to predators or prey.[240]

In a hierarchy, individuals are either dominant or submissive. A despotic hierarchy is where one individual is dominant while the others are submissive, as in wolves and lemurs,[241] and a pecking order is a linear ranking of individuals where there is a top individual and a bottom individual. Pecking orders may also be ranked by sex, where the lowest individual of a sex has a higher ranking than the top individual of the other sex, as in hyenas.[242] Dominant individuals, or alphas, have a high chance of reproductive success, especially in harems where one or a few males (resident males) have exclusive breeding rights to females in a group.[243] Non-resident males can also be accepted in harems, but some species, such as the common vampire bat (Desmodus rotundus), may be more strict.[244]
Some mammals are perfectly monogamous, meaning that they mate for life and take no other partners (even after the original mate's death), as with wolves, Eurasian beavers, and otters.[245][246] There are three types of polygamy: either one or multiple dominant males have breeding rights (polygyny), multiple males that females mate with (polyandry), or multiple males have exclusive relations with multiple females (polygynandry). It is much more common for polygynous mating to happen, which, excluding leks, are estimated to occur in up to 90% of mammals.[247] Lek mating occurs when males congregate around females and try to attract them with various courtship displays and vocalizations, as in harbor seals.[248]
All higher mammals (excluding monotremes) share two major adaptations for care of the young: live birth and lactation. These imply a group-wide choice of a degree of parental care. They may build nests and dig burrows to raise their young in, or feed and guard them often for a prolonged period of time. Many mammals are K-selected, and invest more time and energy into their young than do r-selected animals. When two animals mate, they both share an interest in the success of the offspring, though often to different extremes. Mammalian females exhibit some degree of maternal aggression, another example of parental care, which may be targeted against other females of the species or the young of other females; however, some mammals may "aunt" the infants of other females, and care for them. Mammalian males may play a role in child rearing, as with tenrecs, however this varies species to species, even within the same genus. For example, the males of the southern pig-tailed macaque (Macaca nemestrina) do not participate in child care, whereas the males of the Japanese macaque (M. fuscata) do.[249]
Humans and other mammals
В человеческой культуре

Нечеловеческие млекопитающие играют самые разнообразные роли в человеческой культуре. Они являются самыми популярными домашними животными : десятки миллионов собак, кошек и других животных, включая кроликов и мышей, содержатся семьями по всему миру. [250] [251] [252] Млекопитающие, такие как мамонты , лошади и олени, являются одними из самых ранних предметов искусства, их можно найти в верхнего палеолита, наскальных рисунках таких как в Ласко . [253] Крупные художники, такие как Альбрехт Дюрер , Джордж Стаббс и Эдвин Ландсир, известны своими портретами млекопитающих. [254] На многие виды млекопитающих охотились ради развлечения и ради еды; Олени и кабаны особенно популярны в качестве охотничьих животных . [255] [256] [257] Млекопитающие, такие как лошади и собаки, широко участвуют в спортивных гонках, часто в сочетании со ставками на результат . [258] [259] Существует противоречие между ролью животных как товарищей по отношению к человеку и их существованием как личностей с собственными правами . [260] Млекопитающие также играют самые разнообразные роли в литературе. [261] [262] [263] фильм, [264] мифология и религия. [265] [266] [267]
Использование и важность

Одомашнивание млекопитающих сыграло важную роль в развитии сельского хозяйства и цивилизации в эпоху неолита , заставив фермеров заменить охотников-собирателей по всему миру. [б] [269] Этот переход от охоты и собирательства к выпасу стад и выращиванию сельскохозяйственных культур стал важным шагом в истории человечества. Новая сельскохозяйственная экономика, основанная на одомашненных млекопитающих, вызвала «радикальную реструктуризацию человеческого общества, глобальные изменения в биоразнообразии и значительные изменения в формах рельефа Земли и ее атмосфере... важные результаты». [270]
Домашние млекопитающие составляют большую часть поголовья скота , выращиваемого на мясо во всем мире. В их число входят (2009 г.) около 1,4 миллиарда крупного рогатого скота , 1 миллиард овец , 1 миллиард домашних свиней , [271] [272] и (1985) более 700 миллионов кроликов. [273] Рабочие домашние животные, включая крупный рогатый скот и лошадей, использовались для работы и транспорта с самого начала сельского хозяйства, их численность сокращается с появлением механизированного транспорта и сельскохозяйственных машин . В 2004 году они по-прежнему обеспечивали около 80% электроэнергии преимущественно мелких ферм в странах третьего мира и около 20% мирового транспорта, опять же в основном в сельских районах. В горных районах, непригодных для колесного транспорта, вьючные животные . грузы продолжают перевозить [274] Из шкур млекопитающих делают кожу для обуви , одежды и обивки . Шерсть млекопитающих, включая овец, коз и альпак, веками использовалась для изготовления одежды. [275] [276]
Млекопитающие играют важную роль в науке в качестве экспериментальных животных , как в фундаментальных биологических исследованиях, таких как генетика, так и в области генетики. [278] и в разработке новых лекарств, которые должны быть тщательно протестированы, чтобы продемонстрировать их безопасность . [279] используются миллионы млекопитающих, особенно мышей и крыс. в экспериментах Ежегодно [280] — Нокаутная мышь это генетически модифицированная мышь с инактивированным геном , замененным или разрушенным искусственным участком ДНК. Они позволяют изучать секвенированные гены, функции которых неизвестны. [281] Небольшой процент млекопитающих — это приматы, не относящиеся к человеку, которые используются в исследованиях из-за их сходства с людьми. [282] [283] [284]
Несмотря на пользу, которую одомашненные млекопитающие принесли для развития человечества, люди оказывают все более пагубное воздействие на диких млекопитающих во всем мире. Подсчитано, что масса всех диких млекопитающих сократилась до 4% от всех млекопитающих, при этом 96% млекопитающих сейчас составляют люди и их домашний скот (см. рисунок). Фактически наземные дикие млекопитающие составляют лишь 2% всех млекопитающих. [285] [286]
Гибриды


Гибриды — это потомство, полученное в результате скрещивания двух генетически различных особей, что обычно приводит к высокой степени гетерозиготности, хотя гибрид и гетерозигота не являются синонимами. Преднамеренная или случайная гибридизация двух или более видов близкородственных животных посредством разведения в неволе — это человеческая деятельность, которая существует на протяжении тысячелетий и развивается в экономических целях. [287] Гибриды между различными подвидами внутри вида (например, между бенгальским тигром и сибирским тигром ) известны как внутривидовые гибриды. Гибриды между разными видами внутри одного рода (например, между львами и тиграми) известны как межвидовые гибриды или скрещивания. Гибриды между разными родами (например, между овцами и козами) известны как межродовые гибриды. [288] Естественные гибриды будут возникать в гибридных зонах , где две популяции видов одного и того же рода или виды, живущие в одних и тех же или соседних областях, будут скрещиваться друг с другом. Некоторые гибриды были признаны видами, например красный волк (хотя это спорный вопрос). [289]
Искусственный отбор , преднамеренное селекционное разведение домашних животных, используется для восстановления недавно вымерших животных в попытке создать породу животных с фенотипом , напоминающим вымершего предка дикого типа . Размножающийся (внутривидовой) гибрид может быть очень похож на вымерший дикий тип по внешнему виду, экологической нише и в некоторой степени генетике, но исходный генофонд этого дикого типа навсегда утрачен с его исчезновением . В результате выведенные породы являются в лучшем случае расплывчатыми двойниками вымерших диких типов, так же как крупный рогатый скот Хека имеет сходство с зубром . [290]
Чистокровные дикие виды, эволюционировавшие в соответствии с определенной экологией, могут оказаться под угрозой исчезновения [291] посредством процесса генетического загрязнения , неконтролируемой гибридизации, интрогрессии генетического затопления, что приводит к гомогенизации или вытеснению гибридных видов из гетерозисных конкуренции . [292] Когда новые популяции завозятся или селективно разводятся людьми, или когда изменение среды обитания приводит к контакту ранее изолированных видов, возможно исчезновение некоторых видов, особенно редких разновидностей. [293] Скрещивание может поглотить более редкий генофонд и создать гибриды, истощая чистокровный генофонд. Например, находящийся под угрозой исчезновения дикий водяной буйвол находится под наибольшей угрозой исчезновения из-за генетического загрязнения домашним водяным буйволом . Такие вымирания не всегда очевидны с морфологической точки зрения. Некоторая степень потока генов является нормальным эволюционным процессом, тем не менее, гибридизация ставит под угрозу существование редких видов. [294] [295]
Угрозы

Утрата видов из экологических сообществ, дефаунация , в первую очередь обусловлена деятельностью человека. [296] Это привело к опустошению лесов , экологическим сообществам, лишенным крупных позвоночных. [297] [298] Во время четвертичного вымирания массовое вымирание представителей мегафауны совпало с появлением людей, что свидетельствует о человеческом влиянии. Одна из гипотез заключается в том, что люди охотились на крупных млекопитающих, таких как шерстистый мамонт , и привели к их исчезновению. [299] [300] 2019 год о глобальной оценке биоразнообразия и экосистемных услуг за В докладе IPBES говорится, что общая биомасса диких млекопитающих сократилась на 82 процента с момента зарождения человеческой цивилизации. [301] [302] Дикие животные составляют всего 4% биомассы млекопитающих на Земле, тогда как люди и их одомашненные животные составляют 96%. [286]
По прогнозам, в ближайшем будущем различные виды вымрут . [303] среди них носорог , [304] жирафы , [305] и виды приматов [306] и панголины . [307] Согласно отчету WWF « Живая планета» за 2020 год , популяции позвоночных диких животных сократились на 68% с 1970 года в результате деятельности человека, особенно чрезмерного потребления , роста населения и интенсивного земледелия , что является свидетельством того, что люди спровоцировали шестое массовое вымирание . [308] [309] Одна только охота угрожает сотням видов млекопитающих по всему миру. [310] [311] Ученые утверждают, что растущий спрос на мясо способствует утрате биоразнообразия , поскольку это является важным фактором вырубки лесов и разрушения среды обитания ; богатые видами места обитания, такие как значительная часть тропических лесов Амазонки , превращаются в сельскохозяйственные угодья для производства мяса. [312] [313] [314] Еще одним фактором влияния является чрезмерная охота и браконьерство , которые могут сократить общую популяцию охотничьих животных. [315] особенно те, что расположены вблизи деревень, [316] как в случае с пекари . [317] Последствия браконьерства особенно заметны в торговле слоновой костью африканских слонов. [318] , подвергаются риску запутывания в рыболовных снастях, Морские млекопитающие, особенно китообразные при этом смертность от выброса колеблется от 65 000 до 86 000 особей в год. [319]
Внимание уделяется исчезающим видам во всем мире, в частности, через Конвенцию о биологическом разнообразии , также известную как Соглашение Рио-де-Жанейро, в которую входят 189 подписавших стран, которые сосредоточены на выявлении исчезающих видов и мест обитания. [320] Другой известной природоохранной организацией является МСОП, в который входят более 1200 правительственных и неправительственных организаций. [321]
Недавние вымирания можно напрямую объяснить влиянием человека. [322] [296] МСОП характеризует «недавнее» вымирание как такое, которое произошло после отсечки в 1500 лет. [323] и около 80 видов млекопитающих вымерли с тех пор и в 2015 году. [324] Некоторые виды, такие как олень Пер-Давида. [325] вымерли в дикой природе и выживают исключительно в популяциях, содержащихся в неволе. Другие виды, такие как пантера Флориды , экологически вымерли , выживая в таких небольших количествах, что практически не оказывают никакого влияния на экосистему. [326] : 318 Другие популяции вымерли лишь локально (искоренены), все еще существуют в других местах, но их распространение сократилось. [326] : 75–77 как и с исчезновением серых китов в Атлантике . [327]
См. также
- Список родов млекопитающих - современные млекопитающие
- Список маммологов
- Список однопроходных и сумчатых
- Список плацентарных млекопитающих
- Список доисторических млекопитающих
- Список млекопитающих, находящихся под угрозой исчезновения
- Списки млекопитающих по численности популяции
- Списки млекопитающих по регионам
- Млекопитающие, описанные в 2000-х годах.
- Млекопитающие в культуре
- Мелкое млекопитающее
Примечания
- ^ Уменьшение задержки подхода к зеркалу, повторяющиеся кружения головой и пристальное рассмотрение отмеченных областей считались признаками самоузнания, поскольку у них нет рук и они не могут коснуться отмеченных областей. [228]
- ↑ Даймонд далее обсуждал этот вопрос в своей книге « Ружья, микробы и сталь» 1997 года . [268]
Ссылки
- ^ Льюис, Чарльтон Т.; Короткий, Чарльз (1879). «мама» . Латинский словарь . Цифровая библиотека Персея. Архивировано из оригинала 29 сентября 2022 года . Проверено 29 сентября 2022 г.
- ^ Воган Т.А., Райан Дж.М., Чаплевски, Нью-Джерси (2013). «Классификация млекопитающих». Маммология (6-е изд.). Джонс и Бартлетт Обучение. ISBN 978-1-284-03209-3 .
- ^ Симпсон Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Американский музей естественной истории . 85 .
- ↑ Перейти обратно: Перейти обратно: а б Салай Ф.С. (1999). «Классификация млекопитающих выше видового уровня: Обзор». Журнал палеонтологии позвоночных . 19 (1): 191–195. дои : 10.1080/02724634.1999.10011133 . JSTOR 4523980 .
- ↑ Перейти обратно: Перейти обратно: а б Уилсон Д.Э. , Ридер Д.М., ред. (2005). «Предисловие и вводный материал» . Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. п. XXVI. ISBN 978-0-8018-8221-0 . OCLC 62265494 .
- ^ «Млекопитающие» . Красный список исчезающих видов МСОП . Международный союз охраны природы (МСОП). Апрель 2010 г. Архивировано из оригинала 3 сентября 2016 г. Проверено 23 августа 2016 г. .
- ^ Бургин С.Дж., Колелла Дж.П., Кан П.Л., Апхэм Н.С. (1 февраля 2018 г.). «Сколько существует видов млекопитающих?» . Журнал маммологии . 99 (1): 1–14. дои : 10.1093/jmammal/gyx147 .
- ^ Роу Т. (1988). «Определение, диагностика и происхождение млекопитающих» (PDF) . Журнал палеонтологии позвоночных . 8 (3): 241–264. Бибкод : 1988JVPal...8..241R . дои : 10.1080/02724634.1988.10011708 . Архивировано (PDF) из оригинала 18 января 2024 года . Проверено 25 января 2024 г.
- ^ Лайель С. (1871 г.). Студенческие элементы геологии . Лондон: Джон Мюррей. п. 347. ИСБН 978-1-345-18248-4 .
- ^ Чифелли Р.Л., Дэвис Б.М. (декабрь 2003 г.). «Палеонтология. Происхождение сумчатых». Наука . 302 (5652): 1899–1900. дои : 10.1126/science.1092272 . ПМИД 14671280 . S2CID 83973542 .
- ^ Кемп Т.С. (2005). Происхождение и эволюция млекопитающих (PDF) . Соединенное Королевство: Издательство Оксфордского университета. п. 3. ISBN 978-0-19-850760-4 . OCLC 232311794 . Архивировано (PDF) из оригинала 26 сентября 2023 года . Проверено 25 января 2024 г.
- ^ Датта ПМ (2005). «Самое раннее млекопитающее с поперечно расширенными верхними коренными зубами из формации Тики позднего триаса (карния), бассейн Южной Ревы Гондваны, Индия». Журнал палеонтологии позвоночных . 25 (1): 200–207. doi : 10.1671/0272-4634(2005)025[0200:EMWTEU]2.0.CO;2 . S2CID 131236175 .
- ^ Луо ZX, Мартин Т (2007). «Анализ молярной структуры и филогении родов докодонтов» (PDF) . Бюллетень Музея естественной истории Карнеги . 39 : 27–47. doi : 10.2992/0145-9058(2007)39[27:AOMSAP]2.0.CO;2 . S2CID 29846648 . Архивировано из оригинала (PDF) 3 марта 2016 года . Проверено 8 апреля 2013 г.
- ^ Аверьянов Александр О.; Войта, Леонид Л. (март 2024 г.). «Предполагаемое триасовое стволовое млекопитающее Tikitherium copyi — неогеновая землеройка» . Журнал эволюции млекопитающих . 31 (1). дои : 10.1007/s10914-024-09703-w . ISSN 1064-7554 .
- ^ Маккенна MC, Bell SG (1997). Классификация млекопитающих выше видового уровня . Нью-Йорк: Издательство Колумбийского университета. ISBN 978-0-231-11013-6 . OCLC 37345734 .
- ^ Нильссон М.А., Чураков Г., Зоммер М., Тран Н.В., Земанн А., Брозиус Дж., Шмитц Дж. (июль 2010 г.). «Отслеживание эволюции сумчатых с помощью архаичных геномных вставок ретропозонов» . ПЛОС Биология . 8 (7): e1000436. дои : 10.1371/journal.pbio.1000436 . ПМЦ 2910653 . ПМИД 20668664 .
- ^ Кригс Дж.О., Чураков Г., Кифманн М., Джордан У., Брозиус Дж., Шмитц Дж. (апрель 2006 г.). «Ретропозированные элементы как архивы эволюционной истории плацентарных млекопитающих» . ПЛОС Биология . 4 (4): е91. doi : 10.1371/journal.pbio.0040091 . ПМЦ 1395351 . ПМИД 16515367 .
- ↑ Перейти обратно: Перейти обратно: а б Нишихара Х., Маруяма С., Окада Н. (март 2009 г.). «Анализ ретропозонов и недавние геологические данные предполагают почти одновременное расхождение трех надотрядов млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (13): 5235–5240. Бибкод : 2009PNAS..106.5235N . дои : 10.1073/pnas.0809297106 . ПМЦ 2655268 . ПМИД 19286970 .
- ^ Спрингер М.С., Мерфи В.Дж., Эйзирик Э., О'Брайен С.Дж. (февраль 2003 г.). «Диверсификация плацентарных млекопитающих и граница мелового и третичного периодов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (3): 1056–1061. Бибкод : 2003PNAS..100.1056S . дои : 10.1073/pnas.0334222100 . ПМК 298725 . ПМИД 12552136 .
- ^ Тарвер Дж.Э., Дос Рейс М., Мирараб С., Моран Р.Дж., Паркер С., О'Рейли Дж.Э. и др. (январь 2016 г.). «Взаимоотношения плацентарных млекопитающих и пределы филогенетических выводов» . Геномная биология и эволюция . 8 (2): 330–344. дои : 10.1093/gbe/evv261 . hdl : 1983/64d6e437-3320-480d-a16c-2e5b2e6b61d4 . ПМЦ 4779606 . ПМИД 26733575 .
- ^ Альварес-Карретеро С., Тамури А.У., Баттини М., Насименто Ф.Ф., Карлайл Э., Ашер Р.Дж., Ян З., Донохью ПК и др. (2022). «Хронология эволюции млекопитающих на уровне вида, объединяющая филогеномные данные» . Природа . 602 (7896): 263–267. Бибкод : 2022Natur.602..263A . дои : 10.1038/s41586-021-04341-1 . hdl : 1983/de841853-d57b-40d9-876f-9bfcf7253f12 . ПМИД 34937052 . S2CID 245438816 .
- ^ Альварес-Карретеро, Сандра; Тамури, Асиф; Баттини, Маттео; Насименто, Фабрисия Ф.; Карлайл, Эмили; Ашер, Роберт; Ян, Цзыхэн; Донохью, Филип; дос Рейс, Марио (2021). «Данные для временной шкалы эволюции млекопитающих на уровне видов с интеграцией филогеномных данных» . Фиговая доля . doi : 10.6084/m9.figshare.14885691.v1 . Архивировано из оригинала 16 декабря 2023 года . Проверено 11 ноября 2023 г.
- ^ Мэн Дж, Ван Ю, Ли С (апрель 2011 г.). «Переходное среднее ухо млекопитающих от нового эвтриконодонта Джехол мелового периода». Природа . 472 (7342): 181–185. Бибкод : 2011Natur.472..181M . дои : 10.1038/nature09921 . ПМИД 21490668 . S2CID 4428972 .
- ^ Альберг П.Е., Милнер А.Р. (апрель 1994 г.). «Происхождение и ранняя диверсификация четвероногих». Природа . 368 (6471): 507–514. Бибкод : 1994Natur.368..507A . дои : 10.1038/368507a0 . S2CID 4369342 .
- ^ «Амниота-Палеос» . Архивировано из оригинала 20 декабря 2010 года.
- ^ «Обзор Synapsida – Палеос» . Архивировано из оригинала 20 декабря 2010 года.
- ↑ Перейти обратно: Перейти обратно: а б с Кемп Т.С. (июль 2006 г.). «Происхождение и раннее распространение терапсидных млекопитающих рептилий: палеобиологическая гипотеза» (PDF) . Журнал эволюционной биологии . 19 (4): 1231–1247. дои : 10.1111/j.1420-9101.2005.01076.x . ПМИД 16780524 . S2CID 3184629 . Архивировано из оригинала (PDF) 8 марта 2021 года . Проверено 14 января 2012 г.
- ↑ Перейти обратно: Перейти обратно: а б Беннетт А.Ф., Рубен Дж.А. (1986). «Метаболический и терморегуляторный статус терапсидов». В Hotton III N, MacLean JJ, Roth J, Roth EC (ред.). Экология и биология млекопитающих рептилий . Вашингтон, округ Колумбия: Издательство Смитсоновского института. стр. 207–218. ISBN 978-0-87474-524-5 .
- ^ Кермак Д.М., Кермак К.А. (1984). Эволюция характеров млекопитающих . Вашингтон, округ Колумбия: Крум Хелм. ISBN 978-0-7099-1534-8 . ОСЛК 10710687 .
- ^ Араужо; и др. (28 июля 2022 г.). «Биомеханика внутреннего уха показывает позднетриасовое происхождение эндотермии млекопитающих». Природа . 607 (7920): 726–731. Бибкод : 2022Natur.607..726A . дои : 10.1038/s41586-022-04963-z . ПМИД 35859179 . S2CID 236245230 .
- ^ Таннер Л.Х., Лукас С.Г., Чепмен М.Г. (2004). «Оценка данных и причин вымираний в позднем триасе» (PDF) . Обзоры наук о Земле . 65 (1–2): 103–139. Бибкод : 2004ESRv...65..103T . дои : 10.1016/S0012-8252(03)00082-5 . Архивировано из оригинала (PDF) 25 октября 2007 года.
- ^ Брусатте С.Л., Бентон М.Дж., Рута М., Ллойд Г.Т. (сентябрь 2008 г.). «Превосходство, конкуренция и оппортунизм в эволюционном развитии динозавров» (PDF) . Наука . 321 (5895): 1485–1488. Бибкод : 2008Sci...321.1485B . дои : 10.1126/science.1161833 . hdl : 20.500.11820/00556baf-6575-44d9-af39-bdd0b072ad2b . ПМИД 18787166 . S2CID 13393888 . Архивировано (PDF) из оригинала 19 июля 2018 года . Проверено 12 октября 2019 г.
- ^ Готье Ж.А. (1986). «Монофилия ящеров и происхождение птиц». В Падиане К. (ред.). Происхождение птиц и эволюция полета. Мемуары Калифорнийской академии наук . Том. 8. Сан-Франциско: Калифорнийская академия наук. стр. 1–55.
- ^ Серено ПК (1991). «Базальные архозавры: филогенетические связи и функциональные последствия». Мемуары Общества палеонтологии позвоночных . 2 :1–53. дои : 10.2307/3889336 . JSTOR 3889336 .
- ^ МакЛауд Н., Роусон П.Ф., Фори П.Л., Баннер Ф.Т., Будагер-Фадель М.К., Баун П.Р. и др. (1997). «Мел-третичный биотический переход». Журнал Геологического общества . 154 (2): 265–292. Бибкод : 1997JGSoc.154..265M . дои : 10.1144/gsjgs.154.2.0265 . S2CID 129654916 .
- ^ Хант Д.М., Ханкинс М.В., Коллин С.П., Маршалл, Нью-Джерси (2014). Эволюция визуальных и невизуальных пигментов . Лондон: Спрингер. п. 73. ИСБН 978-1-4614-4354-4 . OCLC 892735337 .
- ^ Бакалар Н. (2006). «Найден юрский бобр; переписывает историю млекопитающих» . Национальные географические новости . Архивировано из оригинала 3 марта 2006 года . Проверено 28 мая 2016 г.
- ^ Холл М.И., Камилар Дж.М., Кирк Э.К. (декабрь 2012 г.). «Форма глаз и ночное узкое место млекопитающих» . Труды Королевского общества B: Биологические науки . 279 (1749): 4962–4968. дои : 10.1098/rspb.2012.2258 . ПМЦ 3497252 . ПМИД 23097513 .
- ^ Луо ZX (декабрь 2007 г.). «Трансформация и диверсификация ранней эволюции млекопитающих». Природа . 450 (7172): 1011–1019. Бибкод : 2007Natur.450.1011L . дои : 10.1038/nature06277 . ПМИД 18075580 . S2CID 4317817 .
- ^ Пикрелл Дж (2003). «Самое древнее ископаемое сумчатого существа, найденное в Китае» . Национальные географические новости. Архивировано из оригинала 17 декабря 2003 года . Проверено 28 мая 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б Луо ZX, Юань CX, Мэн QJ, Цзи Q (август 2011 г.). «Юрское плацентарное млекопитающее и дивергенция сумчатых и плацентарных». Природа . 476 (7361): 442–5. Бибкод : 2011Natur.476..442L . дои : 10.1038/nature10291 . ПМИД 21866158 . S2CID 205225806 .
- ^ Цзи Q, Луо ZX, Юань CX, Вибл-младший, Чжан Дж. П., Георгий Дж. А. (апрель 2002 г.). «Самое раннее известное эвтерийное млекопитающее». Природа . 416 (6883): 816–822. Бибкод : 2002Natur.416..816J . дои : 10.1038/416816a . ПМИД 11976675 . S2CID 4330626 .
- ↑ Перейти обратно: Перейти обратно: а б Новачек М.Дж., Ружье Г.В., Уибл-младший, Маккенна М.К., Дашевег Д., Горовиц I (октябрь 1997 г.). «Надлобковые кости плацентарных млекопитающих позднего мела Монголии». Природа . 389 (6650): 483–486. Бибкод : 1997Natur.389..483N . дои : 10.1038/39020 . ПМИД 9333234 . S2CID 205026882 .
- ^ Power ML, Schulkin J (2012). «Эволюция живорождения у млекопитающих» . Эволюция плаценты человека . Балтимор: Издательство Университета Джонса Хопкинса. п. 68. ИСБН 978-1-4214-0643-5 .
- ^ Роу Т., Рич Т.Х., Викерс-Рич П., Спрингер М., Вудберн М.О. (январь 2008 г.). «Самый старый утконос и его влияние на время расхождения клад утконоса и ехидны» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (4): 1238–1242. Бибкод : 2008PNAS..105.1238R . дои : 10.1073/pnas.0706385105 . ПМК 2234122 . ПМИД 18216270 .
- ^ Грант Т. (1995). «Репродукция» . Утконос: уникальное млекопитающее . Сидней: Университет Нового Южного Уэльса. п. 55. ИСБН 978-0-86840-143-0 . OCLC 33842474 .
- ^ Goldman AS (июнь 2012 г.). «Эволюция иммунных функций молочной железы и защита ребенка». Медицина грудного вскармливания . 7 (3): 132–142. дои : 10.1089/bfm.2012.0025 . ПМИД 22577734 .
- ↑ Перейти обратно: Перейти обратно: а б Роуз К.Д. (2006). Начало эпохи млекопитающих . Балтимор: Издательство Университета Джонса Хопкинса. стр. 82–83. ISBN 978-0-8018-8472-6 . OCLC 646769601 .
- ^ Бринк А.С. (1955). «Исследование скелета Диадемодона ». Палеонтология Африканская . 3 :3–39.
- ^ Кемп Т.С. (1982). Млекообразные рептилии и происхождение млекопитающих . Лондон: Академическая пресса. п. 363. ИСБН 978-0-12-404120-2 . ОСЛК 8613180 .
- ^ Эстес Р. (1961). «Краниальная анатомия рептилии-цинодонта Thrinaxodon liorhinus ». Бюллетень Музея сравнительной зоологии (1253 г.): 165–180.
- ^ « Тринаксодон: новое млекопитающее» . Ежедневные новости National Geographic. 11 февраля 2009 года. Архивировано из оригинала 14 февраля 2009 года . Проверено 26 августа 2012 г.
- ↑ Перейти обратно: Перейти обратно: а б Байдек П., Кварнстрем М., Овоцкий К., Сулей Т., Сенников А.Г., Голубев В.К., Недзведски Г. (2015). «Микробиота и остатки пищи, включая возможные свидетельства присутствия волос до млекопитающих в копролитах верхней перми из России». Летайя . 49 (4): 455–477. дои : 10.1111/лет.12156 .
- ^ Бота-Бринк Дж., Ангельчик К.Д. (2010). «Объясняют ли необычайно высокие темпы роста пермско-триасовых дицинодонтов (Therapsida, Anomodontia) их успех до и после вымирания в конце пермского периода?» . Зоологический журнал Линнеевского общества . 160 (2): 341–365. дои : 10.1111/j.1096-3642.2009.00601.x .
- ^ Пол Г.С. (1988). Хищные динозавры мира . Нью-Йорк: Саймон и Шустер. п. 464 . ISBN 978-0-671-61946-6 . OCLC 18350868 .
- ^ Уотсон Дж. М., Грейвс Дж. А. (1988). «Монотремные клеточные циклы и эволюция гомеотермии». Австралийский журнал зоологии . 36 (5): 573–584. дои : 10.1071/ZO9880573 .
- ^ Макнаб Б.К. (1980). «Энергетика и пределы умеренного распространения броненосцев». Журнал маммологии . 61 (4): 606–627. дои : 10.2307/1380307 . JSTOR 1380307 .
- ^ Килан-Яворовска З., Хурум Дж.Х. (2006). «Положение конечностей у ранних млекопитающих: растянутое или парасагиттальное» (PDF) . Acta Palaeontologica Polonica . 51 (3): 10237–10239. Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Лиллегравен Дж.А., Килан-Яворовска З., Клеменс В.А. (1979). Мезозойские млекопитающие: первые две трети истории млекопитающих . Издательство Калифорнийского университета. п. 321. ИСБН 978-0-520-03951-3 . OCLC 5910695 .
- ^ Офтедал ОТ (июль 2002 г.). «Молочная железа и ее происхождение в ходе эволюции синапсид». Журнал биологии молочной железы и неоплазии . 7 (3): 225–252. дои : 10.1023/А:1022896515287 . ПМИД 12751889 . S2CID 25806501 .
- ^ Офтедал ОТ (июль 2002 г.). «Происхождение лактации как источника воды для яиц в пергаментной скорлупе». Журнал биологии молочной железы и неоплазии . 7 (3): 253–266. дои : 10.1023/А:1022848632125 . ПМИД 12751890 . S2CID 8319185 .
- ^ «Развитие груди» . Техасская детская больница . Архивировано из оригинала 13 января 2021 года . Проверено 13 января 2021 г.
- ^ Пфафф, Кэтрин; Нагель, Дорис; Ганнелл, Грегг; Вебер, Герхард В.; Жажда, Юрген; Блэк, Майкл; Бастл, Кэтрин (2017). «Палеобиология Hyaenodon exiguus (Hyaenodonta, Mammalia) на основе морфометрического анализа костного лабиринта » Журнал анатомии . 230 (2): 282–289. дои : 10.1111/joa.12545 . ПМК 5244453 . ПМИД 27666133 .
- ↑ Перейти обратно: Перейти обратно: а б Сахни С., Бентон М.Дж., Ферри, Пенсильвания (август 2010 г.). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Смит Ф.А., Бойер А.Г., Браун Дж.Х., Коста Д.П., Даян Т., Эрнест С.К. и др. (ноябрь 2010 г.). «Эволюция максимального размера тела наземных млекопитающих». Наука . 330 (6008): 1216–1219. Бибкод : 2010Sci...330.1216S . CiteSeerX 10.1.1.383.8581 . дои : 10.1126/science.1194830 . ПМИД 21109666 . S2CID 17272200 .
- ^ Симмонс Н.Б., Сеймур К.Л., Хаберсетцер Дж., Ганнелл Г.Ф. (февраль 2008 г.). «Примитивная летучая мышь раннего эоцена из Вайоминга и эволюция полета и эхолокации» . Природа . 451 (7180): 818–821. Бибкод : 2008Natur.451..818S . дои : 10.1038/nature06549 . hdl : 2027.42/62816 . ПМИД 18270539 . S2CID 4356708 . Архивировано из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Бининда-Эмондс О.Р., Кардилло М., Джонс К.Е., Макфи Р.Д., Бек Р.М., Греньер Р. и др. (март 2007 г.). «Замедленный рост современных млекопитающих» (PDF) . Природа . 446 (7135): 507–512. Бибкод : 2007Natur.446..507B . дои : 10.1038/nature05634 . ПМИД 17392779 . S2CID 4314965 . Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б Уибл-младший, Ружье Г.В., Новачек М.Дж., Ашер Р.Дж. (июнь 2007 г.). «Меловые эвтерии и лавразийское происхождение плацентарных млекопитающих вблизи границы K/T». Природа . 447 (7147): 1003–1006. Бибкод : 2007Natur.447.1003W . дои : 10.1038/nature05854 . ПМИД 17581585 . S2CID 4334424 .
- ^ О'Лири М.А., Блох Дж.И., Флинн Дж.Дж., Годен Т.Дж., Джалломбардо А., Джаннини Н.П. и др. (февраль 2013 г.). «Предок плацентарных млекопитающих и пост-K-Pg-излучение плаценты» . Наука . 339 (6120): 662–667. Бибкод : 2013Sci...339..662O . дои : 10.1126/science.1229237 . hdl : 11336/7302 . ПМИД 23393258 . S2CID 206544776 . Архивировано из оригинала 10 ноября 2021 года . Проверено 30 июня 2022 г.
- ^ Холлидей Т.Дж., Апчерч П., Госвами А. (февраль 2017 г.). «Разрешение взаимоотношений плацентарных млекопитающих палеоцена» . Биологические обзоры Кембриджского философского общества . 92 (1): 521–550. дои : 10.1111/brv.12242 . ПМК 6849585 . ПМИД 28075073 .
- ^ Холлидей Т.Дж., Апчерч П., Госвами А. (июнь 2016 г.). «Эвтерийцы испытали повышенные темпы эволюции сразу после массового вымирания в мел-палеогеновом периоде» . Слушания. Биологические науки . 283 (1833): 20153026. doi : 10.1098/rspb.2015.3026 . ПМЦ 4936024 . ПМИД 27358361 .
- ↑ Перейти обратно: Перейти обратно: а б с Ни Х, Гебо Д.Л., Дагосто М., Мэн Дж., Таффоро П., Флинн Дж.Дж., Бирд К.С. (июнь 2013 г.). «Самый старый известный скелет примата и ранняя эволюция гаплорина». Природа . 498 (7452): 60–64. Бибкод : 2013Natur.498...60N . дои : 10.1038/nature12200 . ПМИД 23739424 . S2CID 4321956 .
- ^ Ромер С.А., Парсонс Т.С. (1977). Тело позвоночного . Филадельфия: Холт-Сондерс Интернэшнл. стр. 129–145. ISBN 978-0-03-910284-5 . OCLC 60007175 .
- ^ Пурвес В.К., Садава Д.Е., Орианс Г.Х., Хелле Х.К. (2001). Жизнь: наука биологии (6-е изд.). Нью-Йорк: Sinauer Associates, Inc., с. 593. ИСБН 978-0-7167-3873-2 . OCLC 874883911 .
- ^ Антвал Н., Джоши Л., Такер А.С. (январь 2013 г.). «Эволюция среднего уха и челюсти млекопитающих: адаптации и новые структуры» . Журнал анатомии . 222 (1): 147–160. дои : 10.1111/j.1469-7580.2012.01526.x . ПМЦ 3552421 . ПМИД 22686855 .
- ^ ван Нивельт А.Ф., Смит К.К. (2005). «Заменять или не заменять: значение снижения функциональной замены зубов у сумчатых и плацентарных млекопитающих». Палеобиология . 31 (2): 324–346. doi : 10.1666/0094-8373(2005)031[0324:trontr]2.0.co;2 . S2CID 37750062 .
- ^ Либертини Дж., Феррара Н. (апрель 2016 г.). «Старение многолетних клеток и частей органов согласно парадигме программируемого старения» . Возраст . 38 (2): 35. дои : 10.1007/s11357-016-9895-0 . ПМЦ 5005898 . ПМИД 26957493 .
- ^ Мао Ф., Ван Ю, Мэн Дж (2015). «Систематическое исследование микроструктур зубной эмали Lambdopsalis Bulla (Multituberculate, Mammalia) – значение для мультитуберкулезной биологии и филогении» . ПЛОС ОДИН . 10 (5): e0128243. Бибкод : 2015PLoSO..1028243M . дои : 10.1371/journal.pone.0128243 . ПМЦ 4447277 . ПМИД 26020958 .
- ^ Осборн Х.Ф. (1900). «Происхождение млекопитающих, III. Затылочные мыщелки рептильного трехраздельного типа» . Американский натуралист . 34 (408): 943–947. дои : 10.1086/277821 . JSTOR 2453526 .
- ^ Кромптон А.В., Дженкинс-младший Ф.А. (1973). «Млекопитающие от рептилий: обзор происхождения млекопитающих». Ежегодный обзор наук о Земле и планетах . 1 : 131–155. Бибкод : 1973AREPS...1..131C . doi : 10.1146/annurev.ea.01.050173.001023 .
- ↑ Перейти обратно: Перейти обратно: а б Power ML, Schulkin J (2013). Эволюция плаценты человека . Балтимор: Издательство Университета Джонса Хопкинса. стр. 1890–1891. ISBN 978-1-4214-0643-5 . OCLC 940749490 .
- ^ Линденфорс П., Гиттлман Дж.Л., Джонс К.Е. (2007). «Половой размерный диморфизм млекопитающих». Пол, размер и гендерные роли: эволюционные исследования полового диморфизма размеров . Оксфорд: Издательство Оксфордского университета. стр. 16–26. ISBN 978-0-19-920878-4 . Архивировано из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Диерауф Л.А., Гулланд FM (2001). Справочник CRC по медицине морских млекопитающих: здоровье, болезни и реабилитация (2-е изд.). Бока-Ратон: CRC Press. п. 154. ИСБН 978-1-4200-4163-7 . OCLC 166505919 .
- ^ Луи Дж. Х., Хансен Д. В., Кригштейн А. Р. (июль 2011 г.). «Развитие и эволюция неокортекса человека» . Клетка . 146 (1): 18–36. дои : 10.1016/j.cell.2011.06.030 . ПМК 3610574 . ПМИД 21729779 .
- ^ Киллер CE (июнь 1933 г.). «Отсутствие мозолистого тела как менделизирующего признака у домовой мыши» . Труды Национальной академии наук Соединенных Штатов Америки . 19 (6): 609–611. Бибкод : 1933PNAS...19..609K . дои : 10.1073/pnas.19.6.609 . JSTOR 86284 . ПМК 1086100 . ПМИД 16587795 .
- ^ Стэндринг С., Борли Н.Р. (2008). Анатомия Грея: анатомические основы клинической практики (40-е изд.). Лондон: Черчилль Ливингстон. стр. 960–962. ISBN 978-0-8089-2371-8 . OCLC 213447727 .
- ^ Беттс Дж. Ф., Дезе П., Джонсон Э., Джонсон Дж. Э., Корол О., Круз Д. и др. (2013). Анатомия и физиология . Хьюстон: Издательство Университета Райса. стр. 787–846. ISBN 978-1-938168-13-0 . OCLC 898069394 . Архивировано из оригинала 23 февраля 2022 года . Проверено 25 января 2024 г.
- ^ Левицкий М.Г. (2013). «Механика дыхания». Легочная физиология (8-е изд.). Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-179313-1 . OCLC 940633137 .
- ↑ Перейти обратно: Перейти обратно: а б Умеш КБ (2011). «Легочная анатомия и физиология» . Справочник по механической вентиляции . Нью-Дели: Медицинское издательство Jaypee Brothers. п. 12. ISBN 978-93-80704-74-6 . OCLC 945076700 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я Фельдхамер Г.А., Дрикамер Л.С., Весси С.Х., Мерритт Дж.Ф., Краевски С. (2007). Маммология: адаптация, разнообразие, экология (3-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-8695-9 . OCLC 124031907 .
- ^ Тинкер СВ (1988). Киты мира . Архив Брилла. п. 51. ИСБН 978-0-935848-47-2 .
- ^ Ромер А.С. (1959). История позвоночных (4-е изд.). Чикаго: Издательство Чикагского университета. ISBN 978-0-226-72490-4 .
- ^ де Мюзон К., Ланге-Бадре Б. (1997). «Плотоядные зубные адаптации трибосфенических млекопитающих и филогенетическая реконструкция». Летайя . 30 (4): 353–366. Бибкод : 1997Лета..30..353Д . дои : 10.1111/j.1502-3931.1997.tb00481.x .
- ↑ Перейти обратно: Перейти обратно: а б Лангер П. (июль 1984 г.). «Сравнительная анатомия желудка травоядных млекопитающих». Ежеквартальный журнал экспериментальной физиологии . 69 (3): 615–625. doi : 10.1113/expphysicalol.1984.sp002848 . ПМИД 6473699 . S2CID 30816018 .
- ^ Воан Т.А., Райан Дж.М., Чаплевски, Нью-Джерси (2011). «Периссодактилия» . Маммология (5-е изд.). Джонс и Бартлетт. п. 322. ИСБН 978-0-7637-6299-5 . OCLC 437300511 .
- ^ Флауэр WH, Лидеккер Р. (1946). Введение в изучение ныне живущих и вымерших млекопитающих . Лондон: Адам и Чарльз Блэк. п. 496. ИСБН 978-1-110-76857-8 .
- ^ Шрикумар С (2010). Базовая физиология . PHI Learning Pvt. ООО, стр. 180–181. ISBN 978-81-203-4107-4 .
- ^ Чейфец А.С. (2010). Оксфордский американский справочник по гастроэнтерологии и гепатологии . Оксфорд: Издательство Оксфордского университета, США. п. 165. ИСБН 978-0-19-983012-1 .
- ^ Кунц Э (2008). Гепатология: Учебник и атлас . Германия: Шпрингер. п. 38. ISBN 978-3-540-76838-8 .
- ^ Ортис Р.М. (июнь 2001 г.). «Осморегуляция у морских млекопитающих» . Журнал экспериментальной биологии . 204 (Часть 11): 1831–1844. дои : 10.1242/jeb.204.11.1831 . ПМИД 11441026 . Архивировано из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б с д Роман А.С., Парсонс Т.С. (1977). Тело позвоночного . Филадельфия: Холт-Сондерс Интернэшнл. стр. 396–399. ISBN 978-0-03-910284-5 .
- ^ Линзи, Дональд В. (2020). Биология позвоночных: систематика, таксономия, естественная история и охрана . Издательство Университета Джонса Хопкинса. п. 306. ИСБН 978-1-42143-733-0 . Архивировано из оригинала 22 января 2024 года . Проверено 22 января 2024 г.
- ^ «Биологические обзоры – Кембриджские журналы» . Архивировано из оригинала 22 ноября 2015 года . Проверено 21 января 2017 г.
- ^ Докинз Р., Вонг Ю. (2016). Рассказ предка: Паломничество на заре эволюции (2-е изд.). Бостон: Mariner Books. п. 281. ИСБН 978-0-544-85993-7 .
- ↑ Перейти обратно: Перейти обратно: а б с Фитч WT (2006). «Производство вокализаций у млекопитающих» (PDF) . В Брауне К. (ред.). Энциклопедия языка и лингвистики . Оксфорд: Эльзевир. стр. 115–121. Архивировано из оригинала 1 июня 2024 года . Проверено 25 января 2024 г.
- ^ Ланжевен П., Барклай Р.М. (1990). « Гипсигнатус чудовищный » . Виды млекопитающих (357): 1–4. дои : 10.2307/3504110 . JSTOR 3504110 .
- ^ Вайсенгрубер Г.Е., Форстенпойнтнер Г., Петерс Г., Кюббер-Хейсс А., Fitch WT (сентябрь 2002 г.). «Подъязычный аппарат и глотка у льва (Panthera leo), ягуара (Panthera onca), тигра (Panthera tigris), гепарда (Acinonyxjubatus) и домашней кошки (Felis silvestris f. catus)» . Журнал анатомии . 201 (3): 195–209. дои : 10.1046/j.1469-7580.2002.00088.x . ПМК 1570911 . ПМИД 12363272 .
- ^ Штегер А.С., Хайльманн Г., Цеппельзауер М., Гансвиндт А., Хенсман С., Чарльтон Б.Д. (2012). «Визуализация звукового излучения слонов: свидетельства двух типов грохота» . ПЛОС ОДИН . 7 (11): е48907. Бибкод : 2012PLoSO...748907S . дои : 10.1371/journal.pone.0048907 . ПМЦ 3498347 . ПМИД 23155427 .
- ^ Кларк К.В. (2004). «Инфразвуковые звуки усатых китов: естественная изменчивость и функция». Журнал Акустического общества Америки . 115 (5): 2554. Бибкод : 2004ASAJ..115.2554C . дои : 10.1121/1.4783845 .
- ↑ Перейти обратно: Перейти обратно: а б Доусон Т.Дж., Вебстер К.Н., Мэлони С.К. (февраль 2014 г.). «Мех млекопитающих в открытой среде; обязательно ли конфликтуют крипсис и тепловые потребности? Сравнение белого медведя и сумчатой коалы». Журнал сравнительной физиологии Б. 184 (2): 273–284. дои : 10.1007/s00360-013-0794-8 . ПМИД 24366474 . S2CID 9481486 .
- ^ Сломински А., Тобин Д.Д., Шибахара С., Вортсман Дж. (октябрь 2004 г.). «Меланиновая пигментация в коже млекопитающих и ее гормональная регуляция». Физиологические обзоры . 84 (4): 1155–1228. doi : 10.1152/physrev.00044.2003 . ПМИД 15383650 . S2CID 21168932 .
- ^ Хилтон-младший Б (1996). «Дикая природа Южной Каролины» . Цвета животных . 43 (4). Хилтон Понд Центр: 10–15. Архивировано из оригинала 25 января 2024 года . Проверено 26 ноября 2011 г.
- ↑ Перейти обратно: Перейти обратно: а б Прум Р.О., Торрес Р.Х. (май 2004 г.). «Структурная окраска кожи млекопитающих: конвергентная эволюция когерентно рассеивающихся массивов дермального коллагена» (PDF) . Журнал экспериментальной биологии . 207 (Часть 12): 2157–2172. дои : 10.1242/jeb.00989 . hdl : 1808/1599 . ПМИД 15143148 . S2CID 8268610 . Архивировано (PDF) из оригинала 5 июня 2024 года . Проверено 25 января 2024 г.
- ^ Суутари М., Маянева М., Фьюер Д.П., Войрин Б., Айелло А., Фридл Т. и др. (март 2010 г.). «Молекулярные доказательства разнообразного сообщества зеленых водорослей, растущих в волосах ленивцев, и специфической связи с Trichophilus welckeri (Chlorophyta, Ulvophyceae)» . Эволюционная биология BMC . 10 (86): 86. Бибкод : 2010BMCEE..10...86S . дои : 10.1186/1471-2148-10-86 . ПМЦ 2858742 . ПМИД 20353556 .
- ^ Каро Т (2005). «Адаптационное значение окраски млекопитающих» . Бионаука . 55 (2): 125–136. doi : 10.1641/0006-3568(2005)055[0125:tasoci]2.0.co;2 .
- ^ Миллс Л.С., Зимова М., Ойлер Дж., Бегущий С., Абацоглу Дж.Т., Лукач П.М. (апрель 2013 г.). «Несоответствие камуфляжа сезонной окраски шерсти из-за уменьшения продолжительности снега» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (18): 7360–7365. Бибкод : 2013PNAS..110.7360M . дои : 10.1073/pnas.1222724110 . ПМЦ 3645584 . ПМИД 23589881 .
- ^ Каро Т. (февраль 2009 г.). «Контрастная окраска у наземных млекопитающих» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 364 (1516): 537–548. дои : 10.1098/rstb.2008.0221 . ПМК 2674080 . ПМИД 18990666 .
- ^ Плавкан Дж.М. (2001). «Половой диморфизм в эволюции приматов» . Американский журнал физической антропологии . Приложение 33 (33): 25–53. дои : 10.1002/ajpa.10011 . ПМИД 11786990 . S2CID 31722173 .
- ^ Брэдли Б.Дж., Джеральд М.С., Виддиг А., Манди Н.И. (2012). «Изменение цвета шерсти и экспрессия генов пигментации у макак-резус ( Macaca Mulatta )» (PDF) . Журнал эволюции млекопитающих . 20 (3): 263–270. дои : 10.1007/s10914-012-9212-3 . S2CID 13916535 . Архивировано из оригинала (PDF) 24 сентября 2015 года.
- ^ Каро Т. , Иззо А., Райнер Р.К., Уокер Х., Станкович Т. (апрель 2014 г.). «Функция полосок зебры» . Природные коммуникации . 5 : 3535. Бибкод : 2014NatCo...5.3535C . дои : 10.1038/ncomms4535 . ПМИД 24691390 . S2CID 9849814 .
- ^ Кобаяши К., Китано Т., Ивао Ю., Кондо М. (2018). Репродуктивные стратегии и стратегии развития: непрерывность жизни . Спрингер. п. 290. ИСБН 978-4-431-56609-0 .
- ^ Либби Генриетта Хайман (15 сентября 1992 г.). Сравнительная анатомия позвоночных Хаймана . Издательство Чикагского университета. стр. 583–. ISBN 978-0-226-87013-7 .
- ^ Леонард, Джанет; Кордова-Агилар, Алекс (19 июля 2010 г.). Эволюция первичных половых признаков у животных . Издательство Оксфордского университета, США. ISBN 978-0-19-532555-3 .
- ^ Ломбарди Дж. (1998). Сравнительное размножение позвоночных . Springer Science & Business Media. ISBN 978-0-7923-8336-9 .
- ^ Тиндейл-Биско Х., Ренфри М. (1987). Репродуктивная физиология сумчатых . Издательство Кембриджского университета. ISBN 978-0-521-33792-2 .
- ^ Джонстон С.Д., Смит Б., Пайн М., Стензель Д., Холт В.В. (2007). «Односторонняя эякуляция пучков сперматозоидов ехидны» (PDF) . Американский натуралист . 170 (6): E162–E164. дои : 10.1086/522847 . ПМИД 18171162 . S2CID 40632746 .
- ^ Бача-младший, Уильям Дж.; Бача, Линда М. (2012). Цветной атлас ветеринарной гистологии . Уайли. п. 308. ИСБН 978-1-11824-364-0 . Проверено 28 ноября 2023 г.
- ^ Кук, Фред; Брюс, Дженни (2004). Энциклопедия животных: полное наглядное руководство . Издательство Калифорнийского университета. п. 79. ИСБН 978-0-52024-406-1 . Проверено 28 ноября 2023 г.
- ^ Максвелл К.Е. (2013). Секс-императив: эволюционная история сексуального выживания . Спрингер. стр. 112–113. ISBN 978-1-4899-5988-1 .
- ^ Воган Т.А., Райан Дж.П., Чаплевски, Нью-Джерси (2011). Маммология . Издательство Джонс и Бартлетт. п. 387. ИСБН 978-0-03-025034-7 .
- ↑ Перейти обратно: Перейти обратно: а б Хоффман Э.А., Роу ТБ (сентябрь 2018 г.). «Перинаты стволовых млекопитающих юрского периода и происхождение размножения и роста млекопитающих». Природа . 561 (7721): 104–108. Бибкод : 2018Natur.561..104H . дои : 10.1038/s41586-018-0441-3 . ПМИД 30158701 . S2CID 205570021 .
- ^ Уоллис М.К., Уотерс П.Д., Делбридж М.Л., Кирби П.Дж., Паск А.Дж., Грюцнер Ф. и др. (2007). «Определение пола у утконоса и ехидны: аутосомное расположение SOX3 подтверждает отсутствие SRY у однопроходных». Хромосомные исследования . 15 (8): 949–959. дои : 10.1007/s10577-007-1185-3 . ПМИД 18185981 . S2CID 812974 .
- ^ Маршалл Грейвс, Дж. А. (2008). «Странные геномы животных и эволюция пола и половых хромосом позвоночных» (PDF) . Ежегодный обзор генетики . 42 : 565–586. дои : 10.1146/annurev.genet.42.110807.091714 . ПМИД 18983263 . Архивировано из оригинала (PDF) 4 сентября 2012 года . Проверено 25 января 2024 г.
- ^ Салли М (2005). «Поведение и образ жизни млекопитающих» . Млекопитающие . Чикаго: Рейнтри. п. 6. ISBN 978-1-4109-1050-9 . OCLC 53476660 .
- ^ Верма П.С., Панди Б.П. (2013). Книга ISC по биологии ISC для XI класса . Нью-Дели: С. Чанд и компания. п. 288. ИСБН 978-81-219-2557-0 .
- ^ Офтедал ОТ (июль 2002 г.). «Молочная железа и ее происхождение в ходе эволюции синапсид». Журнал биологии молочной железы и неоплазии . 7 (3): 225–252. дои : 10.1023/а:1022896515287 . ПМИД 12751889 . S2CID 25806501 .
- ^ Крокенбергер А (2006). «Лактация». Дикман Ч.Р., Армати П.Дж., Хьюм И.Д. (ред.). Сумчатые . Издательство Кембриджского университета. п. 109. ИСБН 978-1-139-45742-2 .
- ^ Щулкин Дж. Power ML (2016). Молоко: биология лактации . Издательство Университета Джонса Хопкинса. п. 66. ИСБН 978-1-4214-2042-4 .
- ^ Томпсон К.В., Бейкер А.Дж., Бейкер А.М. (2010). «Отцовская забота и развитие поведения млекопитающих в неволе». Клейман Д.Г., Томпсон К.В., Баер К.К. (ред.). Дикие млекопитающие в неволе. Принципы и методы управления зоопарком (2-е изд.). Издательство Чикагского университета. п. 374. ИСБН 978-0-226-44011-8 .
- ^ Кэмпбелл Н.А., Рис Дж.Б. (2002). Биология (6-е изд.). Бенджамин Каммингс. п. 845 . ISBN 978-0-8053-6624-2 . OCLC 47521441 .
- ^ Буффенштейн Р., Яхав С. (1991). «Является ли голый землекоп Hererocephalus glaber эндотермическим, но пойкилотермным млекопитающим?». Журнал термической биологии . 16 (4): 227–232. Бибкод : 1991JTBio..16..227B . дои : 10.1016/0306-4565(91)90030-6 .
- ^ Шмидт-Нильсен К., Дюк Дж.Б. (1997). «Температурные эффекты» . Физиология животных: адаптация и окружающая среда (5-е изд.). Кембридж: Издательство Кембриджского университета. п. 218. ИСБН 978-0-521-57098-5 . OCLC 35744403 .
- ↑ Перейти обратно: Перейти обратно: а б Лоренцини А., Джонсон Ф.Б., Оливер А., Трезини М., Смит Дж.С., Хдейб М. и др. (2009). «Значительная корреляция продолжительности жизни видов с распознаванием двухцепочечных разрывов ДНК, но не с длиной теломер» . Механизмы старения и развития . 130 (11–12): 784–792. дои : 10.1016/j.mad.2009.10.004 . ПМК 2799038 . ПМИД 19896964 .
- ^ Харт Р.В., Сетлоу Р.Б. (июнь 1974 г.). «Корреляция между эксцизионным восстановлением дезоксирибонуклеиновой кислоты и продолжительностью жизни у ряда видов млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (6): 2169–2173. Бибкод : 1974PNAS...71.2169H . дои : 10.1073/pnas.71.6.2169 . ПМЦ 388412 . ПМИД 4526202 .
- ^ Ма С., Упнея А., Галецкий А., Цай Ю.М., Бурант К.Ф., Раскинд С. и др. (ноябрь 2016 г.). «Профилирование млекопитающих на основе клеточных культур показывает, что восстановление ДНК и метаболизм являются определяющими факторами долголетия вида» . электронная жизнь . 5 . doi : 10.7554/eLife.19130 . ПМК 5148604 . ПМИД 27874830 .
- ^ Грубе К., Бюркле А. (декабрь 1992 г.). «Активность поли(АДФ-рибозы)-полимеразы в мононуклеарных лейкоцитах 13 видов млекопитающих коррелирует с видоспецифичной продолжительностью жизни» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (24): 11759–11763. Бибкод : 1992PNAS...8911759G . дои : 10.1073/pnas.89.24.11759 . ПМК 50636 . ПМИД 1465394 .
- ^ Фрэнсис А.А., Ли В.Х., Риган Дж.Д. (июнь 1981 г.). «Связь эксцизионной репарации ДНК поражений, вызванных ультрафиолетом, с максимальной продолжительностью жизни млекопитающих». Механизмы старения и развития . 16 (2): 181–189. дои : 10.1016/0047-6374(81)90094-4 . ПМИД 7266079 . S2CID 19830165 .
- ^ Третон Дж. А., Куртуа Ю. (март 1982 г.). «Корреляция между эксцизионной репарацией ДНК и продолжительностью жизни млекопитающих в эпителиальных клетках хрусталика». Международные отчеты по клеточной биологии . 6 (3): 253–260. doi : 10.1016/0309-1651(82)90077-7 (неактивен 31 января 2024 г.). ПМИД 7060140 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Маслански CJ, Williams GM (февраль 1985 г.). «Синтез репарации ДНК, индуцированный ультрафиолетовым светом, в гепатоцитах видов с разной продолжительностью жизни». Механизмы старения и развития . 29 (2): 191–203. дои : 10.1016/0047-6374(85)90018-1 . ПМИД 3974310 . S2CID 23988416 .
- ^ Уокер В.Ф., Хомбергер Д.Г. (1998). Анатомия и вскрытие плода свиньи (5-е изд.). Нью-Йорк: WH Freeman and Company. п. 3. ISBN 978-0-7167-2637-1 . OCLC 40576267 .
- ^ Орр CM (ноябрь 2005 г.). «Муравьед, ходящий на костяшках пальцев: тест конвергенции адаптации предполагаемых особенностей ходьбы на костяшках пальцев африканских гоминид». Американский журнал физической антропологии . 128 (3): 639–658. дои : 10.1002/ajpa.20192 . ПМИД 15861420 .
- ^ Фиш Ф.Э., Фраппелл П.Б., Бодинетт Р.В., Макфарлейн П.М. (февраль 2001 г.). «Энергетика наземного передвижения утконоса Ornithorhynchus anatinus» (PDF) . Журнал экспериментальной биологии . 204 (Часть 4): 797–803. дои : 10.1242/jeb.204.4.797 . hdl : 2440/12192 . ПМИД 11171362 . Архивировано (PDF) из оригинала 14 марта 2024 года . Проверено 25 января 2024 г.
- ^ Дхингра П. (2004). «Сравнительное прямохождение – как остальные представители животного мира ходят на двух ногах» . Антропологическая наука . 131 (231). Архивировано из оригинала 21 апреля 2021 года . Проверено 11 марта 2017 г.
- ^ Александр Р.М. (май 2004 г.). «Двуногие животные и их отличия от человека» . Журнал анатомии . 204 (5): 321–330. дои : 10.1111/j.0021-8782.2004.00289.x . ПМЦ 1571302 . ПМИД 15198697 .
- ↑ Перейти обратно: Перейти обратно: а б Дагг А.И. (1973). «Походка млекопитающих». Обзор млекопитающих . 3 (4): 135–154. дои : 10.1111/j.1365-2907.1973.tb00179.x .
- ^ Робертс ТД (1995). Понимание баланса: механика позы и передвижения . Сан-Диего: Нельсон Торнс. п. 211. ИСБН 978-1-56593-416-0 . OCLC 33167785 .
- ↑ Перейти обратно: Перейти обратно: а б с Картмилл М (1985). «Восхождение». Хильдебранд М., Брамбл Д.М., Лием К.Ф., Wake DB (ред.). Функциональная морфология позвоночных . Кембридж: Белкнап Пресс. стр. 73–88. ISBN 978-0-674-32775-7 . ОСЛК 11114191 .
- ^ Верн К. (2001). «Скольжение северной летяги ( Glaucomys sabrinus ) в зрелом смешанном лесу восточной Канады» . Журнал маммологии . 82 (4): 1026–1033. doi : 10.1644/1545-1542(2001)082<1026:GPOTNF>2.0.CO;2 . S2CID 78090049 .
- ^ Барба Л.А. (октябрь 2011 г.). «Летучие мыши – единственные летающие млекопитающие» . Биовоздушное передвижение . Архивировано из оригинала 14 мая 2016 года . Проверено 20 мая 2016 г. .
- ^ «Летучие мыши в полете демонстрируют неожиданную аэродинамику» . ScienceDaily . 2007. Архивировано из оригинала 19 декабря 2019 года . Проверено 12 июля 2016 г.
- ^ Хеденстрем А., Йоханссон LC (март 2015 г.). «Полет летучей мыши: аэродинамика, кинематика и морфология полета» (PDF) . Журнал экспериментальной биологии . 218 (Часть 5): 653–663. дои : 10.1242/jeb.031203 . ПМИД 25740899 . S2CID 21295393 . Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ «Летучие мыши экономят энергию, сжимая крылья при взмахе вверх» . ScienceDaily . 2012. Архивировано из оригинала 31 мая 2021 года . Проверено 12 июля 2016 г.
- ^ Карен Т. (2008). Встреча с летучими мышами: экобаты, вампиры и кинозвезды . Альбукерке: Издательство Университета Нью-Мексико. п. 14. ISBN 978-0-8263-4403-8 . OCLC 191258477 .
- ^ Стербинг-Д'Анджело С., Чадха М., Чиу С., Фальк Б., Сиань В., Барсело Дж. и др. (июль 2011 г.). «Датчики крыльев летучей мыши поддерживают управление полетом» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (27): 11291–11296. Бибкод : 2011PNAS..10811291S . дои : 10.1073/pnas.1018740108 . ПМК 3131348 . ПМИД 21690408 .
- ^ Дамиани, Р., 2003, Самые ранние свидетельства рытья нор цинодонтов, Издательство Королевского общества, том 270, выпуск 1525.
- ^ Шимер Х.В. (1903). «Адаптации к водным, древесным, ископаемым и бегущим привычкам млекопитающих. III. Ископаемые адаптации» . Американский натуралист . 37 (444): 819–825. дои : 10.1086/278368 . JSTOR 2455381 . S2CID 83519668 . Архивировано из оригинала 9 апреля 2023 года . Проверено 23 августа 2020 г. .
- ^ Стэнхоуп М.Дж., Уодделл В.Г., Мэдсен О., де Йонг В., Хеджес С.Б., Клевен Г.К. и др. (август 1998 г.). «Молекулярные доказательства множественного происхождения насекомоядных и нового порядка эндемичных африканских насекомоядных млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (17): 9967–9972. Бибкод : 1998PNAS...95.9967S . дои : 10.1073/pnas.95.17.9967 . ПМК 21445 . ПМИД 9707584 .
- ^ Перри Д.А. (1949). «Анатомические основы плавания китов». Журнал зоологии . 119 (1): 49–60. дои : 10.1111/j.1096-3642.1949.tb00866.x .
- ^ Фиш Ф.Е., Хуэй, Калифорния (1991). «Плавание дельфинов – обзор» (PDF) . Обзор млекопитающих . 21 (4): 181–195. дои : 10.1111/j.1365-2907.1991.tb00292.x . Архивировано из оригинала (PDF) 29 августа 2006 года.
- ^ Марш Х (1989). «Глава 57: Дюгонгиды» (PDF) . Фауна Австралии . Том. 1. Канберра: Публикации правительства Австралии. ISBN 978-0-644-06056-1 . ОСЛК 27492815 . Архивировано из оригинала (PDF) 11 мая 2013 года.
- ↑ Перейти обратно: Перейти обратно: а б Берта А (2012). «Разнообразие ластоногих: эволюция и адаптация». Возвращение в море: жизнь и эпоха эволюции морских млекопитающих . Издательство Калифорнийского университета. стр. 62–64. ISBN 978-0-520-27057-2 .
- ↑ Перейти обратно: Перейти обратно: а б Фиш Ф.Е., Херли Дж., Коста Д.П. (февраль 2003 г.). «Маневренность морского льва Zalophus Californianus: поворотные качества нестабильной конструкции тела» . Журнал экспериментальной биологии . 206 (Часть 4): 667–674. дои : 10.1242/jeb.00144 . ПМИД 12517984 .
- ↑ Перейти обратно: Перейти обратно: а б Ридман М (1990). Ластоногие: тюлени, морские львы и моржи . Издательство Калифорнийского университета. ISBN 978-0-520-06497-3 . OCLC 19511610 .
- ^ Фиш ФЭ (1996). «Переход от движения на основе сопротивления к подъему в плавании млекопитающих» . Интегративная и сравнительная биология . 36 (6): 628–641. дои : 10.1093/icb/36.6.628 .
- ^ Рыба ФЭ (2000). «Биомеханика и энергетика водных и околоводных млекопитающих: от утконоса до кита» (PDF) . Физиологическая и биохимическая зоология . 73 (6): 683–698. CiteSeerX 10.1.1.734.1217 . дои : 10.1086/318108 . ПМИД 11121343 . S2CID 49732160 . Архивировано из оригинала (PDF) 4 августа 2016 года.
- ^ Элтрингем СК (1999). «Анатомия и физиология». Бегемоты . Лондон: T & AD Poyser Ltd., с. 8. ISBN 978-0-85661-131-5 . OCLC 42274422 .
- ^ «Гиппопотам Бегемот-амфибия » . Нэшнл Географик . Архивировано из оригинала 25 ноября 2014 года . Проверено 30 апреля 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б Сейфарт Р.М., Чейни Д.Л., Марлер П. (1980). «Тревожные звонки верветок: семантическая коммуникация у свободно живущего примата» . Поведение животных . 28 (4): 1070–1094. дои : 10.1016/S0003-3472(80)80097-2 . S2CID 53165940 . Архивировано из оригинала 12 сентября 2019 года . Проверено 22 сентября 2018 г.
- ^ Зубербюлер К (2001). «Тревожные сигналы, характерные для хищников, у обезьян Кэмпбелла, Cercopithecus Campbelli ». Поведенческая экология и социобиология . 50 (5): 414–442. дои : 10.1007/s002650100383 . JSTOR 4601985 . S2CID 21374702 .
- ^ Слаббекоорн Х., Смит Т.Б. (апрель 2002 г.). «Пение птиц, экология и видообразование» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 357 (1420): 493–503. дои : 10.1098/rstb.2001.1056 . ПМК 1692962 . ПМИД 12028787 .
- ^ Баннистер Дж.Л. (2008). «Усатые киты (Mysticetes)» . В Perrin WF, Würsig B, Thewissen JG (ред.). Энциклопедия морских млекопитающих (2-е изд.). Академическая пресса. стр. 80–89. ISBN 978-0-12-373553-9 .
- ^ Скотт Н. (2002). «Существа культуры? Обоснование культурных систем у китов и дельфинов» . Бионаука . 52 (1): 9–14. doi : 10.1641/0006-3568(2002)052[0009:COCMTC]2.0.CO;2 . S2CID 86121405 .
- ^ Бофман Дж.В. (февраль 1998 г.). «Вокальное обучение больших остроносых летучих мышей» . Слушания. Биологические науки . 265 (1392): 227–233. дои : 10.1098/rspb.1998.0286 . ПМК 1688873 . ПМИД 9493408 .
- ^ «Язык луговых собачек расшифрован учеными» . Новости Си-Би-Си. 21 июня 2013 года. Архивировано из оригинала 22 мая 2015 года . Проверено 20 мая 2015 г.
- ^ Мэйелл Х. (3 марта 2004 г.). «Слоны звонят по междугородной связи в нерабочее время» . Нэшнл Географик . Архивировано из оригинала 5 марта 2004 года . Проверено 15 ноября 2016 г.
- ^ Смит Дж. М. , Харпер Д. (2003). Сигналы животных . Оксфордская серия по экологии и эволюции. Издательство Оксфордского университета. стр. 61–63. ISBN 978-0-19-852684-1 . OCLC 54460090 .
- ^ Компакт-диск ФитцГиббона, Фэншоу Дж.Х. (1988). «Стоттинг в газелях Томсона: честный сигнал о состоянии» (PDF) . Поведенческая экология и социобиология . 23 (2): 69–74. дои : 10.1007/bf00299889 . S2CID 2809268 . Архивировано из оригинала (PDF) 25 февраля 2014 года.
- ^ Бильдштейн К.Л. (май 1983 г.). «Почему белохвостые олени машут хвостами». Американский натуралист . 121 (5): 709–715. дои : 10.1086/284096 . JSTOR 2460873 . S2CID 83504795 .
- ^ Гослинг Л.М. (январь 1982 г.). «Переоценка функции ароматической маркировки территорий» . Zeitschrift für Tierpsychologie . 60 (2): 89–118. дои : 10.1111/j.1439-0310.1982.tb00492.x . Архивировано (PDF) из оригинала 27 марта 2018 г. Проверено 12 октября 2019 г.
- ^ Зала С.М., Поттс В.К., Пенн Диджей (март 2004 г.). «Дисплеи с ароматической маркировкой дают честные сигналы о здоровье и инфекции». Поведенческая экология . 15 (2): 338–344. дои : 10.1093/beheco/arh022 . hdl : 10.1093/beheco/arh022 .
- ^ Джонсон Р.П. (август 1973 г.). «Ароматная маркировка млекопитающих». Поведение животных . 21 (3): 521–535. дои : 10.1016/S0003-3472(73)80012-0 .
- ^ Шевилл В.Е., Макбрайд А.Ф. (1956). «Доказательства эхолокации китообразных». Глубоководные исследования . 3 (2): 153–154. Бибкод : 1956DSR.....3..153S . дои : 10.1016/0146-6313(56)90096-х .
- ^ Уилсон В., Мосс С. (2004). Томас Дж. (ред.). Эхолокация у летучих мышей и дельфинов . Издательство Чикагского университета. п. 22. ISBN 978-0-226-79599-7 . OCLC 50143737 .
- ^ Au WW (1993). Сонар дельфинов . Спрингер-Верлаг. ISBN 978-3-540-97835-0 . OCLC 26158593 .
- ^ Сандерс Дж.Г., Бейхман А.С., Роман Дж., Скотт Дж.Дж., Эмерсон Д., Маккарти Дж.Дж., Гиргис П.Р. (сентябрь 2015 г.). «У усатых китов имеется уникальный кишечный микробиом, сходный как с плотоядными, так и с травоядными» . Природные коммуникации . 6 : 8285. Бибкод : 2015NatCo...6.8285S . дои : 10.1038/ncomms9285 . ПМЦ 4595633 . ПМИД 26393325 .
- ^ Представитель JR (1996). «Энергетика и эволюция размеров тела мелких наземных млекопитающих» (PDF) . Симпозиумы Лондонского зоологического общества (69): 69–81. Архивировано из оригинала (PDF) 2 июня 2021 года . Проверено 31 мая 2016 г.
- ↑ Перейти обратно: Перейти обратно: а б Уилсон Д.Е., Берни Д., ред. (2001). Животные: Полный визуальный путеводитель по дикой природе мира . Издательство ДК. стр. 86–89. ISBN 978-0-7894-7764-4 . OCLC 46422124 .
- ↑ Перейти обратно: Перейти обратно: а б Ван Валкенбург Б. (июль 2007 г.). «Дежа вю: эволюция морфологии питания хищных животных» . Интегративная и сравнительная биология . 47 (1): 147–163. дои : 10.1093/icb/icm016 . ПМИД 21672827 .
- ^ Сакко Т., ван Валкенбург Б. (2004). «Экоморфологические показатели пищевого поведения медведей (Carnivora: Ursidae)». Журнал зоологии . 263 (1): 41–54. дои : 10.1017/S0952836904004856 .
- ^ Певица М.С., Бернейс Е.А. (2003). «Понимание всеядности требует поведенческой точки зрения» . Экология . 84 (10): 2532–2537. Бибкод : 2003Ecol...84.2532S . дои : 10.1890/02-0397 .
- ^ Хатсон Дж. М., Берк К.С., Хейнс Дж. (1 декабря 2013 г.). «Остеофагия и модификации костей жирафа и других крупных копытных». Журнал археологической науки . 40 (12): 4139–4149. Бибкод : 2013JArSc..40.4139H . дои : 10.1016/j.jas.2013.06.004 .
- ^ «Почему кошки едят траву?» . Пет доктор медицины. Архивировано из оригинала 10 декабря 2016 года . Проверено 13 января 2017 г.
- ^ Гейзер Ф (2004). «Скорость обмена веществ и снижение температуры тела во время спячки и ежедневного оцепенения». Ежегодный обзор физиологии . 66 : 239–274. doi : 10.1146/annurev.phyol.66.032102.115105 . ПМИД 14977403 . S2CID 22397415 .
- ^ Хамфрис М.М., Томас Д.В., Крамер Д.Л. (2003). «Роль доступности энергии в спячке млекопитающих: подход с точки зрения затрат и выгод». Физиологическая и биохимическая зоология . 76 (2): 165–179. дои : 10.1086/367950 . ПМИД 12794670 . S2CID 14675451 .
- ^ Барнс Б.М. (июнь 1989 г.). «Избежание замерзания у млекопитающих: температура тела ниже 0 градусов Цельсия в арктической спячке». Наука . 244 (4912): 1593–1595. Бибкод : 1989Sci...244.1593B . дои : 10.1126/science.2740905 . ПМИД 2740905 .
- ^ Фриц Г. (2010). «Праздник млекопитающих и птиц» . В Навасе, Калифорния, Карвалью Дж.Э. (ред.). Летний период: молекулярные и физиологические аспекты . Прогресс молекулярной и субклеточной биологии. Том. 49. Шпрингер-Верлаг. стр. 95–113. дои : 10.1007/978-3-642-02421-4 . ISBN 978-3-642-02420-7 .
- ^ Майер, с. 59.
- ^ Гроув Дж.С., Грей Л.А., Ла Санта Медина Н., Сивакумар Н., Ан Дж.С., Корпус ТВ, Берке Дж.Д., Крейцер AC, Найт З.А. (июль 2022 г.). «Дофаминовые подсистемы, отслеживающие внутренние состояния» . Природа . 608 (7922): 374–380. Бибкод : 2022Natur.608..374G . дои : 10.1038/s41586-022-04954-0 . ПМЦ 9365689 . ПМИД 35831501 .
- ↑ Перейти обратно: Перейти обратно: а б с д и Веник, с. 105.
- ^ Смит, с. 238.
- ^ «Кошачьи языки используют сложную физику» . 12 ноября 2010 г.
- ^ Смит, с. 237.
- ^ Майер, с. 54.
- ^ «Как жирафы пьют воду?» . Февраль 2016.
- ^ Манн Дж., Паттерсон Э.М. (ноябрь 2013 г.). «Использование орудий водными животными» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 368 (1630): 20120424. doi : 10.1098/rstb.2012.0424 . ПМК 4027413 . ПМИД 24101631 .
- ^ Раффаэле П. (2011). Среди человекообразных обезьян: Приключения по следам наших ближайших сородичей . Нью-Йорк: Харпер. п. 83. ИСБН 978-0-06-167184-5 . OCLC 674694369 .
- ^ Келер В. (1925). Менталитет обезьян . Ливерайт. ISBN 978-0-87140-108-3 . OCLC 2000769 .
- ^ Макгоуэн Р.Т., Рен Т., Норлинг Ю., Килинг Л.Дж. (май 2014 г.). «Позитивный эффект и обучение: изучение «эффекта Эврики» у собак». Познание животных . 17 (3): 577–587. дои : 10.1007/s10071-013-0688-x . ПМИД 24096703 . S2CID 15216926 .
- ^ Карбовски Дж. (май 2007 г.). «Глобальное и региональное масштабирование метаболизма мозга и его функциональные последствия» . БМК Биология . 5 (18): 18. arXiv : 0705.2913 . Бибкод : 2007arXiv0705.2913K . дои : 10.1186/1741-7007-5-18 . ПМК 1884139 . ПМИД 17488526 .
- ^ Марино Л. (июнь 2007 г.). «Мозг китообразных: насколько они водные?» . Анатомическая запись . 290 (6): 694–700. дои : 10.1002/ar.20530 . ПМИД 17516433 . S2CID 27074107 . Архивировано из оригинала 20 марта 2020 года . Проверено 5 октября 2019 г.
- ^ Галоп Г.Г. (январь 1970 г.). «Шимпанзе: самопознание». Наука . 167 (3914): 86–87. Бибкод : 1970Sci...167...86G . дои : 10.1126/science.167.3914.86 . ПМИД 4982211 . S2CID 145295899 .
- ^ Плотник Дж. М., де Ваал Ф. Б., Рейсс Д. (ноябрь 2006 г.). «Самоузнавание в азиатском слоне» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 103 (45): 17053–17057. Бибкод : 2006PNAS..10317053P . дои : 10.1073/pnas.0608062103 . ПМЦ 1636577 . ПМИД 17075063 . Архивировано (PDF) из оригинала 25 января 2024 года . Проверено 25 января 2024 г.
- ^ Роберт С. (1986). «Онтогенез зеркального поведения у двух видов человекообразных обезьян». Американский журнал приматологии . 10 (2): 109–117. дои : 10.1002/ajp.1350100202 . ПМИД 31979488 . S2CID 85330986 .
- ^ Вальравен В., ван Эльсакер Л., Верхейен Р. (1995). «Реакция группы карликовых шимпанзе (Pan paniscus) на свои зеркальные изображения: свидетельства самопознания». Приматы . 36 : 145–150. дои : 10.1007/bf02381922 . S2CID 38985498 .
- ^ Лики Р. (1994). «Происхождение разума» . Происхождение человечества . Нью-Йорк: BasicBooks. п. 150. ИСБН 978-0-465-05313-1 . ОСЛК 30739453 .
- ^ Арчер Дж (1992). Этология и развитие человека . Роуман и Литтлфилд. стр. 215–218. ISBN 978-0-389-20996-6 . OCLC 25874476 .
- ↑ Перейти обратно: Перейти обратно: а б Мартен К., Псаракос С. (1995). «Свидетельства самосознания у афалин ( Tursiops truncatus )». В Паркер С.Т., Митчелл Р., Бочча М. (ред.). Самосознание у животных и людей: перспективы развития . Кембридж: Издательство Кембриджского университета. стр. 361–379. ISBN 978-0-521-44108-7 . ОСЛК 28180680 .
- ↑ Перейти обратно: Перейти обратно: а б Дельфур Ф., Мартен К. (апрель 2001 г.). «Обработка зеркальных изображений трех видов морских млекопитающих: косаток (Orcinus orca), ложных косаток (Pseudorca crassidens) и калифорнийских морских львов (Zalophus Californianus)». Поведенческие процессы . 53 (3): 181–190. дои : 10.1016/s0376-6357(01)00134-6 . ПМИД 11334706 . S2CID 31124804 .
- ^ Джарвис Дж. Ю. (май 1981 г.). «Эусоциальность млекопитающих: совместное размножение в колониях голых землекопов». Наука . 212 (4494): 571–573. Бибкод : 1981Sci...212..571J . дои : 10.1126/science.7209555 . JSTOR 1686202 . ПМИД 7209555 . S2CID 880054 .
- ^ Джейкобс Д.С., Беннетт, Северная Каролина, Джарвис Дж., Кроу Т.М. (1991). «Структура колонии и иерархия доминирования дамаралендского землекопа Cryptomys damarensis (Rodentia: Bathyergidae) из Намибии». Журнал зоологии . 224 (4): 553–576. дои : 10.1111/j.1469-7998.1991.tb03785.x .
- ^ Харди С.Б. (2009). Матери и другие: эволюционные истоки взаимопонимания . Бостон: Belknap Press издательства Гарвардского университета. стр. 92–93.
- ^ Харлоу HF, Суоми SJ (июль 1971 г.). «Социальное восстановление обезьян, выращенных в изоляции» . Труды Национальной академии наук Соединенных Штатов Америки . 68 (7): 1534–1538. Бибкод : 1971PNAS...68.1534H . дои : 10.1073/pnas.68.7.1534 . ПМЦ 389234 . ПМИД 5283943 .
- ^ ван Шайк CP (январь 1999 г.). «Социоэкология социальности деления-слияния у орангутанов». Приматы; Журнал приматологии . 40 (1): 69–86. дои : 10.1007/BF02557703 . ПМИД 23179533 . S2CID 13366732 .
- ^ Арчи Э.А., Мосс СиДжей, Альбертс СК (март 2006 г.). «Связующие узы: генетическое родство предсказывает деление и слияние социальных групп у диких африканских слонов» . Слушания. Биологические науки . 273 (1586): 513–522. дои : 10.1098/rspb.2005.3361 . ПМК 1560064 . ПМИД 16537121 .
- ^ Смит Дж. Э., Меминис С. К., Холекамп К. Э. (2007). «Выбор партнера в зависимости от ранга в обществе деления-слияния пятнистой гиены ( Crocuta crocuta )» (PDF) . Поведенческая экология и социобиология . 61 (5): 753–765. дои : 10.1007/s00265-006-0305-y . S2CID 24927919 . Архивировано из оригинала (PDF) 25 апреля 2014 года.
- ^ Матоба Т., Куцукаке Н., Хасегава Т. (2013). Хейворд М. (ред.). «Потирание и облизывание головы укрепляют социальные связи в группе содержащихся в неволе африканских львов Panthera leo» . ПЛОС ОДИН . 8 (9): e73044. Бибкод : 2013PLoSO...873044M . дои : 10.1371/journal.pone.0073044 . ПМЦ 3762833 . ПМИД 24023806 .
- ^ Круцен М., Барре Л.М., Коннор Р.К., Манн Дж., Шервин В.Б. (июль 2004 г.). « О отец, где ты?» – Оценка отцовства в открытом обществе деления-слияния диких афалин (Tursiops sp.) в заливе Шарк, Западная Австралия». Молекулярная экология . 13 (7): 1975–1990. Бибкод : 2004МолЭк..13.1975К . дои : 10.1111/j.1365-294X.2004.02192.x . ПМИД 15189218 . S2CID 4510393 .
- ^ Мартин С. (1991). Тропические леса Западной Африки: экология – угрозы – сохранение . Спрингер. дои : 10.1007/978-3-0348-7726-8 . ISBN 978-3-0348-7726-8 .
- ^ ле Ру А., Черри М.И., Гайгакс Л. (5 мая 2009 г.). «Поведение бдительности и последствия фитнеса: сравнение одиночного добывания пищи и млекопитающего, обязательного группового добывания пищи». Поведенческая экология и социобиология . 63 (8): 1097–1107. дои : 10.1007/s00265-009-0762-1 . S2CID 21961356 .
- ^ Палаги Э, Норша I (2015). Сэмондс К.Э. (ред.). «Сезон мира: примирение деспотического вида (Lemur catta)» . ПЛОС ОДИН . 10 (11): e0142150. Бибкод : 2015PLoSO..1042150P . дои : 10.1371/journal.pone.0142150 . ПМЦ 4646466 . ПМИД 26569400 .
- ^ Ист МЛ, Хофер Х (2000). «Самцы пятнистых гиен ( Crocuta crocuta ) стоят в очереди за статусом в социальных группах, где доминируют самки» . Поведенческая экология . 12 (15): 558–568. дои : 10.1093/beheco/12.5.558 .
- ^ Сэмюэлс А., Силк Дж.Б. , Родман П. (1984). «Изменения в ранге доминирования и репродуктивном поведении самцов шляпных макак ( Macaca radiate )». Поведение животных . 32 (4): 994–1003. дои : 10.1016/s0003-3472(84)80212-2 . S2CID 53186523 .
- ^ Дельпьетро Х.А., Руссо Р.Г. (2002). «Наблюдения за обыкновенной летучей мышью-вампиром ( Desmodus rotundus ) и летучей мышью-вампиром с волосатыми ногами ( Diphylla ecaudata ) в неволе». Биология млекопитающих . 67 (2): 65–78. дои : 10.1078/1616-5047-00011 .
- ^ Клейман Д.Г. (март 1977 г.). «Моногамия у млекопитающих». Ежеквартальный обзор биологии . 52 (1): 39–69. дои : 10.1086/409721 . ПМИД 857268 . S2CID 25675086 .
- ^ Холланд Б., Райс В.Р. (февраль 1998 г.). «Перспектива: преследование сексуального отбора: антагонистическое соблазнение против сопротивления» (PDF) . Эволюция; Международный журнал органической эволюции . 52 (1): 1–7. дои : 10.2307/2410914 . JSTOR 2410914 . ПМИД 28568154 . Архивировано из оригинала (PDF) 8 июня 2019 года . Проверено 8 июля 2016 г.
- ^ Клаттон-Брок, TH (май 1989 г.). «Системы спаривания млекопитающих» . Труды Лондонского королевского общества. Серия Б, Биологические науки . 236 (1285): 339–372. Бибкод : 1989РСПСБ.236..339С . дои : 10.1098/rspb.1989.0027 . ПМИД 2567517 . S2CID 84780662 .
- ^ Бонесс DJ, Боуэн Д., Бюлейер Б.М., Маршалл Дж.Дж. (2006). «Тактика спаривания и система спаривания водно-спаривающихся ластоногих: обыкновенный тюлень Phoca vitulina » . Поведенческая экология и социобиология . 61 : 119–130. дои : 10.1007/s00265-006-0242-9 . S2CID 25266746 .
- ^ Клопфер П.Х. (1981). «Истоки родительской заботы» . В Губернике DJ (ред.). Родительская забота у млекопитающих . Нью-Йорк: Пленум Пресс. ISBN 978-1-4613-3150-6 . OCLC 913709574 .
- ^ Мерти Р., Бирман Г., Браун С., Брайант К., Чинн Р., Хьюлетт А. и др. (май 2015 г.). «Животные в медицинских учреждениях: рекомендации по минимизации потенциальных рисков» (PDF) . Инфекционный контроль и госпитальная эпидемиология . 36 (5): 495–516. дои : 10.1017/ice.2015.15 . ПМИД 25998315 . S2CID 541760 . Архивировано (PDF) из оригинала 3 ноября 2023 года.
- ^ Гуманное общество США. «Статистика владения домашними животными в США» . Архивировано из оригинала 7 апреля 2012 года . Проверено 27 апреля 2012 г.
- ^ Министерство сельского хозяйства США. «Профиль кролиководческой отрасли США» (PDF) . Архивировано из оригинала (PDF) 7 августа 2019 года . Проверено 10 июля 2013 г.
- ^ Маккай Р. (26 мая 2013 г.). «Доисторическое наскальное искусство в Дордони» . Хранитель . Архивировано из оригинала 31 мая 2021 года . Проверено 9 ноября 2016 г.
- ^ Джонс Дж. (27 июня 2014 г.). «10 лучших портретов животных в искусстве» . Хранитель . Архивировано из оригинала 18 мая 2016 года . Проверено 24 июня 2016 г.
- ^ «Охота на оленей в Соединенных Штатах: анализ демографии и поведения охотников, дополнение к отчету о национальном обследовании рыболовства, охоты и отдыха, связанного с дикой природой, 2001–2006 гг.» . Служба рыболовства и дикой природы (США). Архивировано из оригинала 13 августа 2016 года . Проверено 24 июня 2016 г.
- ^ Шелтон Л. (5 апреля 2014 г.). «Рост популярности рекреационной охоты на свиней» . Натчезский демократ . Гранд-Вью на открытом воздухе. Архивировано из оригинала 12 декабря 2017 года . Проверено 24 июня 2016 г.
- ^ Нгуен Дж., Уитли Р. (2015). Охота за едой: Руководство по сбору урожая, полевой обработке и приготовлению дичи . Ф+В Медиа. стр. 6–77. ISBN 978-1-4403-3856-4 . Главы об охоте на оленя, кабана (кабана), кролика и белку.
- ^ «Скачки» . Британская энциклопедия . Архивировано из оригинала 21 декабря 2013 года . Проверено 6 мая 2014 г.
- ^ Гендеры Р (1981). Энциклопедия собачьих бегов . Книги Пелхэма. ISBN 978-0-7207-1106-6 . OCLC 9324926 .
- ^ Плюс С (1993). «Роль животных в человеческом обществе». Журнал социальных проблем . 49 (1): 1–9. дои : 10.1111/j.1540-4560.1993.tb00906.x .
- ^ Фаулер К.Дж. (26 марта 2014 г.). «10 лучших книг об умных животных» . Хранитель . Архивировано из оригинала 28 мая 2021 года . Проверено 9 ноября 2016 г.
- ^ Гэмбл Н., Йейтс С. (2008). Изучение детской литературы (2-е изд.). Лос-Анджелес: Сейдж. ISBN 978-1-4129-3013-0 . OCLC 71285210 .
- ^ «Книги для взрослых» . Натурщики тюленей . Архивировано из оригинала 11 июля 2016 года . Проверено 9 ноября 2016 г.
- ^ Патерсон Дж (2013). «Животные в кино и медиа». Оксфордские библиографии . дои : 10.1093/обо/9780199791286-0044 .
- ^ Джонс С. (2011). Крупный рогатый скот: история, миф, искусство . Лондон: Издательство Британского музея. ISBN 978-0-7141-5084-0 . OCLC 665137673 .
- ^ ван Гулик Р.Х. Хаягрива: Мантраянский аспект культа лошади в Китае и Японии . Архив Брилла. п. 9.
- ^ Грейнджер Р. (24 июня 2012 г.). «Изображение льва в древних и современных религиях» . ТРЕВОГА. Архивировано из оригинала 23 сентября 2016 года . Проверено 6 ноября 2016 г.
- ^ Даймонд Дж. М. (1997). «Часть 2: Возникновение и распространение производства продуктов питания» . Оружие, микробы и сталь: судьбы человеческих обществ . Нью-Йорк: WW Norton & Company. ISBN 978-0-393-03891-0 . ОСЛК 35792200 .
- ^ Ларсон Дж., Бургер Дж. (апрель 2013 г.). «Взгляд на приручение животных с точки зрения популяционной генетики» (PDF) . Тенденции в генетике . 29 (4): 197–205. дои : 10.1016/j.tig.2013.01.003 . ПМИД 23415592 . Архивировано из оригинала (PDF) 8 июня 2019 года . Проверено 9 ноября 2016 г.
- ^ Зедер М.А. (август 2008 г.). «Одомашнивание и раннее сельское хозяйство в Средиземноморском бассейне: происхождение, распространение и влияние» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (33): 11597–11604. Бибкод : 2008PNAS..10511597Z . дои : 10.1073/pnas.0801317105 . ПМЦ 2575338 . ПМИД 18697943 .
- ^ «Графическая деталь. Графики, карты и инфографика. Подсчет кур» . Экономист . 27 июля 2011 г. Архивировано из оригинала 15 июля 2016 г. Проверено 6 ноября 2016 г.
- ^ «Породы крупного рогатого скота в КРС СЕГОДНЯ» . Крупный рогатый скот сегодня . Крупный рогатый скот-сегодня.com. Архивировано из оригинала 15 июля 2011 года . Проверено 6 ноября 2016 г.
- ^ Люкфар С.Д., Чик PR. «Стратегии развития кролиководческих проектов в системах натурального хозяйства» . Продовольственная и сельскохозяйственная организация . Архивировано из оригинала 6 мая 2016 года . Проверено 6 ноября 2016 г.
- ^ Пруд РГ (2004). Энциклопедия зоотехники . ЦРК Пресс. стр. 248–250. ISBN 978-0-8247-5496-9 . OCLC 57033325 . Архивировано из оригинала 23 января 2023 года . Проверено 5 октября 2018 г.
- ^ Бротен А.В. (2005). "Шерсть" . В Стиле V (ред.). Энциклопедия одежды и моды . Том. 3. Томсон Гейл . стр. 441–443 . ISBN 978-0-684-31394-8 . ОСЛК 963977000 .
- ^ Quiggle C (осень 2000 г.). «Альпака: древняя роскошь». Переплетение лиц : 74–76.
- ^ «Дикие млекопитающие составляют лишь несколько процентов млекопитающих мира» . Наш мир в данных . Проверено 8 августа 2023 г.
- ^ «Генетические исследования» . Фонд здоровья животных. Архивировано из оригинала 12 декабря 2017 года . Проверено 6 ноября 2016 г.
- ^ «Разработка лекарств» . Исследования животных.info. Архивировано из оригинала 8 июня 2016 года . Проверено 6 ноября 2016 г.
- ^ «Статистика ЕС показывает снижение количества исследований на животных» . Кстати об исследованиях. 2013. Архивировано из оригинала 24 апреля 2019 года . Проверено 6 ноября 2016 г.
- ^ Пилчер Х.Р. (2003). «Это нокаут» . Природа . дои : 10.1038/news030512-17 . Архивировано из оригинала 10 ноября 2016 года . Проверено 6 ноября 2016 г.
- ^ «Поставка и использование приматов в ЕС» . Европейская ассоциация биомедицинских исследований. 1996. Архивировано из оригинала 17 января 2012 года.
- ^ Карлссон Х.Э., Шапиро С.Дж., Фарах И., Хау Дж. (август 2004 г.). «Использование приматов в исследованиях: глобальный обзор». Американский журнал приматологии . 63 (4): 225–237. дои : 10.1002/ajp.20054 . ПМИД 15300710 . S2CID 41368228 .
- ^ Уэзералл Д. и др. (2006). Использование приматов в исследованиях (PDF) (Отчет). Лондон: Академия медицинских наук. Архивировано из оригинала (PDF) 23 марта 2013 года.
- ^ Ричи Х., Розер М. (15 апреля 2021 г.). «Биоразнообразие» . Наш мир в данных . Архивировано из оригинала 11 декабря 2022 года . Проверено 29 августа 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б Бар-Он Ю.М., Филлипс Р., Майло Р. (июнь 2018 г.). «Распределение биомассы на Земле» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (25): 6506–6511. Бибкод : 2018PNAS..115.6506B . дои : 10.1073/pnas.1711842115 . ПМК 6016768 . ПМИД 29784790 .
- ^ Прайс Е (2008). Принципы и применение поведения домашних животных: вводный текст . Сакраменто: Издательство Кембриджского университета. ISBN 978-1-84593-398-2 . ОСЛК 226038028 .
- ^ Таупитц Дж., Весшка М. (2009). Химбриды – химеры и гибриды в сравнительных европейских и международных исследованиях . Гейдельберг: Спрингер. п. 13. ISBN 978-3-540-93869-9 . OCLC 495479133 .
- ^ Чемберс С.М., Фейн С.Р., Фацио Б., Амарал М. (2012). «Отчет о систематике североамериканских волков на основе морфологического и генетического анализа» . Североамериканская фауна . 77 :2. дои : 10.3996/нафа.77.0001 . Архивировано из оригинала 31 мая 2021 года . Проверено 12 октября 2019 г.
- ^ ван Вюр Т (2005). По следам зубра – история, морфология и экология вымершего дикого быка . Издательство Пенсофт. ISBN 978-954-642-235-4 . OCLC 940879282 .
- ^ Муни Х.А., Клеланд Э.Э. (май 2001 г.). «Эволюционное влияние инвазивных видов» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (10): 5446–5451. Бибкод : 2001PNAS...98.5446M . дои : 10.1073/pnas.091093398 . ПМЦ 33232 . ПМИД 11344292 .
- ^ Ле Ру Дж. Дж., Фокскрофт Л. К., Хербст М., Макфадьен С. (январь 2015 г.). «Генетический анализ показывает низкий уровень гибридизации между африканскими дикими кошками (Felis silvestris lybica) и домашними кошками (F. s. catus) в Южной Африке» . Экология и эволюция . 5 (2): 288–299. Бибкод : 2015EcoEv...5..288L . дои : 10.1002/ece3.1275 . ПМК 4314262 . ПМИД 25691958 .
- ^ Уилсон А. (2003). Отчет о состоянии лесов Австралии . п. 107.
- ^ Раймер Дж. М., Симберлофф Д. (ноябрь 1996 г.). «Вымирание путем гибридизации и интрогрессии». Ежегодный обзор экологии и систематики . 27 : 83–109. doi : 10.1146/annurev.ecolsys.27.1.83 .
- ^ Поттс Б.М. (2001). Барбур Р.К., Хингстон AB (ред.). Генетическое загрязнение в результате фермерского лесного хозяйства с использованием видов и гибридов эвкалипта: отчет для программы совместного предприятия RIRDC/L&WA/FWPRDC по агролесомелиорации . Корпорация сельских промышленных исследований и развития Австралии. ISBN 978-0-642-58336-9 . OCLC 48794104 .
- ↑ Перейти обратно: Перейти обратно: а б Дирзо Р., Янг Х.С., Галетти М., Себальос Г., Исаак Н.Дж., Коллен Б. (июль 2014 г.). «Дефаунация в антропоцене» (PDF) . Наука . 345 (6195): 401–406. Бибкод : 2014Sci...345..401D . дои : 10.1126/science.1251817 . ПМИД 25061202 . S2CID 206555761 . Архивировано (PDF) из оригинала 7 августа 2019 года . Проверено 25 января 2024 г.
- ^ Примак Р. (2014). Основы биологии сохранения (6-е изд.). Сандерленд, Массачусетс: Издательство Sinauer Associates, Inc. стр. 217–245. ISBN 978-1-60535-289-3 . OCLC 876140621 .
- ^ Виньери С (июль 2014 г.). «Исчезающая фауна. Введение» . Наука . 345 (6195): 392–395. Бибкод : 2014Sci...345..392V . дои : 10.1126/science.345.6195.392 . ПМИД 25061199 .
- ^ Берни Д.А., Фланнери Т.Ф. (июль 2005 г.). «Пятьдесят тысячелетий катастрофических вымираний после контакта с человеком» (PDF) . Тенденции в экологии и эволюции . 20 (7): 395–401. дои : 10.1016/j.tree.2005.04.022 . ПМИД 16701402 . Архивировано из оригинала (PDF) 10 июня 2010 года.
- ^ Даймонд Дж (1984). «Исторические вымирания: Розеттский камень для понимания доисторических вымираний». Мартин П.С., Кляйн Р.Г. (ред.). Четвертичные вымирания: доисторическая революция . Тусон: Издательство Университета Аризоны. стр. 824–862. ISBN 978-0-8165-1100-6 . ОСЛК 10301944 .
- ^ Уоттс Дж. (6 мая 2019 г.). «Человеческое общество находится под серьезной угрозой утраты естественной жизни на Земле» . Хранитель . Архивировано из оригинала 14 июня 2019 года . Проверено 1 июля 2019 г.
- ^ МакГрат М. (6 мая 2019 г.). «Природный кризис: люди угрожают исчезновением миллиону видов » . Би-би-си . Архивировано из оригинала 30 июня 2019 года . Проверено 1 июля 2019 г.
- ^ Главный D (22 ноября 2013 г.). «7 знаковых животных, которых люди ведут к вымиранию» . Живая наука . Архивировано из оригинала 6 января 2023 года . Проверено 25 января 2024 г.
- ^ Платт-младший (25 октября 2011 г.). «Браконьеры привели к вымиранию яванского носорога во Вьетнаме» . Научный американец . Архивировано из оригинала 6 апреля 2015 года.
- ^ Кэррингтон Д. (8 декабря 2016 г.). «Жирафам грозит исчезновение после разрушительного сокращения, предупреждают эксперты» . Хранитель . Архивировано из оригинала 13 августа 2021 года . Проверено 4 февраля 2017 г.
- ^ Эстрада А., Гарбер П.А., Райландс А.Б., Роос С., Фернандес-Дюке Е., Ди Фиоре А. и др. (январь 2017 г.). «Надвигающийся кризис вымирания приматов в мире: почему приматы имеют значение» . Достижения науки . 3 (1): e1600946. Бибкод : 2017SciA....3E0946E . дои : 10.1126/sciadv.1600946 . ПМЦ 5242557 . ПМИД 28116351 .
- ^ Флетчер М. (31 января 2015 г.). «Панголины: почему это милое доисторическое млекопитающее находится на грани исчезновения» . Телеграф . Архивировано из оригинала 10 января 2022 года.
- ^ Гринфилд П. (9 сентября 2020 г.). «Люди эксплуатируют и уничтожают природу в беспрецедентных масштабах – доклад» . Хранитель . Архивировано из оригинала 21 октября 2021 года . Проверено 13 октября 2020 г.
- ^ Маккарти Д. (1 октября 2020 г.). «Ужасающие потери дикой природы показывают, что конец игры по вымиранию начался, но еще не поздно измениться» . Независимый . Архивировано из оригинала 7 апреля 2023 года . Проверено 13 октября 2020 г.
- ^ Пенниси Э (18 октября 2016 г.). «Люди истребляют приматов, летучих мышей и других млекопитающих» . Наука . Архивировано из оригинала 20 октября 2021 года . Проверено 3 февраля 2017 г.
- ^ Риппл У.Дж., Абернети К., Беттс М.Г., Чапрон Г., Дирзо Р., Галетти М. и др. (октябрь 2016 г.). «Охота на мясо диких животных и риск исчезновения млекопитающих мира» . Королевское общество открытой науки . 3 (10): 160498. Бибкод : 2016RSOS....360498R . дои : 10.1098/rsos.160498 . hdl : 1893/24446 . ПМК 5098989 . ПМИД 27853564 .
- ^ Уильямс М., Заласевич Дж., Хафф П.К., Швегерль С., Барноски А.Д. , Эллис ЕС (2015). «Антропоценовая биосфера». Обзор антропоцена . 2 (3): 196–219. Бибкод : 2015AntRv...2..196W . дои : 10.1177/2053019615591020 . S2CID 7771527 .
- ^ Морелл В. (11 августа 2015 г.). «Мясоеды могут ускорить исчезновение видов во всем мире, предупреждает исследование» . Наука . Архивировано из оригинала 20 декабря 2016 года . Проверено 3 февраля 2017 г.
- ^ Маховина Б., Фили К.Дж., Риппл У.Дж. (декабрь 2015 г.). «Сохранение биоразнообразия: главное — сократить потребление мяса». Наука об общей окружающей среде . 536 : 419–431. Бибкод : 2015ScTEn.536..419M . doi : 10.1016/j.scitotenv.2015.07.022 . ПМИД 26231772 .
- ^ Редфорд К.Х. (1992). «Пустой лес» (PDF) . Бионаука . 42 (6): 412–422. дои : 10.2307/1311860 . JSTOR 1311860 . Архивировано (PDF) оригинала 28 февраля 2021 года . Проверено 4 февраля 2017 г.
- ^ Перес К.А., Насименто Х.С. (2006). «Влияние охоты на дичь каяпо на юго-востоке Амазонии: последствия для сохранения дикой природы в местных заповедниках тропических лесов». Человеческая эксплуатация и сохранение биоразнообразия . Том. 3. Спрингер. стр. 287–313. ISBN 978-1-4020-5283-5 . OCLC 207259298 .
- ^ Альтрихтер М., Боальо Г. (2004). «Распространение и относительная численность пекари в аргентинском Чако: связь с человеческим фактором». Биологическая консервация . 116 (2): 217–225. Бибкод : 2004BCons.116..217A . дои : 10.1016/S0006-3207(03)00192-7 .
- ^ Гобуш К. «Влияние браконьерства на африканских слонов» . Центр биологии сохранения . Университет Вашингтона. Архивировано из оригинала 8 декабря 2021 года . Проверено 12 мая 2021 г.
- ^ Алверсон Д.Л., Фрибург М.Х., Муравски С.А., Поуп Дж.Г. (1996) [1994]. «Прилов морских млекопитающих» . Глобальная оценка рыболовного прилова и выбросов . Рим: Продовольственная и сельскохозяйственная организация Объединенных Наций. ISBN 978-92-5-103555-9 . ОСЛК 31424005 . Архивировано из оригинала 17 февраля 2019 года . Проверено 25 января 2024 г.
- ^ Гловка Л., Бурхенне-Гильмин Ф., Synge HM, Макнили Дж.А., Гюндлинг Л. (1994). Документ по экологической политике и законодательству МСОП . Путеводитель по Конвенции о биоразнообразии. Международный союз охраны природы. ISBN 978-2-8317-0222-3 . ОСЛК 32201845 .
- ^ «О МСОП» . Международный союз охраны природы. 3 декабря 2014 г. Архивировано из оригинала 15 апреля 2020 г. . Проверено 3 февраля 2017 г.
- ^ Себальос Г., Эрлих П.Р., Барноски А.Д., Гарсиа А., Прингл Р.М., Палмер Т.М. (июнь 2015 г.). «Ускоренная антропогенная утрата видов: на пороге шестого массового вымирания» . Достижения науки . 1 (5): e1400253. Бибкод : 2015SciA....1E0253C . дои : 10.1126/sciadv.1400253 . ПМК 4640606 . ПМИД 26601195 .
- ^ Фишер Д.О., Бломберг С.П. (апрель 2011 г.). «Корреляты повторного открытия и возможность обнаружения вымирания млекопитающих» . Слушания. Биологические науки . 278 (1708): 1090–1097. дои : 10.1098/rspb.2010.1579 . ПМК 3049027 . ПМИД 20880890 .
- ^ Себальос Г., Эрлих А.Х., Эрлих П.Р. (2015). Уничтожение природы: вымирание птиц и млекопитающих человеком . Балтимор: Издательство Университета Джонса Хопкинса. п. 69. ИСБН 978-1-4214-1718-9 .
- ^ Цзян, З.; Харрис, РБ (2016). « Элафурус Давидианус » . Красный список исчезающих видов МСОП . 2016 : e.T7121A22159785. doi : 10.2305/IUCN.UK.2016-2.RLTS.T7121A22159785.en . Проверено 12 ноября 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б МакКинни М.Л., Шох Р., Йонавьяк Л. (2013). «Сохранение биологических ресурсов» . Наука об окружающей среде: системы и решения (5-е изд.). Джонс и Бартлетт Обучение. ISBN 978-1-4496-6139-7 . OCLC 777948078 .
- ^ Перрин В.Ф., Вюрсиг Б.Ф., Тьювиссен Дж.Г. (2009). Энциклопедия морских млекопитающих . Академическая пресса. п. 404. ИСБН 978-0-12-373553-9 . OCLC 455328678 .
- На момент редактирования в этой статье используется контент из раздела «Анатомия и физиология» , который лицензируется таким образом, чтобы его можно было повторно использовать по лицензии Creative Commons Attribution-ShareAlike 3.0 Unported License , но не по GFDL . Все соответствующие условия должны быть соблюдены.
Дальнейшее чтение
- Браун ВМ (2001). «Естественный отбор компонентов мозга млекопитающих». Тенденции экологии и эволюции . 16 (9): 471–473. дои : 10.1016/S0169-5347(01)02246-7 .
- Маккенна MC, Bell SK (1997). Классификация млекопитающих выше видового уровня . Нью-Йорк: Издательство Колумбийского университета. ISBN 978-0-231-11013-6 . OCLC 37345734 . [ постоянная мертвая ссылка ]
- Новак Р.М. (1999). Млекопитающие мира Уокера (6-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-5789-8 . OCLC 937619124 .
- Симпсон Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Бюллетень Американского музея естественной истории . 85 : 1–350.
- Мерфи В.Дж., Эйзирик Э., О'Брайен С.Дж., Мэдсен О., Скалли М., Дуади С.Дж. и др. (декабрь 2001 г.). «Разрешение ранней плацентарной радиации млекопитающих с использованием байесовской филогенетики». Наука . 294 (5550): 2348–2351. Бибкод : 2001Sci...294.2348M . дои : 10.1126/science.1067179 . ПМИД 11743200 . S2CID 34367609 .
- Спрингер М.С., Стэнхоуп М.Дж., Мэдсен О., де Йонг В.В. (август 2004 г.). «Молекулы консолидируют плацентарное дерево млекопитающих» (PDF) . Тенденции в экологии и эволюции . 19 (8): 430–438. дои : 10.1016/j.tree.2004.05.006 . ПМИД 16701301 . S2CID 1508898 . Архивировано из оригинала (PDF) 29 июля 2016 года . Проверено 21 января 2005 г.
- Воган Т.А., Райан Дж.М., Капзаплевски, Нью-Джерси (2000). Маммология (4-е изд.). Форт-Уэрт, Техас: Издательство Saunders College. ISBN 978-0-03-025034-7 . OCLC 42285340 .
- Кригс Дж.О., Чураков Г., Кифманн М., Джордан У., Брозиус Дж., Шмитц Дж. (апрель 2006 г.). «Ретропозированные элементы как архивы эволюционной истории плацентарных млекопитающих» . ПЛОС Биология . 4 (4): е91. doi : 10.1371/journal.pbio.0040091 . ПМЦ 1395351 . ПМИД 16515367 .
- Макдональд Д.В., Норрис С. (2006). Энциклопедия млекопитающих (3-е изд.). Лондон: Справочная группа Брауна. ISBN 978-0-681-45659-4 . OCLC 74900519 .
Внешние ссылки
- База данных ASM по разнообразию млекопитающих. Архивировано 25 декабря 2022 г. на Wayback Machine.
- Biodiversitymapping.org - Все отряды млекопитающих в мире с картами распространения. Архивировано 26 сентября 2016 г. в Wayback Machine.
- Палеоценовые млекопитающие. Архивировано 3 февраля 2024 года на Wayback Machine , сайте, освещающем возникновение млекопитающих, paleocene-mammals.de.
- Эволюция млекопитающих. Архивировано 25 января 2024 года в Wayback Machine , краткое введение в ранних млекопитающих, enchantedlearning.com.
- Европейский атлас млекопитающих EMMA. Архивировано 25 января 2024 г. в Wayback Machine от Societas Europaea Mammalogica, European-mammals.org.
- Морские млекопитающие мира. Архивировано 8 июня 2019 года в Wayback Machine . Обзор всех морских млекопитающих, включая описания, как полностью водных, так и полуводных, noaa.gov.
- Mammalogy.org. Архивировано 1 марта 2020 года в Wayback Machine. Американское общество маммологов было основано в 1919 году с целью содействия изучению млекопитающих, и этот веб-сайт включает библиотеку изображений млекопитающих.










