Птерозавры
| Птерозавры Временной диапазон: поздний триас – поздний мел , | |
|---|---|
 | |
| Шесть птерозавров (слева сверху вниз справа): Диморфодон , Птеродактиль , Анурогнат , Кетцалькоатль , Сорд , Тропеогнат. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Орнитодира |
| Клэйд : | † Птерозавроморфа |
| Заказ: | † Птерозаврия Покупка , 1834 г. |
| Подгруппы [1] [2] | |
| |
 | |
| Распределение местонахождений окаменелостей птерозавров . Цветные названия видов или родов соответствуют их таксономической группе. [а] | |
| Синонимы | |
Орнитозаврия Сили , 1870 г. | |
Птерозавры ( / ˈ t ɛr ə s ɔːr , ˈ t ɛr oʊ -/ ; [5] [6] от греческого pteron и sauros , что означает «крылатая ящерица») [7] — вымершая группа летающих рептилий отряда птерозавров . Они существовали на протяжении большей части мезозоя : от позднего триаса до конца мела ( 228–66 миллионов лет назад). [8] Птерозавры — самые ранние позвоночные, которые, как известно, развили двигательный полет . Их крылья были образованы мембраной из кожи, мышц и других тканей, простирающейся от лодыжек до значительно удлинившегося безымянного пальца. [9]
Существовало два основных типа птерозавров. Базальные птерозавры (также называемые «нептеродактилоидными птерозаврами» или « рамфоринхоидами ») были более мелкими животными с полностью зубчатыми челюстями и, как правило, длинными хвостами. Их широкие перепонки крыльев, вероятно, включали и соединяли задние ноги. На земле они могли бы принять неуклюжую распростертую позу, но анатомия их суставов и сильные когти сделали бы их эффективными альпинистами, а некоторые, возможно, даже жили на деревьях. Базальные птерозавры были насекомоядными или хищниками мелких позвоночных. Позже птерозавры ( птеродактилоиды ) развили различные размеры, формы и образ жизни. У птеродактилоидов были более узкие крылья со свободными задними конечностями, сильно редуцированные хвосты и длинные шеи с большими головами. По земле они хорошо ходили на всех четырех конечностях, сохраняя вертикальное положение, стоя стопоходя на задних лапах и загибая крыльевой палец вверх, чтобы ходить на трехпалой «руке». Они могли взлетать с земли, и ископаемые следы показывают, что по крайней мере некоторые виды были способны бегать, переходить вброд или плавать. [10] На их челюстях были роговые клювы, а у некоторых групп отсутствовали зубы. У некоторых групп развились сложные гребни на голове с половым диморфизмом .
У птерозавров была оболочка из волосоподобных нитей, известных как пикноволокна , которые покрывали их тела и части крыльев. Пикноволокна росли в нескольких формах: от простых нитей до ветвящихся перьев . Они могут быть гомологичны пуховым перьям, обнаруженным как у птиц , так и у некоторых нептичьих динозавров , что позволяет предположить, что ранние перья развились у общего предка птерозавров и динозавров, возможно, в качестве изоляции. [11] При жизни птерозавры имели бы гладкую или пушистую шерсть, не похожую на птичьи перья. Это были теплокровные (эндотермные) активные животные. Дыхательная система имела эффективное однонаправленное «проточное» дыхание с использованием воздушных мешков , которые до чрезвычайной степени опустошали их кости. Птерозавры охватывали широкий диапазон взрослых размеров : от очень маленьких анурогнатид до крупнейших известных летающих существ, включая кетцалькоатля и хацегоптерикса . [12] [13] [14] который достигал размаха крыльев не менее девяти метров. Сочетание эндотермии , хорошего снабжения кислородом и сильных мышц сделало птерозавров сильными и способными летать.
или широкая общественность часто называют птерозавров «летающими динозаврами», но динозавры определяются как потомки последнего общего предка Saurischia Популярные средства массовой информации и Ornithischia , что исключает птерозавров. [15] Тем не менее птерозавры более тесно связаны с птицами и другими динозаврами, чем с крокодилами или любыми другими живыми рептилиями, хотя они и не являются предками птиц. Птерозавров также в разговорной речи называют птеродактилями , особенно в художественной литературе и журналистике. [16] Однако технически птеродактиль может относиться к членам рода Pterodactylus и, в более широком смысле, к членам подотряда Pterodactyloidea . птерозавров [17]
Птерозавры вели разнообразный образ жизни. Традиционно считавшаяся рыбоядными, теперь считается, что в эту группу также входили охотники на наземных животных, насекомоядные, питающиеся фруктами и даже хищники других птерозавров. Они размножаются яйцами , некоторые окаменелости которых были обнаружены.
Описание
[ редактировать ]Анатомия птерозавров сильно отличалась от их предков-рептилий из-за адаптации к полету. птерозавров Кости были полыми и наполненными воздухом, как у птиц . Это обеспечило более высокую поверхность прикрепления мышц при данном скелетном весе. Костяные стены зачастую были тонкими, как бумага. У них была большая килевидная грудная кость для летательных мышц и увеличенный мозг, способный координировать сложное летное поведение. [18] Скелеты птерозавров часто демонстрируют значительное слияние. В черепе исчезли швы между элементами. У некоторых более поздних птерозавров позвоночник над плечами сросся в структуру, известную как нотариум , которая служила для придания туловищу жесткости во время полета и обеспечивала стабильную поддержку лопатки . Точно так же крестцовые позвонки могли образовывать единый синкрестец , в то время как кости таза также срастались.
Базальные птерозавры включают клады Dimorphodontidae ( Dimorphodon ), Campylognathididae ( Eudimorphodon , Campyognathoides ) и Rhamphorhynchidae ( Rhamphorhynchus , Scaphognathus ).
К птеродактилоидам относятся клады Ornithocheiroidea ( Istiodactylus , Ornithocheirus , Pteranodon ), Ctenochasmatoidea ( Ctenochasma , Pterodactylus ), Dsungaripteroidea ( Germanodactylus , Dsungaripterus ) и Azhdarchoidea ( Tapejara , Tupuxuara , Quetzalcoatlus ).
Эти две группы перекрывались во времени, но самые ранние птерозавры в летописи окаменелостей — это базальные птерозавры, а самые поздние птерозавры — птеродактилоиды. [19]
Положение клады Anurognathidae ( Anurognathus , Jeholopterus , Vesperopterylus ) дискутируется. [20] Анурогнатиды были высокоспециализированными. Небольшие летуны с укороченными челюстями и широким зевом, у некоторых были большие глаза, указывающие на ночной или сумеречный образ жизни, щетина во рту и ступни, приспособленные для цепляния. Параллельные адаптации наблюдаются у птиц и летучих мышей, которые охотятся на насекомых в полете.
Размер
[ редактировать ]
Птерозавры имели широкий диапазон размеров, хотя в целом они были крупными. Самый маленький вид имел размах крыльев не менее 25 сантиметров (10 дюймов). [12] Самые крупные формы представляют собой самых крупных известных животных, когда-либо летавших, с размахом крыльев до 10–11 метров (33–36 футов). [21]
Стоя, такие гиганты могли достигать роста современного жирафа . Традиционно считалось, что птерозавры были чрезвычайно легкими по сравнению с их размером. Позже стало понятно, что это будет означать нереально низкую плотность их мягких тканей. Поэтому по некоторым современным оценкам вес самых крупных видов может достигать 250 килограммов (550 фунтов). [22]
Череп, зубы и гребни
[ редактировать ]
По сравнению с другими летающими группами позвоночных, птицами и летучими мышами, черепа птерозавров обычно были довольно большими. [23] Большинство черепов птерозавров имели удлиненные челюсти. [23] Кости черепа у взрослых особей имеют тенденцию срастаться. [23] У ранних птерозавров часто были гетеродонтные зубы разного строения, а у некоторых все еще были зубы на небе. В более поздних группах зубы преимущественно стали коническими. [24] Передние зубы часто были длиннее, образуя «хватку добычи» в поперечно расширенных кончиках челюстей, но размер и положение у разных видов сильно различались. [25] У производных Pterodactyloidea черепа стали еще более удлиненными, иногда превосходя по длине вместе взятые шею и туловище. Это было вызвано растяжением и сращением передней кости морды, предчелюстной кости , с костью верхней челюсти, верхней челюсти . В отличие от большинства архозавров , носовое и анторбитальное отверстия птеродактилоидных птерозавров слились в одно большое отверстие, называемое носоанторбитальным окном . [26] Эта особенность, вероятно, возникла для того, чтобы облегчить череп для полета. [24] Напротив, кости позади глазницы сжимались и вращались, сильно наклоняя заднюю часть черепа и выдвигая челюстной сустав вперед. [27] Мозговая коробка была относительно большой для рептилий. [28]
В некоторых случаях окаменевшая ороговевшая ткань клюва, хотя у зубчатых форм клюв небольшой, ограничен кончиками челюстей и не затрагивает зубы. сохранилась [29] Некоторые продвинутые формы с клювами были беззубыми, такими как Pteranodontidae и Azhdarchidae , и имели более крупные, обширные и более птичьи клювы. [24] Некоторые группы имели специализированные формы зубов. были У Istiodactylidae загнутые зубы, позволяющие есть мясо. Ctenochasmatidae для фильтрационного питания использовали гребни с многочисленными игольчатыми зубцами; У Птеродаустро могло быть более тысячи щетинообразных зубов. Dsungaripteridae покрывали свои зубы тканью челюстной кости для выполнения дробящей функции. Если зубы присутствовали, их помещали в отдельные лунки. [26] Замещающие зубы были созданы позади, а не ниже старых зубов. [25]

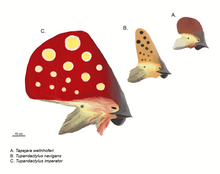
Общественный имидж птерозавров определяется их сложными гребнями на голове. [30] На это повлиял характерный обращенный назад гребень известного птеранодона . Основными положениями таких гребней являются передняя часть морды, как вырост предчелюстных костей, или задняя часть черепа, как продолжение теменных костей , и в этом случае его называют «надзатылочным гребнем». [28] Передние и задние гребни могут присутствовать одновременно и могут быть слиты в одну более крупную структуру, наиболее обширная из которых представлена Tapejaridae . Никтозавр имел причудливый гребень, похожий на рог. Гребни были толщиной всего несколько миллиметров в поперечном направлении. Основание костного гребня обычно расширено кератином или другой мягкой тканью. [28]
С 1990-х годов новые открытия и более тщательное изучение старых экземпляров показали, что гребни гораздо более распространены среди птерозавров, чем предполагалось ранее. То, что они были расширены или полностью состоят из кератина, который нелегко окаменевает, ввело в заблуждение более ранние исследования. [31] Для Pterorhynchus и Pterodactylus истинная протяженность этих гребней была обнаружена только с помощью ультрафиолетовой фотографии. [29] [32] В то время как ископаемые гребни раньше были ограничены более развитыми Pterodactyloidea, Pterorhynchus и Austriadactylus показывают, что они принадлежали даже некоторым ранним птерозаврам. [31]
Как и верхние челюсти, парные нижние челюсти птерозавров были сильно удлинены. [33] У продвинутых форм они, как правило, были короче верхней части черепа, поскольку челюстной сустав находился в более переднем положении. Передние кости нижней челюсти, dentaries или ossa dentalia , на кончике плотно срослись с центральным симфизом. Это позволило нижним челюстям функционировать как единое целое – нижняя челюсть . Симфиз часто был очень тонким в поперечном направлении и длинным, составляя значительную часть длины челюсти, до 60%. [27] Если на морде присутствовал гребень, симфиз мог иметь соответствующий гребень нижней челюсти, выступающий вниз. [27] Зубатые виды также имели зубы в зубных рядах. Нижняя челюсть открывалась и закрывалась простым вертикальным или «ортальным» движением вверх и вниз.
Позвоночник
[ редактировать ]
Позвоночный столб птерозавров насчитывал от тридцати четырех до семидесяти позвонков . Позвонки перед хвостом были «процелозными»: семядоля (передняя часть тела позвонка ) была вогнутой и в нее вписывался выпуклый выступ в задней части предыдущего позвонка — мыщелок . Развитые птерозавры уникальны тем, что обладают особыми отростками, выступающими рядом с их мыщелком и семядолей, эксапофизами . [34] и семядоля также может иметь небольшой зубец на средней линии, называемый гипапофизом. [35]

Шеи птерозавров были относительно длинными и прямыми. У птеродактилоидов шея обычно длиннее туловища. [36] Эта длина не вызвана увеличением числа позвонков, которых неизменно семь. Некоторые исследователи включают два переходных «цервидорсала», в результате чего их число достигает девяти. [36] Вместо этого сами позвонки стали более вытянутыми, в восемь раз длиннее своей ширины. Тем не менее, шейные позвонки были шире, чем высота, что подразумевало лучшую вертикальную, чем горизонтальную подвижность шеи. Птеродактилоиды потеряли все шейные ребра. [35] Шеи птерозавров, вероятно, были довольно толстыми и мускулистыми. [37] особенно вертикально. [38]
Туловище было относительно коротким и яйцеобразным. Первоначально количество позвонков на спине птерозавров могло насчитывать восемнадцать. У продвинутых видов все большее их число имело тенденцию располагаться в крестце . У таких видов также часто наблюдается слияние передних спинных позвонков в жесткое целое, которое называется нотариумом по сходному строению у птиц. Это была адаптация, позволяющая противостоять силам, вызванным взмахами крыльев. [36] Нотариум включал от трех до семи позвонков, в зависимости от вида, а также от возраста человека. Эти позвонки могли быть соединены сухожилиями или слиянием их нервных отростков в «надневральную пластинку». Их ребра также будут плотно сращены с нотарием. [39] Обычно ребра двуглавые. [40] Крестец состоял из трех-десяти крестцовых позвонков. Их тоже можно было соединить через надневральную пластину, которая, однако, не будет контактировать с нотариусом. [39]

Хвосты птерозавров всегда были довольно тонкими. Это означает, что мышца-ретрактор caudofemoralis , которая у большинства базальных архозаврий обеспечивает основную движущую силу задней конечности, была относительно неважной. [38] Хвостовые позвонки амфицельные, тела позвонков на обоих концах вогнутые. Ранние виды имели длинные хвосты, содержащие до пятидесяти хвостовых позвонков, средние из которых укреплялись удлиненными сочленяющимися отростками, зигапофизами и шевронами . [41] Такие хвосты выполняли роль рулей направления, иногда заканчиваясь сзади вертикальным ромбовидным или овальным флюгером. [42] У птеродактилоидов хвосты были сильно редуцированы и никогда не затвердевали. [42] у некоторых видов насчитывается всего десять позвонков. [39]
Shoulder girdle
[edit]The shoulder girdle was a strong structure that transferred the forces of flapping flight to the thorax. It was probably covered by thick muscle layers.[43] The upper bone, the shoulder blade, was a straight bar. It was connected to a lower bone, the coracoid that is relatively long in pterosaurs. In advanced species, their combined whole, the scapulocoracoid, was almost vertically oriented. The shoulder blade in that case fitted into a recess in the side of the notarium, while the coracoid likewise connected to the breastbone. This way, both sides together made for a rigid closed loop, able to withstand considerable forces.[40] A peculiarity was that the breastbone connections of the coracoids often were asymmetrical, with one coracoid attached in front of the other. In advanced species the shoulder joint had moved from the shoulder blade to the coracoid.[44] The joint was saddle-shaped and allowed considerable movement to the wing.[40] It faced sideways and somewhat upwards.[42]
The breastbone, formed by fused paired sterna, was wide. It had only a shallow keel. Via sternal ribs, it was at its sides attached to the dorsal ribs.[41] At its rear, a row of belly ribs or gastralia was present, covering the entire belly.[42] To the front, a long point, the cristospina, jutted obliquely upwards. The rear edge of the breastbone was the deepest point of the thorax.[44] Clavicles or interclavicles were completely absent.[42]
Wings
[edit]
Pterosaur wings were formed by bones and membranes of skin and other tissues. The primary membranes attached to the extremely long fourth finger of each arm and extended along the sides of the body. Where they ended has been very controversial but since the 1990s a dozen specimens with preserved soft tissue have been found that seem to show they attached to the ankles. The exact curvature of the trailing edge, however, is still equivocal.[45]

While historically thought of as simple leathery structures composed of skin, research has since shown that the wing membranes of pterosaurs were highly complex dynamic structures suited to an active style of flight.[46] The outer wings (from the tip to the elbow) were strengthened by closely spaced fibers called actinofibrils.[47] The actinofibrils themselves consisted of three distinct layers in the wing, forming a crisscross pattern when superimposed on one another. The function of the actinofibrils is unknown, as is the exact material from which they were made. Depending on their exact composition (keratin, muscle, elastic structures, etc.), they may have been stiffening or strengthening agents in the outer part of the wing.[48] The wing membranes also contained a thin layer of muscle, fibrous tissue, and a unique, complex circulatory system of looping blood vessels.[31] The combination of actinofibrils and muscle layers may have allowed the animal to adjust the wing slackness and camber.[46]
As shown by cavities in the wing bones of larger species and soft tissue preserved in at least one specimen, some pterosaurs extended their system of respiratory air sacs into the wing membrane.[49]
Parts of the wing
[edit]
The pterosaur wing membrane is divided into three basic units.[50] The first, called the propatagium ("fore membrane"), was the forward-most part of the wing and attached between the wrist and shoulder, creating the "leading edge" during flight. The brachiopatagium ("arm membrane") was the primary component of the wing, stretching from the highly elongated fourth finger of the hand to the hindlimbs. Finally, at least some pterosaur groups had a membrane that stretched between the legs, possibly connecting to or incorporating the tail, called the uropatagium;[50] the extent of this membrane is not certain, as studies on Sordes seem to suggest that it simply connected the legs but did not involve the tail (rendering it a cruropatagium). A common interpretation is that non-pterodactyloid pterosaurs had a broader uro/cruropatagium stretched between their long fifth toes, with pterodactyloids, lacking such toes, only having membranes running along the legs.[51]
There has been considerable argument among paleontologists about whether the main wing membranes (brachiopatagia) attached to the hindlimbs, and if so, where. Fossils of the rhamphorhynchoid Sordes,[52] the anurognathid Jeholopterus,[53] and a pterodactyloid from the Santana Formation seem to demonstrate that the wing membrane did attach to the hindlimbs, at least in some species.[54] However, modern bats and flying squirrels show considerable variation in the extent of their wing membranes and it is possible that, like these groups, different species of pterosaur had different wing designs. Indeed, analysis of pterosaur limb proportions shows that there was considerable variation, possibly reflecting a variety of wing-plans.[55]
The bony elements of the arm formed a mechanism to support and extend the wing. Near the body, the humerus or upper arm bone is short but powerfully built.[56] It sports a large deltopectoral crest, to which the major flight muscles are attached.[56] Despite the considerable forces exerted on it, the humerus is hollow or pneumatised inside, reinforced by bone struts.[44] The long bones of the lower arm, the ulna and radius, are much longer than the humerus.[57] They were probably incapable of pronation.
A bone unique to pterosaurs,[58] known as the pteroid, connected to the wrist and helped to support the forward membrane (the propatagium) between the wrist and shoulder. Evidence of webbing between the three free fingers of the pterosaur forelimb suggests that this forward membrane may have been more extensive than the simple pteroid-to-shoulder connection traditionally depicted in life restorations.[31] The position of the pteroid bone itself has been controversial. Some scientists, notably Matthew Wilkinson, have argued that the pteroid pointed forward, extending the forward membrane and allowing it to function as an adjustable flap.[59] This view was contradicted in a 2007 paper by Chris Bennett, who showed that the pteroid did not articulate as previously thought and could not have pointed forward, but rather was directed inward toward the body as traditionally interpreted.[60] Specimens of Changchengopterus pani and Darwinopterus linglongtaensis show the pteroid in articulation with the proximal syncarpal, suggesting that the pteroid articulated with the 'saddle' of the radiale (proximal syncarpal) and that both the pteroid and preaxial carpal were migrated centralia.[61][62]
The pterosaur wrist consists of two inner (proximal, at the side of the long bones of the arm) and four outer (distal, at the side of the hand) carpals (wrist bones), excluding the pteroid bone, which may itself be a modified distal carpal. The proximal carpals are fused together into a "syncarpal" in mature specimens, while three of the distal carpals fuse to form a distal syncarpal. The remaining distal carpal, referred to here as the medial carpal, but which has also been termed the distal lateral, or pre-axial carpal, articulates on a vertically elongate biconvex facet on the anterior surface of the distal syncarpal. The medial carpal bears a deep concave fovea that opens anteriorly, ventrally and somewhat medially, within which the pteroid articulates, according to Wilkinson.[63]
In derived pterodactyloids like pteranodontians and azhdarchoids, metacarpals I-III are small and do not connect to the carpus, instead hanging in contact with the fourth metacarpal.[64] With these derived species, the fourth metacarpal has been enormously elongated, typically equalling or exceeding the length of the long bones of the lower arm.[65] The fifth metacarpal had been lost.[56] In all species, the first to third fingers are much smaller than the fourth, the "wingfinger", and contain two, three and four phalanges respectively.[64] The smaller fingers are clawed, with the ungual size varying among species. In nyctosaurids the forelimb digits besides the wingfinger have been lost altogether. The wingfinger accounts for about half or more of the total wing length.[64] It normally consists of four phalanges. Their relative lengths tend to vary among species, which has often been used to distinguish related forms.[64] The fourth phalanx is usually the shortest. It lacks a claw and has been lost completely by nyctosaurids. It is curved to behind, resulting in a rounded wing tip, which reduces induced drag. The wingfinger is also bent somewhat downwards.[65]
When standing, pterosaurs probably rested on their metacarpals, with the outer wing folded to behind. In this position, the "anterior" sides of the metacarpals were rotated to the rear. This would point the smaller fingers obliquely to behind. According to Bennett, this would imply that the wingfinger, able to describe the largest arc of any wing element, up to 175°, was not folded by flexion but by an extreme extension. The wing was automatically folded when the elbow was bowed.[38][66]
A laser-simulated fluorescence scan on Pterodactylus also identified a membranous "fairing" (area conjunctioning the wing with the body at the neck), as opposed to the feathered or fur-composed "fairing" seen in birds and bats respectively.[67]
Pelvis
[edit]
The pelvis of pterosaurs was of moderate size compared to the body as a whole. Often the three pelvic bones were fused.[65] The ilium was long and low, its front and rear blades projecting horizontally beyond the edges of the lower pelvic bones. Despite this length, the rod-like form of these processes indicates that the hindlimb muscles attached to them were limited in strength.[38] The, in side view narrow, pubic bone fused with the broad ischium into an ischiopubic blade. Sometimes, the blades of both sides were also fused, closing the pelvis from below and forming the pelvic canal. The hip joint was not perforated and allowed considerable mobility to the leg.[64] It was directed obliquely upwards, preventing a perfectly vertical position of the leg.[65]
The front of the pubic bones articulated with a unique structure, the paired prepubic bones. Together these formed a cusp covering the rear belly, between the pelvis and the belly ribs. The vertical mobility of this element suggests a function in breathing, compensating the relative rigidity of the chest cavity.[64]
Hindlimbs
[edit]The hindlimbs of pterosaurs were strongly built, yet relative to their wingspans smaller than those of birds. They were long in comparison to the torso length.[68] The thighbone was rather straight, with the head making only a small angle with the shaft.[64] This implies that the legs were not held vertically below the body but were somewhat sprawling.[68] The shinbone was often fused with the upper ankle bones into a tibiotarsus that was longer than the thighbone.[68] It could attain a vertical position when walking.[68] The calf bone tended to be slender, especially at its lower end that in advanced forms did not reach the ankle, sometimes reducing total length to a third. Typically, it was fused to the shinbone.[64] The ankle was a simple, "mesotarsal", hinge.[68] The, rather long and slender,[69] metatarsus was always splayed to some degree.[70] The foot was plantigrade, meaning that during the walking cycle the sole of the metatarsus was pressed onto the soil.[69]
There was a clear difference between early pterosaurs and advanced species regarding the form of the fifth digit. Originally, the fifth metatarsal was robust and not very shortened. It was connected to the ankle in a higher position than the other metatarsals.[69] It bore a long, and often curved, mobile clawless fifth toe consisting of two phalanges.[70] The function of this element has been enigmatic. It used to be thought that the animals slept upside-down like bats, hanging from branches and using the fifth toes as hooks. Another hypothesis held that they stretched the brachiopatagia, but in articulated fossils the fifth digits are always flexed towards the tail.[69] Later it became popular to assume that these toes extended an uropatagium or cruropatagium between them. As the fifth toes were on the outside of the feet, such a configuration would only have been possible if these rotated their fronts outwards in flight.[69] Such a rotation could be caused by an abduction of the thighbone, meaning that the legs would be spread. This would also turn the feet into a vertical position.[69] They then could act as rudders to control yaw. Some specimens show membranes between the toes,[71] allowing them to function as flight control surfaces. The uropatagium or cruropatagium would control pitch. When walking the toes could flex upwards to lift the membrane from the ground. In Pterodactyloidea, the fifth metatarsal was much reduced and the fifth toe, if present, little more than a stub.[72] This suggests that their membranes were split, increasing flight maneuverability.[51]
The first to fourth toes were long. They had two, three, four and five phalanges respectively.[68] Often the third toe was longest; sometimes the fourth. Flat joints indicate a limited mobility. These toes were clawed but the claws were smaller than the hand claws.[70]
Soft tissues
[edit]The rare conditions that allowed for the fossilisation of pterosaur remains, sometimes also preserved soft tissues. Modern synchrotron or ultraviolet light photography has revealed many traces not visible to the naked eye.[73] These are often imprecisely called "impressions" but mostly consist of petrifications, natural casts and transformations of the original material. They may include horn crests, beaks or claw sheaths as well as the various flight membranes. Exceptionally, muscles were preserved.[74] Skin patches show small round non-overlapping scales on the soles of the feet, the ankles and the ends of the metatarsals.[75] They covered pads cushioning the impact of walking. Scales are unknown from other parts of the body.[76]
Pycnofibers
[edit]
Most or all pterosaurs had hair-like filaments known as pycnofibers on the head and torso.[77] The term "pycnofiber", meaning "dense filament", was coined by palaeontologist Alexander Kellner and colleagues in 2009.[48] Pycnofibers were unique structures similar to, but not homologous (sharing a common origin) with, mammalian hair, an example of convergent evolution.[52] A fuzzy integument was first reported from a specimen of Scaphognathus crassirostris in 1831 by Georg August Goldfuss,[78] but had been widely doubted. Since the 1990s, pterosaur finds and histological and ultraviolet examination of pterosaur specimens have provided incontrovertible proof: pterosaurs had pycnofiber coats. Sordes pilosus (which translates as "hairy demon") and Jeholopterus ninchengensis show pycnofibers on the head and body.

The presence of pycnofibers strongly indicates that pterosaurs were endothermic (warm-blooded). They aided thermoregulation, as is common in warm-blooded animals who need insulation to prevent excessive heat-loss.[77] Pycnofibers were flexible, short filaments, about five to seven millimetres long and rather simple in structure with a hollow central canal.[77] Pterosaur pelts might have been comparable in density to many Mesozoic mammals.[b][77]
Relation with feathers
[edit]Pterosaur filaments could share a common origin with feathers, as speculated in 2002 by Czerkas and Ji.[32] In 2009, Kellner concluded that pycnofibers were structured similarly to theropod proto-feathers.[48] Others were unconvinced, considering the difference with the "quills" found on many of the bird-like maniraptoran specimens too fundamental.[77]
A 2018 study of the remains of two small Jurassic-age pterosaurs from Inner Mongolia, China, found that pterosaurs had a wide array of pycnofiber shapes and structures, as opposed to the homogeneous structures that had generally been assumed to cover them. Some of these had frayed ends, very similar in structure to four different feather types known from birds or other dinosaurs but almost never known from pterosaurs prior to the study, suggesting homology.[79][80] A response to this study was published in 2020, where it was suggested that the structures seen on the anurognathids were actually a result of the decomposition of aktinofibrils: a type of fibre used to strengthen and stiffen the wing.[81] However, in a response to this, the authors of the 2018 paper point to the fact that the presence of the structures extend past the patagium, and the presence of both aktinofibrils and filaments on Jeholopterus ningchengensis[82] and Sordes pilosus.[83] The various forms of filament structure present on the anurognathids in the 2018 study would also require a form of decomposition that would cause the different 'filament' forms seen. They therefore conclude that the most parsimonious interpretation of the structures is that they are filamentous protofeathers.[84] But Liliana D'Alba points out that the description of the preserved integumentary structures on the two anurognathid specimens is still based upon gross morphology. She also points out that Pterorhynchus was described to have feathers to support the claim that feathers had a common origin with Ornithodirans but was argued against by several authors. The only method to assure if it was homologous to feathers is to use a scanning electron microscope.[85]
In 2022, a new fossil of Tupandactylus cf. imperator[86] was found to have melanosomes in forms that signal an earlier than anticipated development of the patterns found in extant feathers than previously thought. In these fossils, it appears as though the feather melanosomes took on a more complex form than the melanosome organization in scales that near relatives of Tupandactylus had. This discovery is one of many that leads us away from many previous theories of feathers evolving directly from scales in reptiles, given the significant distinction of melanosome organization and content between the two. This indicates a distinct form of melanosomes within feather structures at the time, different from other contemporary feathers that did not carry this formation. The feather fossils obtained from this specimen also suggested the presence of Stage IIIa feathers, a new discovery which may also suggest that more complex feather structures were present at this time. Previously, no Stage III feather forms had been discovered in this time. This study contains multiple indications about the development of feather forms. These include a more precise estimate for the development of avian feather forms, as well as a more ancient ancestor that contained the origins of feather-specific melanosome signaling found in extant birds.
History of discovery
[edit]First finds
[edit]
Pterosaur fossils are very rare, due to their light bone construction. Complete skeletons can generally only be found in geological layers with exceptional preservation conditions, the so-called Lagerstätten. The pieces from one such Lagerstätte, the Late Jurassic Solnhofen Limestone in Bavaria,[87] became much sought after by rich collectors.[88] In 1784, Italian naturalist Cosimo Alessandro Collini was the first scientist to describe a pterosaur fossil.[89] At that time the concepts of evolution and extinction were imperfectly developed. The bizarre build of the pterosaur was shocking, as it could not clearly be assigned to any existing animal group.[90] The discovery of pterosaurs would thus play an important role in the progress of modern paleontology and geology.[91] Scientific opinion at the time was that if such creatures were still alive, only the sea was a credible habitat; Collini suggested it might be a swimming animal that used its long front limbs as paddles.[92] A few scientists continued to support the aquatic interpretation even until 1830, when German zoologist Johann Georg Wagler suggested that Pterodactylus used its wings as flippers and was affiliated with Ichthyosauria and Plesiosauria.[93]

In 1800, Johann Hermann first suggested that it represented a flying creature in a letter to Georges Cuvier. Cuvier agreed in 1801, understanding it was an extinct flying reptile.[94] In 1809, he coined the name Ptéro-Dactyle, "wing-finger".[95] This was in 1815 Latinised to Pterodactylus.[96] At first most species were assigned to this genus and ultimately "pterodactyl" was popularly and incorrectly applied to all members of Pterosauria.[16] Today, paleontologists limit the term to the genus Pterodactylus or members of the Pterodactyloidea.[17]
In 1812 and 1817, Samuel Thomas von Soemmerring redescribed the original specimen and an additional one.[97] He saw them as affiliated to birds and bats. Although he was mistaken in this, his "bat model" would be influential during the 19th century.[98] In 1843, Edward Newman thought pterosaurs were flying marsupials.[99] Ironically, as the "bat model" depicted pterosaurs as warm-blooded and furred, it would turn out to be more correct in certain aspects than Cuvier's "reptile model" in the long run. In 1834, Johann Jakob Kaup coined the term Pterosauria.[100]
Expanding research
[edit]
In 1828, Mary Anning found in England the first pterosaur genus outside Germany,[101] named as Dimorphodon by Richard Owen, also the first non-pterodactyloid pterosaur known.[102] Later in the century, the Early Cretaceous Cambridge Greensand produced thousands of pterosaur fossils, that however, were of poor quality, consisting mostly of strongly eroded fragments.[103] Nevertheless, based on these, numerous genera and species would be named.[91] Many were described by Harry Govier Seeley, at the time the main English expert on the subject, who also wrote the first pterosaur book, Ornithosauria,[104] and in 1901 the first popular book,[91] Dragons of the Air. Seeley thought that pterosaurs were warm-blooded and dynamic creatures, closely related to birds.[105] Earlier, the evolutionist St. George Jackson Mivart had suggested pterosaurs were the direct ancestors of birds.[106] Owen opposed the views of both men, seeing pterosaurs as cold-blooded "true" reptiles.[107]
In the US, Othniel Charles Marsh in 1870 discovered Pteranodon in the Niobrara Chalk, then the largest known pterosaur,[107] the first toothless one and the first from America.[108] These layers too rendered thousands of fossils,[108] also including relatively complete skeletons that were three-dimensionally preserved instead of being strongly compressed as with the Solnhofen specimens. This led to a much better understanding of many anatomical details,[108] such as the hollow nature of the bones.
Meanwhile, finds from the Solnhofen had continued, accounting for the majority of complete high-quality specimens discovered.[109] They allowed to identify most new basal taxa, such as Rhamphorhynchus, Scaphognathus and Dorygnathus.[109] This material gave birth to a German school of pterosaur research, which saw flying reptiles as the warm-blooded, furry and active Mesozoic counterparts of modern bats and birds.[110] In 1882, Marsh and Karl Alfred Zittel published studies about the wing membranes of specimens of Rhamphorhynchus.[111][112] German studies continued well into the 1930s, describing new species such as Anurognathus. In 1927, Ferdinand Broili discovered hair follicles in pterosaur skin,[113] and paleoneurologist Tilly Edinger determined that the brains of pterosaurs more resembled those of birds than modern cold-blooded reptiles.[114]
In contrast, English and American paleontologists by the middle of the twentieth century largely lost interest in pterosaurs. They saw them as failed evolutionary experiments, cold-blooded and scaly, that hardly could fly, the larger species only able to glide, being forced to climb trees or throw themselves from cliffs to achieve a take-off. In 1914, for the first-time pterosaur aerodynamics were quantitatively analysed, by Ernest Hanbury Hankin and David Meredith Seares Watson, but they interpreted Pteranodon as a pure glider.[115] Little research was done on the group during the 1940s and 1950s.[91]
Pterosaur renaissance
[edit]
The situation for dinosaurs was comparable. From the 1960s onwards, a dinosaur renaissance took place, a quick increase in the number of studies and critical ideas, influenced by the discovery of additional fossils of Deinonychus, whose spectacular traits refuted what had become entrenched orthodoxy. In 1970, likewise the description of the furry pterosaur Sordes began what Robert Bakker named a renaissance of pterosaurs.[116] Kevin Padian especially propagated the new views, publishing a series of studies depicting pterosaurs as warm-blooded, active and running animals.[117][118][119] This coincided with a revival of the German school through the work of Peter Wellnhofer, who in 1970s laid the foundations of modern pterosaur science.[87] In 1978, he published the first pterosaur textbook,[120] the Handbuch der Paläoherptologie, Teil 19: Pterosauria,[121] and in 1991 the second ever popular science pterosaur book,[120] the Encyclopedia of Pterosaurs.[122]

This development accelerated through the exploitation of two new Lagerstätten.[120] During the 1970s, the Early Cretaceous Santana Formation in Brazil began to produce chalk nodules that, though often limited in size and the completeness of the fossils they contained, perfectly preserved three-dimensional pterosaur skeletal parts.[120] German and Dutch institutes bought such nodules from fossil poachers and prepared them in Europe, allowing their scientists to describe many new species and revealing a whole new fauna. Soon, Brazilian researchers, among them Alexander Kellner, intercepted the trade and named even more species.
Even more productive was the Early Cretaceous Chinese Jehol Biota of Liaoning that since the 1990s has brought forth hundreds of exquisitely preserved two-dimensional fossils, often showing soft tissue remains. Chinese researchers such as Lü Junchang have again named many new taxa. As discoveries also increased in other parts of the world, a sudden surge in the total of named genera took place. By 2009, when they had increased to about ninety, this growth showed no sign of levelling-off.[123] In 2013, M.P. Witton indicated that the number of discovered pterosaur species had risen to 130.[124] Over ninety percent of known taxa has been named during the "renaissance". Many of these were from groups the existence of which had been unknown.[120] Advances in computing power enabled researchers to determine their complex relationships through the quantitative method of cladistics. New and old fossils yielded much more information when subjected to modern ultraviolet light or roentgen photography, or CAT-scans.[125] Insights from other fields of biology were applied to the data obtained.[125] All this resulted in a substantial progress in pterosaur research, rendering older accounts in popular science books completely outdated.
In 2017 a fossil from a 170-million-year-old pterosaur, later named as the species Dearc sgiathanach in 2022, was discovered on the Isle of Skye in Scotland. The National Museum of Scotland claims that it is the largest of its kind ever discovered from the Jurassic period, and it has been described as the world's best-preserved skeleton of a pterosaur.[126]
Evolution and extinction
[edit]Origins
[edit]
Because pterosaur anatomy has been so heavily modified for flight, and immediate transitional fossil predecessors have not so far been described, the ancestry of pterosaurs is not fully understood.[127] The oldest known pterosaurs were already fully adapted to a flying lifestyle. Since Seeley, it was recognised that pterosaurs were likely to have had their origin in the "archosaurs", what today would be called the Archosauromorpha. In the 1980s, early cladistic analyses found that they were Avemetatarsalians (archosaurs closer to dinosaurs than to crocodilians). As this would make them also rather close relatives of the dinosaurs, these results were seen by Kevin Padian as confirming his interpretation of pterosaurs as bipedal warm-blooded animals. Because these early analyses were based on a limited number of taxa and characters, their results were inherently uncertain. Several influential researchers who rejected Padian's conclusions offered alternative hypotheses. David Unwin proposed an ancestry among the basal Archosauromorpha, specifically long-necked forms ("protorosaurs") such as tanystropheids. A placement among basal archosauriforms like Euparkeria was also suggested.[24] Some basal archosauromorphs seem at first glance to be good candidates for close pterosaur relatives due to their long-limbed anatomy; one example is Sharovipteryx, a "protorosaur" with skin membranes on its hindlimbs likely used for gliding.[128] A 1999 study by Michael Benton found that pterosaurs were avemetatarsalians closely related to Scleromochlus, and named the group Ornithodira to encompass pterosaurs and dinosaurs.[129]


Two researchers, S. Christopher Bennett in 1996,[130] and paleoartist David Peters in 2000, published analyses finding pterosaurs to be protorosaurs or closely related to them. However, Peters gathered novel anatomical data using an unverified technique called "Digital Graphic Segregation" (DGS), which involves digitally tracing over images of pterosaur fossils using photo editing software.[131] Bennett only recovered pterosaurs as close relatives of the protorosaurs after removing characteristics of the hindlimb from his analysis, to test the possibility of locomotion-based convergent evolution between pterosaurs and dinosaurs. A 2007 reply by Dave Hone and Michael Benton could not reproduce this result, finding pterosaurs to be closely related to dinosaurs even without hindlimb characters. They also criticized David Peters for drawing conclusions without access to the primary evidence, that is, the pterosaur fossils themselves.[132] Hone and Benton concluded that, although more basal pterosauromorphs are needed to clarify their relationships, current evidence indicates that pterosaurs are avemetatarsalians, as either the sister group of Scleromochlus or a branch between the latter and Lagosuchus.[132] An 2011 archosaur-focused phylogenetic analysis by Sterling Nesbitt benefited from far more data and found strong support for pterosaurs being avemetatarsalians, though Scleromochlus was not included due to its poor preservation.[133] A 2016 archosauromorph-focused study by Martin Ezcurra included various proposed pterosaur relatives, yet also found pterosaurs to be closer to dinosaurs and unrelated to more basal taxa.[134] Working from his 1996 analysis, Bennett published a 2020 study on Scleromochlus which argued that both Scleromochlus and pterosaurs were non-archosaur archosauromorphs, albeit not particularly closely related to each other.[135] By contrast, a later 2020 study proposed that lagerpetid archosaurs were the sister clade to pterosauria.[136] This was based on newly described fossil skulls and forelimbs showing various anatomical similarities with pterosaurs and reconstructions of lagerpetid brains and sensory systems based on CT scans also showing neuroanatomical similarities with pterosaurs.[137][138] The results of the latter study were subsequently supported by an independent analysis of early pterosauromorph interrelationships.[139]
A related problem is the origin of pterosaur flight.[140] Like with birds, hypotheses can be ordered into two main varieties: "ground up" or "tree down". Climbing a tree would cause height and gravity to provide both the energy and a strong selection pressure for incipient flight.[clarification needed] Rupert Wild in 1983 proposed a hypothetical "propterosaurus": a lizard-like arboreal animal developing a membrane between its limbs, first to safely parachute and then, gradually elongating the fourth finger, to glide.[141] However, subsequent cladistic results did not fit this model well. Neither protorosaurs nor ornithodirans are biologically equivalent to lizards. Furthermore, the transition between gliding and flapping flight is not well-understood. More recent studies on basal pterosaur hindlimb morphology seem to vindicate a connection to Scleromochlus. Like this archosaur, basal pterosaur lineages have plantigrade hindlimbs that show adaptations for saltation.[142]
At least one study found that the early Triassic ichnofossil Prorotodactylus is anatomically similar to that of early pterosaurs.[136]
Extinction
[edit]
It was once thought that competition with early bird species might have resulted in the extinction of many of the pterosaurs.[143] It was thought that by the end of the Cretaceous, only large species of pterosaurs were present (no longer true; see below). The smaller species were thought to have become extinct, their niche filled by birds.[144] However, pterosaur decline (if actually present) seems unrelated to bird diversity, as ecological overlap between the two groups appears to be minimal.[145] In fact, at least some avian niches were reclaimed by pterosaurs prior to the Cretaceous–Paleogene extinction event.[146] It seems that the K-Pg extinction event at the end of the Cretaceous, which wiped out all non-avian dinosaurs and many other animals, was the direct cause of the extinction of the pterosaurs.
In the early 2010s, several new pterosaur taxa were discovered dating to the Campanian/Maastrichtian, such as the ornithocheirids Piksi and "Ornithocheirus", possible pteranodontids and nyctosaurids, several tapejarids and the indeterminate non-azhdarchid Navajodactylus.[147][148] Small azhdarchoid pterosaurs were also present in the Campanian. This suggests that late Cretaceous pterosaur faunas were far more diverse than previously thought, possibly not even having declined significantly from the early Cretaceous.
Small-sized pterosaur species apparently were present in the Csehbánya Formation, indicating a higher diversity of Late Cretaceous pterosaurs than previously accounted for.[149] The recent findings of a small cat-sized adult azhdarchid further indicate that small pterosaurs from the Late Cretaceous might actually have simply been rarely preserved in the fossil record, helped by the fact that there is a strong bias against terrestrial small sized vertebrates such as juvenile dinosaurs, and that their diversity might actually have been much larger than previously thought.[150]
At least some non-pterodactyloid pterosaurs survived into the Late Cretaceous, postulating a Lazarus taxa situation for late Cretaceous pterosaur faunas.[151]
A 2021 study showcases that niches previously occupied by small pterosaurs were increasingly occupied by the juvenile stages of larger species in the Late Cretaceous. Rather than being outcompeted by birds, pterosaurs essentially specialized a trend already occurring in previous eras of the Mesozoic.[152]
Classification and phylogeny
[edit]In phylogenetic taxonomy, the clade Pterosauria has usually been defined as node-based and anchored to several extensively studied taxa as well as those thought to be primitive. One 2003 study defined Pterosauria as "The most recent common ancestor of the Anurognathidae, Preondactylus and Quetzalcoatlus and all their descendants."[153] However, these types of definition would inevitably leave any related species that are slightly more primitive out of the Pterosauria. To remedy this, a new definition was proposed that would anchor the name not to any particular species but to an anatomical feature, the presence of an enlarged fourth finger that supports a wing membrane.[154] This "apomorophy-based" definition was adopted by the PhyloCode in 2020 as "[T]he clade characterized by the apomorphy fourth manual digit hypertrophied to support a wing membrane, as inherited by Pterodactylus (originally Ornithocephalus) antiquus (Sömmerring 1812)".[155] A broader clade, Pterosauromorpha, has been defined as all ornithodirans more closely related to pterosaurs than to dinosaurs.[156]
The internal classification of pterosaurs has historically been difficult, because there were many gaps in the fossil record. Starting from the 21st century, new discoveries are now filling in these gaps and giving a better picture of the evolution of pterosaurs. Traditionally, they were organized into two suborders: the Rhamphorhynchoidea, a "primitive" group of long-tailed pterosaurs, and the Pterodactyloidea, "advanced" pterosaurs with short tails.[24] However, this traditional division has been largely abandoned. Rhamphorhynchoidea is a paraphyletic (unnatural) group, since the pterodactyloids evolved directly from them and not from a common ancestor, so, with the increasing use of cladistics, it has fallen out of favor among most scientists.[124][157]
The precise relationships between pterosaurs is still unsettled. Many studies of pterosaur relationships in the past have included limited data and were highly contradictory. However, newer studies using larger data sets are beginning to make things clearer. The cladogram (family tree) below follows a phylogenetic analysis presented by Longrich, Martill and Andres in 2018, with clade names after Andres et al. (2014).[146][1]
| Pterosauria | |
Paleobiology
[edit]

Flight
[edit]The mechanics of pterosaur flight are not completely understood or modeled at this time.[158][159][needs update]
Katsufumi Sato, a Japanese scientist, did calculations using modern birds and concluded that it was impossible for a pterosaur to stay aloft.[158] In the book Posture, Locomotion, and Paleoecology of Pterosaurs it is theorized that they were able to fly due to the oxygen-rich, dense atmosphere of the Late Cretaceous period.[160] However, both Sato and the authors of Posture, Locomotion, and Paleoecology of Pterosaurs based their research on the now-outdated theories of pterosaurs being seabird-like, and the size limit does not apply to terrestrial pterosaurs, such as azhdarchids and tapejarids. Furthermore, Darren Naish concluded that atmospheric differences between the present and the Mesozoic were not needed for the giant size of pterosaurs.[161]

Another issue that has been difficult to understand is how they took off. Earlier suggestions were that pterosaurs were largely cold-blooded gliding animals, deriving warmth from the environment like modern lizards, rather than burning calories. In this case, it was unclear how the larger ones of enormous size, with an inefficient cold-blooded metabolism, could manage a bird-like takeoff strategy, using only the hind limbs to generate thrust for getting airborne. Later research shows them instead as being warm-blooded and having powerful flight muscles, and using the flight muscles for walking as quadrupeds.[162] Mark Witton of the University of Portsmouth and Mike Habib of Johns Hopkins University suggested that pterosaurs used a vaulting mechanism to obtain flight.[163] The tremendous power of their winged forelimbs would enable them to take off with ease.[162] Once aloft, pterosaurs could reach speeds of up to 120 km/h (75 mph) and travel thousands of kilometres.[163]
In 1985, the Smithsonian Institution commissioned aeronautical engineer Paul MacCready to build a half-scale working model of Quetzalcoatlus northropi. The replica was launched with a ground-based winch. It flew several times in 1986 and was filmed as part of the Smithsonian's IMAX film On the Wing.[164][165]
Large-headed species are thought to have forwardly swept their wings in order to better balance.[166]
Air sacs and respiration
[edit]A 2009 study showed that pterosaurs had a lung-and-air-sac system and a precisely controlled skeletal breathing pump, which supports a flow-through pulmonary ventilation model in pterosaurs, analogous to that of birds. The presence of a subcutaneous air sac system in at least some pterodactyloids would have further reduced the density of the living animal.[49] Like modern crocodilians, pterosaurs appeared to have had a hepatic piston, seeing as their shoulder-pectoral girdles were too inflexible to move the sternum as in birds, and they possessed strong gastralia.[167] Thus, their respiratory system had characteristics comparable to both modern archosaur clades.
Nervous system
[edit]An X-ray study of pterosaur brain cavities revealed that the animals (Rhamphorhynchus muensteri and Anhanguera santanae) had massive flocculi. The flocculus is a brain region that integrates signals from joints, muscles, skin and balance organs.[18] The pterosaurs' flocculi occupied 7.5% of the animals' total brain mass, more than in any other vertebrate. Birds have unusually large flocculi compared with other animals, but these only occupy between 1 and 2% of total brain mass.[18]
The flocculus sends out neural signals that produce small, automatic movements in the eye muscles. These keep the image on an animal's retina steady. Pterosaurs may have had such a large flocculus because of their large wing size, which would mean that there was a great deal more sensory information to process.[18] The low relative mass of the flocculi in birds is also a result of birds having a much larger brain overall; though this has been considered an indication that pterosaurs lived in a structurally simpler environment or had less complex behaviour compared to birds,[168] recent studies of crocodilians and other reptiles show that it is common for sauropsids to achieve high intelligence levels with small brains.[169] Studies on the endocast of Allkaruen show that brain evolution in pterodactyloids was a modular process.[170]
Ground movement
[edit]
Pterosaurs' hip sockets are oriented facing slightly upwards, and the head of the femur (thigh bone) is only moderately inward facing, suggesting that pterosaurs had an erect stance. It would have been possible to lift the thigh into a horizontal position during flight, as gliding lizards do.
There was considerable debate whether pterosaurs ambulated as quadrupeds or as bipeds. In the 1980s, paleontologist Kevin Padian suggested that smaller pterosaurs with longer hindlimbs, such as Dimorphodon, might have walked or even run bipedally, in addition to flying, like road runners.[119] However, a large number of pterosaur trackways were later found with a distinctive four-toed hind foot and three-toed front foot; these are the unmistakable prints of pterosaurs walking on all fours.[171][172]
Fossil footprints show that pterosaurs stood with the entire foot in contact with the ground (plantigrade), in a manner similar to many mammals like humans and bears. Footprints from azhdarchids and several unidentified species show that pterosaurs walked with an erect posture with their four limbs held almost vertically beneath the body, an energy-efficient stance used by most modern birds and mammals, rather than the sprawled limbs of modern reptiles.[71][162] Indeed, erect-limbs may be omnipresent in pterosaurs.[142]

Though traditionally depicted as ungainly and awkward when on the ground, the anatomy of some pterosaurs (particularly pterodactyloids) suggests that they were competent walkers and runners.[173] Early pterosaurs have long been considered particularly cumbersome locomotors due to the presence of large cruropatagia, but they too appear to have been generally efficient on the ground.[142]
The forelimb bones of azhdarchids and ornithocheirids were unusually long compared to other pterosaurs, and, in azhdarchids, the bones of the arm and hand (metacarpals) were particularly elongated. Furthermore, as a whole, azhdarchid front limbs were proportioned similarly to fast-running ungulate mammals. Their hind limbs, on the other hand, were not built for speed, but they were long compared with most pterosaurs, and allowed for a long stride length. While azhdarchid pterosaurs probably could not run, they would have been relatively fast and energy efficient.[71]
The relative size of the hands and feet in pterosaurs (by comparison with modern animals such as birds) may indicate the type of lifestyle pterosaurs led on the ground. Azhdarchid pterosaurs had relatively small feet compared to their body size and leg length, with foot length only about 25–30% the length of the lower leg. This suggests that azhdarchids were better adapted to walking on dry, relatively solid ground. Pteranodon had slightly larger feet (47% the length of the tibia), while filter-feeding pterosaurs like the ctenochasmatoids had very large feet (69% of tibial length in Pterodactylus, 84% in Pterodaustro), adapted to walking in soft muddy soil, similar to modern wading birds.[71] Though clearly forelimb-based launchers, basal pterosaurs have hindlimbs well adapted for hopping, suggesting a connection with archosaurs such as Scleromochlus.[142]
Swimming
[edit]Следы, оставленные гребневиками, указывают на то, что эти птерозавры плавали, используя задние конечности. В целом у них большие задние лапы и длинное туловище, что указывает на то, что они, вероятно, были более приспособлены к плаванию, чем другие птерозавры. [174] У птеранодонтов, напротив, есть несколько видов плечевых костей, которые интерпретируются как наводящие на мысль о водной версии типичного четвероногого запуска, а некоторые подобные бореоптеридам, должно быть, добывали пищу во время плавания, поскольку они, похоже, неспособны к воздушному ястребу, подобно фрегатам . [174] Эти адаптации наблюдаются и у наземных птерозавров, таких как аждархиды , которым, по-видимому, все еще нужно было запускаться из воды на случай, если они в ней окажутся. Никтозаврид олуши Alcione может иметь приспособления для ныряния с использованием крыльев, как современные и тропические птицы . [146]
Диета и пищевые привычки
[ редактировать ]Традиционно почти всех птерозавров считали рыбоядными или рыбоядными, питающимися на поверхности, и эта точка зрения до сих пор доминирует в популярной науке. Сегодня считается, что многие группы птерозавров были наземными хищниками, всеядными или насекомоядными.
Вначале было признано, что мелкие Anurognathidae были ночными воздушными насекомоядными. Обладая очень гибкими суставами на пальце крыла, широкой треугольной формой крыла, большими глазами и коротким хвостом, эти птерозавры, вероятно, были аналогом козодоев или современных насекомоядных летучих мышей, будучи способными к высокой маневренности на относительно низких скоростях. [175]
Интерпретации привычек базальных групп глубоко изменились. Диморфодон , который в прошлом считался аналогом тупика , отличается строением челюстей, походкой и плохими способностями к полету как наземный/полудревесный хищник мелких млекопитающих, чешуйчатых и крупных насекомых. [176] Из-за мощного зубного ряда Campylognathoides считался универсалом или наземным хищником мелких позвоночных, но очень крепкая плечевая кость и морфология крыльев с большим удлинением позволяют предположить, что он, возможно, был способен хватать добычу на крыле. [177] Маленький насекомоядный Carniadactylus и более крупный Eudimorphodon были очень воздушными животными, быстрыми и маневренными летающими животными с длинными крепкими крыльями. Эвдиморфодон был обнаружен с остатками рыбы в желудке, но его зубной ряд предполагает оппортунистическую диету. Тонкокрылые Austriadactylus и Caviramus , вероятно, были наземными/полудревесными универсалами. Кавирамус , вероятно, имел сильную силу укуса, что указывает на адаптацию к твердой пище, которую можно было жевать из-за изнашивания зубов. [178]
Некоторые Rhamphorhynchidae , такие как сам Rhamphorhynchus или Dorygnathus , питались рыбой с длинными тонкими крыльями, игольчатыми зубными рядами и длинными тонкими челюстями. Sericipterus , Scaphognathus и Harpactognathus были зифодонтными, кинжаловидными имели более крепкие челюсти и зубы (которые у Sericipterus ), а также более короткие и широкие крылья. Это были либо наземные/воздушные хищники позвоночных животных. [179] или врановые универсалы. [180] Wukongopteridae , такие как Darwinopterus, сначала считались воздушными хищниками. Из-за отсутствия прочной структуры челюстей и мощных летательных мышц они теперь считаются древесными или полуназемными насекомоядными. В частности, Darwinopterusrobustidens , по-видимому, был специалистом по жукам. [181]
Среди птеродактилоидов наблюдается большее разнообразие рациона питания. Pteranodontia содержала множество таксонов рыбоядных, таких как Ornithocheirae , Boreopteridae , Pteranodontidae и Nyctosauridae. Разделение ниш привело к тому, что орнитохейраны и более поздние никтозавриды питались с воздуха, как современные фрегаты (за исключением адаптированного к нырянию Alcione elainus ), в то время как бореоптериды были пресноводными ныряющими животными, похожими на бакланов , а птеранодонты пелагическими ныряльщиками, похожими на олушей. и олуши . Анализ Lonchodraco обнаружил скопления отверстий на кончике его клюва; птицы со столь же многочисленными отверстиями имеют чувствительные клювы, используемые для нащупывания пищи, поэтому Лончодрако , возможно, использовал свой клюв, чтобы нащупывать рыбу или беспозвоночных на мелководье. [182] Истиодактилиды , вероятно, были в первую очередь падальщиками. [183] Archaeopterodactyloidea добывали пищу в прибрежных или пресноводных местообитаниях. Germanodactylus и Pterodactylus были рыбоядными животными, а Ctenochasmatidae питались взвешенными веществами, используя свои многочисленные мелкие зубы для фильтрации мелких организмов с мелководья. Птеродаустро был приспособлен для фламинго . фильтрационного питания, подобно [184]
Напротив, Azhdarchoidea в основном были наземными птерозаврами. Tapejaridae были древесными всеядными животными, дополняя семена и плоды мелкими насекомыми и позвоночными. [174] [185] Dsungaripteridae были специализированными моллюскоядными животными, которые использовали свои мощные челюсти, чтобы раздавливать раковины моллюсков и ракообразных. Thalassodromidae, вероятно, были наземными хищниками. Сам Thalassodromeus был назван в честь метода ловли рыбы, известного как «обезжиренное кормление», который позже оказался биомеханически невозможным. Возможно, он преследовал относительно крупную добычу ввиду усиленных челюстных суставов и относительно высокой силы укуса. [186] Сейчас аждархиды считаются наземными хищниками, похожими на наземных птиц-носорогов или некоторых аистов , поедающих любую добычу, которую они могут проглотить целиком. [187] Хацегоптерикс был крепко сложенным хищником, охотившимся на относительно крупную добычу, включая динозавров среднего размера. [188] [189] Аланка, возможно, была специализированным моллюскоядным животным. [190]
Исследование 2021 года реконструировало приводящую мускулатуру черепов птеродактилоидов , оценив силу укуса и потенциальные пищевые привычки девяти выбранных видов. [191] Исследование подтвердило мнение о птеранодонтидах , никтозавридах и анхануэридах как о рыбоядных животных, основываясь на том, что они относительно слабы, но быстро кусаются, и позволяет предположить, что Tropeognathus mesembrinus специализировался на поедании относительно крупной добычи по сравнению с Anhanguera . Dsungaripterus Было подтверждено, что является дурофагом , а Thalassodromeus предложили разделить эту привычку питания на основе высоких расчетных коэффициентов силы укуса (BFQ) и абсолютных значений силы укуса. [191] Было подтверждено, что Tapejara wellnhoferi является специализированным потребителем твердого растительного материала с относительно высоким BFQ и высоким механическим преимуществом, а Caupedactylus ybaka и Tupuxuara leonardii были предложены как универсалы, питающиеся с земли, с промежуточными значениями силы укуса и менее специализированными челюстями. [191]
Естественные хищники
[ редактировать ]Известно, что птерозавры были съедены тероподами . В выпуске журнала Nature от 1 июля 2004 года палеонтолог Эрик Бюффето обсуждает раннемеловую окаменелость трёх шейных позвонков птерозавра со сломанным зубом спинозавра , скорее всего, Ирритатора , вмонтированным в него. Известно, что позвонки не были съедены и не подверглись перевариванию, поскольку суставы все еще сочленены. [192] На окаменелостях птеранодона были обнаружены следы зубов таких акул, как Скваликоракс. [193] , а окаменелость со следами зубов из формации Тулебук была интерпретирована как подвергшаяся нападению или уничтожению ихтиозавром ] (скорее всего, Platypterygius ).
Размножение и история жизни
[ редактировать ]
Хотя о размножении птерозавров известно очень мало, считается, что, как и все динозавры, все птерозавры размножались путем откладывания яиц, хотя такие находки очень редки. Первые известные яйца птерозавров были найдены в карьерах Ляонина, того же места, где добывались пернатые динозавры, и в Лома-дель-Птеродаустро ( формация Лагарсито , Аргентина ). Яйца из Ляонина были раздавлены, без признаков растрескивания, поэтому, очевидно, яйца имели кожистую скорлупу, как у современных ящериц. [194] Яйцо из формации Лагарсито было отложено птеродаустро . [195] [196] птерозавр, известный по обильному материалу. [197] Это было подтверждено описанием еще одного яйца птерозавра, принадлежащего к роду Darwinopterus , описанного в 2011 году, которое также имело кожистую оболочку и, как и современные рептилии, но в отличие от птиц, было довольно маленьким по сравнению с размером матери. [198] В 2014 году пять несплюснутых яиц вида Hamipterus tianshanensis были обнаружены в отложениях раннего мела на северо-западе Китая. Исследование скорлупы с помощью сканирующей электронной микроскопии показало наличие тонкого известкового слоя яичной скорлупы с мембраной под ней. [199] Исследование структуры и химического состава яичной скорлупы птерозавров, опубликованное в 2007 году, показало, что, скорее всего, птерозавры закапывали свои яйца, как современные крокодилы и черепахи . Захоронение яиц было бы полезно для ранней эволюции птерозавров, поскольку оно позволяет добиться большей адаптации к снижению веса, но этот метод размножения также наложил бы ограничения на разнообразие сред, в которых могли жить птерозавры, и, возможно, поставил бы их в невыгодное положение, когда они начали сталкиваться с экологической конкуренцией со стороны птиц . [200]
Образец Darwinopterus демонстрирует, что, по крайней мере, у некоторых птерозавров была пара функциональных яичников , в отличие от единственного функционального яичника у птиц, что отвергает уменьшение функциональных яичников как необходимое условие для полета с приводом. [201]
Крыльевые перепонки, сохранившиеся у эмбрионов птерозавров, хорошо развиты, что позволяет предположить, что птерозавры были готовы летать вскоре после рождения. [202] Однако томография окаменелых яиц Hamipterus позволяет предположить, что у молодых птерозавров были хорошо развиты бедренные кости для ходьбы, но слабая грудь для полета. [203] Неизвестно, справедливо ли это для других птерозавров. Были обнаружены окаменелости птерозавров возрастом всего от нескольких дней до недели (так называемые «взмахи»), представляющие несколько семейств птерозавров, включая птеродактилид, рамфоринхид, ктенохазматид и аждархид. [24] У всех сохранились кости, демонстрирующие относительно высокую степень затвердевания ( окостенения ) для своего возраста, а пропорции крыльев аналогичны взрослым особям. Фактически, многие птерозавры в прошлом считались взрослыми особями и относились к отдельным видам. Кроме того, птенцы обычно встречаются в тех же отложениях, что и взрослые особи, и молодые особи того же вида, например, птенцы Pterodactylus и Rhamphorhynchus, найденные в известняке Зольнхофен в Германии, и птенцы Pterodaustro из Аргентины. Все они обитают в глубоководной среде вдали от берега. [204]
Для большинства видов птерозавров неизвестно, практиковали ли они какую-либо форму родительской заботы, но их способность летать, как только они вышли из яйца, и многочисленные взмахи крыльев, обнаруженные в среде, далекой от гнезд и рядом со взрослыми особями, побудили большинство исследователей , в том числе Кристофер Беннетт и Дэвид Анвин, пришли к выводу, что птенцы зависели от своих родителей в течение относительно короткого периода времени, в период быстрого роста, когда крылья вырастали достаточно длинными, чтобы летать, а затем покидали гнездо, чтобы позаботиться о себе. , возможно, в течение нескольких дней после вылупления. [24] [205] Альтернативно, они могли использовать продукты из накопленного желтка для питания в течение первых нескольких дней жизни, как современные рептилии, вместо того, чтобы зависеть от родителей в питании. [204] Было показано, что в окаменелых гнездах Hamipterus сохранилось множество самцов и самок птерозавров вместе с их яйцами, что аналогично тому, как это происходит в современных морских птиц колониях . [199] [206] Из-за того, что грудь детенышей была недостаточно развита для полета, было высказано предположение, что Hamipterus , возможно, практиковал некоторую форму родительской заботы. [203] Однако с тех пор это исследование подверглось критике. [207] Большинство данных в настоящее время склоняются к тому, что вылупившиеся птерозавры являются сверхранними , подобно птицам- мегаподам , которые после вылупления летают, не нуждаясь в родительской заботе. Дальнейшее исследование сравнивает доказательства сверхраннего развития и «позднего полета» и в подавляющем большинстве случаев предполагает, что большинство, если не все, птерозавры были способны летать вскоре после вылупления. [208] Более позднее исследование показало, что, хотя птерозавры с меньшим телом, скорее всего, были сверхранними или ранними из-за постоянного или уменьшающегося соотношения сторон крыла во время роста, некоторые птерозавры с крупным телом, такие как птеранодон, продемонстрировали возможные доказательства того, что их молодые особи были альтрициальными , из-за После вылупления кости конечностей, ближайшие к телу, росли быстрее, чем любой другой элемент их скелета. Другими упомянутыми факторами были пределы яиц с мягкой скорлупой и размер тазового отверстия крупных самок птерозавров. [209] [210]
Темпы роста птерозавров после их вылупления различались в разных группах. У более примитивных длиннохвостых птерозавров (« рамфоринхоидов »), таких как Rhamphorhynchus , средняя скорость роста в течение первого года жизни составляла от 130% до 173%, что немного выше, чем скорость роста аллигаторов . потребовалось бы более трех лет, После половой зрелости рост этих видов замедлился, и Rhamphorhynchus чтобы достичь максимального размера. [205] Напротив, более продвинутые, крупные птеродактилоидные птерозавры, такие как птеранодон , вырастали до взрослых размеров в течение первого года жизни. Кроме того, птеродактилоиды имели детерминированный рост , то есть животные достигали фиксированного максимального размера взрослой особи и прекращали расти. [204]
Исследование 2021 года показывает, что молодые птерозавры более крупных видов все чаще берут на себя роли, ранее занимаемые взрослыми мелкими птерозаврами. [152]
Ежедневная активность
[ редактировать ]Сравнение склеральных колец птерозавров и современных птиц и рептилий было использовано для определения режима повседневной активности птерозавров. Роды птерозавров Pterodactylus , Scaphognathus и Tupuxuara, как предполагается, ведут дневной образ жизни , Ctenochasma , Pterodaustro и Rhamphorhynchus — как ночной образ жизни , а Tapejara — как катемерный , будучи активным в течение дня в течение коротких промежутков времени. В результате возможно питающиеся рыбой Ctenochasma и Rhamphorhynchus могли иметь сходный характер активности с современными ночными морскими птицами, а фильтрующий Pterodaustro, возможно, имел сходный характер активности с современными гусеобразными птицами, которые кормятся ночью. Различия в характере активности Solnhofen птерозавров Ctenochasma , Rhamphorhynchus , Scaphognathus и Pterodactylus также могут указывать на разделение ниш между этими родами. [211]
Культурное значение
[ редактировать ]
Птерозавры были основным продуктом массовой культуры с тех пор, как их двоюродные братья динозавры, хотя они обычно не занимают столь заметного места в фильмах, литературе или другом искусстве. Хотя изображение динозавров в популярных средствах массовой информации радикально изменилось в ответ на достижения палеонтологии, с середины 20-го века сохраняется в основном устаревшее представление о птерозаврах. [212]

Для этих существ часто используется расплывчатый общий термин «птеродактиль». Животные, изображенные в художественной литературе и поп-культуре, часто представляют собой либо птеранодона , либо (нептеродактилоида) рамфоринха , либо вымышленный гибрид этих двух. [212] Во многих детских игрушках и мультфильмах изображены «птеродактили» с гребнями, как у птеранодонов , и длинными хвостами и зубами, как у рамфоринхов , - комбинация, которая никогда не существовала в природе. Тем не менее, по крайней мере, у одного птерозавра действительно были и гребень, и зубы, как у птеранодона : Ludodactylus , чье имя означает «игрушечный палец» из-за его сходства со старыми, неточными детскими игрушками. [213] Птерозавров иногда неправильно идентифицировали как (предков) птиц , хотя птицы являются динозаврами -тероподами , а не потомками птерозавров.
Птерозавры использовались в художественной литературе в романе сэра Артура Конан Дойля 1912 года «Затерянный мир» и его экранизации 1925 года . С тех пор они появлялись во многих фильмах и телевизионных программах, в том числе в фильме 1933 года «Кинг-Конг» в фильме 1966 года «Миллион лет до нашей эры » и . мембраны не разваливались, хотя эта конкретная ошибка была распространена в искусстве еще до того, как был снят фильм. Родан , вымышленный гигантский монстр (или кайдзю ), впервые появившийся в фильме 1956 года «Родан» , изображается как огромный облученный вид птеранодонов . [214] [215] Родан появлялся в нескольких японских о Годзилле фильмах , выпущенных в 1960-х, 1970-х, 1990-х и 2000-х годах, а также появился в фильме американского производства 2019 года «Годзилла: Король монстров» . [216] [217] [215]
После 1960-х годов птерозавры практически не появлялись в заметных американских фильмах до выхода в 2001 году « Парка Юрского периода III» . Палеонтолог Дэйв Хоун отметил, что птерозавры в этом фильме не были существенно обновлены с учетом современных исследований. Сохраняющимися ошибками были зубы, в то время как беззубый птеранодон должен был быть изображен, поведение при гнездовании, которое, как было известно к 2001 году, было неточным, и кожистые крылья, а не тугие мембраны мышечных волокон, необходимые для полета птерозавра. [212] Петри из «Земли до начала времен» (1988) — яркий пример анимационного фильма. [218]
В большинстве публикаций в средствах массовой информации птерозавры изображаются как рыбоядные , не отражая их полного разнообразия в питании. Их также часто изображают в виде воздушных хищников, похожих на хищных птиц , хватающих человеческие жертвы когтями на ногах. Однако только небольшой анурогнатид Vesperopterylus и небольшой вуконгоптерид Kunpengopterus [219] известно, что они обладают цепкими ногами и руками соответственно; У всех других известных птерозавров плоские стопоходящие ступни без противостоящих пальцев, а ступни обычно пропорционально малы, по крайней мере, в случае Pteranodontia. [16]
См. также
[ редактировать ]- Летающие и планирующие животные
- Графическая хронология птерозавров
- Список стратиграфических подразделений, содержащих птерозавров
- Список родов птерозавров
- Филогения птерозавров
- Пляж Птерозавров
- Размер птерозавра
- Хронология исследований птерозавров
Пояснительные примечания
[ редактировать ]- ^ Адаптировано из Witton (2013). [3] Таксономические группы на основе Unwin et al. (2010). [4]
- ^ См . Расширение экологических ниш в мезозое.
Ссылки
[ редактировать ]- ^ Jump up to: а б Андрес, Б.; Кларк, Дж.; Сюй, X. (2014). «Самый ранний птеродактилоид и происхождение группы» . Современная биология . 24 (9): 1011–16. Бибкод : 2014CBio...24.1011A . дои : 10.1016/j.cub.2014.03.030 . ПМИД 24768054 .
- ^ Барон, Мэтью Г. (2020). «Тестирование внутригрупповых взаимоотношений птерозавров посредством более широкой выборки таксонов и признаков avemetatarsalian, а также ряда методов филогенетического анализа» . ПерДж . 8 : е9604. дои : 10.7717/peerj.9604 . ПМЦ 7512134 . ПМИД 33005485 .
- ^ Марк П. Виттон (2013), Птерозавры: естественная история, эволюция, анатомия , Princeton University Press, Bibcode : 2013pnhe.book.....W , ISBN 978-0-691-15061-1
- ^ Дэвид М. Анвин (2010), « Darwinopterus и его значение для филогении птерозавров», Acta Geoscientica Sinica , 31 (1): 68–69.
- ^ Джонс, Дэниел (2003) [1917], Питер Роуч; Джеймс Хартманн; Джейн Сеттер (ред.), Словарь английского произношения , Кембридж: Издательство Кембриджского университета, ISBN 978-3-12-539683-8
- ^ «Птерозавр» . Словарь Merriam-Webster.com .
- ^ Кольбер, Эдвин Х. (Эдвин Харрис); Найт, Чарльз Роберт (1951). Книга динозавров: правящие рептилии и их родственники . Нью-Йорк: МакГроу-Хилл. п. 153.
- ^ «Распределение птерозавров во времени и пространстве: атлас» (PDF) . Зиттелиана : 61–107. 2008.
- ^ Элгин Р.А., Хоун Д.В., Фрей Э. (2011). «Размеры летной мембраны птерозавра» . Acta Palaeontologica Polonica . 56 (1): 99–111. дои : 10.4202/app.2009.0145 .
- ^ "Pterosaur.net :: Земное передвижение" . pterosaur.net . Проверено 01 февраля 2020 г.
- ^ Геггель 2018-12-17T19:23:17Z, Лаура (17 декабря 2018 г.). «Официально: летающие рептилии, называемые птерозаврами, были покрыты пушистыми перьями» . www.livscience.com . Проверено 01 февраля 2020 г.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Jump up to: а б Ван, X.; Келлнер, AWA; Чжоу, З.; Кампос, Д.А. (2008). «Обнаружение редкой древесно-лесной летающей рептилии (Pterosauria, Pterodactyloidea) из Китая» . Труды Национальной академии наук . 105 (6): 1983–87. Бибкод : 2008PNAS..105.1983W . дои : 10.1073/pnas.0707728105 . ПМЦ 2538868 . PMID 18268340 .
- ^ Лоусон Д.А. (март 1975 г.). «Птерозавр из позднего мелового периода Западного Техаса: открытие самого крупного летающего существа». Наука . 187 (4180): 947–948. Бибкод : 1975Sci...187..947L . дои : 10.1126/science.187.4180.947 . ПМИД 17745279 . S2CID 46396417 .
- ^ Буффето Э., Григореску Д., Чики З. (апрель 2002 г.). «Новый гигантский птерозавр с крепким черепом из позднего мелового периода Румынии» (PDF) . Naturwissenschaften . 89 (4): 180–84. Бибкод : 2002NW.....89..180B . дои : 10.1007/s00114-002-0307-1 . ПМИД 12061403 . S2CID 15423666 .
- ^ Бентон, Майкл Дж. (2004). «Происхождение и родство динозавров». В Вейшампеле, Дэвид Б.; Додсон, Питер; Осмольска, Гальшка (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 7–19 . ISBN 978-0-520-24209-8 .
- ^ Jump up to: а б с Нэйш, Даррен. «Птерозавры: мифы и заблуждения» . Птерозавр.нет . Проверено 18 июня 2011 г.
- ^ Jump up to: а б Александр, Дэвид Э. и Фогель, Стивен (2004). Летуны природы: птицы, насекомые и биомеханика полета . Джу Пресс. п. 191. ИСБН 978-0-8018-8059-9 .
- ^ Jump up to: а б с д Уитмер Л.М., Чаттерджи С., Франзоса Дж., Роу Т. (2003). «Нейроанатомия летающих рептилий и ее значение для полета, позы и поведения» (PDF) . Природа . 425 (6961): 950–53. Бибкод : 2003Natur.425..950W . дои : 10.1038/nature02048 . ПМИД 14586467 . S2CID 4431861 .
- ^ «Pterosaur.net :: Происхождение и взаимоотношения» . pterosaur.net . Проверено 01 февраля 2020 г.
- ^ Андрес, Брайан; Кларк, Джеймс М.; Син, Сюй (29 января 2010 г.). «Новый рамфоринхидный птерозавр из верхней юры Синьцзяна, Китай, и филогенетические отношения базальных птерозавров» (PDF) . Журнал палеонтологии позвоночных . 30 (1): 163–187. Бибкод : 2010JVPal..30..163A . дои : 10.1080/02724630903409220 . S2CID 53688256 .
- ^ Виттон, Марк П.; Мартилл, Дэвид М.; Ловеридж, Роберт Ф. (2010). «Подрезание крыльев гигантских птерозавров: комментарии к оценкам размаха крыльев и разнообразию». Acta Geoscientica Sinica . 31 : 79–81.
- ^ Виттон 2013 , с. 58.
- ^ Jump up to: а б с Уиттон 2013 , с. 23.
- ^ Jump up to: а б с д и ж г Анвин, Дэвид М. (2006). Птерозавры: Из глубин времени . Нью-Йорк: Пи Пресс. п. 246. ИСБН 978-0-13-146308-0 .
- ^ Jump up to: а б Уиттон 2013 , с. 27.
- ^ Jump up to: а б Веллнхофер 1991 , с. 47.
- ^ Jump up to: а б с Уиттон 2013 , с. 26.
- ^ Jump up to: а б с Уиттон 2013 , с. 24.
- ^ Jump up to: а б Фрей Э., Мартилл Д.М. (1998). «Сохранение мягких тканей экземпляра Pterodactylus kochi (Wagner) из верхней юры Германии». Новогодний альбом по геологии и палеонтологии, Трактаты . 210 (3): 421–41. дои : 10.1127/njgpa/210/1998/421 .
- ^ Веллнхофер 1991 , с. 48.
- ^ Jump up to: а б с д Нэйш Д., Мартилл Д.М. (2003). «Птерозавры – успешное вторжение в доисторическое небо». Биолог . 50 (5): 213–16.
- ^ Jump up to: а б Черкас С.А. и Джи К. (2002). Новый рамфоринхоид с гребнем на голове и сложными покровными структурами. В: Черкас, С.Дж. (Ред.). Пернатые динозавры и происхождение полета . Музей динозавров: Бландинг, Юта, 15–41. ISBN 1-932075-01-1 .
- ^ Веллнхофер 1991 , с. 49.
- ^ С. Кристофер Беннетт (1994). «Таксономия и систематика позднемелового птерозавра Pteranodon (Pterosauria, Pterodactyloidea)» . Случайные статьи Музея естественной истории Канзасского университета . 169 : 1–70.
- ^ Jump up to: а б Уиттон 2013 , с. 28.
- ^ Jump up to: а б с Веллнхофер 1991 , с. 50.
- ^ Виттон 2013 , с. 45.
- ^ Jump up to: а б с д Уиттон 2013 , с. 46.
- ^ Jump up to: а б с Уиттон 2013 , с. 30.
- ^ Jump up to: а б с Уиттон 2013 , с. 31.
- ^ Jump up to: а б Веллнхофер 1991 , с. 51.
- ^ Jump up to: а б с д и Веллнхофер 1991 , с. 52.
- ^ Виттон 2013 , с. 44.
- ^ Jump up to: а б с Уиттон 2013 , с. 32.
- ^ Виттон 2013 , с. 54.
- ^ Jump up to: а б Уиттон 2013 , с. 53.
- ^ Беннетт СК (2000). «Полет птерозавра: роль актинофибрилл в функции крыльев». Историческая биология . 14 (4): 255–84. Бибкод : 2000HBio...14..255B . дои : 10.1080/10292380009380572 . S2CID 85185457 .
- ^ Jump up to: а б с Келлнер, AWA; Ван, X.; Тишлингер, Х.; Кампос, Д.; Хон, DWE; Мэн, X. (2009). «Мягкие ткани Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) и строение мембраны крыла птерозавра» . Труды Королевского общества Б. 277 (1679): 321–29. дои : 10.1098/rspb.2009.0846 . ПМЦ 2842671 . ПМИД 19656798 .
- ^ Jump up to: а б Классенс LP, О'Коннор PM, Анвин DM (2009). Серено П. (ред.). «Дыхательная эволюция способствовала возникновению полета птерозавров и воздушного гигантизма» . ПЛОС ОДИН . 4 (2): е4497. Бибкод : 2009PLoSO...4.4497C . дои : 10.1371/journal.pone.0004497 . ПМК 2637988 . ПМИД 19223979 .
- ^ Jump up to: а б Уиттон 2013 , с. 52.
- ^ Jump up to: а б Уиттон 2013 , с. 55.
- ^ Jump up to: а б Анвин Д.М., Бахурина Н.Н. (1994). « Sordes pilosus и природа летательного аппарата птерозавров». Природа . 371 (6492): 62–64. Бибкод : 1994Natur.371...62U . дои : 10.1038/371062a0 . S2CID 4314989 .
- ^ Ван X, Чжоу Z, Чжан Ф, Сюй X (2002). «Почти полностью сочлененный рамфоринхоидный птерозавр с исключительно хорошо сохранившимися перепонками крыльев и «волосами» из Внутренней Монголии, северо-восточный Китай». Китайский научный бюллетень . 47 (3):3. Бибкод : 2002ЧСБу..47..226Вт . дои : 10.1360/02tb9054 . S2CID 86641794 .
- ^ Фрей, Э.; Тишлингер, Х.; Буши, М.-К.; Мартилл, DM (2003). «Новые экземпляры птерозавров (рептилий) с мягкими частями, имеющие значение для анатомии и передвижения птерозавров». Геологическое общество, Лондон, специальные публикации . 217 (1): 233–66. Бибкод : 2003GSLSP.217..233F . дои : 10.1144/ГСЛ.СП.2003.217.01.14 . S2CID 130462931 .
- ^ Дайк Г.Дж., Наддс Р.Л., Рейнер Дж.М. (июль 2006 г.). «Несоответствие конечностей и форма крыльев птерозавров» . Дж. Эвол. Биол . 19 (4): 1339–42. дои : 10.1111/j.1420-9101.2006.01096.x . ПМИД 16780534 . S2CID 30516133 .
- ^ Jump up to: а б с Веллнхофер 1991 , с. 53.
- ^ Виттон 2013 , с. 33.
- ^ Виттон 2013 , с. 34.
- ^ Уилкинсон М.Т., Анвин Д.М., Эллингтон К.П. (2006). «Высокая подъемная функция птероидной кости и переднего крыла птерозавров» . Труды Королевского общества Б. 273 (1582): 119–26. дои : 10.1098/rspb.2005.3278 . ПМК 1560000 . ПМИД 16519243 .
- ^ Беннетт СК (2007). «Сочленение и функция птероидной кости птерозавров» (PDF) . Журнал палеонтологии позвоночных . 27 (4): 881–91. doi : 10.1671/0272-4634(2007)27[881:AAFOTP]2.0.CO;2 . S2CID 86326537 .
- ^ Чжоу, Чан Фу; Шох, Райнер Р. (2011). «Новый материал о нептеродактилоидном птерозавре Changchengopterus pani Lü, 2009 из позднеюрской формации Тяоцзишань на западе Ляонина». Новогодний альбом по геологии и палеонтологии, Трактаты . 260 (3): 265–75. дои : 10.1127/0077-7749/2011/0131 .
- ^ Ван, Сяо-Линь; Келлнер, Александр В.А.; Цзян, Шунь-Син; Ченг, Синь; Мэн, Си; Родригес, Таисса (2010). «Новые длиннохвостые птерозавры (Wukongopteridae) из западного Ляонина, Китай» . Анналы Бразильской академии наук . 82 (4): 1045–62. дои : 10.1590/s0001-37652010000400024 . ПМИД 21152776 .
- ^ Уилкинсон М.Т.; Анвин ДМ; Эллингтон CP (2006). «Высокая подъемная функция птероидной кости и переднего крыла птерозавров» . Труды Королевского общества Б. 273 (1582): 119–26. дои : 10.1098/rspb.2005.3278 . ПМК 1560000 . ПМИД 16519243 .
- ^ Jump up to: а б с д и ж г час Уиттон 2013 , с. 35.
- ^ Jump up to: а б с д Веллнхофер 1991 , с. 55.
- ^ Wellnhofer 1991 , стр. 53–54.
- ^ Питтман, Майкл; Барлоу, Люк А.; Кэй, Томас Г.; Хабиб, Майкл Б. (2021). «У птерозавров развилось мускулистое соединение крыла и тела, обеспечивающее многогранные преимущества летных характеристик: улучшенное аэродинамическое сглаживание, сложное управление корнем крыла и создание силы крыла» . Труды Национальной академии наук . 118 (44): e2107631118. Бибкод : 2021PNAS..11807631P . дои : 10.1073/pnas.2107631118 . ISSN 0027-8424 . ПМЦ 8612209 . ПМИД 34663691 . S2CID 239028043 .
- ^ Jump up to: а б с д и ж Веллнхофер 1991 , с. 56.
- ^ Jump up to: а б с д и ж Веллнхофер 1991 , с. 57.
- ^ Jump up to: а б с Уиттон 2013 , с. 36.
- ^ Jump up to: а б с д Член парламента Уиттона, Нэйш Д. (2008). Макклейн CR (ред.). «Переоценка функциональной морфологии и палеоэкологии аждархид-птерозавров» . ПЛОС ОДИН . 3 (5): е2271. Бибкод : 2008PLoSO...3.2271W . дои : 10.1371/journal.pone.0002271 . ПМК 2386974 . ПМИД 18509539 .
- ^ Виттон 2013 , с. 37.
- ^ Виттон 2013 , с. 39.
- ^ Виттон 2013 , с. 43.
- ^ Виттон 2013 , с. 47.
- ^ Виттон 2013 , с. 48.
- ^ Jump up to: а б с д и Уиттон 2013 , с. 51.
- ^ Гольдфус, А. (1831). «Вклад в познание различных рептилий доисторического мира». Nova Acta Academiae Leopoldinae . 15 :61–128.
- ^ Ян, Цзысяо; Цзян, Баоюй; Макнамара, Мария Э.; Кернс, Стюарт Л.; Питтман, Майкл; Кэй, Томас Г.; Орр, Патрик Дж.; Сюй, Син; Бентон, Майкл Дж. (январь 2019 г.). «Покровные структуры птерозавра со сложным перистым ветвлением» . Экология и эволюция природы . 3 (1): 24–30. дои : 10.1038/s41559-018-0728-7 . hdl : 1983/1f7893a1-924d-4cb3-a4bf-c4b1592356e9 . ПМИД 30568282 . S2CID 56480710 .
- ^ Бриггс, Хелен (17 декабря 2018 г.). «Мех летит над новыми окаменелостями птерозавров» . Новости Би-би-си . Проверено 19 декабря 2018 г.
- ^ Анвин, Дэвид М.; Мартилл, Дэвид М. (декабрь 2020 г.). «У птерозавров нет протоперьев». Экология и эволюция природы . 4 (12): 1590–1591. Бибкод : 2020NatEE...4.1590U . дои : 10.1038/s41559-020-01308-9 . ПМИД 32989266 . S2CID 222168569 .
- ^ Келлнер; и др. (2009). «Мягкая ткань Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) и строение мембраны крыла птерозавра» . Труды Королевского общества B: Биологические науки . 277 (1679): 321–29. дои : 10.1098/rspb.2009.0846 . ПМЦ 2842671 . ПМИД 19656798 .
- ^ Анвин, Дэвид М.; Бахурина, Наташа Н. (сентябрь 1994 г.). «Sordes pilosus и природа летного аппарата птерозавров». Природа . 371 (6492): 62–64. Бибкод : 1994Natur.371...62U . дои : 10.1038/371062a0 . S2CID 4314989 .
- ^ Ян, Цзысяо; Цзян, Баоюй; Макнамара, Мария Э.; Кернс, Стюарт Л.; Питтман, Майкл; Кэй, Томас Г.; Орр, Патрик Дж.; Сюй, Син; Бентон, Майкл Дж. (декабрь 2020 г.). «Ответ на: У птерозавров нет протоперьев». Экология и эволюция природы . 4 (12): 1592–1593. Бибкод : 2020NatEE...4.1592Y . дои : 10.1038/s41559-020-01309-8 . hdl : 10468/11874 . ПМИД 32989267 . S2CID 222163211 .
- ^ Д'Альба, Лилиана (январь 2019 г.). «Оперение птерозавра» . Экология и эволюция природы . 3 (1): 12–13. дои : 10.1038/s41559-018-0767-0 . ПМИД 30568284 . S2CID 56480834 .
- ^ Чинкотта; и др. (2022). «Меланосомы птерозавров поддерживают сигнальные функции ранних перьев» . Природа . 604 (7907): 684–688. Бибкод : 2022Natur.604..684C . дои : 10.1038/s41586-022-04622-3 . ПМК 9046085 . ПМИД 35444275 .
- ^ Jump up to: а б Уиттон 2013 , с. 5.
- ^ Веллнхофер 1991 , с. 22.
- ^ Виттон 2013 , с. 6.
- ^ Виттон 2013 , стр. 6–7.
- ^ Jump up to: а б с д Уиттон 2013 , с. 7.
- ^ Коллини, Калифорния (1784 г.). «О некоторых зоолитах из кабинета естествознания SASE Палатин и Баварии в Мангейме». Acta Theodoro-Palatinae Mannheim 5 Pars Physica , стр. 58–103 (1 пластина).
- ^ Ваглер, Дж. (1830). Естественная система амфибий Мюнхен, 1830: 1–354.
- ^ Кювье Ж (1801). «[Летающая рептилия]. В: Отрывок из работы о видах четвероногих, кости которых были найдены в недрах Земли». Журнал физики, химии и естествознания . 52 :253–67.
- ^ Кювье, Г., 1809, «Мемуары об ископаемом скелете летающей рептилии из окрестностей Айхштедта, которого некоторые натуралисты приняли за птицу, и поэтому мы образуем род ящеров под названием Ptero-Dactyle», Анналы Музея естественной истории , Париж, 13 стр. 424–37
- ^ Рафинеск, CS, 1815, Анализ природы или таблица Вселенной и организованных тел , Палермо
- ^ Фон Земмерринг, ST, 1812, «Об орнитоцефале или о неизвестном животном доисторического мира, чей ископаемый скелет Коллини описал в 5-м томе Actorum Academiae Theodoro-Palatinae вместе с иллюстрацией в натуральную величину в 1784 году, и который скелет в настоящее время находится в коллекции естественной истории Королевской академии наук в Мюнхене», Меморандумы Королевской Баварской академии наук , Мюнхен: Математико-физический класс 3 : 89–158.
- ^ Веллнхофер 1991 , с. 27.
- ^ Ньюман, Э. (1843). «Заметка о племени птеродактилей, рассматриваемом как сумчатые летучие мыши». Зоолог . 1 : 129–31.
- ^ Кауп, Дж. (1834 г.). «Попытайтесь разделить млекопитающих на 6 триб, а земноводных на 6 отрядов» . Исида из Окена . 1834 : 311–315.
- ^ Веллнхофер 1991 , с. 28.
- ^ Веллнхофер 1991 , с. 29.
- ^ Веллнхофер 1991 , с. 33.
- ^ Сили, Х.Г., 1870, Орнитозаврия - элементарное исследование костей птеродактилей , Cambridge University Press.
- ^ Сили, Х.Г., 1901, Воздушные драконы: отчет о вымерших летающих рептилиях , Лондон: Метуэн.
- ^ Миварт, Г. (1881). «Популярный рассказ о хамелеонах». Природа . 24 (615): 309–38. Бибкод : 1881Natur..24..335. . дои : 10.1038/024335f0 . S2CID 30819954 .
- ^ Jump up to: а б Веллнхофер 1991 , с. 35.
- ^ Jump up to: а б с Веллнхофер 1991 , с. 36.
- ^ Jump up to: а б Веллнхофер 1991 , с. 31.
- ^ Wellnhofer 1991 , стр. 37–38.
- ^ Марш, О.К. (1882 г.). «Крылья Птеродактиля». Американский научный журнал . 3 (16): 223.
- ^ Зиттель, К.А. (1882). «О птерозаврах с литографского сланца Баварии». Палеонтографика . 29 :47–80.
- ^ Бройли, Ф., 1927, «Рамфоринх со следами волосяного покрова», Труды Баварской академии наук , с. 49-67
- ^ Эдингер, Т (1927). «Мозг птерозавра» (PDF) . Журнал анатомии и истории развития . 83 (1/3): 105–12. дои : 10.1007/bf02117933 . S2CID 19084773 . Архивировано из оригинала (PDF) 28 июля 2020 г. Проверено 27 октября 2019 г.
- ^ Ханкин Э.Х. и Watson DSM; «О полете птеродактилей», Аэронавтический журнал , октябрь 1914 г., стр. 324–35.
- ^ Баккер, Роберт, 1986, Ереси динозавров , Лондон: Penguin Books, 1988, стр. 283
- ^ Падиан, К. (1979). «Крылья птерозавров: новый взгляд». Открытие . 14 : 20–29.
- ^ Падиан, К., 1980, Исследования строения, эволюции и полета птерозавров (рептилий: Pterosauria) , доктор философии. дисс., кафедра биологии Йельского университета
- ^ Jump up to: а б Падиан К. (1983). «Функциональный анализ полета и ходьбы птерозавров». Палеобиология . 9 (3): 218–39. Бибкод : 1983Pbio....9..218P . дои : 10.1017/S009483730000765X . JSTOR 2400656 . S2CID 88434056 .
- ^ Jump up to: а б с д и Уиттон 2013 , с. 9.
- ^ Веллнхофер, П., 1978, Справочник по палеогерпетологии XIX. Птерозаврия , Urban & Fischer, Мюнхен
- ^ Wellnhofer 1991 , стр. 1–192.
- ^ Дайк, Дж. Дж. Макгоуэн; Наддс, РЛ; Смит, Д. (2009). «Форма эволюции птерозавров: свидетельства окаменелостей» . Журнал эволюционной биологии . 22 (4): 890–98. дои : 10.1111/j.1420-9101.2008.01682.x . ПМИД 19210587 . S2CID 32518380 .
- ^ Jump up to: а б Виттон 2013
- ^ Jump up to: а б Уиттон 2013 , с. 10.
- ^ « Великолепно сохранившаяся окаменелость птерозавра, обнаруженная в Шотландии» . Ассошиэйтед Пресс (АП) . 22 февраля 2022 г.
- ^ Виттон 2013 , с. 13.
- ^ Виттон 2013 , стр. 14, 17.
- ^ Бентон, MJ (1999). « Scleromochlus taylori и происхождение динозавров и птерозавров» . Философские труды Королевского общества B: Биологические науки . 354 (1388): 1423–46. дои : 10.1098/rstb.1999.0489 . ПМЦ 1692658 .
- ^ Беннетт, С. Кристофер (1996). «Филогенетическое положение птерозавров внутри архозавроморф» . Зоологический журнал Линнеевского общества . 118 (3): 261–308. дои : 10.1111/j.1096-3642.1996.tb01267.x .
- ^ Ирмис, РБ; Несбитт, С.Дж.; Падиан, К.; Смит, Северная Дакота; Тернер, А.Х.; Вуди, Д.; Даунс, А. (2007). «Сборка динозавров позднего триаса из Нью-Мексико и появление динозавров» (PDF) . Наука . 317 (5836): 358–61. Бибкод : 2007Sci...317..358I . дои : 10.1126/science.1143325 . ПМИД 17641198 . S2CID 6050601 .
- ^ Jump up to: а б Хонинговать DWE; Бентон М.Дж. (2007). «Оценка филогенетических отношений птерозавров с рептилиями-архозавроморфами». Журнал систематической палеонтологии . 5 (4): 465–69. дои : 10.1017/S1477201907002064 . S2CID 86145645 .
- ^ Несбитт, SJ (2011). «Ранняя эволюция архозавров: взаимоотношения и происхождение основных клад» . Бюллетень Американского музея естественной истории . 352 : 1–292. дои : 10.1206/352.1 . hdl : 2246/6112 . S2CID 83493714 .
- ^ Эскурра, Мартин Д. (28 апреля 2016 г.). «Филогенетические взаимоотношения базальных архозавроморфов с акцентом на систематике протерозуховых архозавроформ» . ПерДж . 4 : е1778. дои : 10.7717/peerj.1778 . ПМЦ 4860341 . ПМИД 27162705 .
- ^ Беннетт, Южная Каролина (2020). «Переоценка триасового архозаврообразного Scleromochlus taylori : ни бегуна, ни двуногого, а прыгуна» . ПерДж . 8 : е8418. дои : 10.7717/peerj.8418 . ПМЦ 7035874 . ПМИД 32117608 .
- ^ Jump up to: а б Эскурра, Мартин Д.; Несбитт, Стерлинг Дж.; Бронзати, Марио; Далла Веккья, Фабио Марко; Аньолин, Федерико Л.; Бенсон, Роджер Б.Дж.; Бриссон Эгли, Федерико; Кабрейра, Серхио Ф.; Эверс, Сережа В.; Джентиль, Адриэль Р.; Ирмис, Рэндалл Б.; Мартинелли, Агустин Г.; Новас, Фернандо Э.; Роберто да Силва, Лусио; Смит, Натан Д.; Стокер, Мишель Р.; Тернер, Алан Х.; Лангер, Макс К. (17 декабря 2020 г.). «Загадочные предшественники динозавров устраняют пробел в происхождении птерозаврии» (PDF) . Природы . 588 (7838): 445–449. Бибкод : 2020Natur.588..445E . дои : 10.1038/s41586-020-3011-4 . ПМИД 33299179 . S2CID 228077525 .
- ^ «Палеонтологи находят предшественников птерозавров, которые заполняют пробел в ранней эволюционной истории» . физ.орг . Проверено 14 декабря 2020 г.
- ^ Блэк, Райли. «Происхождение птерозавров становится в фокусе» . Научный американец . Проверено 14 декабря 2020 г.
- ^ Барон, Мэтью Г. (октябрь 2021 г.). «Происхождение птерозавров». Обзоры наук о Земле . 221 : 103777. Бибкод : 2021ESRv..22103777B . doi : 10.1016/j.earscirev.2021.103777 .
- ^ Виттон 2013 , с. 18.
- ^ Руперт Уайлд, 1983, «О происхождении птерозавров», Академия Вельтенбергера, Эрвин Рютте-Фестшрифт , стр. 231–38.
- ^ Jump up to: а б с д Виттон, Марк П. (2015). «Были ли ранние птерозавры неумелыми наземными двигательными аппаратами?» . ПерДж . 3 : е1018. дои : 10.7717/peerj.1018 . ПМЦ 4476129 . ПМИД 26157605 .
- ^ Документальный фильм BBC: Прогулка с динозаврами (серия 4) - Небесный гигант на 22 футах, Тим Хейнс, 1999 г.
- ^ Слэк К.Э., Джонс С.М., Андо Т. и др. (июнь 2006 г.). «Ранние окаменелости пингвинов, а также митохондриальные геномы определяют эволюцию птиц» . Молекулярная биология и эволюция . 23 (6): 1144–55. дои : 10.1093/molbev/msj124 . ПМИД 16533822 .
- ^ Батлер, Ричард Дж.; Барретт, Пол М.; Новбат, Стивен и Апчерч, Пол (2009). «Оценка влияния систематических ошибок выборки на структуру разнообразия птерозавров: последствия для гипотез о конкурентной замене птиц и птерозавров». Палеобиология . 35 (3): 432–46. Бибкод : 2009Pbio...35..432B . дои : 10.1666/0094-8373-35.3.432 . S2CID 84324007 .
- ^ Jump up to: а б с Лонгрич, Северная Каролина; Мартилл, DM; Андрес, Б. (2018). «Позднемаастрихтские птерозавры из Северной Африки и массовое вымирание птерозавров на рубеже мела и палеогена» . ПЛОС Биология . 16 (3): e2001663. дои : 10.1371/journal.pbio.2001663 . ПМЦ 5849296 . ПМИД 29534059 .
- ^ Андрес, Б.; Майерс, Т.С. (2013). «Одинокая звезда Птерозавры». Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 103 (3–4): 1. Бибкод : 2012EESTR.103..383A . дои : 10.1017/S1755691013000303 . S2CID 84617119 .
- ^ Аньолин, Федерико Л. и Варриккио, Дэвид (2012). «Систематическая реинтерпретация Пикси барбарульны Варриккио, 2002 г. из формации Ту Медисин (верхний мел) на западе США (Монтана) как птерозавра, а не птицы» (PDF) . Геодиверситас . 34 (4): 883–94. дои : 10.5252/g2012n4a10 . S2CID 56002643 . Архивировано из оригинала (PDF) 15 января 2013 г. Проверено 29 декабря 2012 г.
- ^ Прондвай, Э.; Бодор, скорая помощь; Оси, А. (2014). «Отражает ли морфология онтогенез, основанный на остеогистологии? Исследование симфизов челюстей птерозавров позднего мела из Венгрии выявляет скрытое таксономическое разнообразие» (PDF) . Палеобиология . 40 (2): 288–321. Бибкод : 2014Pbio...40..288P . дои : 10.1666/13030 . S2CID 85673254 .
- ^ Мартин-Сильверстоун, Элизабет; Виттон, Марк П.; Арбур, Виктория М.; Карри, Филип Дж. (2016). «Небольшой аждархоидный птерозавр из позднего мела, эпохи летающих гигантов» . Королевское общество открытой науки . 3 (8): 160333. Бибкод : 2016RSOS....360333M . дои : 10.1098/rsos.160333 . ПМК 5108964 . ПМИД 27853614 .
- ^ Халуза, А.; Апестегия, С. (2007). «Останки птерозавров (Архозаврия, Орнитодира) из раннего позднего мела Ла-Буитрера, Рио-Негро, Аргентина» . XXIII Аргентинская конференция по палеонтологии позвоночных .
- ^ Jump up to: а б Смит, Рой Э.; Чинсами, Анусуя; Анвин, Дэвид М.; Ибрагим, Низар; Зухри, Самир; Мартилл, Дэвид М. (16 октября 2021 г.). «Маленькие незрелые птерозавры из мелового периода Африки: последствия тафономической предвзятости и структуры палеосообщества летающих рептилий» . Меловые исследования . 130 : 105061. doi : 10.1016/j.cretres.2021.105061 . S2CID 239257717 .
- ^ Келлнер, AW (2003). «Филогения птерозавров и комментарии к эволюционной истории группы». Геологическое общество, Лондон, специальные публикации . 217 (1): 105–37. Бибкод : 2003GSLSP.217..105K . дои : 10.1144/gsl.sp.2003.217.01.10 . S2CID 128892642 .
- ^ Несбитт, С.Дж., Дезохо, Дж.Б., и Ирмис, Р.Б. (2013). Анатомия, филогения и палеобиология ранних архозавров и их родственников . Геологическое общество Лондона. ISBN 1862393613
- ^ де Кейрос, К.; Кантино, PD; Готье, Ж.А., ред. (2020). Филонимы: дополнение к Филокоду . CRC Press Бока-Ратон, Флорида. п. 2072. ИСБН 9780429821202 .
- ^ Падиан, К. (1997). «Pterosauromorpha», стр. 617–18 в Карри, П.Дж. и Падиан, К. Энциклопедия динозавров . Академическая пресса. ISBN 0122268105 .
- ^ Люй Дж.; Анвин ДМ; Сюй Л.; Чжан С. (2008). «Новый аждархоидный птерозавр из нижнего мела Китая и его значение для филогении и эволюции птерозавров». Naturwissenschaften . 95 (9): 891–97. Бибкод : 2008NW.....95..891L . дои : 10.1007/s00114-008-0397-5 . ПМИД 18509616 . S2CID 13458087 .
- ^ Jump up to: а б Аллейн, Ричард (1 октября 2008 г.). «Птеродактили были слишком тяжелы для полета, — утверждают учёные» . Телеграф . Архивировано из оригинала 31 октября 2009 года . Проверено 2 марта 2012 г.
- ^ Пауэлл, Девин (2 октября 2008 г.). «Были ли птерозавры слишком большими, чтобы летать?» . НовыйУченый . Проверено 2 марта 2012 г.
- ^ Темплин, Р.Дж.; Чаттерджи, Санкар (2004). Поза, передвижение и палеоэкология птерозавров . Боулдер, Колорадо: Геологическое общество Америки. п. 60. ИСБН 978-0-8137-2376-1 .
- ^ Нэйш, Даррен (18 февраля 2009 г.). «Птерозавры дышали подобно птицам и имели в крыльях надувные воздушные мешки» . Научные блоги . Архивировано из оригинала 21 февраля 2009 года . Проверено 3 апреля 2016 г.
- ^ Jump up to: а б с «Почему птерозавры все-таки не были такими страшными» . Газета «Обозреватель» . 11 августа 2013 года . Проверено 12 августа 2013 г.
- ^ Jump up to: а б Хехт, Джефф (16 ноября 2010 г.). «Гигантские птерозавры прыгали ввысь, как летучие мыши-вампиры?» . НовыйУченый . Проверено 2 марта 2012 г.
- ^ Маккриди, П. (1985). «Великий проект птеродактиля» (PDF) . Инженерия и наука . 49 (2): 18–24.
- ^ Молоцкий, Ирвин (28 января 1986 г.). «Модель птеродактиля взмахивает крыльями и поднимается в воздух» . Нью-Йорк Таймс .
- ^ «Кончики крыльев птерозавров: анатомия, воздухоплавательная функция и три экологических значения» (PDF) . Qmro.qmul.ac.uk. Проверено 25 июня 2022 г.
- ^ Гейст, Н.; Хиллениус, В.; Фрей, Э.; Джонс, Т.; Элгин, Р. (2014). «Дыхание в коробке: ограничения вентиляции легких у гигантских птерозавров» . Анатомическая запись . 297 (12): 2233–53. дои : 10.1002/ar.22839 . ПМИД 24357452 . S2CID 27659270 .
- ^ Хопсон Дж. А. (1977). «Относительный размер мозга и поведение архозавровых рептилий». Ежегодный обзор экологии и систематики . 8 : 429–48. doi : 10.1146/annurev.es.08.110177.002241 .
- ^ Антес, Эмили (18 ноября 2013 г.). «Хладнокровный не значит глупый» . Нью-Йорк Таймс .
- ^ Кодорню, Лаура; Полина Карабахал, Ариана; Пол, Диего; Анвин, Дэвид; Раухут, Оливер В.М. (2016). «Юрский птерозавр из Патагонии и происхождение птеродактилоидного нейрокраниума» . ПерДж . 4 : е2311. дои : 10.7717/peerj.2311 . ПМК 5012331 . ПМИД 27635315 .
- ^ Падиан К. (2003). «Стойка и походка птерозавра и интерпретация следов» (PDF) . Ихнос . 10 (2–4): 115–26. Бибкод : 2003Ично..10..115П . дои : 10.1080/10420940390255501 . S2CID 129113446 .
- ^ Хван К., Ха М., Локли М.Г., Анвин Д.М., Райт Дж.Л. (2002). «Новые следы птерозавров (Pteraichnidae) из позднемеловой формации Уханри, юго-западная Корея» . Геологический журнал . 139 (4): 421–35. Бибкод : 2002GeoM..139..421H . дои : 10.1017/S0016756802006647 . S2CID 54996027 .
- ^ Анвин Д.М. (1997). «Следы птерозавров и наземные способности птерозавров» (PDF) . Летайя . 29 (4): 373–86. дои : 10.1111/j.1502-3931.1996.tb01673.x .
- ^ Jump up to: а б с Уиттон 2013 , с. 51
- ^ Беннетт, Южная Каролина (2007). «Второй экземпляр птерозавра Anurognathus ammoni ». Палеонтологический журнал . 81 (4): 376–98. Бибкод : 2007PalZ...81..376B . дои : 10.1007/bf02990250 . S2CID 130685990 .
- ^ Виттон 2013 , с. 103.
- ^ Виттон 2013 , с. 121.
- ^ Виттон 2013 , с. 122.
- ^ Андрес, Б.; Кларк, Дж. М.; Син, X. (2010). «Новый рамфоринхидный птерозавр из верхней юры Синьцзяна, Китай, и филогенетические отношения базальных птерозавров» (PDF) . Журнал палеонтологии позвоночных . 30 (1): 163–87. Бибкод : 2010JVPal..30..163A . дои : 10.1080/02724630903409220 . S2CID 53688256 .
- ^ Виттон 2013 , с. 134.
- ^ Люй Дж.; Сюй Л.; Чанг Х.; Чжан С. (2011). «Новый птерозавр-дарвиноптерид из средней юры западного Ляонина, северо-восточного Китая и его экологические последствия». Acta Geologica Sinica — английское издание . 85 (3): 507–14. Бибкод : 2011AcGlS..85..507L . дои : 10.1111/j.1755-6724.2011.00444.x . S2CID 128545851 .
- ^ Мартилл, Дэвид М.; Смит, Рой Э.; Лонгрич, Николас; Браун, Джеймс (01 января 2021 г.). «Свидетельства тактильного поиска пищи у птерозавров: чувствительный кончик клюва Lonchodraco giganteus (Pterosauria, Lonchodectidae) из верхнего мела на юге Англии» . Меловые исследования . 117 : 104637. Бибкод : 2021CrRes.11704637M . doi : 10.1016/j.cretres.2020.104637 . ISSN 0195-6671 . S2CID 225130037 .
- ^ Виттон 2013 , стр. 150–51.
- ^ Виттон 2013 , с. 199.
- ^ У, Вэнь-Хао; Чжоу, Чан-Фу; Андрес, Брайан (2017). «Беззубый птерозавр Jidapterus edentus (Pterodactyloidea: Azhdarchoidea) из раннемеловой биоты Джехол и его палеоэкологические последствия» . ПЛОС ОДИН . 12 (9): e0185486. Бибкод : 2017PLoSO..1285486W . дои : 10.1371/journal.pone.0185486 . ПМК 5614613 . ПМИД 28950013 .
- ^ Пегас, Р.В., и Келлнер, AW (2015). Предварительная миологическая реконструкция нижней челюсти Thalassodromeus sethi (Pterodactyloidea: Tapejaridae). Flugsaurier 2015 Портсмут, тезисы, 47–48
- ^ Уиттон, член парламента; Нэйш, Д. (2015). «Аждархидские птерозавры: водотралые имитаторы пеликанов или «наземные сталкеры»?» . Acta Palaeontologica Polonica . 60 (3). дои : 10.4202/app.00005.2013 .
- ^ Нэйш, Д.; Виттон, член парламента (2017). «Биомеханика шеи указывает на то, что гигантские трансильванские птерозавры-аждархиды были хищниками с короткой шеей» . ПерДж . 5 : e2908. дои : 10.7717/peerj.2908 . ПМЦ 5248582 . ПМИД 28133577 .
- ^ Уиттон, М.; Брусатте, С.; Дайк, Г.; Нэйш, Д.; Норелл, М.; Времир, М. (2013). Повелители птерозавров Трансильвании: гигантские аждархиды с короткой шеей в позднемеловой Румынии . Ежегодный симпозиум по палеонтологии и сравнительной анатомии позвоночных. Эдинбург. Архивировано из оригинала 6 апреля 2016 г.
- ^ Мартилл, Дэвид М.; Ибрагим, Низар (март 2015 г.). «Необычная модификация челюстей у Alanqa, птерозавра-аждархида среднего мела из кем-кемских отложений Марокко» . Меловые исследования . 53 : 59–67. Бибкод : 2015CrRes..53...59M . дои : 10.1016/j.cretres.2014.11.001 .
- ^ Jump up to: а б с Пегас, Родриго V; Коста, Фабиана Р; Келлнер, Александр В.А. (24 сентября 2021 г.). «Реконструкция приводящей камеры и прогнозируемая сила укуса у птеродактилоидов (Pterosauria)». Зоологический журнал Линнеевского общества . 193 (2): 602–635. doi : 10.1093/zoolinnean/zlaa163 .
- ^ Баффето Э., Мартий Д., Эскуилье Ф. (июль 2004 г.). «Птерозавры как часть диеты спинозавров» . Природа . 430 (6995): 33. Бибкод : 2004Natur.429...33B . дои : 10.1038/430033а . ПМИД 15229562 . S2CID 4398855 .
- ^ «Доисторические акулы питались летающими рептилиями, как показывают окаменелости» . Наука и инновации . 3 октября 2018 г. Архивировано из оригинала 3 октября 2018 г.
- ^ Цзи Ц, Цзи С.А., Ченг Юнь и др. (декабрь 2004 г.). «Палеонтология: яйцо птерозавра с кожистой скорлупой» (PDF) . Природа . 432 (7017): 572. Бибкод : 2004Natur.432..572J . дои : 10.1038/432572a . ПМИД 15577900 . S2CID 4416203 .
- ^ Кодорню, Л.; Чиаппе, Л.; Риварола, Д. (2014). «Первое сообщение об эмбрионе птерозавра (нижний мел, Сан-Луис, Аргентина)» . Амегиниана . 41 (4 (приложение, тезисы из XX Jornadas Argentinas de Paleontologia de Vertebrados, Ла-Плата, 26-28 мая 2004 г.)): 40R.
- ^ Кьяппе, Луис М.; Кодорню, Лаура; Греллет-Тиннер, Джеральд; Риварола, Дэвид (декабрь 2004 г.). «Окаменелость невылупившегося аргентинского птерозавра» . Природа . 432 (7017): 571–572. дои : 10.1038/432571a . hdl : 11336/156308 . ISSN 1476-4687 . ПМИД 15577899 .
- ^ Кодорню, Лаура; Кьяппе, Луис М.; Сид, Фабрисио Д. (май 2013 г.). «Первое появление камней в желудке у птерозавров» . Журнал палеонтологии позвоночных . 33 (3): 647–654. Бибкод : 2013JVPal..33..647C . дои : 10.1080/02724634.2013.731335 . hdl : 11336/4391 . ISSN 0272-4634 .
- ^ Люй Дж.; Анвин ДМ; Считая DC; Джин Х.; Лю Ю.; Джи К. (2011). «Ассоциация яйцо-взрослая особь, пол и размножение птерозавров». Наука . 331 (6015): 321–24. Бибкод : 2011Sci...331..321L . дои : 10.1126/science.1197323 . ПМИД 21252343 . S2CID 206529739 .
- ^ Jump up to: а б Ван, Сяолинь (2014). «Сексуально-диморфные, трехмерно сохранившиеся птерозавры и их яйца из Китая» . Современная биология . 24 (12): 1323–30. Бибкод : 2014CBio...24.1323W . дои : 10.1016/j.cub.2014.04.054 . ПМИД 24909325 . [ постоянная мертвая ссылка ]
- ^ Грелле-Тиннер Дж., Роу С., Томпсон М.Б., Джи Кью (2007). «Заметка о гнездовом поведении птерозавров». Историческая биология . 19 (4): 273–77. Бибкод : 2007HBio...19..273G . дои : 10.1080/08912960701189800 . S2CID 85055204 .
- ^ Сяолинь Ван, Келлнер Александр В.А.; Ченг, Синь; Цзян, Шуньсин; Ван, Цян; Саяо Хулиана, М.; Рордригес Таисса, Коста Фабиана Р.; Оболочка; Мэн, Си; Чжоу, Чжунхэ (2015). «Яичная скорлупа и гистология дают представление об истории жизни птерозавра с двумя функциональными яичниками» . Анаис да Академия Бразилиа де Сиенсиас . 87 (3): 1599–1609. дои : 10.1590/0001-3765201520150364 . ПМИД 26153915 .
- ^ Ван X, Чжоу Z (июнь 2004 г.). «Палеонтология: эмбрион птерозавра из раннего мела» . Природа . 429 (6992): 621. Бибкод : 2004Natur.429..621W . дои : 10.1038/429621a . ПМИД 15190343 . S2CID 4428545 .
- ^ Jump up to: а б «Детеныши птерозавров нуждались в своих родителях, как показывает клад яиц (обновление)» . физ.орг . Проверено 21 марта 2020 г.
- ^ Jump up to: а б с Беннетт СК (1995). «Статистическое исследование Rhamphorhynchus из известняка Зольнхофен в Германии: годовые классы одного крупного вида». Журнал палеонтологии . 69 (3): 569–80. Бибкод : 1995JPal...69..569B . дои : 10.1017/S0022336000034946 . JSTOR 1306329 . S2CID 88244184 .
- ^ Jump up to: а б Прондвай, Э.; Штейн, К.; Оси, А.; Сандер, член парламента (2012). Соарес, Дафна (ред.). «История жизни рамфоринха , выведенная на основе гистологии костей и разнообразия стратегий роста птерозавров» . ПЛОС ОДИН . 7 (2): e31392. Бибкод : 2012PLoSO...731392P . дои : 10.1371/journal.pone.0031392 . ПМК 3280310 . ПМИД 22355361 .
- ^ «Первые 3D-яйца птерозавров найдены вместе с их родителями» . физ.орг . Проверено 21 марта 2020 г.
- ^ Анвин, Дэвид Майкл; Диминг, Д. Чарльз (2019). «Пренатальное развитие птерозавров и его влияние на их постнатальную двигательную способность» . Труды Королевского общества B: Биологические науки . 286 (1904). дои : 10.1098/rspb.2019.0409 . ПМК 6571455 . ПМИД 31185866 .
- ^ Нэйш, Даррен; Виттон, Марк П.; Мартин-Сильверстоун, Элизабет (2021). «Полет с электроприводом у птерозавров: данные по форме крыльев и прочности костей» . Научные отчеты . 11 (1): 13130. Бибкод : 2021NatSR..1113130N . дои : 10.1038/s41598-021-92499-z . ПМЦ 8298463 . ПМИД 34294737 .
- ^ Ян, Цзысяо; Цзян, Баоюй; Бентон, Майкл Дж.; Сюй, Син; Макнамара, Мария Э.; Хоун, Дэвид ВЕ (26 июля 2023 г.). «Рост аллометрических крыльев связывает родительскую заботу с гигантизмом птерозавров» . Труды Королевского общества B: Биологические науки . 290 (2003). дои : 10.1098/rspb.2023.1102 . ISSN 0962-8452 . ПМЦ 10354479 . PMID 37464754 .
- ^ Бристоль, Университет. «Июль: Родители птерозавров | Новости и статьи | Бристольский университет» . www.bristol.ac.uk . Проверено 22 августа 2023 г.
- ^ Шмитц, Л.; Мотани, Р. (2011). «Ночной образ жизни динозавров, выведенный на основании морфологии склерального кольца и орбиты». Наука . 332 (6030): 705–08. Бибкод : 2011Sci...332..705S . дои : 10.1126/science.1200043 . ПМИД 21493820 . S2CID 33253407 .
- ^ Jump up to: а б с Хон, Д. (2010). «Птерозавры в популярной культуре». Pterosaur.net , по состоянию на 27 августа 2010 г.
- ^ Фрей Э., Мартилл Д. и Бьючи М. (2003). «Новый хохлатый орнитохейрид из нижнего мела на северо-востоке Бразилии и необычная смерть необычного птерозавра» в: Буффето, Э., и Мазин, Ж.-М. (ред.). Эволюция и палеобиология птерозавров . Специальная публикация Геологического общества 217 : 56–63. ISBN 1-86239-143-2 .
- ^ Берри 2005 , с. 452.
- ^ Jump up to: а б Томас, HN (2020). «Рожденный из огня: птерозаврологический анализ Родана» . Журнал компьютерных исследований 7 : 53–59.
- ^ Гонсалес, Дэйв (12 октября 2016 г.). «Монстровская разбивка каждого безумного фильма о Годзилле» . Триллерист . Проверено 11 июля 2019 г.
- ^ Шарф, Зак (10 декабря 2018 г.). « Трейлер «Годзиллы: Король монстров» превращает Мотру, Родана и многих других в эпическое зрелище» (видео) . ИндиВайр . Проверено 11 июля 2019 г.
- ^ Мансур, Дэвид (2005). От Abba до Zoom Энциклопедия поп-культуры конца 20 века . Издательство Эндрюса МакМила. п. 272.
- ^ Чжоу, X.; Пегас, Р.В.; Ма, В.; Вешать.; Джин, X.; Леал, MEC; Бонде, Н.; Кобаяши, Ю.; Лаутеншлагер, С.; Вэй, X.; Шен, К.; Джи, С. (2021). «Новый дарвиноптеран-птерозавр демонстрирует древесный реализм и противоположный большой палец» . Современная биология . 31 (11): 2429–2436.e7. Бибкод : 2021CBio...31E2429Z . дои : 10.1016/j.cub.2021.03.030 . ПМИД 33848460 .
Источники
[ редактировать ]- Берри, Марк Ф. (2005). Фильмография Динозавра . МакФарланд и компания. ISBN 978-0-7864-2453-5 .
- Веллнхофер, Питер (1991). Иллюстрированная энциклопедия птерозавров: иллюстрированная естественная история летающих рептилий мезозойской эры . Книги Полумесяца. ISBN 978-0-517-03701-0 .
- Виттон, Марк (2013). Птерозавры: естественная история, эволюция, анатомия . Издательство Принстонского университета. ISBN 978-0-691-15061-1 .
Внешние ссылки
[ редактировать ]- Pterosaur.net , сайт с участием нескольких авторов, посвященный всем аспектам науки о птерозаврах.
- База данных птерозавров , автор Пол Персглов
- «Комментарии к филогении птеродактилодея» Александра В.А. Келлнера (технический)
















