Водное передвижение

Водное передвижение или плавание среде с биологическим — это движение в жидкой приводом . Простейшие двигательные системы состоят из ресничек и жгутиков . Плавание неоднократно развивалось у целого ряда организмов, включая членистоногих , рыб , моллюсков , амфибий , рептилий , птиц и млекопитающих .
Эволюция плавания
[ редактировать ]
Плавание развивалось несколько раз в несвязанных линиях. Предполагаемые окаменелости медуз встречаются в Эдиакарском периоде , но первые свободно плавающие животные появляются в раннем и среднем кембрии . Они в основном связаны с членистоногими и включают Anomalocaridids , которые плавали с помощью боковых лопастей, напоминая сегодняшних каракатиц . к числу активных пловцов ( нектона ) пополнились головоногие моллюски. В позднем кембрии [1] а хордовые, вероятно, заплыли из раннего кембрия. [2] Многие наземные животные сохраняют некоторую способность плавать, однако некоторые вернулись в воду и развили способности к водному передвижению. Однако большинство обезьян (включая человека) утратили инстинкт плавания . [3]
В 2013 году Педро Ренато Бендер, научный сотрудник Института эволюции человека Университета Витватерсранда , предложил теорию, объясняющую утрату этого инстинкта. Названная гипотезой последнего общего предка Сачи (в честь Саси , бразильского фольклорного персонажа, который не может пересекать водные преграды), она утверждает, что потеря инстинктивной способности плавания у обезьян лучше всего объясняется как следствие ограничений, связанных с адаптацией к древесной жизни у обезьян. последний общий предок обезьян. [4] Бендер предположил, что предки обезьян все чаще избегали глубоководных водоемов, когда риск подвергнуться воздействию воды был явно выше, чем преимущества от их пересечения. [4] Уменьшение контакта с водоемами тогда могло привести к исчезновению собачьего инстинкта весла . [4]
Микроорганизмы
[ редактировать ]Микробные пловцы, иногда называемые микропловцами , представляют собой микроскопические существа, способные перемещаться в жидкой или водной среде. [5] Естественные микропловцы встречаются повсюду в мире природы в виде биологических микроорганизмов , таких как бактерии , археи , протисты , сперматозоиды и микроживотные .
Бактериальный
[ редактировать ]Инфузории
[ редактировать ]Инфузории используют маленькие жгутики, называемые ресничками, для передвижения по воде. Одна инфузория обычно имеет от сотен до тысяч ресничек, плотно упакованных вместе в массивы. Во время движения отдельная ресничка деформируется с помощью рабочего хода с высоким коэффициентом трения, за которым следует восстановительный ход с низким коэффициентом трения. Поскольку в отдельном организме имеется множество ресничек, упакованных вместе, они демонстрируют коллективное поведение в метахрональном ритме . Это означает, что деформация одной реснички синфазна с деформацией соседней, вызывая волны деформации, распространяющиеся по поверхности организма. Эти распространяющиеся волны ресничек позволяют организму скоординировано использовать реснички для движения. Типичным примером реснитчатого микроорганизма является Paramecium , одноклеточное реснитчатое простейшее, покрытое тысячами ресничек. Сбивающиеся вместе реснички позволяют Paramecium двигаться в воде со скоростью 500 микрометров в секунду. [6]


Жгутиконосцы
[ редактировать ]У некоторых организмов, таких как бактерии и сперма животных, есть жгутики , которые научились передвигаться в жидкой среде. Модель роторного двигателя показывает, что бактерии используют протоны электрохимического градиента для перемещения своих жгутиков. Крутящий момент в жгутиках бактерий создается частицами, которые проводят протоны вокруг основания жгутика. Направление вращения жгутиков у бактерий обусловлено заполненностью протонных каналов по периметру жгутикового мотора. [7]
Движение сперматозоидов называется подвижностью сперматозоидов . В середине сперматозоида млекопитающих содержатся митохондрии, обеспечивающие движение жгутика сперматозоида. Двигатель вокруг основания создает крутящий момент, как у бактерий при движении в водной среде. [8]
Псевдоподии
[ редактировать ]Движение с помощью псевдоподии осуществляется за счет увеличения давления в одной точке клеточной мембраны . Это увеличение давления является результатом полимеризации актина между корой и мембраной. По мере увеличения давления клеточная мембрана выталкивается наружу, образуя ложноножку. Когда ложноножка движется наружу, остальная часть тела вытягивается вперед за счет коркового напряжения. Результатом является движение клеток через жидкую среду. Кроме того, направление движения определяется хемотаксисом . Когда хемоаттракция происходит в определенной области клеточной мембраны , может начаться полимеризация актина , которая переместит клетку в этом направлении. [9] Прекрасным примером организма, использующего ложноножки, является Naegleria fowleri . [10]
Беспозвоночные
[ редактировать ]
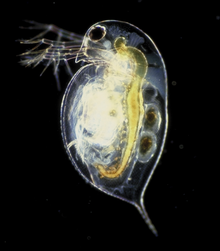
Среди лучистых медуз и их родственников основной формой плавания является сгибание их чашеобразных тел. Все медузы свободно плавают, хотя многие из них проводят большую часть времени пассивно, плавая. Пассивное плавание похоже на скольжение; организм плавает, используя потоки там, где это возможно, и не затрачивает никакой энергии на управление своим положением или движением. Активное плавание, напротив, предполагает расход энергии для перемещения в желаемое место.
У билатерий существует множество способов плавания. Черви -стрелы ( chaetognatha ) имеют волнистые плавники, мало чем отличаясь от рыб. Нематоды плавают, покачивая своим телом без плавников. Некоторые группы членистоногих умеют плавать, в том числе многие ракообразные . Большинство ракообразных, таких как креветки , обычно плавают, гребля специальными плавательными ногами ( плеоподами ). Крабы-плаватели плавают с видоизмененными шагающими ногами ( переоподами ). Дафния , ракообразное, вместо этого плавает, тряся усиками.
Существует также ряд форм плавающих моллюсков . Многие свободно плавающие морские слизни , такие как морские ангелы , имеют плавниковые структуры. Некоторые моллюски с панцирем, такие как гребешки, могут ненадолго плавать, открывая и закрывая две раковины. Наиболее развитыми для плавания моллюсками являются головоногие моллюски . Фиолетовые морские улитки используют плавучий пенопластовый плот, стабилизированный амфифильными муцинами , чтобы плавать на поверхности моря. [11] [12]
Среди Deuterostomia также есть некоторое количество пловцов. Перьевые звезды могут плавать, размахивая своими многочисленными руками. Сальпы передвигаются, прокачивая воду через свои студенистые тела. Второротые, наиболее развитые для плавания, встречаются среди позвоночных , особенно у рыб .
Реактивное движение
[ редактировать ]


Реактивное движение — это метод водного передвижения, при котором животные заполняют мышечную полость и выплескивают воду, чтобы подтолкнуть их в направлении, противоположном направлению брызг воды. Большинство организмов оснащены одной из двух конструкций реактивного движения; они могут черпать воду сзади и выбрасывать ее сзади, как медузы, или черпать воду спереди и выбрасывать ее сзади, как сальпы. Заполнение полости приводит к увеличению как массы, так и сопротивления животного. Из-за обширности сжимающейся полости скорость животного колеблется при движении по воде: ускоряясь при выбрасывании воды и замедляясь при всасывании воды пылесосом. Несмотря на то, что эти колебания сопротивления и массы можно игнорировать, если частота циклов реактивного движения достаточно высока, реактивное движение является относительно неэффективным методом передвижения в воде.
Все головоногие моллюски могут передвигаться с помощью реактивного движения , но это очень энергозатратный способ передвижения по сравнению с хвостовым движением, используемым рыбами. [13] Относительная эффективность реактивного движения снижается по мере увеличения размера животного. Начиная с палеозоя, когда конкуренция с рыбами создала среду, в которой эффективное движение имело решающее значение для выживания, реактивное движение отошло на второй план, а плавники и щупальца использовались для поддержания постоянной скорости. [14] Однако движение «стоп-старт», обеспечиваемое форсунками, по-прежнему полезно для обеспечения высокоскоростных рывков – не в последнюю очередь при поимке добычи или уклонении от хищников. [14] Действительно, это делает головоногих моллюсков самыми быстрыми морскими беспозвоночными. [15] : Предисловие и они могут ускорить большинство рыб. [16] Окисленная вода забирается в мантийную полость к жабрам и посредством мышечного сокращения этой полости выбрасывается израсходованная вода через гипоном , образованный складкой мантии. Движение головоногих моллюсков обычно происходит назад, поскольку вода вытесняется вперед через гипоном, но направление можно в некоторой степени контролировать, направляя ее в разные стороны. [17] Большинство головоногих моллюсков плавают (то есть имеют нейтральную плавучесть ), поэтому им не нужно плавать, чтобы оставаться на плаву. [13] Кальмары плавают медленнее, чем рыбы, но используют больше энергии для создания скорости. Потеря эффективности связана с количеством воды, которое кальмар может ускорить из мантийной полости. [18]
Медузы используют одностороннюю конструкцию водной полости, которая генерирует фазу непрерывных циклов реактивного движения, за которой следует фаза покоя. Эффективность Фруда составляет около 0,09, что указывает на очень дорогостоящий метод передвижения. Метаболические затраты на транспортировку медуз высоки по сравнению с рыбой такой же массы.
Другие животные с реактивными двигателями имеют аналогичные проблемы с эффективностью. Гребешки , конструкция которых аналогична медузам, плавают, быстро открывая и закрывая свои раковины, которые втягивают воду и выбрасывают ее со всех сторон. Это передвижение используется как средство спасения от хищников, таких как морские звезды . После этого раковина действует как подводное крыло, противодействуя склонности гребешка тонуть. Эффективность Фруда для этого типа движения невысока, около 0,3, поэтому его используют как механизм аварийного спасения от хищников. Однако объем работы, которую приходится выполнять гребешку, смягчается эластичным шарниром, соединяющим две раковины двустворчатого моллюска. Кальмары плавают, втягивая воду в мантийную полость и выбрасывая ее через сифон. КПД Фруда их реактивной двигательной установки составляет около 0,29, что значительно ниже, чем у рыбы той же массы.
Большая часть работы, выполняемой мышцами гребешка по закрытию раковины, сохраняется в виде упругой энергии в отводящей ткани, которая действует как пружина, открывая раковину. Эластичность приводит к тому, что работа, совершаемая против воды, становится низкой из-за больших отверстий, в которые вода должна войти, и маленьких отверстий, через которые вода должна выйти. Инерционная работа гребешкового реактивного движителя также невелика. Из-за малой инерционной работы экономия энергии, создаваемая эластичной тканью, настолько мала, что ею можно пренебречь. Медузы также могут использовать свою эластичную мезоглею, чтобы увеличить свой колокол. Их мантия содержит слой мышц, зажатый между эластичными волокнами. Мышечные волокна проходят вокруг колокола по окружности, а эластические волокна проходят через мышцу и по бокам колокола, чтобы предотвратить удлинение. После однократного сокращения колокольчик пассивно вибрирует на резонансной частоте, чтобы снова наполнить колокол. Однако, в отличие от гребешков, инерционная работа аналогична гидродинамической за счет того, как медузы выбрасывают воду – через большое отверстие с малой скоростью. Из-за этого отрицательное давление, создаваемое колеблющейся полостью, ниже положительного давления струи, а значит, инерционная работа мантии мала. Таким образом, водометное движение оказывается неэффективным методом плавания. [18]
Рыба
[ редактировать ]

Многие рыбы плавают в воде, создавая волны своим телом или покачивая плавниками . Волнистость создает компоненты тяги вперед, дополненные силой, направленной назад, боковыми силами, которые представляют собой потраченную впустую часть энергии, и нормальной силой, которая находится между тягой вперед и боковой силой. Разные рыбы плавают, двигая разными частями тела. Рыбы в форме угрей ритмично колеблются всем своим телом. У обтекаемых рыб, таких как лосось, хвостовая часть тела волнистая. Некоторые рыбы, например акулы, используют жесткие и сильные плавники, чтобы создавать динамическую подъемную силу и двигаться вперед. Рыбы обычно используют более одного способа движения, хотя они демонстрируют один доминирующий способ плавания. [19] Изменения походки наблюдались даже у молоди рифовых рыб разных размеров. В зависимости от своих потребностей рыбы могут быстро переключаться между синхронными и попеременными движениями плавников. [20]
Согласно Книге рекордов Гиннеса 2009 года , Hippocampus zosterae (карликовый морской конёк) — самая медленная рыба, максимальная скорость которой составляет около 5 футов (150 см) в час. [21] Они очень плохо плавают, быстро взмахивая спинным плавником и используя грудные плавники (расположенные за глазами) для управления. У морских коньков нет хвостового плавника .
Двигательная установка «Тело-хвостовой плавник» (BCF)
[ редактировать ]- Ангилиформные: Ангилиформные пловцы обычно медленные пловцы. Они двигают большую часть своего тела волнообразно и используют голову как точку опоры для груза, который они перемещают. В любой момент волн их тело имеет амплитуду 0,5–1,0 длины волны. Амплитуда, которую они перемещают своим телом, позволяет им плыть назад. Ангилиформное передвижение обычно наблюдается у рыб с длинными тонкими телами, таких как угри , миноги , весла и ряд видов сомов .
- Подгульчатые, карангиформные, тунниформные: эти пловцы колеблют заднюю половину своего тела и намного быстрее, чем ангилиформные пловцы. В любой момент во время плавания на волнообразном рисунке тела можно увидеть длину волны <1. Среди некоторых карангиформных пловцов есть акулы-няньки, бамбуковые акулы и рифовые акулы. Тоннообразные пловцы очень быстрые, и среди наиболее распространенных тунниформных пловцов можно назвать тунца, белых акул, лосося, джеков и акул мако. [ нужна ссылка ] . Тонкообразные пловцы колеблют только хвостовой плавник с большим удлинением , поэтому они обычно очень жесткие, чтобы отталкивать больше воды.
- Острациеформные: пловцы острациевидной формы колеблют хвостовую область, что делает их относительно медленными пловцами. Самшит, скаты-торпеды и момыры используют острациеобразную локомоцию. Рыба-корова использует Osctraciiform движения, чтобы парить в толще воды. [22]
Движительная установка срединного парного плавника (MPF)
[ редактировать ]- Tetraodoniform, Balistiform, Diodontiform: эти пловцы колеблют свои срединные плавники. Обычно они медленно плавают, и некоторые известные примеры включают океаническую солнечную рыбу (у которой чрезвычайно модифицированы анальный и спинной плавники), рыбу-фугу и спинорога.
- Rajiform, Amiiform, Gymnotiform: этот двигательный режим достигается за счет волнистости грудных и срединных плавников. Во время их волнообразного рисунка в их плавниках можно увидеть длину волны>1. Обычно они плавают медленно или умеренно, а некоторые примеры включают скатов, боуфинов и рыб-ножей. Черная рыба-призрак-нож — гимнотиформный пловец с очень длинным брюшным ленточным плавником. Тяга создается за счет прохождения волн по ленточному плавнику, в то время как тело остается жестким. Это также позволяет рыбе-призраку-ножу плавать задним ходом.
- Labriform: Пловцы Labriform также являются медленными пловцами. Они колеблют грудные плавники, создавая тягу. Колеблющиеся плавники создают тягу, когда начальный вихрь выходит из задней кромки плавника. Когда фольга отходит от исходного вихря, эффект этого вихря уменьшается, но связанная циркуляция сохраняется, создавая подъемную силу. Лабриформное плавание можно рассматривать как непрерывное начало и остановку. Губаны и серфинговые окуни — обычные пловцы Labriform. [22]
Суда на подводных крыльях
[ редактировать ]
Подводные крылья или плавники используются для отталкивания воды, чтобы создать нормальную силу, обеспечивающую тягу, продвигающую животное через воду. Морские черепахи и пингвины машут своими парными подводными крыльями, чтобы создать подъемную силу. Некоторые парные плавники, например грудные плавники леопардовых акул, могут располагаться под разным углом, что позволяет животному подниматься, опускаться или сохранять свое положение в толще воды. Уменьшение площади поверхности плавников помогает минимизировать сопротивление и, следовательно, повысить эффективность. Независимо от размера животного, при любой конкретной скорости максимально возможная подъемная сила пропорциональна (площадь крыла) х (скорость). 2 . У дельфинов и китов есть большие горизонтальные хвостовые крылья на подводных крыльях, тогда как у многих рыб и акул есть вертикальные хвостовые крылья на подводных крыльях. Морские свиньи (наблюдаемые у китообразных, пингвинов и ластоногих) могут экономить энергию, если они двигаются быстро. Поскольку сопротивление увеличивается со скоростью, работа, необходимая для преодоления единицы расстояния, увеличивается на более высоких скоростях, но работа, необходимая для преодоления единицы расстояния, не зависит от скорости. Тюлени движутся по воде с помощью хвостового хвоста, а морские львы создают толчок исключительно с помощью грудных ласт. [19]
Плавание с дрэг-приводом
[ редактировать ]
Как и при движении через любую жидкость, при столкновении молекул жидкости с организмом возникает трение. Столкновение вызывает сопротивление движущейся рыбы, поэтому многие рыбы имеют обтекаемую форму. Обтекаемые формы уменьшают сопротивление, ориентируя удлиненные объекты параллельно силе сопротивления, что позволяет течению проходить через конец рыбы и сужать его. Эта обтекаемая форма позволяет более эффективно использовать энергию движения. Некоторые рыбы плоской формы могут воспользоваться сопротивлением давления, имея плоскую нижнюю поверхность и изогнутую верхнюю поверхность. Создаваемое сопротивление давления позволяет поднять рыбу вверх. [19]
Придатки водных организмов приводят их в движение двумя основными биомеханически крайними механизмами. Некоторые используют плавание с подъемной силой, которое можно сравнить с полетом, поскольку придатки машут, как крылья, и уменьшают сопротивление на поверхности придатков. Другие используют плавание с приводом от сопротивления, которое можно сравнить с греблей в лодке на веслах с движением в горизонтальной плоскости или греблей с движением в парасагиттальной плоскости.
Пловцы-дрэг используют циклическое движение, при котором они отталкивают воду назад во время рабочего гребка и возвращают конечность вперед во время обратного или восстановительного гребка. Когда они толкают воду прямо назад, это перемещает их тело вперед, но когда они возвращают конечности в исходное положение, они толкают воду вперед, что, таким образом, в некоторой степени тянет их назад и, таким образом, противодействует направлению движения тела. Эта противодействующая сила называется сопротивлением . Сопротивление обратного хода заставляет пловцов-дрэг-пловцов использовать иные стратегии, чем пловцов-подъемников. Уменьшение сопротивления при обратном ходе имеет важное значение для оптимизации эффективности. Например, утки гребут по воде, расправляя перепонки на ногах, когда они отводят воду назад, а затем, когда они возвращают ноги вперед, они стягивают перепонки вместе, чтобы уменьшить последующее притяжение воды вперед. На ногах водяных жуков есть маленькие волоски, которые расправляются, чтобы улавливать и возвращать воду при обратном ходе, но лежат ровно, когда придаток движется вперед при обратном ходе. Кроме того, одна сторона ноги водяного жука шире других и удерживается перпендикулярно движению при движении назад, но нога вращается, когда конечность возвращается вперед, поэтому более тонкая сторона захватывает меньше воды. [23]
У дрэг-пловцов снижается эффективность плавания из-за сопротивления, которое влияет на их оптимальную скорость. Чем меньшее сопротивление испытывает рыба, тем больше она сможет поддерживать более высокие скорости. Морфологию рыбы можно изменить таким образом, чтобы уменьшить сопротивление, например, за счет обтекаемости тела. Стоимость транспорта у дрэг-пловец значительно выше, и при отклонении от оптимальной скорости дрэг-пловец энергетически напрягается гораздо сильнее, чем пловец-подъемник. Существуют естественные процессы оптимизации использования энергии, и считается, что корректировка скорости метаболизма может частично компенсировать механические недостатки. [24]
У полуводных животных по сравнению с полностью водными животными наблюдается усиление сопротивления. Конструкция, позволяющая им работать вне воды, ограничивает эффективность, которую можно достичь в воде. Плавание в воде на поверхности подвергает их воздействию резистивного волнового сопротивления и связано с более высокими затратами, чем плавание под водой. Плавание под водой подвергает их сопротивлению из-за обратных ударов и давления, но в первую очередь трения. Сопротивление трения обусловлено вязкостью жидкости и морфологическими характеристиками. Сопротивление давлению обусловлено разницей потока воды вокруг тела, а также зависит от морфологии тела. Полуводные организмы сталкиваются с повышенными силами сопротивления, находясь в воде или вне ее, поскольку они не приспособлены ни к одной среде обитания. Например, морфология выдр и бобров должна отвечать потребностям обеих сред. Их мех уменьшает обтекаемость и создает дополнительное сопротивление. Утконос может быть хорошим примером промежуточного звена между пловцами-перетаскивателями и пловцами с подъемом, поскольку было показано, что у него есть механизм гребли, аналогичный грудным колебаниям, основанным на подъеме. Конечности полуводных организмов предназначены для использования на суше, и использование их в воде не только увеличивает стоимость передвижения, но и ограничивает их режимами, основанными на перетаскивании. [25] Хотя они менее эффективны, пловцы-дрэг могут развивать большую тягу на низких скоростях, чем пловцы-подъемники. Также считается, что они обладают большей маневренностью из-за большей тяги. [26]
Земноводные
[ редактировать ]
Большинство амфибий имеют личиночное состояние, унаследовавшее от предков рыб ангиловидное движение и сопутствующий ему сжатый с боков хвост.Соответствующие взрослые формы четвероногих, даже у сохраняющего хвост подкласса Urodeles , иногда ведут водный образ жизни лишь в незначительной степени (как в роде Salamandra , хвост которой утратил пригодность к водному движению), но большинство Urodeles , от тритонов до гигантской саламандры Megalobatrachus,сохраняют сжатый с боков хвост для жизни, которая в значительной степени ведет водный образ жизни, который может использоваться в гуглевидных движениях. [27]
Из бесхвостых земноводных ( лягушки и жабы подкласса Anura ) большинство во взрослой жизни ведут водный образ жизни в незначительной степени, но у того значительного меньшинства, ведущих преимущественно водный образ жизни, мы впервые сталкиваемся с проблемой адаптации бесхвостых. конструкция тетрапода для водного движения. Режим, который они используют, не связан с режимом, используемым рыбами. Благодаря своим гибким задним ногам и перепончатым ступням они выполняют движения ног, близкие к человеческому «гребку брассом», но гораздо более эффективно, поскольку ноги имеют более обтекаемую форму. [27]
Рептилии
[ редактировать ]
С точки зрения водного движения происхождение современных представителей класса Reptilia от архаичных хвостатых амфибий наиболее очевидно в случае отряда Crocodilia ( крокодилы и аллигаторы ), которые используют свои глубокие, сжатые с боков хвосты в форме каракуля. способ движения (см. Передвижение рыб#Carangiform ). [27]
Наземные змеи , несмотря на свою «плохую» гидромеханическую форму с примерно круглым поперечным сечением и постепенным сужением задней части, при необходимости плавают довольно легко, используя ангиллиформные движения (см. Локомоция рыб#Anguilliform ). [27]
Cheloniidae (морские черепахи ) нашли решение проблемы плавания четвероногих за счет развития их передних конечностей в ласты с большим удлинением формы крыла, с помощью которых они более точно имитируют двигательный режим птицы, чем сами скаты-орляки. [ нужна ссылка ]


Движение плавников и ласт
[ редактировать ]
Водные рептилии, такие как морские черепахи (см. также черепахи ) и вымершие виды, такие как плиозавроиды, преимущественно используют грудные ласты, чтобы двигаться по воде, и тазовые ласты для маневрирования. грудными ластами Во время плавания они совершают дорзо-вентральные движения , вызывая движение вперед. Во время плавания они вращают передние ласты, чтобы уменьшить сопротивление в толще воды и повысить эффективность. Только что вылупившиеся морские черепахи демонстрируют несколько поведенческих навыков, которые помогают им ориентироваться в океане, а также определять переход от песка к воде. При вращении по тангажу, рысканию или крену вылупившиеся птенцы способны противодействовать действующим на них силам, корректируя грудные или тазовые ласты и перенаправляясь в сторону открытого океана.
Среди млекопитающих отарииды ( морские котики ) плавают преимущественно передними ластами, используя для управления задние ласты, а фоциды ( настоящие тюлени ) перемещают задние ласты вбок, толкая животное в воде.
Реакции побега
[ редактировать ]
Некоторые членистоногие, такие как омары и креветки , могут быстро двигаться назад, взмахивая хвостом, что известно как омаровая реакция или каридоидная реакция побега .
Разновидности рыб, например костистые, также используют быстрый старт, чтобы спастись от хищников. Быстрый старт характеризуется сокращением мышц на одной стороне рыбы, скручивающим рыбу в С-образную форму. После этого на противоположной стороне происходит сокращение мышц, что позволяет рыбе войти в состояние устойчивого плавания, а волны волнообразно распространяются вдоль тела. Сила изгибающего движения исходит от быстросокращающихся мышечных волокон, расположенных в центральной части рыбы. Сигнал на выполнение этого сокращения исходит от набора клеток Маутнера , которые одновременно посылают сигнал мышцам на одной стороне рыбы. [28] Клетки Маутнера активируются, когда что-то пугает рыбу, и их можно активировать зрительным путем. [29] или звуковые стимулы.
Быстрый старт разбит на три этапа. Первый этап, называемый подготовительным ходом, характеризуется начальным изгибом в С-образную форму с небольшой задержкой, вызванной гидродинамическим сопротивлением. Второй этап, пропульсивный гребок, включает в себя быстрый наклон тела в другую сторону, что может происходить несколько раз. На третьем этапе, фазе отдыха, рыба возвращается к нормальному устойчивому плаванию, и колебания тела начинают прекращаться. Крупные мышцы, расположенные ближе к центральной части рыбы, сильнее и производят больше силы, чем мышцы хвоста. Эта асимметрия в составе мышц вызывает волнистость тела, которая возникает на стадии 3. Было показано, что после завершения быстрого старта положение рыбы имеет определенный уровень непредсказуемости, что помогает рыбе выжить против хищников. [29]
Скорость, с которой тело может сгибаться, ограничена сопротивлением, заключенным в инерции каждой части тела. Однако эта инерция помогает рыбе создавать движение в результате импульса, создаваемого против воды. Движение вперед, создаваемое C-стартом, и устойчивое плавание в целом является результатом удара тела рыбы о воду. Волны волн создают обратный импульс против воды, обеспечивая толчок вперед, необходимый для подталкивания рыбы вперед. [30]
Эффективность
[ редактировать ]Эффективность движения Фруда определяется как отношение выходной мощности к входной мощности:
nf знак равно 2 U 1 / ( U 1 + U 2 )
где U1 = скорость набегающего потока и U2 = скорость струи. Хороший КПД для гуглевидной тяги составляет от 50 до 80%. [19]
Минимизация сопротивления
[ редактировать ]Перепады давления возникают вне пограничного слоя плавающих организмов из-за нарушения обтекания тела. Разница на поверхностях тела, расположенных выше и ниже по потоку, представляет собой сопротивление давления , которое создает силу, действующую вниз по потоку на объект. С другой стороны, сопротивление трения является результатом вязкости жидкости в пограничном слое . Более высокая турбулентность вызывает большее сопротивление трения. [18]
Число Рейнольдса (Re) является мерой соотношения между инерционными и вязкими силами в потоке ((длина животного x скорость животного)/кинематическая вязкость жидкости). Турбулентный поток можно обнаружить при более высоких значениях Re, когда пограничный слой отделяется и создает след, а ламинарный поток можно обнаружить при более низких значениях Re, когда отделение пограничного слоя задерживается, уменьшая потери следа и кинетической энергии на противоположный импульс воды. [31]
Форма тела плавающего организма влияет на результирующее сопротивление. Длинные, тонкие корпуса уменьшают сопротивление давлению за счет обтекаемости, а короткие круглые корпуса уменьшают сопротивление трения; следовательно, оптимальная форма организма зависит от его ниши. Плавающие организмы веретенообразной формы, вероятно, испытают наибольшее снижение как давления, так и сопротивления трения. [31]
Форма крыла также влияет на величину сопротивления, испытываемого организмом, поскольку при различных методах гребка восстановление положения перед гребком приводит к накоплению сопротивления.
Высокоскоростная напорная вентиляция создает ламинарный поток воды от жабр по телу организма.
Секреция слизи вдоль поверхности тела организма или добавление длинноцепочечных полимеров к градиенту скорости могут уменьшить сопротивление трения, испытываемое организмом.
Плавучесть
[ редактировать ]Многие водные/морские организмы развили органы, позволяющие компенсировать свой вес и контролировать плавучесть в воде. Эти структуры делают плотность их тел очень близкой к плотности окружающей воды. Некоторые гидрозойные, например сифонофоры, имеют газонаполненные поплавки; Наутилус, Сепия и Спирула ( головоногие моллюски ) имеют внутри своих раковин газовые камеры; а большинство костистых рыб и многие рыбы-фонарики (Myctophidae) оснащены плавательными пузырями . Многие водные и морские организмы также могут состоять из материалов низкой плотности. Глубоководные костистые кости, не имеющие плавательного пузыря, имеют мало липидов и белков, глубоко окостеневшие кости и водянистые ткани, сохраняющие плавучесть. Печень некоторых акул состоит из липидов низкой плотности, таких как углеводородный сквален или эфиры воска (также встречаются у Myctophidae без плавательного пузыря), которые обеспечивают плавучесть. [19]
Плавающие животные, плотнее воды, должны создавать подъемную силу или адаптироваться к донному образу жизни. Движение рыбы для создания гидродинамической подъемной силы необходимо для предотвращения погружения. Часто их тела действуют как подводные крылья , и эта задача более эффективна для рыб с плоским телом. При небольшом угле наклона подъемная сила у плоской рыбы больше, чем у рыбы с узким телом. Рыбы с узким телом используют свои плавники как подводные крылья, в то время как их тело остается горизонтальным. У акул гетероцеркальная форма хвоста направляет воду вниз, создавая противодействующую восходящую силу, толкая акулу вперед. Создаваемому подъему способствуют грудные плавники и положение тела под углом вверх. Предполагается, что тунцы в основном используют грудные плавники для подъема. [19]
Поддержание плавучести требует метаболических затрат. Выращивание и поддержание органа плавучести, корректировка биологического состава и физическое напряжение, чтобы оставаться в движении, требуют большого количества энергии. Предполагается, что подъемную силу можно физически генерировать с меньшими затратами энергии за счет плавания вверх и скольжения вниз в движении «подъём и скольжение», а не постоянного плавания по плоскости. [19]
Температура
[ редактировать ]Температура также может сильно влиять на способность водных организмов передвигаться в воде. Это связано с тем, что температура влияет не только на свойства воды, но и на обитающие в ней организмы, поскольку большинство из них имеют идеальный диапазон, специфичный для их тела и метаболических потребностей.
Q10 (температурный коэффициент) — коэффициент, на который скорость увеличивается при повышении температуры на 10 °C, используется для измерения того, как работоспособность организмов зависит от температуры. У большинства из них скорость увеличивается по мере того, как вода становится теплее, но у некоторых есть пределы этому, а другие находят способы изменить такие эффекты, например, за счет эндотермии или более раннего набора более быстрых мышц. [32]
Например, было обнаружено, что Crocodylus porosus , или эстуарные крокодилы, увеличивают скорость плавания с 15 °C до 23 °C, а затем достигают пиковой скорости плавания с 23 °C до 33 °C. Однако производительность начала снижаться по мере того, как температура поднималась выше этой точки, показывая предел диапазона температур, при которых этот вид мог бы идеально работать. [33]
Погружение
[ редактировать ]Чем большая часть тела животного погружена в воду во время плавания, тем меньше энергии оно расходует. Плавание на поверхности требует в два-три раза больше энергии, чем при полном погружении. Это происходит из-за носовой волны, которая образуется спереди, когда животное во время плавания толкает поверхность воды, создавая дополнительное сопротивление. [34]
Вторичная эволюция
[ редактировать ]

Хотя четвероногие утратили многие из своих естественных приспособлений к плаванию, когда они эволюционировали на сушу, многие из них вновь обрели способность плавать или действительно вернулись к полностью водному образу жизни.
В основном или исключительно водные животные неоднократно эволюционировали из наземных четвероногих: примеры включают земноводных , таких как тритоны , рептилий, таких как крокодилы , морские черепахи , ихтиозавры , плезиозавры и мозазавры , морских млекопитающих , таких как киты , тюлени и выдры , и птиц, таких как пингвины . Многие виды змей также являются водными и всю свою жизнь живут в воде. Среди беспозвоночных ряд видов насекомых обладают приспособлениями к водной жизни и передвижению. Примеры водных насекомых включают стрекоз личинки , водников и жуков-ныряльщиков . Существуют также водные пауки , хотя они, как правило, предпочитают другие способы передвижения под водой, чем собственно плавание.
Примеры: Некоторые породы собак плавают для развлечения. Умбра, собака-рекордсменка мира, может проплыть 4 мили (6,4 км) за 73 минуты, что ставит ее в число 25% лучших в соревнованиях по плаванию среди людей на длинные дистанции. [35] Кошка -рыболов — один из диких видов кошек, у которого развились особые приспособления для водного или полуводного образа жизни — перепончатые пальцы. Тигры и некоторые отдельные ягуары — единственные крупные кошки, которые, как известно, с готовностью заходят в воду, хотя наблюдалось плавание и других крупных кошек, в том числе львов. Некоторые породы домашних кошек также любят плавать, например, турецкий ван .
Лошади , лоси и лоси — очень сильные пловцы и могут преодолевать большие расстояния в воде. Слоны также способны плавать даже в глубоких водах. Очевидцы подтвердили, что верблюды , в том числе дромадеры и двугорбые верблюды, умеют плавать, [36] несмотря на то, что в их естественной среде обитания мало глубокой воды.
как домашние, так и дикие кролики Плавать умеют . Домашних кроликов иногда обучают плаванию в качестве циркового аттракциона. Дикий кролик, как известно, напал на лодку президента США Джимми Картера, когда ему угрожали в его естественной среде обитания. [37]
Морская свинка (или морская свинка) обладает отличными способностями к плаванию. [38] Мыши неплохо умеют плавать. Они впадают в панику, когда их помещают в воду, но многих лабораторных мышей используют в водном лабиринте Морриса — тесте для измерения обучаемости. Когда мыши плавают, они используют хвост как жгутики и пинают ногами.
Многие змеи также являются отличными пловцами. Крупные взрослые анаконды проводят большую часть времени в воде и с трудом передвигаются по суше.
Многие обезьяны умеют плавать от природы, а некоторые, например, хоботчатая обезьяна , крабоедная макака и макака-резус, плавают регулярно.
Человеческое плавание
[ редактировать ]Плавание известно людям с доисторических времен; Самое раннее упоминание о плавании относится к рисункам каменного века , сделанным примерно 7000 лет назад. Соревновательное плавание началось в Европе около 1800 года и было частью первых современных летних Олимпийских игр 1896 года в Афинах , хотя и не в форме, сопоставимой с современными соревнованиями. Лишь в 1908 году Международная федерация плавания ввела в действие правила проведения соревнований по плаванию . [39]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Крегер, Б.; Юн-бай, Ю.Б. (2009). «Импульсная диверсификация головоногих моллюсков в ордовике». Палеогеография, Палеоклиматология, Палеоэкология . 273 (1–2): 174–201. Бибкод : 2009PPP...273..174K . дои : 10.1016/j.palaeo.2008.12.015 .
- ^ Шу, Д.Г.; Конвей Моррис, С.; Хан, Дж.; Чжан, ZF; Ясуи, К.; Жанвье, П.; Чен, Л.; Чжан, XL; Лю, JN; Ли, Ю.; Лю, Х. -К. (2003), «Голова и позвоночник раннекембрийских позвоночных Haikouichthys » , Nature , 421 (6922): 526–529, Bibcode : 2003Natur.421..526S , doi : 10.1038/nature01264 , PMID 12556891 , S2CID 4401274
- ^ «Поведение в воде» . Центр академических исследований и обучения в области антропогении . Проверено 2 июля 2017 г.
- ^ Jump up to: Перейти обратно: а б с «Использование конвергенции как инструмента реконструкции человеческого прошлого с особым акцентом на использование воды в эволюции человекообразных» (PDF) . WIReDSpace (институциональный репозиторий Wits на DSpace) . Проверено 2 июля 2017 г.
- ^ Бунеа, Ада-Иоана; Табориски, Рафаэль (2020). «Последние достижения в области микропловцов для биомедицинских применений» . Микромашины . 11 (12): 1048. дои : 10,3390/ми11121048 . ПМЦ 7760273 . ПМИД 33261101 .
- ^ Лауга, Эрик; Томас Р. Пауэрс (25 августа 2009 г.). «Гидродинамика плавающих микроорганизмов». Отчеты о прогрессе в физике . 72 (9): 096601. arXiv : 0812.2887 . Бибкод : 2009RPPH...72i6601L . дои : 10.1088/0034-4885/72/9/096601 . S2CID 3932471 .
- ^ Брэди, Ричард М. (1993). «Крутящий момент и переключение бактериального жгутикового двигателя. Электростатическая модель» . Биофизический журнал . 64 (4): 961–973. Бибкод : 1993BpJ....64..961B . дои : 10.1016/S0006-3495(93)81462-0 . ПМЦ 1262414 . ПМИД 7684268 .
- ^ Мортимер, Шэрон Т. (1997). «Критический обзор физиологической важности и анализ движения сперматозоидов у млекопитающих» . Обновление репродукции человека . 3 (5): 403–439. дои : 10.1093/humupd/3.5.403 . ПМИД 9528908 .
- ^ член парламента Нилсон; Дж. А. Маккензи; С.Д. Уэббс; Р. Х. Инсолл (2011). «Моделирование движения клеток и хемотаксиса с использованием обратной связи на основе псевдоподий» (PDF) . Журнал SIAM по научным вычислениям . 33 (3/4): 1035–1057. Бибкод : 2011ГАО...33.1035Н . дои : 10.1137/100788938 .
- ^ Шустер, Флорида; Вишвешвара, Г.С. (2004). «Свободноживущие амебы как условно-патогенные и неоппортунистические патогены человека и животных». Международный журнал паразитологии . 34 (9): 1001–1027. дои : 10.1016/j.ijpara.2004.06.004 . ПМИД 15313128 .
- ^ Черчилль, Селия К.К.; О Фогил, Диармайд; Стронг, Эллен Э.; Гиттенбергер, Адриан (октябрь 2011 г.). «Самки первыми плавали в плавающих по пузырям улитках» . Современная биология . 21 (19): Р802–Р803. Бибкод : 2011CBio...21.R802C . дои : 10.1016/j.cub.2011.08.011 . ПМИД 21996498 .
- ^ Рюс, Патрик А.; Бергфройнд, Джотам; Берч, Паскаль; Гштёль, Стефан Дж.; Фишер, Питер (2021). «Сложные жидкости в стратегиях выживания животных» . Мягкая материя . 17 (11): 3022–3036. arXiv : 2005.00773 . Бибкод : 2021SMat...17.3022R . дои : 10.1039/D1SM00142F . ПМИД 33729256 .
- ^ Jump up to: Перейти обратно: а б Уилбур, Карл М.; Кларк, MR; Труман, ER, ред. (1985), «11: Эволюция плавучести и передвижения у современных головоногих моллюсков», The Mollusca , vol. 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ Jump up to: Перейти обратно: а б Уилбур, Карл М.; Кларк, MR; Труман, ER, ред. (1985), Моллюска , том. 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ Мэрион Никсон; Джей Зи Янг (2003). Мозг и жизнь головоногих моллюсков . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-852761-9 .
- ^ Дэниел Л. Гилберт; Уильям Дж. Адельман; Джон М. Арнольд (1990). Кальмары как экспериментальные животные . Дэниел Л. Гилберт, Джон М. Арнольд (иллюстрированный ред.). Спрингер. ISBN 978-0-306-43513-3 .
- ^ Кэмпбелл, Рис и Митчелл, стр.612.
- ^ Jump up to: Перейти обратно: а б с Левинтон, Джеффри С. Морская биология: функция, биоразнообразие, экология. 3-е изд. Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета, 2008. Печать.
- ^ Jump up to: Перейти обратно: а б с д и ж г Макнил Александр, Р. Изучение биомеханики: животные в движении. 2-е изд. Нью-Йорк, Нью-Йорк: WH Freeman & Co, 1992. Печать.
- ^ Хейл, Мелина, Р. Дэй, Д. Торсен, М. Вестнит. 2006. Координация грудных плавников и переходы походки у постоянно плавающей молоди рифовых рыб. Журнал экспериментальной биологии (209): 3708-3718.
- ^ Книга рекордов Гиннеса (2009)
- ^ Jump up to: Перейти обратно: а б Сфакиотакис, Майкл, Д. Лейн, Б. Дэвис. 1999. Обзор способов плавания рыб для передвижения в воде. Журнал океанической инженерии (24:2) 237-252.
- ^ Лаудер, Г., и Джейн, Б. Передвижение грудных плавников у рыб: тестирование моделей на основе сопротивления с использованием трехмерной кинематики. Оксфордские журналы. 2011.
- ^ Ольбергер, Дж., Г. Стаакс и Ф. Хокер. Эффективность плавания и влияние морфологии на затраты на плавание рыб. J Comp Physiol B. 176: 17-25. 2005.
- ^ Фиш, Фрэнк Э. Биомеханика и энергетика водных и полуводных млекопитающих: от утконоса до кита. Чикагские журналы. 2011.
- ^ Уокер, Дж., и М. Вестнит. Механические характеристики водной гребли и полета. Королевское общество. 10.1098. 2000.
- ^ Jump up to: Перейти обратно: а б с д Лайтхилл, М.Дж. Гидромеханика движения водных животных-Анну. Преподобный Жидкость. Мех. - 1969 1-413-446.
- ^ Уэйклинг, Джеймс М.; Джонстон, Ян А. (1999). «Изгиб тела рыбы во время быстрого старта можно объяснить с точки зрения мышечного момента и гидродинамического сопротивления». Журнал экспериментальной биологии . 202 (6): 675–682. дои : 10.1242/jeb.202.6.675 . ПМИД 10021321 .
- ^ Jump up to: Перейти обратно: а б Итон, Роберт С.; и др. (1977). «Реакция испуга у костистых рыб, инициированная Маутнером». Журнал экспериментальной биологии . 66 (1): 65–81. дои : 10.1242/jeb.66.1.65 . ПМИД 870603 .
- ^ Уэйклинг, Дж. М. «Биомеханика быстрого плавания рыб». Сравнительная биохимия и физиология, часть A.131 (2001): 31-40.
- ^ Jump up to: Перейти обратно: а б Фельдкамп, С.Д. «Плавание калифорнийского морского льва: морфометрия, сопротивление и энергетика». Дж. эксп. Биол. 131, 117–135 (1987).
- ^ Марцинек, Дэвид Дж.; Сюзанна Б. Блэквелл; Хайди Дьюар; Эллен В. Фройнд; Чарльз Фарвелл; Дэниел Дау; Эндрю К. Зейтц; Барбара А. Блок (1 апреля 2001 г.). «Глубина и температура мышц тихоокеанского голубого тунца исследованы с помощью акустических и всплывающих спутниковых архивных меток». Морская биология . 138 (4): 869–885. Бибкод : 2001МарБи.138..869М . дои : 10.1007/s002270000492 . S2CID 84807808 .
- ^ Элсворт, Питер Г.; Фрэнк Сибахер; Крейг Э. Франклин (апрель 2003 г.). «Устойчивое плавание крокодилов (Crocodylus porosus): влияние размера тела и температуры». Журнал герпетологии . 37 (2): 363–368. CiteSeerX 10.1.1.667.6790 . doi : 10.1670/0022-1511(2003)037[0363:sspicc]2.0.co;2 .
- ^ Александр, Дэвид Э. (17 ноября 2004 г.). Летчики природы: птицы, насекомые и биомеханика полета . Джу Пресс. ISBN 978-0-8018-8059-9 – через Google Книги.
- ^ «ВИДЕО ПЛАВАЮЩИХ СОБАК… Фон плавания» . www.sdogv.com . Архивировано из оригинала 24 августа 2017 г. Проверено 30 марта 2009 г.
- ^ «Натуральная почта с наркотиками: Настоящая почта с наркотиками: Верблюд — единственное животное, которое не умеет плавать?» .
- ^ «Сегодня в странной истории: на Джимми Картера напал кролик-убийца (20 апреля 1979 г.)» . www.newsoftheodd.com .
- ^ Харкнесс, Джон Э.; Вагнер, Джозеф Э. (1995). Биология и медицина кроликов и грызунов . Уильямс и Уилкинс. стр. 30–39. ISBN 978-0-683-03919-1 .
- ^ Кем, Г. (2007). Олимпийское плавание и прыжки в воду: великие моменты в олимпийской истории . Издательская группа Розен. стр. 14 . ISBN 978-1-4042-0970-1 .
| до |  | |
|---|---|---|
| Конечности | ||
| Крылья | ||
| Эволюция | ||
| Связанный | ||