Головоногие моллюски
| Головоногие моллюски Временной диапазон: | |
|---|---|
 | |
| Современные и вымершие головоногие моллюски; по часовой стрелке сверху слева: осьминог обыкновенный ( Octopus vulgaris ), карибский рифовый кальмар ( Sepioteuthis sepioidea ), наутилус с камерами ( Nautilus pompilius ), Orthosphynctes , Clarkeiteuthis conocauda и каракатица обыкновенная ( Sepia officinalis ). | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Моллюска |
| Подтип: | Конхифера |
| Сорт: | Головоногие моллюски Кювье , 1797 г. |
| Подклассы | |
| |
Головоногие моллюски / ˈ s ɛ f ə l ə p ɒ d / — любой представитель моллюсков класса Головоногие / s ɛ f ə ˈ l ɒ p ə d ə / ( греческое множественное число κεφαλόποδες , головоногие ; «голова-ноги») [3] например , кальмар , осьминог , каракатица или наутилус . Эти исключительно морские животные характеризуются двусторонней симметрией тела , выдающейся головой и набором рук или щупалец ( мышечных гидростатов ), видоизмененных из примитивной стопы моллюска. Рыбаки иногда называют головоногих моллюсков « чернильными рыбами », имея в виду их общую способность брызгать чернилами . Изучение головоногих — раздел малакологии, известный как тевтология .
период доминирующими стали головоногие В ордовикский , представленные примитивными наутилоидами . Класс теперь содержит два, лишь отдаленно родственных, существующих подкласса: Coleoidea , который включает осьминогов , кальмаров и каракатиц ; и Nautiloidea , представленная Nautilus и Allonautilus . У Coleoidea раковина моллюска интернализирована или отсутствует, тогда как у Nautiloidea внешняя оболочка сохраняется. около 800 современных видов Выявлено головоногих моллюсков. Двумя важными вымершими таксонами являются Ammonoidea (аммониты) и Belemnoidea (белемниты). Размер современных головоногих варьируется от Idiosepius thailandicus размером 10 мм (0,3 дюйма) до колоссального кальмара длиной 14 м (45,1 фута) , самого крупного из ныне живущих беспозвоночных .
Распространение [ править ]


Справа: осьминог ( Benthoctopus sp.) на подводной горе Дэвидсон на глубине 2422 м.
насчитывается более 800 На сегодняшний день видов головоногих моллюсков. [4] хотя новые виды продолжают описываться. Было описано около 11 000 вымерших таксонов , хотя мягкотелая природа головоногих означает, что их нелегко окаменеть. [5]
Головоногие моллюски встречаются во всех океанах Земли. Ни один из них не переносит пресную воду , но кальмар Lolliguncula brevis , обитающий в Чесапикском заливе , является заметным частичным исключением, поскольку он переносит солоноватую воду . [6] Считается, что головоногие моллюски не могут жить в пресной воде из-за многочисленных биохимических ограничений, и за время своего существования более 400 миллионов лет ни разу не рискнули перейти в полностью пресноводную среду обитания. [7]
Головоногие моллюски занимают большую часть глубины океана, от абиссальных равнин до морской поверхности, а также обнаружены в хадаловой зоне . [8] Их разнообразие наиболее велико вблизи экватора (около 40 видов, пойманных сетями на 11° с.ш. в результате исследования разнообразия) и уменьшается к полюсам (~5 видов, пойманных на 60° с.ш.). [9]
Биология [ править ]
Нервная система и поведение [ править ]


Справа: гавайский бобтейл-кальмар , сколопы Euprymna , зарываются в песок, оставляя открытыми только глаза.
Головоногие моллюски считаются самыми умными из беспозвоночных , у них хорошо развиты чувства и большой мозг (больше, чем у брюхоногих моллюсков ). [10] Нервная система головоногих моллюсков – самая сложная из беспозвоночных. [11] [12] а соотношение массы мозга к массе тела у них находится между таковым у эндотермных и экзотермных позвоночных. [9] : 14 Также известно, что содержащиеся в неволе головоногие моллюски вылезают из своих аквариумов, проходят расстояние от пола лаборатории, заходят в другой аквариум, чтобы питаться крабами в неволе, и возвращаются в свой собственный аквариум. [13]
Мозг защищен хрящевым черепом. Гигантские нервные головоногих моллюсков волокна мантии в течение многих лет широко используются как экспериментальный материал в нейрофизиологии ; их большой диаметр (из-за отсутствия миелинизации ) делает их относительно легкими для изучения по сравнению с другими животными. [14]
Многие головоногие — социальные существа; Некоторые виды, изолированные от себе подобных, скапливались с рыбой. [15]
Некоторые головоногие моллюски способны летать по воздуху на расстояние до 50 метров (160 футов). Хотя головоногие моллюски не обладают особой аэродинамичностью, они достигают таких впечатляющих дальностей за счет реактивного движения; вода продолжает выбрасываться из воронки, пока организм находится в воздухе. [16] Животные расправляют плавники и щупальца, образуя крылья, и активно контролируют подъемную силу с помощью положения тела. [17] У одного вида, Todarodes pacificus , наблюдалось распространение щупалец в форме плоского веера с пленкой слизи между отдельными щупальцами. [17] [18] в то время как у другого вида, Sepioteuthis sepioidea , щупальца располагались по кругу. [19]
Чувства [ править ]
Головоногие моллюски обладают развитым зрением, могут определять гравитацию с помощью статоцист и имеют множество химических органов чувств. [9] : 34 Осьминоги используют свои руки, чтобы исследовать окружающую среду, и могут использовать их для восприятия глубины. [9]
Видение [ править ]

Большинство головоногих полагаются на зрение, чтобы обнаруживать хищников и добычу, а также общаться друг с другом. [20] Следовательно, зрение головоногих моллюсков острое: обучающие эксперименты показали, что обыкновенный осьминог способен различать яркость, размер, форму, горизонтальную или вертикальную ориентацию предметов. Морфологическое строение придает глазам головоногих моллюсков те же характеристики, что и глазам акулы; однако их конструкция отличается: у головоногих нет роговицы и вывернутая сетчатка. [20] Глаза головоногих также чувствительны к плоскости поляризации света. [21] В отличие от многих других головоногих моллюсков, наутилусы не обладают хорошим зрением; строение глаз у них сильно развито, но отсутствует твердый хрусталик . У них есть простой глазок- обскура , через который может проходить вода. Считается, что вместо зрения животное использует обоняние в качестве основного чувства для поиска пищи , а также для поиска или идентификации потенциальных партнеров.

Все осьминоги [22] и большинство головоногих моллюсков [23] [24] считаются дальтониками . Колеоидные головоногие моллюски (осьминоги, кальмары, каракатицы) имеют один тип фоторецепторов и лишены способности определять цвет путем сравнения интенсивности обнаруженных фотонов по нескольким спектральным каналам. Маскируя которые себя, они используют свои хроматофоры для изменения яркости и рисунка в зависимости от видимого фона, но их способность соответствовать определенному цвету фона может исходить от таких клеток, как иридофоры и лейкофоры, отражают свет из окружающей среды. [25] Они также производят зрительные пигменты по всему телу и могут чувствовать уровень освещенности непосредственно своим телом. [26] Свидетельства цветового зрения были обнаружены у сверкающего кальмара-энопы ( Watasenia scintillans ). [23] [27] Цветное зрение достигается с помощью трех фоторецепторов , которые основаны на одном и том же опсине , но используют разные молекулы сетчатки в качестве хромофоров: А1 (ретиналь), А3 (3-дегидроретиналь) и А4 (4-гидроксиретиналь). Фоторецептор А1 наиболее чувствителен к зелено-синему (484 нм), фоторецептор А2 — к сине-зеленому (500 нм), а фоторецептор А4 — к синему (470 нм) свету. [28]
В 2015 году был описан новый механизм спектральной дискриминации у головоногих моллюсков. Это основано на использовании хроматической аберрации (зависимости фокусного расстояния от длины волны). Численное моделирование показывает, что хроматическая аберрация может дать полезную хроматическую информацию за счет зависимости остроты изображения от аккомодации. Необычная внеосевая щель и кольцевидная форма зрачков головоногих моллюсков усиливают эту способность, действуя как призмы, рассеивающие белый свет во всех направлениях. [29] [30]
Фотоприем [ править ]
В 2015 году были опубликованы молекулярные данные, указывающие на то, что хроматофоры головоногих моллюсков светочувствительны; Полимеразная цепная реакция с обратной транскрипцией (ОТ-ПЦР) выявила транскрипты, кодирующие родопсин и ретинохром, в сетчатке и коже длинноплавникового прибрежного кальмара ( Doryteuthis pealeii ), а также обыкновенной каракатицы ( Sepia officinalis ) и каракатицы широкопалой ( Sepia latimanus ). Авторы утверждают, что это первое свидетельство того, что кожные ткани головоногих моллюсков могут обладать необходимой комбинацией молекул, чтобы реагировать на свет. [31]
Слушание [ править ]
Было показано, что некоторые кальмары улавливают звук с помощью своих статоцист . [32] но вообще головоногие глухие.
Использование света [ править ]

Большинство головоногих обладают набором компонентов кожи, которые взаимодействуют со светом. Они могут включать иридофоры, лейкофоры , хроматофоры и (у некоторых видов) фотофоры . Хроматофоры — это цветные пигментные клетки, которые расширяются и сжимаются в зависимости от цвета и рисунка, которые они могут использовать самым поразительным образом. [9] [31] Некоторые головоногие не только маскируют свой фон, но и биолюминесцируют, направляя свет вниз, чтобы замаскировать свои тени от хищников, которые могут скрываться внизу. [9] Биолюминесценция ; производится бактериальными симбионтами головоногие моллюски-хозяева способны улавливать свет, излучаемый этими организмами. [33] Биолюминесценцию также можно использовать для привлечения добычи, а некоторые виды используют красочные изображения, чтобы произвести впечатление на партнеров, напугать хищников или даже общаться друг с другом. [9]
Окраска [ править ]
Головоногие могут менять свои цвета и узоры за миллисекунды, будь то для подачи сигнала (как внутри вида , так и для предупреждения ) или для активного камуфляжа . [9] по мере того, как их хроматофоры расширяются или сжимаются. [34] Хотя изменение цвета, по-видимому, зависит в первую очередь от зрительного воздействия, есть свидетельства того, что клетки кожи, в частности хроматофоры , могут обнаруживать свет и приспосабливаться к условиям освещения независимо от глаз. [35] Осьминог меняет цвет и текстуру кожи во время циклов спокойного и активного сна. [36]
Головоногие моллюски могут использовать хроматофоры как мышцы, поэтому они могут так же быстро менять оттенок кожи.Окраска обычно сильнее у прибрежных видов, чем у видов, обитающих в открытом океане, чьи функции, как правило, ограничиваются разрушительным камуфляжем . [9] : 2 Эти хроматофоры расположены по всему телу осьминога, однако ими управляет та же часть мозга, которая контролирует удлинение во время реактивного движения для уменьшения сопротивления. Таким образом, летающие осьминоги могут побледнеть, потому что мозг не может одновременно контролировать удлинение и хроматофоры. [37] Большинство осьминогов имитируют избранные структуры в поле зрения, а не становятся составным цветом своего всего фона. [38]
Доказательства первоначальной окраски были обнаружены в окаменелостях головоногих моллюсков, датируемых еще силурийским периодом ; эти ортоконические особи имели концентрические полосы, которые, как полагают, служили камуфляжем. [39] Девонские головоногие моллюски имеют более сложные цветовые узоры, функция которых неизвестна. [40]
Хроматофоры [ править ]
Колеоиды, подкласс головоногих моллюсков (кальмаров, каракатиц и осьминогов), не имеющих панциря, имеют сложные пигментсодержащие клетки, называемые хроматофорами, которые способны создавать быстро меняющиеся цветовые узоры. Эти клетки хранят пигмент в эластичном мешочке, который и придает цвет, видимый этими клетками. Колеоиды могут изменять форму этого мешочка, называемого цитоэластичным саккулюсом, что затем вызывает изменения прозрачности и непрозрачности клетки. Быстро меняя множество хроматофоров разного цвета, головоногие способны менять цвет своей кожи с поразительной скоростью. Эта адаптация особенно заметна у организма, который видит черно-белое изображение. Известно, что хроматофоры содержат только три пигмента: красный, желтый и коричневый, которые не могут создать полный цветовой спектр. [41] Однако у головоногих также есть клетки, называемые иридофоры, тонкие, слоистые белковые клетки, которые отражают свет способами, которые могут производить цвета, которые хроматофоры не могут. [42] Механизм контроля иридофоров неизвестен, но хроматофоры находятся под контролем нервных путей, что позволяет головоногим моллюскам координировать сложные проявления. Вместе хроматофоры и иридофоры способны создавать широкий спектр цветов и узоров.
Адаптивное значение [ править ]
Головоногие моллюски используют способность хроматофоров менять цвет, чтобы маскироваться. Хроматофоры позволяют колеоидам сливаться с самыми разными средами: от коралловых рифов до песчаного морского дна. Изменение цвета хроматофоров происходит совместно с сосочками, эпителиальной тканью, которая растет и деформируется за счет гидростатического движения, изменяя текстуру кожи. Хроматофоры способны выполнять два типа камуфляжа: мимикрию и подбор цвета. Мимикрия — это когда организм меняет свой внешний вид, чтобы выглядеть как другой организм. Было задокументировано, что кальмар Sepioteuthis sepioide изменил свой внешний вид и стал выглядеть как не представляющая угрозы травоядная рыба-попугай, приближающаяся к ничего не подозревающей добыче. Известно, что осьминог Thaumoctopus mimicus имитирует ряд различных ядовитых организмов, с которыми он сосуществует, чтобы отпугивать хищников. [43] При совпадении фона головоногие меняют свой внешний вид, чтобы напоминать окружающую среду, прячась от хищников или прячась от добычи. Способность имитировать другие организмы и соответствовать внешнему виду окружающей среды примечательна, учитывая, что зрение головоногих моллюсков монохроматично.
Головоногие моллюски также используют свой тонкий контроль над окраской и рисунком тела для отображения сложных сигналов как для видовой, так и для внутривидовой коммуникации. Окраска используется вместе с передвижением и текстурой для передачи сигналов другим организмам. Внутривидово это может служить предупреждением для потенциальных хищников. Например, когда осьминогу Callistoctopus macropus угрожает опасность, он приобретает ярко-красно-коричневый цвет с белыми точками в качестве высококонтрастного изображения, чтобы отпугнуть хищников. В частности, изменение цвета используется как для демонстрации спаривания, так и для социального общения. Каракатицы демонстрируют сложную демонстрацию спаривания самцов и самок. Существует также передача сигналов от мужчины к мужчине, которая происходит во время конкуренции за партнеров, и все они являются продуктом проявления окраски хроматофора.
Происхождение [ править ]
Существуют две гипотезы об эволюции изменения окраски у головоногих моллюсков. Одна из гипотез состоит в том, что способность менять цвет могла развиться для выполнения социальных, сексуальных и сигнальных функций. Другое объяснение состоит в том, что оно впервые появилось из-за давления отбора, поощряющего избегание хищников и скрытную охоту.
Чтобы изменение цвета возникло в результате социального отбора, среда обитания предков головоногих моллюсков должна была соответствовать ряду критериев. Во-первых, должен быть какой-то брачный ритуал, включающий передачу сигналов. Во-вторых, им придется испытать явно высокий уровень полового отбора. И в-третьих, предку необходимо было бы общаться, используя сексуальные сигналы, видимые получателю своего вида. Чтобы изменение цвета возникло в результате естественного отбора, должны быть соблюдены различные параметры. Во-первых, вам понадобится некоторое фенотипическое разнообразие в телосложении среди населения. Этому виду также придется сосуществовать с хищниками, которые полагаются на зрение для идентификации добычи. Эти хищники должны обладать высоким диапазоном зрительной чувствительности, распознавая не только движение или контраст, но и цвета. Места обитания, которые они занимают, также должны отражать разнообразие происхождения. [44] Эксперименты, проведенные на карликовых хамелеонах для проверки этой гипотезы, показали, что таксоны хамелеонов с большей способностью к изменению цвета имели более визуально заметные социальные сигналы, но не происходили из более визуально разнообразных мест обитания, что позволяет предположить, что способность к изменению цвета, вероятно, развилась для облегчения передачи социальных сигналов, в то время как камуфляж является полезный побочный продукт. [45] Поскольку камуфляж используется головоногими моллюсками для множества адаптивных целей, изменение цвета могло возникнуть для одного использования, а другое — позже, или же оно эволюционировало для регулирования компромиссов внутри обоих.
Конвергентная эволюция [ править ]
Изменение цвета широко распространено у эктотермных животных, включая анолисов, лягушек, моллюсков, многих рыб, насекомых и пауков. [46] Механизм изменения цвета может быть морфологическим или физиологическим. Морфологические изменения являются результатом изменения плотности пигментсодержащих клеток и имеют тенденцию меняться в течение более длительных периодов времени. Физиологические изменения, наблюдаемые у головоногих моллюсков, обычно являются результатом движения пигмента внутри хроматофора, изменяющего места локализации различных пигментов внутри клетки. Это физиологическое изменение обычно происходит в гораздо более короткие сроки по сравнению с морфологическими изменениями. Головоногие моллюски обладают редкой формой физиологического изменения цвета, при которой используется нервный контроль мышц для изменения морфологии их хроматофоров. Этот нейронный контроль хроматофоров развился конвергентно как у головоногих, так и у костистых рыб. [47]
Чернила [ править ]
За исключением Nautilidae и видов осьминогов, принадлежащих к подотряду Cirrina , [48] У всех известных головоногих есть чернильный мешок, с помощью которого можно выбрасывать облако темных чернил, чтобы сбить с толку хищников . [22] Этот мешок представляет собой мышечный мешок, возникший как продолжение задней кишки. Он расположен под кишечником и открывается в задний проход, куда его содержимое – почти чистый меланин можно впрыскивать ; его близость к основанию воронки означает, что чернила могут распределяться за счет выбрасываемой воды, поскольку головоногие моллюски используют реактивное движение. [22] Выброшенное облако меланина при изгнании обычно смешивается со слизью , вырабатываемой в других частях мантии, и поэтому образует густое облако, приводящее к зрительным (и, возможно, хемосенсорным) нарушениям у хищника, наподобие дымовой завесы . Однако наблюдалось более сложное поведение, при котором головоногие выпускают облако с большим содержанием слизи, примерно напоминающее выпустившее его головоногие моллюски (эту приманку называют псевдоморфом ) . Эта стратегия часто приводит к тому, что хищник атакует псевдоморф, а не быстро убегающую добычу. [22] Дополнительные сведения см. в разделе Поведение рукописного ввода .
Чернильный мешок головоногих моллюсков привел к общему названию «чернильница». [49] ранее рыба, написанная пером и чернилами. [50]
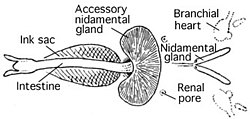

Кровеносная система [ править ]
Головоногие моллюски — единственные моллюски с закрытой кровеносной системой. Колеоиды имеют два жаберных сердца (также известных как жаберные сердца ), которые перемещают кровь через капилляры жабр . Затем одно системное сердце перекачивает насыщенную кислородом кровь через остальную часть тела. [51]
Как и большинство моллюсков, головоногие моллюски используют для транспортировки кислорода гемоцианин , медьсодержащий белок, а не гемоглобин . В результате их кровь бесцветна при деоксигенации и становится синей при связывании с кислородом. [52] В средах, богатых кислородом, и в кислой воде гемоглобин более эффективен, но в средах с низким содержанием кислорода и при низких температурах преимущество имеет гемоцианин. [53] [54] [55] Молекула гемоцианина намного больше молекулы гемоглобина, что позволяет ей связываться с 96 Молекулы O 2 или CO 2 вместо всего четырех у гемоглобина. Но в отличие от гемоглобина, который миллионами прикреплен к поверхности одного эритроцита, молекулы гемоцианина свободно плавают в кровотоке. [56]
Дыхание [ править ]
Головоногие обмениваются газами с морской водой, проталкивая воду через жабры, прикрепленные к крыше организма. [57] : 488 [58] Вода поступает в мантийную полость снаружи жабр, и вход в мантийную полость закрывается. Когда мантия сжимается, вода выталкивается через жабры, расположенные между мантийной полостью и воронкой. Выброс воды через воронку можно использовать для реактивного движения. Если дыхание используется одновременно с реактивным движением, можно ожидать больших потерь в скорости или выработке кислорода. [59] [60] К вентральной поверхности мантийной полости прикрепляются жабры, значительно более эффективные, чем у других моллюсков. [58] В зависимости от образа жизни существует компромисс с размером жабр. Чтобы достичь высоких скоростей, жабры должны быть маленькими — вода будет проходить через них быстро, когда потребуется энергия, что компенсирует их небольшой размер. Однако организмы, которые проводят большую часть своего времени медленно, перемещаясь по дну, естественным образом не пропускают через свою полость много воды для передвижения; таким образом, у них более крупные жабры, а также сложные системы, обеспечивающие постоянное промывание воды через жабры, даже когда организм неподвижен. [57] Поток воды контролируется сокращениями радиальных и круговых мышц мантийной полости. [61]
Жабры головоногих поддерживаются скелетом из прочных волокнистых белков; отсутствие мукополисахаридов отличает этот матрикс от хряща. [62] [63] Считается, что жабры также участвуют в выделении NH 4 + меняется на К + из морской воды. [58]
Передвижение и плавучесть [ править ]

Хотя большинство головоногих моллюсков могут передвигаться с помощью реактивного движения, это очень энергозатратный способ передвижения по сравнению с движением хвоста, используемым рыбами. [64] КПД винтовым приводом с водомета (т.е. эффективность Фруда ) выше, чем у ракеты . [65] Относительная эффективность реактивного движения снижается по мере увеличения размера животного; параличинки гораздо более эффективны, чем молодые и взрослые особи. [66] Начиная с палеозойской эры , когда конкуренция с рыбами создала среду, в которой эффективное движение имело решающее значение для выживания, реактивное движение отошло на второй план, а плавники и щупальца использовались для поддержания постоянной скорости. [5] Реактивное движение никогда не является единственным способом передвижения. [5] : 208 движение «стоп-старт», обеспечиваемое форсунками, по-прежнему полезно для обеспечения высокоскоростных рывков – не в последнюю очередь при поимке добычи или уклонении от хищников . [5] Действительно, это делает головоногих моллюсков самыми быстрыми морскими беспозвоночными. [9] : Предисловие и они могут обогнать большинство рыб. [57] Струя дополняется движением киля; у кальмаров плавники хлопают каждый раз, когда выпускается струя, усиливая тягу; затем они выдвигаются между струями (предположительно, чтобы избежать затопления). [66] Окисленная вода забирается в мантийную полость к жабрам и посредством мышечного сокращения этой полости выбрасывается израсходованная вода через гипоном , образованный складкой мантии. Разница в размерах между задним и передним концами этого органа контролирует скорость струи, которую может производить организм. [67] Скорость организма можно точно предсказать для заданной массы и морфологии животного. [68] Движение головоногих моллюсков обычно происходит назад, поскольку вода вытесняется вперед через гипоном, но направление можно в некоторой степени контролировать, направляя ее в разные стороны. [69] Некоторые головоногие моллюски сопровождают выброс воды хлопающим звуком, похожим на выстрел, который, как полагают, служит отпугиванию потенциальных хищников. [70]
Головоногие моллюски используют аналогичный метод движения, несмотря на то, что их увеличивающиеся размеры (по мере роста) меняют динамику воды, в которой они находятся. Таким образом, их параличинки не используют широко свои плавники (которые менее эффективны при низких числах Рейнольдса ) и в основном используют свои струи для продвижения вверх, тогда как крупные взрослые головоногие моллюски имеют тенденцию плавать менее эффективно и больше полагаться на свои плавники. [66]

Считается, что ранние головоногие моллюски создавали струи, втягивая свое тело в раковины, как это делает сегодня Наутилус . [71] Наутилус также способен создавать струю за счет волнистости своей воронки; этот более медленный поток воды больше подходит для извлечения кислорода из воды. [71] В неподвижном состоянии Наутилус может извлечь из воды только 20% кислорода. [59] Скорость струи у Наутилуса гораздо медленнее, чем у колеоидов , но в ее производстве задействовано меньше мускулатуры и энергии. [72] Реактивная тяга у головоногих определяется в первую очередь максимальным диаметром отверстия воронки (или, возможно, средним диаметром воронки). [73] : 440 и диаметр мантийной полости. [74] Изменения размера отверстия чаще всего используются на промежуточных скоростях. [73] Достигаемая абсолютная скорость ограничена потребностью головоногих вдыхать воду для выбрасывания; этот прием ограничивает максимальную скорость восемью длинами тела в секунду - скоростью, которую большинство головоногих моллюсков могут достичь после двух ударов воронки. [73] Вода наполняет полость, попадая не только через отверстия, но и через воронку. [73] Кальмары могут выбрасывать до 94% жидкости из своей полости за один реактивный толчок. [65] Чтобы приспособиться к быстрым изменениям в заборе и выведении воды, отверстия очень гибкие и могут изменять свой размер в двадцать раз; радиус воронки, наоборот, изменяется лишь примерно в 1,5 раза. [73]
Некоторые виды осьминогов также способны ходить по морскому дну. Кальмары и каракатицы могут перемещаться на короткие расстояния в любом направлении, перемещая мышечный лоскут вокруг мантии.
Хотя большинство головоногих плавает (то есть имеют нейтральную или почти плавучесть; на самом деле плотность большинства головоногих примерно на 2–3 % выше, чем у морской воды). [15] ), они добиваются этого по-разному. [64] Некоторые, такие как Наутилус , позволяют газу диффундировать в зазор между мантией и оболочкой; другие позволяют более чистой воде сочиться из почек, вытесняя более плотную соленую воду из полости тела; [64] другие, как некоторые рыбы, накапливают жиры в печени; [64] а у некоторых осьминогов студенистое тело с более легкими хлорида ионами , заменяющими сульфат в химическом составе тела. [64]
Кальмары являются основными жертвами отрицательной плавучести головоногих моллюсков. Отрицательная плавучесть означает, что некоторым кальмарам, особенно тем, глубина обитания которых довольно невелика, приходится активно регулировать свое вертикальное положение. Это означает, что им приходится расходовать энергию, часто за счет струи или волнистости, чтобы поддерживать одинаковую глубину. Таким образом, стоимость транспортировки многих кальмаров довольно высока. При этом кальмары и другие головоногие моллюски, обитающие в глубоких водах, имеют тенденцию иметь более нейтральную плавучесть, что устраняет необходимость регулирования глубины и повышает их двигательную эффективность. [75] [59]
Было замечено, что Macrotritopus defilippi , или обитающий в песке осьминог, имитирует окраску и плавательные движения обитающей в песке камбалы Bothus lunatus , чтобы избежать хищников. Осьминоги смогли сплющить свои тела и отвести руки назад, чтобы выглядеть так же, как камбалы, а также двигаться с той же скоростью и движениями. [76]
У самок двух видов, Ocythoe tuberculata и Haliphron atlanticus , развился настоящий плавательный пузырь . [77]
осьминога кальмара против Передвижение
Две категории головоногих моллюсков, осьминоги и кальмары, сильно различаются в своих движениях, несмотря на то, что принадлежат к одному и тому же классу. Осьминоги обычно не считаются активными пловцами; их часто можно встретить очищающими морское дно вместо того, чтобы плавать по воде на большие расстояния. С другой стороны, кальмары могут преодолевать огромные расстояния: некоторые из них преодолевают расстояние до 2000 км за 2,5 месяца со средней скоростью 0,9 длины тела в секунду. [78] Существует основная причина разницы в типе и эффективности движений: анатомия.
И у осьминогов, и у кальмаров есть мантия (упомянутая выше), которая обеспечивает дыхание и передвижение в виде струи. Однако состав этих мантий у этих двух семейств различается. У осьминогов мантия состоит из трех типов мышц: продольных, радиальных и кольцевых. Продольные мышцы проходят параллельно длине осьминога и используются для поддержания одинаковой длины мантии на протяжении всего процесса выброса струи. Учитывая, что это мышцы, можно отметить, что это означает, что осьминог должен активно сгибать продольные мышцы во время выброса струи, чтобы поддерживать мантию постоянной длины. Радиальные мышцы проходят перпендикулярно продольным мышцам и используются для утолщения и утончения стенки мантии. Наконец, круговые мышцы используются в качестве основных активаторов при струях. Это мышечные группы, окружающие мантию и расширяющие/сжимающие полость. Все три типа мышц работают в унисон, создавая реактивную струю как двигательный механизм. [78]
У кальмаров нет продольных мышц, которые есть у осьминогов. Вместо этого у них туника. [78] Эта туника состоит из слоев коллагена и окружает верхнюю и нижнюю часть мантии. Поскольку туники состоят из коллагена, а не из мышц, они представляют собой твердые тела, которые намного прочнее, чем их мышечные аналоги. Это дает кальмарам некоторые преимущества при водометном плавании. Жесткость означает, что нет необходимости сгибать мышцы, чтобы поддерживать мантию того же размера. Кроме того, туники занимают лишь 1% толщины стенки мантии кальмара, тогда как у осьминогов продольные мышечные волокна занимают до 20% толщины стенки мантии. [78] Кроме того, из-за жесткости туники радиальные мышцы кальмара могут сокращаться сильнее.
Мантия — не единственное место, где у кальмаров есть коллаген. Коллагеновые волокна расположены среди других мышечных волокон мантии. Эти коллагеновые волокна действуют как эластики и иногда называются «коллагеновыми пружинами». [78] Как следует из названия, эти волокна действуют как пружины. Когда радиальные и круговые мышцы мантии сокращаются, они достигают точки, в которой сокращение больше не способствует движению существа вперед. В таких случаях избыточное сокращение сохраняется в коллагене, который затем эффективно начинается или способствует расширению мантии в конце струи. В некоторых тестах было показано, что коллаген способен начать повышать мантийное давление за 50 мс до начала мышечной активности. [78]
Эти анатомические различия между кальмарами и осьминогами могут помочь объяснить, почему кальмары плавают так же, как и рыбы, в то время как осьминоги обычно полагаются на другие формы передвижения по морскому дну, такие как ходьба на двух ногах, ползание и плавание без струи. [79]
Шелл [ править ]


Наутилусы — единственные современные головоногие моллюски с настоящей внешней оболочкой. Однако все раковины моллюсков образуются из эктодермы (наружного слоя зародыша); у каракатиц ( Sepia spp.), например, в эмбриональном периоде формируется инвагинация эктодермы, в результате чего образуется панцирь ( каракатица ), находящийся у взрослой особи внутренним. [80] То же самое и с хитиновым гладиусом кальмара. [80] и осьминоги. [81] Цирратные осьминоги имеют дугообразные хрящевые опоры плавников . [82] которые иногда называют «остатком ракушки» или «гладиусом». [83] либо У Инциррины есть пара палочковидных стилетов , либо нет остатков внутренней раковины. [84] а у некоторых кальмаров отсутствует гладиус. [85] Колеоиды с панцирем не образуют ни клады, ни даже парафилетической группы. [86] Оболочка спирулы изначально представляет собой органическую структуру, а затем очень быстро минерализуется. [87] «Потерянные» оболочки могут быть потеряны в результате резорбции компонента карбоната кальция. [88]
Самки осьминогов рода Argonauta выделяют специализированную яйцевую оболочку толщиной с бумагу, в которой они живут, и ее в народе называют «скорлупой», хотя она не прикреплена к телу животного и имеет отдельное эволюционное происхождение.
Самая большая группа панцирных головоногих моллюсков — аммониты — вымерла, но их панцири очень распространены в виде ископаемых .
Отложение карбоната, приводящее к образованию минерализованной раковины, по-видимому, связано с кислотностью органического матрикса раковины (см. Раковина моллюска ); раковинообразующие головоногие моллюски имеют кислую матрицу, тогда как гладиусы кальмара имеют основную матрицу. [89] Основное устройство наружной стенки головоногих моллюсков следующее: наружный (сферулитовый) призматический слой, ламинарный (перламутровый) слой и внутренний призматический слой. Толщина каждого слоя зависит от таксона. [90] У современных головоногих карбонат кальция представляет собой арагонит. Что касается других раковин моллюсков или скелетов кораллов, то самые мелкие видимые единицы представляют собой гранулы неправильной округлой формы. [91]


Справа: деталь щупальцевой булавы Abraliopsis morisi.
Придатки головы [ править ]
Головоногие, как следует из названия, имеют мышечные придатки, отходящие от головы и окружающие рот. Они используются для питания, передвижения и даже размножения. В колеоидах их восемь или десять. У десятиногих моллюсков, таких как каракатицы и кальмары, пять пар. Два более длинных, называемые щупальцами , активно участвуют в захвате добычи; [1] : 225 они могут быстро удлиняться (всего за 15 миллисекунд). [1] : 225 ). У гигантских кальмаров они могут достигать в длину 8 метров. Они могут заканчиваться расширенной булавой, покрытой присосками. [1] : 225 Четыре более короткие пары называются руками и участвуют в удержании и манипулировании захваченным организмом. [1] : 225 У них тоже есть присоски на стороне, ближайшей ко рту; они помогают удерживать добычу. [1] : 226 Как следует из названия, у осьминогов есть только четыре пары рук, покрытых присосками, хотя аномалии развития могут изменять количество выраженных рук. [92]
Щупальца состоят из толстого центрального нервного тяжа (который должен быть толстым, чтобы можно было управлять каждой присоской независимо). [93] окружен круговыми и радиальными мышцами. Поскольку объем щупальца остается постоянным, сокращение круговых мышц уменьшает радиус и позволяет быстро увеличивать длину. Обычно удлинение на 70% достигается за счет уменьшения ширины на 23%. [1] : 227 У более коротких рук такая возможность отсутствует.
Размер щупальца связан с размером ротовой полости; более крупные и сильные щупальца могут удерживать добычу, делая от нее небольшие укусы; с более многочисленными и меньшими щупальцами добыча заглатывается целиком, поэтому ротовая полость должна быть больше. [94]
с внешним панцирем Наутилиды ( Nautilus и Allonautilus ) имеют порядка 90 пальцеобразных придатков, называемых щупальцами , у которых нет присосок, но они липкие и частично втягиваются.
Кормление [ править ]

состоит из двух частей У всех современных головоногих клюв ; [9] : 7 у большинства из них есть радула , хотя у большинства осьминогов она редуцирована, а у Spirula она вообще отсутствует . [9] : 7 [95] : 110 Они питаются, захватывая добычу щупальцами, втягивая ее в рот и откусывая от нее укусы. [22] У них есть смесь токсичных пищеварительных соков, некоторые из которых производятся симбиотическими водорослями, которые они выбрасывают из слюнных желез на пойманную добычу, удерживаемую во рту. Эти соки отделяют плоть добычи от костей или панциря. [22] На конце слюнной железы есть небольшой зуб, который можно вонзить в организм, чтобы переварить его изнутри. [22]
Сама пищеварительная железа довольно короткая. [22] Он состоит из четырех элементов: пища проходит через зоб, желудок и слепую кишку, прежде чем попасть в кишечник. Большая часть пищеварения, а также всасывания питательных веществ происходит в пищеварительной железе, иногда называемой печенью. Обмен питательными веществами и отходами между кишечником и пищеварительной железой осуществляется через пару соединений, соединяющих железу с местом соединения желудка и слепой кишки. [22] Клетки пищеварительной железы непосредственно выделяют пигментированные экскреторные химические вещества в просвет кишечника, которые затем связываются со слизью, проходящей через задний проход в виде длинных темных нитей, выбрасываемых с помощью выдыхаемой воды из воронки. [22] Головоногие моллюски склонны концентрировать поступившие с пищей тяжелые металлы в тканях своего тела. [96] Однако руки осьминогов используют семейство хемотактильных рецепторов (CR), специфичных для головоногих моллюсков, в качестве системы «вкуса на ощупь». [97]
Радула [ править ]

Радула головоногих моллюсков состоит из нескольких симметричных рядов, насчитывающих до девяти зубов. [98] - тринадцать в классах ископаемых. [99] Этот орган редуцирован или даже рудиментарен у некоторых видов осьминогов и отсутствует у Spirula . [99] Зубы могут быть гомодонтными (т.е. одинаковыми по форме по всему ряду), гетеродонтными (в противном случае) или ктенодонтными (гребневидными). [99] Их высота, ширина и количество выступов варьируются в зависимости от вида. [99] Рисунок зубов повторяется, но каждый ряд может быть не идентичен предыдущему; например, у осьминога последовательность повторяется каждые пять рядов. [99] : 79
Головоногие радулы известны из ископаемых отложений ордовика. [100] Обычно они сохраняются в камере тела головоногих моллюсков, обычно вместе с нижними челюстями; но это не всегда так; [101] многие радулы сохранились в различных местах Мейсон-Крик. [102] Радулы обычно трудно обнаружить, даже если они сохранились в окаменелостях, поскольку порода должна выветриться и расколоться именно таким образом, чтобы они обнажились; например, радулы были обнаружены только у девяти из 43 родов аммонитов. [103] [ нужны разъяснения ] еще реже они встречаются в неаммоноидных формах: только три домезозойских вида обладают одной. [100]
Выделительная система [ править ]
У большинства головоногих имеется одна пара крупных нефридиев . Отфильтрованные азотистые отходы образуются в перикарда полости жаберных сердец , каждая из которых соединена с нефридием узким каналом. Канал доставляет экскременты в пузыреподобный почечный мешок, а также резорбирует излишнюю воду из фильтрата. Несколько отростков латеральной полой вены выступают в почечный мешок, непрерывно надуваясь и сдуваясь при сокращении жаберных сердец. Это действие помогает перекачивать секретируемые отходы в мешочки и выбрасывать их в мантийную полость через поры. [104]
Наутилус , что необычно, обладает четырьмя нефридиями, ни один из которых не связан с полостями перикарда.
Включение аммиака важно для формирования раковин у наземных моллюсков и других немоллюсковых линий. [105] Поскольку белок (т. е. мясо) является основным компонентом рациона головоногих моллюсков, большое количество ионов аммония в виде отходов образуется . Основным органом, участвующим в высвобождении избытка аммония, являются жабры. [106] Скорость выделения самая низкая у головоногих моллюсков Nautilus и Sepia с панцирем , поскольку они используют азот для наполнения своих панцирей газом для увеличения плавучести. [106] Другие головоногие моллюски используют аммоний аналогичным образом, сохраняя ионы (в виде хлорида аммония ), чтобы уменьшить их общую плотность и увеличить плавучесть. [106]
и Размножение цикл жизненный




Головоногие моллюски представляют собой разнообразную группу видов, но имеют общие черты жизненного цикла, например, у них быстрый темп роста и короткая продолжительность жизни. [107] Стернс (1992) предположил, что для производства как можно большего числа жизнеспособного потомства события нереста зависят от экологических факторов окружающей среды организма. Большинство головоногих моллюсков не оказывают родительской заботы о своем потомстве, за исключением, например, осьминога, который помогает этому организму повысить выживаемость своего потомства. [107] На жизненный цикл морских видов влияют различные условия окружающей среды. [108] На развитие эмбриона головоногих моллюсков могут сильно влиять температура, насыщение кислородом, загрязнение окружающей среды, интенсивность света и соленость. [107] Эти факторы важны для скорости эмбрионального развития и успеха вылупления эмбрионов. Доступность пищи также играет важную роль в репродуктивном цикле головоногих моллюсков. Ограничение в пище влияет на сроки нереста, а также на их функцию и рост. [108] Время нереста и нерест различаются у разных морских видов; это коррелирует с температурой, хотя головоногие моллюски на мелководье нерестятся в холодные месяцы, чтобы потомство вылупилось при более высоких температурах. Размножение может длиться от нескольких дней до месяца. [107]
Половая зрелость [ править ]
Головоногие, достигшие половой зрелости и взрослого размера, начинают нереститься и размножаться. После передачи генетического материала следующему поколению взрослые головоногие моллюски у большинства видов умирают. [107] Половое созревание у головоногих самцов и самок можно наблюдать изнутри по увеличению гонад и добавочных желез. [109] Спаривание может быть плохим индикатором полового созревания самок; они могут получать сперму, когда еще не полностью репродуктивно созрели, и хранить ее до тех пор, пока не будут готовы к оплодотворению яйцеклеток. [108] Самцы более агрессивны в соревновании перед спариванием в присутствии неполовозрелых самок, чем в борьбе за половозрелую самку. [110] У большинства самцов головоногих моллюсков развивается гектокотиль - кончик руки, который способен переносить сперматозоиды в мантийную полость самки. Хотя не все виды используют гектокотиль; например, взрослый наутилус выпускает початок . [111] У некоторых самцов кальмаров, в основном глубоководных видов, вместо этого развился пенис, длина которого превышает длину их собственного тела, — самый длинный пенис среди всех свободноживущих животных. Предполагается, что эти самцы просто прикрепляют сперматофор в любом месте тела самки. [112] Признаком половой зрелости самок является развитие плечевых фотофоров для привлечения самок. [113]
Оплодотворение [ править ]
Головоногие моллюски не являются широковещательными производителями . В процессе оплодотворения самки используют сперму, полученную самцом в результате внешнего оплодотворения . Внутреннее оплодотворение наблюдается только у осьминогов. [109] Начало совокупления начинается, когда самец ловит самку и обнимает ее рукой либо в положении «самец к шее самки», либо в положении «рот ко рту», в зависимости от вида. Затем самцы инициируют процесс оплодотворения, несколько раз сжимая мантию для высвобождения сперматозоидов. [114] Головоногие моллюски часто спариваются несколько раз, что заставляет самцов спариваться дольше с самками, которые ранее спаривались почти в три раза чаще, чем количество сокращений мантии. [114] Чтобы обеспечить оплодотворение яйцеклеток, самки головоногих моллюсков выделяют через студенистые слои яйцеклетки пептид, привлекающий сперматозоиды, чтобы направлять сперматозоиды. Самки головоногих откладывают яйца в кладках; каждое яйцо имеет защитную оболочку, обеспечивающую безопасность развивающегося эмбриона при попадании в толщу воды. Репродуктивные стратегии различаются у разных видов головоногих моллюсков. У гигантского тихоокеанского осьминога крупные яйца откладываются в берлоге; на то, чтобы уложить их все, часто уходит несколько дней. [109] После того, как яйца высвобождаются и обычно прикрепляются к защищенному субстрату, самка обычно вскоре умирает, но впоследствии за их яйцами будут присматривать осьминоги и несколько кальмаров. [109] [115] [116] Другие, такие как японский летающий кальмар , выметывают нейтрально плавучие икринные массы, которые плавают на границе слоев воды с немного разной плотностью. [117] или самка будет плавать, неся с собой икру. [118] Большинство видов являются полуродящими (размножаются только один раз перед смертью), единственными известными исключениями являются кальмар-вампир , малый тихоокеанский полосатый осьминог и наутилус , которые являются итерородящими . [119] [120] У некоторых видов головоногих кладки яиц прикрепляются к субстрату с помощью слизистого клейкого вещества. Эти яйца набухают перивителлиновой жидкостью (PVF), гипертонической жидкостью, которая предотвращает преждевременное вылупление. [121] Грозди оплодотворенных яиц имеют нейтральную плавучесть в зависимости от глубины, на которой они были отложены, но их также можно найти в таких субстратах, как песок, матрица кораллов или морские водоросли. [108] Поскольку эти виды не обеспечивают родительскую заботу о своем потомстве, самка может вводить чернила в яйцевые капсулы, чтобы замаскировать эмбрионы от хищников. [108]
Соревнования между мужчинами [ править ]
Большинство головоногих занимаются агрессивным сексом: такое поведение стимулирует белок в оболочке мужской капсулы. Они также участвуют в агрессии между самцами, при этом более крупные самцы, как правило, выигрывают во взаимодействии. [107] Когда самка находится рядом, самцы постоянно атакуют друг друга и размахивают руками. Если ни один самец не отступает, руки вытягиваются назад, обнажая рот, после чего следует кусание кончиков рук. [122] Во время спаривания самцы также используют технику, называемую смыванием. Этот метод используется вторым самцом, пытающимся спариться с самкой. Промывка удаляет из ротовой полости сперматофоры, которые поместил туда первый помощник капитана, путем нагнетания воды в полость. [107] Еще одно поведение, которым занимаются самцы, - это спаривание или мимикрия: более мелкие самцы подстраивают свое поведение под поведение самки, чтобы уменьшить агрессию. Используя эту технику, они могут оплодотворить икру, в то время как более крупный самец отвлекается на другого самца. [122] Во время этого процесса самцы-кроссовки быстро вводят каплевидную сперму в семеприемник. [123]
Выбор партнера [ править ]
Выбор партнера наблюдается у видов каракатиц, у которых самки предпочитают одних самцов другим, хотя характеристики предпочитаемых самцов неизвестны. [107] Гипотеза гласит, что самки отвергают самцов по обонятельным, а не по визуальным сигналам. [107] Некоторые виды головоногих являются полиандрическими: они принимают и хранят несколько мужских сперматофоров, что было идентифицировано с помощью дактилоскопии ДНК. [114] Самки больше не восприимчивы к попыткам спаривания, когда держат яйца на руках. Самки могут хранить сперму в двух местах (1) в ротовой полости, куда недавно спарившиеся самцы помещают свои сперматофоры, и (2) во внутренних резервуарах для хранения спермы, где хранятся пакеты спермы от предыдущих самцов. [107] Хранение сперматофоров приводит к конкуренции сперматозоидов; в котором говорится, что самка контролирует, какой партнер оплодотворяет яйца. Чтобы уменьшить такого рода конкуренцию, самцы развивают агонистическое поведение, такое как охрана партнера и спугивание. [107] Hapalochlaena lunulata , или синекольчатый осьминог, легко спаривается как с самцами, так и с самками. [124]
Половой диморфизм [ править ]
Видно, что у многих морских организмов самки крупнее самцов некоторых близкородственных видов. В некоторых линиях, таких как осьминог-покрывало , самцы становятся все меньше и меньше по структуре, что напоминает термин «карликовость»: карликовые самцы обычно возникают при низкой плотности. [125] Самец покрывалового осьминога является примером сексуально-эволюционной карликовости; самки вырастают в 10 000–40 000 раз крупнее самцов, и соотношение полов между самцами и самками можно определить сразу после вылупления яиц. [125]

Эмбриология [ править ]
Яйца головоногих моллюсков имеют широкий диапазон размеров: от 1 до 30 мм в диаметре. [126] Оплодотворенная яйцеклетка первоначально делится, образуя диск зародышевых клеток на одном полюсе, а желток остается на противоположном полюсе. Зародышевый диск растет, окутывая и в конечном итоге поглощая желток, образуя эмбрион. Щупальца и руки сначала появляются в задней части тела, там, где у других моллюсков была бы нога, и лишь позже мигрируют к голове. [104] [127]
Воронка головоногих развивается на макушке головы, а рот — на противоположной поверхности. [128] : 86 Ранние эмбриологические стадии напоминают предков брюхоногих моллюсков и современных моноплакофор . [127]
Раковины развиваются из эктодермы как органический каркас, который впоследствии минерализуется. [80] У Sepia , имеющей внутреннюю оболочку, эктодерма образует инвагинацию, поры которой закрываются до отложения органического каркаса. [80]
Развитие [ править ]


Справа: зрелый Chiroteuthis veranyi . У этого вида одни из самых длинных щупалец по сравнению с его размером среди всех известных головоногих моллюсков.
Продолжительность времени до вылупления сильно варьируется; Яйца меньшего размера в более теплых водах вылупляются быстрее всего, а новорожденные могут появиться всего через несколько дней. Более крупные икринки в более холодных водах могут развиваться более года, прежде чем вылупятся. [126]
Процесс от нереста до вылупления проходит по одинаковой траектории у всех видов, причем основной переменной является количество желтка, доступного молодняку, и время его поглощения эмбрионом. [126]
В отличие от большинства других моллюсков, головоногие не имеют морфологически четко выраженной личиночной стадии. Вместо этого молодые особи известны как параличинки . Они быстро учатся охотиться, используя встречи с добычей для совершенствования своей стратегии. [126]
Рост молодых особей обычно аллометрический , тогда как рост взрослых особей изометрический . [129]
Эволюция [ править ]
Традиционный взгляд на эволюцию головоногих предполагает, что они произошли в позднем кембрии от моноплакофорам. предка, подобного [130] с изогнутой, сужающейся оболочкой, [131] который был тесно связан с брюхоногими моллюсками (улитками). [132] сходство ранних головоногих моллюсков Plectronoceras В подтверждение этой точки зрения использовалось с панцирем и некоторыми брюхоногими моллюсками. Развитие сифункула позволило бы панцирям этих ранних форм стать наполненными газом (таким образом, плавучими), чтобы поддерживать их и удерживать раковины в вертикальном положении, пока животное ползло по полу, и отделить настоящих головоногих моллюсков от предполагаемых предков, таких как как Knightoconus , у которого отсутствовал сифункул. [132] Нейтральная или положительная плавучесть (то есть способность плавать) появилась позже, за ней последовало плавание у Plectronocerida и, в конечном итоге, реактивное движение у более продвинутых головоногих моллюсков. [133]
Возможные раннекембрийские останки были найдены на полуострове Авалон , что соответствует генетическим данным докембрийского происхождения. [2]
Однако некоторые морфологические данные трудно примирить с этой точкой зрения, а переописание Nectocaris pteryx , который не имел панциря и, по-видимому, обладал реактивным движением наподобие «производных» головоногих моллюсков, усложнило вопрос о порядке, в котором головоногие моллюски развиты особенности – при условии, что Nectocaris вообще является головоногим моллюском. [134]
Ранние головоногие моллюски, вероятно, были хищниками на вершине пищевой цепи. [22] После того как позднекембрийское вымирание привело к исчезновению многих аномалокаридид , хищные ниши стали доступны для других животных. [135] В ордовикский период примитивные головоногие моллюски претерпели импульсы диверсификации. [136] стать разнообразными и доминирующими в палеозойских и мезозойских морях. [137]
В раннем палеозое их ареал был гораздо более ограниченным, чем сегодня; они были приурочены главным образом к сублиторальным районам мелководных шельфов низких широт и обычно встречались в ассоциации с тромболитами . [138] По мере развития ордовика постепенно приобретался более пелагический образ жизни. [138] Глубоководные головоногие хоть и редки, но обнаружены в нижнем ордовике, но только в водах высоких широт. [138] В середине ордовика появились первые головоногие моллюски с перегородками, достаточно сильными, чтобы выдерживать давление, связанное с более глубокими водами, и могли обитать на глубинах более 100–200 м. [136] Направление скручивания раковины окажется решающим для будущего успеха этой линии; Эндогастральная спиральная спираль позволяла достичь больших размеров только с прямой раковиной, тогда как экзогастральная спиральная спираль - первоначально довольно редкая - позволяла развиваться спиралям, знакомым по летописи окаменелостей, с соответствующими им большими размерами и разнообразием. [139] (Эндогастральный означает, что раковина изогнута так, что вентральная или нижняя сторона вогнута в продольном направлении (брюшко внутрь); экзогастральный означает, что раковина изогнута так, что вентральная сторона выпукла в продольном направлении (брюшко наружу), что позволяет воронке быть направленной назад под раковину .) [139]

Предки колеоидов (включая большинство современных головоногих моллюсков) и предки современных наутилусов разошлись во флоианском веке раннего ордовика, более 470 миллионов лет назад. [138] [140] Bactritida , девонско-триасовая группа ортоконусов, широко считается парафилетической без колеоидов и аммоноидей, то есть последние группы возникли внутри Bactritida. [141] : 393 Увеличение разнообразия колеоидов и аммоноидей наблюдается примерно в начале девона и соответствует резкому увеличению разнообразия рыб. Это может отражать происхождение двух производных групп. [141]
В отличие от большинства современных головоногих моллюсков, большинство древних разновидностей имели защитный панцирь. Эти раковины сначала имели коническую форму, но позже превратились в изогнутые наутилоидные формы, наблюдаемые у современных видов наутилусов .Считается, что конкурентное давление со стороны рыбы вынудило формы с панцирем погрузиться в более глубокие воды, что обеспечило эволюционное давление в сторону потери панциря и привело к появлению современных колеоидов, изменение, которое привело к большим метаболическим издержкам, связанным с потерей плавучести, но позволило им повторно заселить мелководье. [132] : 36 Однако некоторые из наутилоидей с прямым панцирем превратились в белемнитов , из которых некоторые превратились в кальмаров и каракатиц . [ нужна проверка ] Потеря панциря также могла быть результатом эволюционного давления, направленного на повышение маневренности, что привело к появлению более рыбьего образа жизни. [1] : 289
Ведутся споры об эмбриологическом происхождении придатков головоногих моллюсков. [142] До середины двадцатого века широко признавалась гипотеза «Руки как голова». Согласно этой теории, руки и щупальца головоногих похожи на придатки головы брюхоногих моллюсков, что позволяет предположить, что они могут быть гомологичными структурами . Придатки головоногих моллюсков окружают рот, поэтому логически они могли произойти из эмбриональных тканей головы. [143] Однако гипотеза «Руки как ноги», предложенная Адольфом Наефом в 1928 году, получает все большее предпочтение; [142] например, картирование судьбы зачатков конечностей у наутилуса с камерами показывает, что зачатки конечностей происходят из эмбриональных тканей «стопы». [144]
Генетика [ править ]
Секвенирование полного генома головоногих моллюсков остается сложной задачей для исследователей из-за длины и повторения их ДНК. [145] Первоначально предполагалось, что характеристики геномов головоногих являются результатом дупликации всего генома . После полного секвенирования калифорнийского двухточечного осьминога геном показал сходство с геномом других морских беспозвоночных со значительными дополнениями к геному, который, как предполагается, уникален для головоногих моллюсков. Никаких доказательств полной дупликации генома обнаружено не было. [146]
В геноме калифорнийского двухточечного осьминога имеются существенные репликации двух семейств генов. Примечательно, что ранее было известно, что расширенные семейства генов демонстрируют репликативное поведение только у позвоночных. [146] Первое семейство генов было идентифицировано как протокадгерины , которые отвечают за развитие нейронов. Протокадгерины действуют как молекулы клеточной адгезии, необходимые для синаптической специфичности . Механизм репликации семейства генов протокадгеринов у позвоночных объясняется сложным сплайсингом или вырезанием и вставкой из локуса. После секвенирования калифорнийского двухточечного осьминога исследователи обнаружили, что семейство генов пророкадгерина у головоногих расширилось в геноме из-за тандемной дупликации генов . Различные механизмы репликации генов протокадгерина указывают на независимую эволюцию экспансии гена протокадгерина у позвоночных и беспозвоночных. [146] Анализ отдельных генов протокадгерина головоногих указывает на независимую эволюцию между видами головоногих. Вид берегового кальмара Doryteuthis pealeii с расширенным семейством генов протокадгерина значительно отличается от такового у калифорнийского двухпятнистого осьминога, что позволяет предположить, что расширение генов не происходило до видообразования внутри головоногих моллюсков. Несмотря на разные механизмы экспансии генов, гены протокадгерина двухпятнистого осьминога были более похожи на гены позвоночных, чем на кальмаров, что предполагает конвергентный механизм эволюции . Второе семейство генов, известное как C 2 H 2 представляют собой небольшие белки, которые действуют как факторы транскрипции цинка . Считается, что C 2 H 2 регулирует функции ДНК, РНК и белка внутри клетки. [145]
Секвенированный геном калифорнийского двухточечного осьминога также показал значительное присутствие мобильных элементов , а также экспрессию транспозонов. Хотя роль мобильных элементов у морских позвоночных все еще относительно неизвестна, наблюдается значительная экспрессия транспозонов в тканях нервной системы. [147] В исследовании, проведенном на позвоночных, экспрессия транспозонов во время развития у плодовой мухи Drosophila melanogaster активировала геномное разнообразие между нейронами. [148] Это разнообразие связано с улучшением памяти и обучения у млекопитающих. Связь между транспозонами и повышенной способностью нейронов может дать представление о наблюдаемом интеллекте, памяти и функциях головоногих моллюсков. [147]
Используя длительное секвенирование, исследователи расшифровали геномы головоногих моллюсков и обнаружили, что они были перемешаны и перемешаны. Гены сравнивали с генами тысяч других видов, и хотя блоки из трех или более генов встречались одновременно у кальмаров и осьминогов, эти блоки генов не были обнаружены вместе ни у каких других животных». Многие из группировок находились в нервной ткани, что указывает на то, как они адаптировали свой интеллект. [149] [150]
Филогения [ править ]
Приблизительный консенсус относительно филогении современных головоногих моллюсков, согласно Whalen & Landman (2022), показан на кладограмме . [151] Минерализованные таксоны выделены жирным шрифтом .
| Головоногие моллюски |
| ||||||||||||||||||






Внутреннюю филогению головоногих трудно определить; было использовано множество молекулярных методов, но полученные результаты противоречивы. [86] [152] Наутилус имеет тенденцию считаться внешней группой, а Вампиротеутис образует внешнюю группу по отношению к другим кальмарам; однако в одном анализе наутилоиды, осьминоги и тевтиды представляют собой политомию . [86] Некоторые молекулярные филогении не обнаруживают минерализованные колеоиды ( Spirula , Sepia и Metasepia ) как кладу; однако другие находят эту, казалось бы, более экономную кладу, причем Spirula является сестринской группой Sepia и Metasepia в кладе, которая, вероятно, разошлась до конца триаса. [153] [154]
Молекулярные оценки дивергенции клад различаются. Согласно одной «статистически достоверной» оценке, ± 24 миллиона лет Наутилус отделился от Осьминога 415 назад . [155]
Таксономия [ править ]





Представленная здесь классификация современных головоногих моллюсков во многом следует из Текущей классификации современных головоногих моллюсков (май 2001 г.), а для ископаемых головоногих взята из Arkell et al. 1957, Тейхерт и Мур, 1964, Тейхерт, 1988 и другие. Эти три подкласса являются традиционными и соответствуют трем отрядам головоногих моллюсков, признанным Батером. [156]
Класс Головоногие († указывает на вымершие группы)
- Подкласс Nautiloidea : основные эктохлиальные головоногие моллюски, которые послужили источником Ammonoidea и Coleoidea.
- Отряд † Plectronocerida : предки головоногих моллюсков кембрийского периода.
- Отряд † Ellesmerocerida ( от 500 до 470 млн лет назад )
- Отряд † Endocerida ( от 485 до 430 млн лет назад )
- Отряд † Actinocerida ( от 480 до 312 млн лет назад ).
- Отряд † Discosorida ( от 482 до 392 млн лет назад )
- Отряд † Pseudorthocerida ( от 432 до 272 млн лет назад )
- Отряд † Tarphycerida ( от 485 до 386 млн лет назад )
- Отряд † Онкоцериды ( от 478,5 до 324 млн лет назад )
- Отряд Nautilida (сохранившийся; от 410,5 млн лет до настоящего времени)
- Отряд † Orthocerida ( от 482,5 до 211,5 млн лет назад )
- Отряд † Ascocerida ( от 478 до 412 млн лет назад )
- Отряд † Бактритиды ( от 418,1 до 260,5 млн лет назад )
- Подкласс † Ammonoidea : аммониты ( от 479 до 66 млн лет назад )
- Отряд † Goniatitida ( от 388,5 до 252 млн лет назад )
- Отряд † Цератитид ( от 254 до 200 млн лет назад )
- Отряд † Аммонитида ( от 215 до 66 млн лет назад )
- Подкласс Coleoidea (410,0 млн лет назад)
- Когорта † Belemnoidea : белемниты и родственники.
- Род † Jeletzkya.
- Отряд † Aulacocerida ( от 265 до 183 млн лет назад )
- Отряд † Phragmoteuthida ( от 189,6 до 183 млн лет назад )
- Отряд † Hematitida ( от 339,4 до 318,1 млн лет назад )
- Отряд † Белемнитида ( от 339,4 до 66 млн лет назад )
- Род † Belemnoteuthis ( от 189,6 до 183 млн лет назад ).
- когорта Неоколооидная
- Надотряд Decapodiformes (также известный как Decabrachia или Decembranchiata)
- Отряд Spirulida : кальмар из бараньего рога.
- Отряд Sepiida : каракатица .
- Отряд Sepiolida : карликовый, бобтейл и бутылочнохвостый кальмар.
- Заказать Идиосепида
- Отряд Oegopsida : неритический кальмар.
- Отряд Myopsida : прибрежный кальмар.
- Заказать Батитеутиду
- Надотряд Octopodiformes (также известный как Vampyropoda)
- Семейство † Trachyteuthididae.
- Отряд Vampyromorphida : кальмар-вампир.
- Отряд Octopoda : осьминоги.
- Надотряд † Palaeoteuthomorpha
- Орден † Болецкиида
- Надотряд Decapodiformes (также известный как Decabrachia или Decembranchiata)
- Когорта † Belemnoidea : белемниты и родственники.
Другие классификации различаются, прежде всего, тем, как связаны различные отряды десятиногих и должны ли они быть отрядами или семействами.
Трактата классификация Надсемейная
Это более старая классификация, которая объединяет классификации, найденные в частях K и L «Трактата о палеонтологии беспозвоночных» , которая составляет основу и в значительной степени сохраняется в классификациях, появившихся позже.
Последовательность наутилоидов в целом (Teichert and Moore, 1964) указана.
- Подкласс † Endoceratoidea . Не используются Флауэром, например Flower and Kummel 1950, интерджоцериды включены в Endocerida.
- Заказать † Эндоцерида
- Заказать † Интерхоцерида
- Подкласс † Actinoceratoidea Не используется Flower, там же
- Отряд † Актиноцериды.
- Подкласс Nautiloidea Nautiloidea в узком смысле.
- Отряд † Ellesmerocerida Plectronocerida впоследствии выделился в отдельный отряд.
- Отряд † Orthocerida Включает ортоцериды и псевдортоцериды.
- Отряд † Аскоцерида
- Заказать † Онкоцерида
- Отряд † Дискосорида
- Отряд † Тарфицериды
- Отряд † Barrandeocerida Полифилетическая группа, теперь включенная в состав Tarphycerida.
- Заказать Наутилиду
- Подкласс † Bactritoidea
- Отряд † Бактритиды.
Палеозойские аммоноидеи (Миллер, Ферниш и Шиндевольф, 1957)
- Подотряд † Anarcestina
- Подотряд † Clymeniina
- Подотряд † Goniatitina
- Подотряд † Prolecanitina
Мезозойские аммоноидеи (Аркель и др., 1957).
- Подотряд † Кератитин
- Подотряд † Phylloceratina
- Подотряд † Lytoceratina
- Подотряд † Ammonitina
Последующие изменения включают создание трех верхнекембрийских отрядов: Plectronocerida, Protactinocerida и Yanhecerida; выделение псевдортоцерид как Pseudorthocerida и повышение ортоцератид до подкласса Orthoceratoidea.
Классификация Шевырева [ править ]
Шевырев (2005) предложил разделение на восемь подклассов, в основном состоящих из более разнообразных и многочисленных ископаемых форм: [157] [158] хотя эту классификацию критиковали как произвольную, лишенную доказательств и основанную на неправильной интерпретации других статей. [159]




Класс Головоногие
- Подкласс † Ellesmeroceratoidea
- Отряд † Plectronocerida ( от 501 до 490 млн лет назад )
- Отряд † Протактиноцериды
- Орден † Янхесерида
- Отряд † Ellesmerocerida ( от 500 до 470 млн лет назад )
- Подкласс † Endoceratoidea ( от 485 до 430 млн лет назад )
- Отряд † Endocerida ( от 485 до 430 млн лет назад )
- Отряд † Intejocerida ( от 485 до 480 млн лет назад )
- Подкласс † Actinoceratoidea
- Отряд † Actinocerida ( от 480 до 312 млн лет назад ).
- Подкласс Nautiloidea (490,0 млн лет назад)
- Отряд † Basslerocerida ( от 490 до 480 млн лет назад )
- Отряд † Tarphycerida ( от 485 до 386 млн лет назад )
- Отряд † Литуитида ( от 485 до 480 млн лет назад )
- Отряд † Discosorida ( от 482 до 392 млн лет назад )
- Отряд † Онкоцериды ( от 478,5 до 324 млн лет назад )
- Отряд Nautilida (410,5 млн лет назад)
- Подкласс † Orthoceratoidea ( от 482,5 до 211,5 млн лет назад )
- Отряд † Orthocerida ( от 482,5 до 211,5 млн лет назад )
- Отряд † Ascocerida ( от 478 до 412 млн лет назад )
- Отряд † Dissidocerida ( от 479 до 457,5 млн лет назад )
- Отряд † Байкалоцерида.
- Подкласс † Bactritoidea ( от 422 до 252 млн лет назад )
- Подкласс † Ammonoidea ( от 410 до 66 млн лет назад )
- Подкласс Coleoidea (410,0 млн лет назад) [160]
классификация Кладистическая

Другая недавняя система делит всех головоногих моллюсков на две клады . Один включает наутилуса и большинство ископаемых наутилоидей. Другая клада ( Neocephalopoda или Angusteradulata ) ближе к современным колеоидам и включает белемноидов, аммоноидей и многие семейства ортоцерид . Существуют также головоногие стволовой группы традиционных Ellesmerocerida , которые не принадлежат ни к одной кладе. [161] [162]
Колеоиды, несмотря на некоторые сомнения, [1] : 289 Судя по молекулярным данным, они являются монофилетическими. [163]
В культуре [ править ]

Древние мореплаватели знали о головоногих моллюсках, о чем свидетельствуют такие произведения искусства, как резьба по камню, найденная во время археологических раскопок на минойском Крите бронзового века в Кноссе (1900–1100 гг. до н. э.), на которой изображен рыбак, несущий осьминога. [164] Ужасающе могущественная Горгона из греческой мифологии, возможно, была вдохновлена осьминогом или кальмаром: тело осьминога представляет собой отрубленную голову Медузы , клюв — высунутый язык и клыки, а его щупальца — змей. [165]

Кракен . — легендарное морское чудовище гигантских размеров, которое, как говорят, обитает у берегов Норвегии и Гренландии. В искусстве его обычно изображают в виде гигантских головоногих моллюсков, нападающих на корабли Линней включил его в первое издание своей «Системы природы» 1735 года . [166] [167] В гавайском мифе о творении , в котором говорится, что нынешний космос является последним из серии, возникшей поэтапно из руин предыдущей вселенной, осьминог — единственный выживший из предыдущей, чужой вселенной. [168] Аккорокамуи айнов — гигантское чудовище с щупальцами из фольклора . [169]
Битва с осьминогом играет значительную роль в Виктора Гюго книге Travailleurs de la mer ( «Труженики моря »), посвященной его пребыванию в изгнании на Гернси . [170] Сборник рассказов Яна Флеминга 1966 года «Осьминожка и живой свет дневного света» и о Джеймсе Бонде фильм 1983 года были частично вдохновлены книгой Хьюго. [171]
Японское эротическое искусство, сюнга , включает гравюры на дереве укиё-э, такие как гравюра Кацусики Хокусая 1814 года « Тако то ама» ( «Сон жены рыбака» ), на которой ныряльщик ама сексуально переплетается с большим и маленьким осьминогом. [172] [173] Принт является предшественником щупальцевой эротики . [174]
Его многочисленные руки, исходящие из общего центра, означают, что осьминог иногда используется как символ мощной и манипулятивной организации. [175]
См. также [ править ]
- Размер головоногих моллюсков
- Головоногий глаз
- Головоногий интеллект
- Боль у головоногих моллюсков
- Кракен
- Список наутилоидей
- Список аммонитов
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д и ж г час я Уилбур, Карл М.; Труман, скорая помощь; Кларк, MR, ред. (1985), Моллюска , том. 11. Форма и функция, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ Jump up to: Перейти обратно: а б Хильденбранд, Энн; Аустерманн, Грегор; Фукс, Дирк; Бенгтсон, Питер; Стиннесбек, Вольфганг (2021). «Потенциальный головоногий моллюск из раннего кембрия восточного Ньюфаундленда, Канада» . Коммуникационная биология . 4 (1): 388. doi : 10.1038/s42003-021-01885-w . ПМЦ 7987959 . ПМИД 33758350 .
- ^ Кейруш, К.; Кантино, PD; Готье, JA (2020). Филонимы: дополнение к Филокоду . ЦРК Пресс. п. 1843. ISBN 978-1-138-33293-5 .
- ^ «Добро пожаловать в CephBase» . CephBase . Архивировано из оригинала 12 января 2016 года . Проверено 29 января 2016 г.
- ^ Jump up to: Перейти обратно: а б с д Уилбур, Карл М.; Кларк, MR; Труман, ER, ред. (1985), Моллюска , том. 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ Бартол, ИК; Манн, Р.; Веккьоне, М. (2002). «Распространение эвригалинного кальмара Lolliguncula brevis в Чесапикском заливе: влияние отдельных абиотических факторов» . Серия «Прогресс в области морской экологии» . 226 : 235–247. Бибкод : 2002MEPS..226..235B . дои : 10.3354/meps226235 .
- ^ «Есть ли пресноводные головоногие моллюски?» . Азбука науки . 16 января 2013 г.
- ^ «Головоногие моллюски наблюдались на рекордных океанских глубинах» .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л Никсон, Мэрион; Янг, Джей Зи (2003). Мозг и жизнь головоногих . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-852761-9 .
- ^ Трикарико, Э.; Амодио, П.; Понте, Г.; Фиорито, Г. (2014). «Познание и распознавание головоногих моллюсков Octopus vulgaris : координация взаимодействия с окружающей средой и сородичами». В Вицани, Г. (ред.). Биокоммуникация животных . Спрингер. стр. 337–349. ISBN 978-94-007-7413-1 .
- ^ Будельманн, Бу (1995). «Нервная система головоногих моллюсков: что эволюция сделала с дизайном моллюсков» . В Брайдбахе, О.; Кач, В. (ред.). Нервные системы беспозвоночных: эволюционный и сравнительный подход . Спрингер. ISBN 978-3-7643-5076-5 .
- ^ Чунг, Вэнь-Сун; Курниаван, Ньоман Д.; Маршалл, Н. Джастин (2020). «К мезомасштабному коннектому мозга кальмара на основе МРТ» . iScience . 23 (1): 100816. Бибкод : 2020iSci...23j0816C . дои : 10.1016/j.isci.2019.100816 . ПМЦ 6974791 . ПМИД 31972515 .
- ^ Рэйвен, Питер; и др. (2003). Биология . Нью-Йорк: Образование Макгроу-Хилл. п. 669. ИСБН 9780073383071 .
- ^ Тасаки, И.; Такенака, Т. (1963). «Покой и потенциал действия гигантских аксонов кальмаров, внутриклеточно перфузированных растворами, богатыми натрием» . Труды Национальной академии наук Соединенных Штатов Америки . 50 (4): 619–626. Бибкод : 1963PNAS...50..619T . дои : 10.1073/pnas.50.4.619 . ПМК 221236 . ПМИД 14077488 .
- ^ Jump up to: Перейти обратно: а б Паккард, А. (1972). «Головоногие моллюски и рыбы: пределы сближения». Биологические обзоры . 47 (2): 241–307. дои : 10.1111/j.1469-185X.1972.tb00975.x . S2CID 85088231 .
- ^ Масия, Сильвия; Робинсон, Майкл П.; Безумие, Пол; Далтон, Роберт; Томас, Джеймс Д. (2004). «Новые наблюдения за воздушно-реактивным движением (полетом) кальмаров с обзором предыдущих отчетов» . Журнал исследований моллюсков . 70 (3): 297–299. дои : 10.1093/моллюс/70.3.297 .
- ^ Jump up to: Перейти обратно: а б Мурамацу, К.; Ямамото, Дж.; Абэ, Т.; Секигути, К.; Хоши, Н.; Сакурай, Ю. (2013). «Океанические кальмары летают». Морская биология . 160 (5): 1171–1175. Бибкод : 2013МарБи.160.1171М . дои : 10.1007/s00227-013-2169-9 . S2CID 84388744 .
- ^ «Ученые разгадали тайну летающего кальмара» . Вид на океан . Нэшнл Географик. 20 февраля 2013 г. Архивировано из оригинала 5 августа 2019 г. . Проверено 5 августа 2019 г.
- ^ Джабр, Феррис (2 августа 2010 г.). «Факт или вымысел: может ли кальмар вылететь из воды?» . Научный американец .
- ^ Jump up to: Перейти обратно: а б Серб, Ю.М.; Эрниссе, диджей (2008). «Изложение траектории эволюции: использование разнообразия глаз моллюсков для понимания параллельной и конвергентной эволюции» . Эволюция: образование и информационно-пропагандистская деятельность . 1 (4): 439–447. дои : 10.1007/s12052-008-0084-1 . S2CID 2881223 .
- ^ Уэллс, Мартин Дж. (2011). «Часть М, Глава 4: Физиология колеоидов» . Трактат онлайн . Лоуренс, Канзас, США. дои : 10.17161/to.v0i0.4226 . Архивировано из оригинала 22 августа 2016 г. Проверено 10 мая 2013 г. (требуется подписка)
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к Бойл, Питер; Родхаус, Пол (2004). Головоногие моллюски: экология и рыболовство . Блэквелл. дои : 10.1002/9780470995310.ch2 . ISBN 978-0-632-06048-1 .
- ^ Jump up to: Перейти обратно: а б Мессенджер, Джон Б.; Хэнлон, Роджер Т. (1998). Поведение головоногих . Кембридж: Издательство Кембриджского университета. стр. 17–21. ISBN 978-0-521-64583-6 .
- ^ Чунг, Вэнь-Сун; Маршалл, Н. Джастин (14 сентября 2016 г.). «Сравнительная визуальная экология головоногих моллюсков из разных местообитаний» . Труды Королевского общества B: Биологические науки . 283 (1838): 20161346. doi : 10.1098/rspb.2016.1346 . ISSN 0962-8452 . ПМК 5031660 . ПМИД 27629028 .
- ^ Хэнлон и Мессенджер, 68.
- ^ Метгер, Л.; Робертс, С.; Хэнлон, Р. (2010). «Доказательства распределенного восприятия света в коже каракатицы Sepia officinalis » . Письма по биологии . 6 (5): 600–603. дои : 10.1098/rsbl.2010.0223 . ПМЦ 2936158 . ПМИД 20392722 .
- ^ Мичиномаэ, М.; Масуда, Х.; Сейду, М.; Кито, Ю. (1994). «Структурная основа распознавания длины волны в сетчатке кальмара-светлячка Watasenia scintillans » . Журнал экспериментальной биологии . 193 (1): 1–12. дои : 10.1242/jeb.193.1.1 . ПМИД 9317205 .
- ^ Сейду, М.; Сугахара, М.; Утияма, Х.; Хираки, К.; Хаманака, Т.; Мичиномаэ, М.; Ёшихара, К.; Кито, Ю. (1990). «О трёх зрительных пигментах сетчатки кальмара-светлячка Watasenia scintillans ». Журнал сравнительной физиологии А. 166 (6). дои : 10.1007/BF00187321 . S2CID 25707481 .
- ^ Стаббс, Алабама; Стаббс, CW (2015). «Новый механизм цветового зрения: форма зрачка и хроматическая аберрация могут обеспечить спектральное распознавание организмов, страдающих цветовой слепотой». биоRxiv 10.1101/017756 .
- ^ «Глаза осьминога безумнее, чем мы себе представляли» . Гизмодо . 06.07.2016. Архивировано из оригинала 30 апреля 2023 г.
- ^ Jump up to: Перейти обратно: а б Кингстон, AC; Кузирян А.М.; Хэнлон, RT; Кронин, Т.В. (2015). «Компоненты зрительной фототрансдукции в хроматофорах головоногих предполагают кожную фоторецепцию» . Журнал экспериментальной биологии . 218 (10): 1596–1602. дои : 10.1242/jeb.117945 . hdl : 11603/13387 . ПМИД 25994635 . S2CID 25431963 .
- ^ «Голоногие моллюски вас слышат» . Новости Би-би-си . 15 июня 2009 г. Проверено 28 апреля 2010 г.
- ^ Тонг, Д.; Розас, С.; Окли, Х.; Митчелл, Дж.; Колли, Дж.; Макфолл-Нгай, Дж. (июнь 2009 г.). «Доказательства восприятия света биолюминесцентным органом» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (24): 9836–9841. Бибкод : 2009PNAS..106.9836T . дои : 10.1073/pnas.0904571106 . ISSN 0027-8424 . ПМК 2700988 . ПМИД 19509343 .
- ^ «Покровы (моллюски)» . Британская энциклопедия. 2009. Британская энциклопедия 2006, DVD Ultimate Reference Suite .
- ^ Рамирес, доктор медицины; Окли, TH (2015). «Глазонезависимое, активируемое светом расширение хроматофора (LACE) и экспрессия генов фототрансдукции в коже Octopus bimaculoides » (PDF) . Журнал экспериментальной биологии . 218 (10): 1513–1520. дои : 10.1242/jeb.110908 . ПМЦ 4448664 . ПМИД 25994633 . Архивировано (PDF) из оригинала 9 августа 2016 г.
- ^ Сильвия Лима де Соуза Медейрос; и др. (2021). «Циклическое чередование состояний спокойного и активного сна у осьминога» . iScience . дои : 10.1016/j.isci.2021.102223 . ПМК 8101055 .
- ^ Хаффард, Кристин Л. (1 октября 2006 г.). «Передвижение Abdopus aculeatus (Cephalopoda: Octopodidae): пройти грань между первичной и вторичной защитой» . Журнал экспериментальной биологии . 209 (19): 3697–3707. дои : 10.1242/jeb.02435 . ISSN 0022-0949 . ПМИД 16985187 . S2CID 26862414 .
- ^ Йозеф, Ноам; Амодио, Пьеро; Фиорито, Грациано; Шашар, Надав (23 мая 2012 г.). «Маскировка в сложной среде: осьминоги используют особенности окружающей среды для сопоставления с фоном» . ПЛОС ОДИН . 7 (5): e37579. Бибкод : 2012PLoSO...737579J . дои : 10.1371/journal.pone.0037579 . ISSN 1932-6203 . ПМК 3359305 . ПМИД 22649542 .
- ^ Манда, Стивен; Турек, Войтех (2009). «Минутные силурийские наутилоиды-онкоцериды с необычной окраской» . Acta Palaeontologica Polonica . 54 (3): 503–512. дои : 10.4202/app.2008.0062 . S2CID 54043278 .
- ^ Турек, Войтех (2009). «Цветовые узоры головоногих моллюсков раннего девона из Баррандия: тафономия и таксономия» . Acta Palaeontologica Polonica . 54 (3): 491–502. дои : 10.4202/app.2007.0064 . S2CID 55851070 .
- ^ Хэнлон, Р.; и др. (2011). «Быстрая адаптивная маскировка у головоногих моллюсков». Камуфляж животных: механизмы и функции . Кембридж, Великобритания: Издательство Кембриджского университета. стр. 145–161.
- ^ Клони, Ричард А.; Брокко, Стивен Л. (1983). «Хроматофорные органы, клетки-рефлекторы, иридоциты и лейкофоры головоногих» . Американский зоолог . 23 (3): 581–592. дои : 10.1093/icb/23.3.581 .
- ^ Норман, Марк Д.; Финн, Джулиан; Трегенза, Том (2001). «Динамическая мимикрия индо-малайского осьминога» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 268 (1478): 1755–1758. дои : 10.1098/rspb.2001.1708 . ПМЦ 1088805 . ПМИД 11522192 .
- ^ Стивенс, М.; Мерайлайта С., ред. (2011). Камуфляж животных: Механизмы и функции . Издательство Кембриджского университета.
- ^ Стюарт-Фокс, Деви; Муссалли, Аднан (2008). «Отбор социальных сигналов стимулирует эволюцию изменения цвета хамелеона» . ПЛОС Биология . 6 (1): е25. doi : 10.1371/journal.pbio.0060025 . ПМК 2214820 . ПМИД 18232740 .
- ^ Баньяра, Джей Ти; Хэдли, Мэн (1973). Хроматофоры и изменение цвета: сравнительная физиология пигментации животных . Прентис-Холл.
- ^ Демский, Лео С. (1992). «Хроматофорные системы костистых и головоногих моллюсков: уровнево-ориентированный анализ конвергентных систем» . Мозг, поведение и эволюция . 40 (2–3): 141–156. дои : 10.1159/000113909 . ПМИД 1422807 .
- ^ Хэнлон, Роджер Т.; Мессенджер, Джон Б. (1999). Поведение головоногих . Издательство Кембриджского университета. п. 2. ISBN 978-0-521-64583-6 .
- ^ «чернильная рыба» . Мерриам-Вебстер . Проверено 1 февраля 2018 г.
- ^ Бикердайк, Джон (1895). Морская рыбалка . Лондон: Longmans, Green and Co. p. 114.
Кальмар обыкновенный или кальмар ( Loligo vulgaris ). Иногда ее называют рыбой-пером и чернилами из-за ее чернильного мешка и нежной удлиненной раковины, находящейся внутри него.
- ^ Уэллс, MJ (1 апреля 1980 г.). «Нервная регуляция сердцебиения у осьминогов» . Журнал экспериментальной биологии . 85 (1): 111–28. дои : 10.1242/jeb.85.1.111 . ПМИД 7373208 .
- ^ Гиретти-Магальди, А. (октябрь 1992 г.). «Предыстория гемоцианина. Открытие меди в крови моллюсков». Клеточные и молекулярные науки о жизни . 48 (10): 971–972. дои : 10.1007/BF01919143 . S2CID 33290596 .
- ^ «Сколько сердец у осьминога?» . Новый учёный .
- ^ «Почему у осьминогов три сердца?» . Наука Азбука . 29 февраля 2020 г.
- ^ «Где бы мы были без крови?» . Научный совет Иллинойса . Архивировано из оригинала 25 апреля 2023 г. Проверено 7 июля 2022 г.
- ^ «Почему кровь осьминога синяя?» . Уилстар .
- ^ Jump up to: Перейти обратно: а б с Гилберт, Дэниел Л.; Адельман, Уильям Дж.; Арнольд, Джон М. (1990). Кальмары как экспериментальные животные (иллюстрированное издание). Спрингер. ISBN 978-0-306-43513-3 .
- ^ Jump up to: Перейти обратно: а б с Шипп, Рудольф; Молленхауэр, Стефан; Болецкий, Сигурд (1979). «Электронно-микроскопические и гистохимические исследования дифференцировки и функции жабр головоногих моллюсков ( Sepia Officinalis L.)». Зооморфология . 93 (3): 193–207. дои : 10.1007/BF00993999 . S2CID 20214206 .
- ^ Jump up to: Перейти обратно: а б с Уэллс, MJ; О'Дор, РК (1 сентября 1991 г.). «Реактивное движение и эволюция головоногих» . Бюллетень морской науки . 49 (1–2): 419–432.
- ^ Паккард, А.; Труман, скорая помощь (октябрь 1974 г.). «Мышечная деятельность мантии Sepia и Loligo (Cephalopoda) при дыхательных движениях и струйных движениях и ее физиологическая интерпретация» . Журнал экспериментальной биологии . 61 (2): 411–419. дои : 10.1242/jeb.61.2.411 . ISSN 0022-0949 . ПМИД 4443736 .
- ^ Боун, К.; Браун, скорая помощь; Трэверс, Г. (1994). «О дыхательном потоке каракатицы Sepia Officinalis » (PDF) . Журнал экспериментальной биологии . 194 (1): 153–165. дои : 10.1242/jeb.194.1.153 . ПМИД 9317534 . Архивировано (PDF) из оригинала 4 марта 2009 г.
- ^ Коул, А.; Холл, Б. (2009). «Дифференциация хряща головоногих моллюсков». Зоология . 112 (1): 2–15. Бибкод : 2009Zool..112....2C . дои : 10.1016/j.zool.2008.01.003 . ПМИД 18722759 .
- ^ См. также http://tolweb.org/articles/?article_id=4200. Архивировано 16 июня 2010 г. на Wayback Machine.
- ^ Jump up to: Перейти обратно: а б с д и Уилбур, Карл М.; Кларк, MR; Труман, ER, ред. (1985), «11: Эволюция плавучести и передвижения у современных головоногих моллюсков», The Mollusca , vol. 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ Jump up to: Перейти обратно: а б Андерсон, Э.; Демонт, М. (2000). «Механика передвижения кальмара Loligo pealei : Локомоторная функция и нестационарная гидродинамика струйного и внутримантийного давления» . Журнал экспериментальной биологии . 203 (18): 2851–2863. дои : 10.1242/jeb.203.18.2851 . ПМИД 10952883 .
- ^ Jump up to: Перейти обратно: а б с Бартол, ИК; Крюгер, PS; Томпсон, Джей Ти; Стюарт, WJ (2008). «Динамика плавания и двигательная эффективность кальмаров на протяжении онтогенеза» . Интегративная и сравнительная биология . 48 (6): 720–733. дои : 10.1093/icb/icn043 . ПМИД 21669828 .
- ^ Ши, Э.; Веккьоне, М. (2002). «Количественная оценка онтогенетических нарушений у трех видов кальмаров-эгопсидов с использованием кусочно-линейной регрессии модели II». Морская биология . 140 (5): 971–979. Бибкод : 2002MarBi.140..971E . дои : 10.1007/s00227-001-0772-7 . S2CID 84822175 .
- ^ Джонсон, В.; Соден, ПД; Труман, скорая помощь (февраль 1972 г.). «Исследование реактивного движения: анализ движения кальмара Loligo vulgaris » . Журнал экспериментальной биологии . 56 (1972): 155–165. дои : 10.1242/jeb.56.1.155 .
- ^ Кэмпбелл, Рис и Митчелл (1999) , стр. 612.
- ^ Герра, А.; Мартинелл, X.; Гонсалес, А.Ф.; Веккьоне, М.; Грасия, Дж.; Мартинелл, Дж. (2007). «В океане обнаружен новый шум». Журнал Морской биологической ассоциации Соединенного Королевства . 87 (5): 1255–1256. Бибкод : 2007JMBUK..87.1255G . дои : 10.1017/S0025315407058225 . hdl : 10261/27009 . S2CID 85770435 .
- ^ Jump up to: Перейти обратно: а б Уэллс, Мартин Дж.; О'Дор, РК (июль 1991 г.). «Реактивное движение и эволюция головоногих». Бюллетень морской науки . 49 (1): 419–432 (14).
- ^ Чемберлен-младший (1993). «Передвижение в древних морях: ограничения и возможности адаптивного дизайна головоногих». Геобиос . 26 (Приложение 1): 49–61. Бибкод : 1993Geobi..26...49C . дои : 10.1016/S0016-6995(06)80360-8 .
- ^ Jump up to: Перейти обратно: а б с д и О'Дор, РК (1988). «Силы, действующие на плавающего кальмара» . Журнал экспериментальной биологии . 137 : 421–442. дои : 10.1242/jeb.137.1.421 .
- ^ О'Дор, РК; Хоар, Дж. А. (2000). «Ограничивает ли геометрия рост кальмаров?» . Журнал морских наук ICES . 57 (1): 8–14. Бибкод : 2000ICJMS..57....8O . дои : 10.1006/jmsc.1999.0502 .
- ^ Сейбель, бакалавр; Туезен, Э.В.; Чилдресс, Джей-Джей; Городецкий, Луизиана (апрель 1997 г.). «Снижение пелагического метаболизма головоногих моллюсков с увеличением глубины обитания отражает различия в двигательной эффективности» . Биологический вестник . 192 (2): 262–278. дои : 10.2307/1542720 . ISSN 1939-8697 . JSTOR 1542720 . ПМИД 28581868 .
- ^ Хэнлон, Роджер Т.; Уотсон, Аня С.; Барбоза, Александра (01 февраля 2010 г.). «Осьминог-мимик» в Атлантике: мимикрия и камуфляж камбалы Macrotritopus defilippi ». Биологический вестник . 218 (1): 15–24. дои : 10.1086/BBLv218n1p15 . hdl : 1912/4811 . ISSN 0006-3185 . ПМИД 20203250 . S2CID 12935620 .
- ^ «Оболочка аргонавта: контроль плавучести с помощью газа у пелагического осьминога» .
- ^ Jump up to: Перейти обратно: а б с д и ж Гослайн, Джон М.; де Мон, М. Эдвин (1985). «Реактивное плавание в кальмарах» . Научный американец . Том. 252, нет. 1. С. 96–103. doi : 10.1038/scientificamerican0185-96 . ISSN 0036-8733 . JSTOR 24967551 .
- ^ Хаффард, Кристин Л. (1 октября 2006 г.). «Передвижение Abdopus aculeatus (Cephalopoda: Octopodidae): Прохождение границы между первичной и вторичной защитой» . Журнал экспериментальной биологии . 209 (19): 3697–3707. дои : 10.1242/jeb.02435 . ISSN 0022-0949 . ПМИД 16985187 . S2CID 26862414 .
- ^ Jump up to: Перейти обратно: а б с д Баратте, С.; Андуш, А.; Бонно, Л. (2007). «Запечатлен в головоногих моллюсках: ключевой ген, связанный с появлением морфологических новинок». Гены развития и эволюция . 217 (5): 353–362. дои : 10.1007/s00427-007-0147-2 . ПМИД 17394016 . S2CID 22241391 .
- ^ фон Болецкий, С. (2004). « Голые аммоноидеи: вызов филогении головоногих моллюсков?» ['Голые аммоноидеи': вызов филогении головоногих моллюсков?]. Геобиос . 37 : 117–118. дои : 10.1016/j.geobios.2003.01.009 .
- ^ Гибсон, Р.Н.; Аткинсон, RJA; Гордон, JDM, ред. (2006). Океанография и морская биология: Ежегодный обзор . ЦРК Пресс. п. 288. ИСБН 978-1420006391 .
- ^ Олдред, Р.Г.; Никсон, М.; Янг, Дж. З. (1983). « Cirrothauma murrayi Chun, осьминог с плавниками» . Философские труды Королевского общества B: Биологические науки . 301 (1103): 1–54. Бибкод : 1983РСТБ.301....1А . дои : 10.1098/rstb.1983.0021 .
- ^ Фукс, Д.; Ифрим, К.; Стиннесбек, В. (2008). «Новый палеоктопус (Cephalopoda: Coleoidea) из позднего мела Вальесильо, северо-восток Мексики, и его значение для эволюции Octopoda» . Палеонтология . 51 (5): 1129–1139. Бибкод : 2008Palgy..51.1129F . дои : 10.1111/j.1475-4983.2008.00797.x .
- ^ фон Болецкий, Сигурд (июль 1991 г.). «Концевой шип детенышей сепиолидов: его развитие и функциональная морфология (Mollusca, Cephalopoda)». Бюллетень морской науки . 49 : 107–112.
- ^ Jump up to: Перейти обратно: а б с Страгнелл, Дж.; Нисигути, МК (2007). «Молекулярная филогения колеоидных головоногих моллюсков (Mollusca: Cephalopoda), выведенная на основе трех митохондриальных и шести ядерных локусов: сравнение выравнивания, подразумеваемого выравнивания и методов анализа» . Журнал исследований моллюсков . 73 (4): 399–410. дои : 10.1093/mollus/eym038 .
- ^ Варнке, К.; Кеупп, Х. (2005). « Спирула – окно в эмбриональное развитие аммоноидей? Морфологические и молекулярные признаки палеонтологической гипотезы» (PDF) . Фации . 51 (1–4): 60–65. Бибкод : 2005Faci...51...60W . дои : 10.1007/s10347-005-0054-9 . S2CID 85026080 .
- ^ Фурухаши, Т.; Шварцингер, К.; Миксик, И.; Смрз, М.; Беран, А. (2009). «Эволюция раковины моллюсков с обзором гипотезы кальцификации раковины». Сравнительная биохимия и физиология Б . 154 (3): 351–371. дои : 10.1016/j.cbpb.2009.07.011 . ПМИД 19665573 .
- ^ Дофин, Ю. (1996). «Органический матрикс колеоидных раковин головоногих моллюсков: молекулярные массы и изоэлектрические свойства растворимого матрикса в связи с процессами биоминерализации» . Морская биология . 125 (3): 525–529. Бибкод : 1996МарБи.125..525Д . дои : 10.1007/BF00353265 . S2CID 83400480 .
- ^ Дофин, Ю. (1983). Основные подразделения класса головоногих: основы современной систематики: вклад микроструктурного анализа . Эти Док. Государственный, Парижский университет Sud. OCLC 972899981 .
- ^ Дофин, Ю. (2001). «Наноструктуры перламутра реальных испытаний головоногих». Палеонтологический журнал . 75 (1): 113–122. дои : 10.1007/bf03022601 . ISSN 0031-0220 . S2CID 126900936 .
- ^ Толл, РБ; Бингер, LC (1991). «Аномалии рук: случаи сверхкомплектного развития и двусторонней агенезии пар рук у осьминоги (Mollusca, Cephalopoda)». Зооморфология . 110 (6): 313–316. дои : 10.1007/BF01668021 . S2CID 34858474 .
- ^ Анатомия обыкновенного кальмара . 1912.
- ^ Никсон, 1988 г., в Виппич, MGE; Леманн, Дж. (2004). « Allocrioceras из сеномана (среднего мела) Ливана и его влияние на палеобиологическую интерпретацию гетероморфных аммонитов» . Палеонтология . 47 (5): 1093–1107. Бибкод : 2004Palgy..47.1093W . дои : 10.1111/j.0031-0239.2004.00408.x .
- ^ Уилбур, Карл М.; Кларк, MR; Труман, ER, ред. (1985), «5», Моллюска , вып. 12. Палеонтология и неонтология головоногих моллюсков, Нью-Йорк: Academic Press, ISBN. 0-12-728702-7
- ^ Саундри, П.; Кливленд, К., ред. (2011). «Кельтское море» . Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде. [ мертвая ссылка ]
- ^ Лена ван Гизен; и др. (2020). «Молекулярные основы хемотаксильной чувствительности у осьминогов». Том. 183, нет. 3. Клетка. стр. 594–604. doi : 10.1016/j.cell.2020.09.008 .
- ^ «Головоногие радулы» . Веб-проект «Древо жизни» .
- ^ Jump up to: Перейти обратно: а б с д и Никсон, М. (1995). «Номенклатура радулы головоногих моллюсков (Mollusca) - живых и ископаемых». Журнал зоологии . 236 : 73–81. дои : 10.1111/j.1469-7998.1995.tb01785.x .
- ^ Jump up to: Перейти обратно: а б Габботт, SE (1999). «Ортоконические головоногие моллюски и связанная с ними фауна из сланцевых сланцев Сум-Лагерштатт позднего ордовика, Южная Африка» . Палеонтология . 42 (1): 123–148. Бибкод : 1999Palgy..42..123G . дои : 10.1111/1475-4983.00065 .
- ^ Ландман, Нил Х.; Дэвис, Ричард Арнольд; Мейпс, Ройал Х., ред. (2007). Головоногие моллюски: настоящее и прошлое: новые идеи и свежие перспективы . Спрингер. ISBN 978-1-4020-6461-6 .
- ^ Ричардсон и ... (1977). Окаменелости Мейсон-Крик .
- ^ Крута, И.; Ландман, Н.; Руже, И.; Чекка, Ф.; Таффоро, П. (2011). «Роль аммонитов в мезозойской морской пищевой сети, выявленная путем сохранения челюстей». Наука . 331 (6013): 70–72. Бибкод : 2011Наука...331...70К . дои : 10.1126/science.1198793 . ПМИД 21212354 . S2CID 206530342 .
- ^ Jump up to: Перейти обратно: а б Барнс, Роберт Д. (1982). Зоология беспозвоночных . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 450–460. ISBN 978-0-03-056747-6 .
- ^ Лоест, Р.А. (1979). «Испарение и поглощение аммиака наземными брюхоногими моллюсками_ сравнение видов с панцирем и без панциря». Физиологическая зоология . 52 (4): 461–469. дои : 10.1086/physzool.52.4.30155937 . JSTOR 30155937 . S2CID 87142440 .
- ^ Jump up to: Перейти обратно: а б с Буше-Родони, Р.; Мангольд, К. (1994). «Продуцирование аммиака у головоногих моллюсков, физиологические и эволюционные аспекты». Морское и пресноводное поведение и физиология . 25 (1–3): 53–60. дои : 10.1080/10236249409378907 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к Видал, Эрика А.Г. Достижения в науке о головоногих: биология, экология, выращивание и рыболовство .
- ^ Jump up to: Перейти обратно: а б с д и Родригес, М.; Герра; Тронкозо (2010). «Эмбриональная фаза и ее влияние на размер и состояние вылупившихся детенышей кальмара атлантического бобтейла Sepiola Atlantica » . Гельголандские морские исследования . 65 (2): 211–216. Бибкод : 2011HMR....65..211R . дои : 10.1007/s10152-010-0217-0 . hdl : 10261/44487 . S2CID 41577834 .
- ^ Jump up to: Перейти обратно: а б с д Архипкин, А.И. (1992). «Структура, развитие и функции репродуктивной системы головоногих моллюсков с новой общей шкалой стадий зрелости» . Журнал науки рыболовства в Северо-Западной Атлантике . 12 : 63–74. дои : 10.2960/j.v12.a7 .
- ^ Моханти, Собхи; Оянгурен, Альфредо Ф.; Фуйман, Ли А. (1 июля 2014 г.). «Агрессивное брачное поведение самцов зависит от зрелости самок Octopus bimaculoides ». Морская биология . 161 (7): 1521–1530. Бибкод : 2014МарБи.161.1521М . дои : 10.1007/s00227-014-2437-3 . ISSN 0025-3162 . S2CID 85256742 .
- ^ Сондерс, В.Б.; Спиноза, К. (1978). «Половой диморфизм у Наутилуса из Палау». Палеобиология . 4 (3): 349–358. Бибкод : 1978Pbio....4..349S . дои : 10.1017/S0094837300006047 . S2CID 85899974 .
- ^ Архипкин А.И.; Лаптиховский, В.В. (2010). «Наблюдение за удлинением пениса у Onykia ingens: значение для переноса сперматофоров у глубоководных кальмаров» . Журнал исследований моллюсков . 76 (3): 299–300. doi : 10.1093/mollus/eyq019 .
- ^ Янг, РБ (1975). «Системный подход к планированию профессиональных программ». Обзор общественного колледжа . 3 (2): 19–25. дои : 10.1177/009155217500300204 . S2CID 145374345 .
- ^ Jump up to: Перейти обратно: а б с Сквайрс, З. Э; Норман, доктор медицины; Стюарт-Фокс, Д. (2013). «Брачное поведение и общие закономерности нереста южного кальмара Euprymna tasmanica » . Журнал исследований моллюсков . 79 (3): 263–269. дои : 10.1093/моллюс/eyt025 .
- ^ «Редкое глубоководное видео показывает, как мама-кальмар несет яйца в целях безопасности» . 9 июля 2022 г.
- ^ Хэнлон, Роджер Т.; Мессенджер, Джон Б. (22 марта 2018 г.). Поведение головоногих . Издательство Кембриджского университета. ISBN 978-0-521-89785-3 .
- ^ Пунита, П.; Виджай, Д.; Ямамото, Дж.; Адачи, К.; Като, Ю.; Сакурай, Ю. (2017). «Строение и свойства яичной массы кальмара-оммастрефида Todarodes pacificus» . ПЛОС ОДИН . 12 (8): e0182261. Бибкод : 2017PLoSO..1282261P . дои : 10.1371/journal.pone.0182261 . ПМК 5540507 . ПМИД 28767686 .
- ^ «Мама глубоководного кальмара несет ослепительную жемчужную нить яиц» . Живая наука . 8 июля 2022 г.
- ^ Ховинг, Х.Дж.; Лаптиховский В.В.; Робисон, Б.Х. (2015). «Репродуктивная стратегия кальмаров-вампиров уникальна среди колеоидных головоногих моллюсков» . Современная биология . 25 (8): Р322-3. Бибкод : 2015CBio...25.R322H . дои : 10.1016/j.cub.2015.02.018 . ПМИД 25898098 .
- ^ Лю, Бенджамин; Песня, Лео; Келкар, Самитра; Рамджи, Анна; Колдуэлл, Рой (2023). «Индивидуально уникальные фиксированные конфигурации полос Octopus chierchiae позволяют проводить фотоидентификацию в долгосрочных исследованиях» . ПЛОС ОДИН . 18 (4): e0265292. Бибкод : 2023PLoSO..1865292L . дои : 10.1371/journal.pone.0265292 . ПМЦ 10096297 . ПМИД 37043461 .
- ^ Марти, HJ; Хаузер, Р; Шолль, А. (1976). «Природный транквилизатор в яйцах головоногих моллюсков». Природа . 261 (5560): 496–7. Бибкод : 1976Natur.261..496M . дои : 10.1038/261496a0 . ПМИД 945466 . S2CID 8693207 .
- ^ Jump up to: Перейти обратно: а б Норман, доктор медицины; Лу, CC (1997). «Переописание южного кальмара-пельмени Euprymna tasmanica и пересмотр рода Euprymna (Cephalopoda: Sepiolidae)». Журнал Морской биологической ассоциации Соединенного Королевства . 77 (4): 1109–1137. Бибкод : 1997JMBUK..77.1109N . дои : 10.1017/s0025315400038662 . S2CID 85748957 .
- ^ Ивата, Ю.; Ито, К.; Сакурай, Ю. (2008). «Влияние низкой температуры на брачное поведение кальмара Loligo bleekeri ». Рыболовная наука . 74 (6): 1345–1347. Бибкод : 2008FisSc..74.1345I . дои : 10.1111/j.1444-2906.2008.01664.x . S2CID 43094931 .
- ^ Ченг, Мэри В.; Колдуэлл, Рой Л. (июль 2000 г.). «Определение пола и спаривание синекольчатого осьминога Hapalochlaena lunulata ». Поведение животных . 60 (1): 27–33. дои : 10.1006/anbe.2000.1447 . ISSN 0003-3472 . ПМИД 10924200 . S2CID 32899443 .
- ^ Jump up to: Перейти обратно: а б Фэйрберн, Д. (2013). «Одеяловой осьминог: дрейфующие самки и карликовые самцы». Странные пары: необычайные различия между полами в животном мире . Издательство Принстонского университета. стр. 104–115.
- ^ Jump up to: Перейти обратно: а б с д Фон Болецкий, С. (2003). Биология ранних стадий жизни головоногих моллюсков . Достижения морской биологии. Том. 44. стр. 143–203. дои : 10.1016/S0065-2881(03)44003-0 . ISBN 978-0-12-026144-4 . ПМИД 12846042 .
- ^ Jump up to: Перейти обратно: а б Сигено, С.; Сасаки, Т.; Моритаки, Т.; Касугай, Т.; Веккьоне, М.; Агата, К. (январь 2008 г.). «Эволюция комплекса головы головоногих моллюсков путем сборки нескольких частей тела моллюсков: данные эмбрионального развития наутилуса ». Журнал морфологии . 269 (1): 1–17. дои : 10.1002/jmor.10564 . ПМИД 17654542 . S2CID 13109195 .
- ^ Гилберт, Дэниел Л.; Адельман, Уильям Дж.; Арнольд, Джон М. (1990). Кальмары как экспериментальные животные . Нью-Йорк: Пленум Пресс. ISBN 978-0-306-43513-3 .
- ^ Молчанивский, Натали А. (2004). «Понимание процесса роста головоногих моллюсков» . Морские и пресноводные исследования . 55 (4): 379–386. дои : 10.1071/MF03147 .
- ^ Лемче, Х.; Вингстранд, КГ (1959). «Анатомия Neopilina galatheae Lemche, 1957 (Mollusca, Tryblidiacea)» (Ссылка на бесплатный полный текст + таблички) . Отчет Галатеи . 3 : 9–73.
- ^ Вингстранд, КГ (1985). «Об анатомии и взаимоотношениях современных моноплакофор» . Отчет Галатеи . 16 :7–94. Архивировано из оригинала (ссылка на бесплатный полный текст + таблички) 3 марта 2016 г. Проверено 23 марта 2009 г.
- ^ Jump up to: Перейти обратно: а б с Бойл, П.; Родхаус, П. (2005). «Происхождение и эволюция». Головоногие . п. 36. дои : 10.1002/9780470995310.ch3 . ISBN 9780470995310 .
- ^ Крёгер, БР (2007). «Некоторые малоизвестные особенности древнего отряда головоногих Ellesmerocerida (Nautiloidea, Cephalopoda)» . Палеонтология . 50 (3): 565–572. Бибкод : 2007Palgy..50..565K . дои : 10.1111/j.1475-4983.2007.00644.x .
- ^ Смит, Мартин Р.; Карон, Жан-Бернар (2010). «Примитивные мягкотелые головоногие моллюски кембрия». Природа . 465 (7297): 427–428. Бибкод : 2010Natur.465..427B . дои : 10.1038/465427a . ПМИД 20505713 . S2CID 205055896 .
- ^ Джайн, Шрипат (2016). Основы палеонтологии беспозвоночных: макрофоссилии . Спрингер. п. 73. ИСБН 978-81-322-3658-0 .
- ^ Jump up to: Перейти обратно: а б Крегер, Б.; Юн-бай, Ю.Б. (2009). «Импульсная диверсификация головоногих моллюсков в ордовике». Палеогеография, Палеоклиматология, Палеоэкология . 273 (1–2): 174–201. Бибкод : 2009PPP...273..174K . дои : 10.1016/j.palaeo.2008.12.015 .
- ^ Дзик, Дж. (1981). «Происхождение головоногих» (PDF) . Acta Palaeontologica Polonica . 26 (2): 161–191. Архивировано (PDF) из оригинала 16 декабря 2008 г.
- ^ Jump up to: Перейти обратно: а б с д Крёгер, БР; Серве, Т.; Чжан, Ю.; Косник, М. (2009). «Происхождение и начальный подъем пелагических головоногих моллюсков в ордовике» . ПЛОС ОДИН . 4 (9): е7262. Бибкод : 2009PLoSO...4.7262K . дои : 10.1371/journal.pone.0007262 . ПМЦ 2749442 . ПМИД 19789709 .
- ^ Jump up to: Перейти обратно: а б Голландия, Швейцария (1987). «Наутилоидные головоногие моллюски: странный успех: юбилейное послание президента 1986 года». Журнал Геологического общества . 144 (1): 1–15. Бибкод : 1987JGSoc.144....1H . дои : 10.1144/gsjgs.144.1.0001 . S2CID 128629737 .
- ^ Крёгер, Бьёрн (2006). «Ранние стадии роста и классификация головоногих ортоцеридан дарривилля (среднего ордовика) Балтоскандии». Летайя . 39 (2): 129–139. Бибкод : 2006Лета..39..129К . дои : 10.1080/00241160600623749 .
- ^ Jump up to: Перейти обратно: а б Янг, Р.Э.; Веккьоне, М.; Донован, Д.Т. (1998). «Эволюция колеоидных головоногих моллюсков и их современное биоразнообразие и экология». Южноафриканский журнал морских наук . 20 (1): 393–420. дои : 10.2989/025776198784126287 .
- ^ Jump up to: Перейти обратно: а б Танабэ, К. (2008). Головоногие моллюски – настоящее и прошлое . Токио: Издательство Университета Токай. [ нужна страница ]
- ^ Бэзил, Дженнифер; Бахтинова Ирина; Куроива, Кристина; Ли, Нанди; Мимс, Дезире; Прейс, Майкл; Сусье, Кристиан (1 сентября 2005 г.). «Функция ринофора и щупалец Nautilus pompilius L. (Cephalopoda, Nautiloidea) в ориентации на запах». Морское и пресноводное поведение и физиология . 38 (3): 209–221. Бибкод : 2005MFBP...38..209B . дои : 10.1080/10236240500310096 . S2CID 33835096 .
- ^ Сигено, Шуичи; Сасаки, Такенори; Моритаки, Такея; Касугай, Такаши; Веккьоне, Майкл; Агата, Киёкадзу (январь 2008 г.). «Эволюция комплекса головы головоногих моллюсков путем сборки нескольких частей тела моллюсков: данные эмбрионального развития наутилуса». Журнал морфологии . 269 (1): 1–17. дои : 10.1002/jmor.10564 . ПМИД 17654542 . S2CID 13109195 .
- ^ Jump up to: Перейти обратно: а б О'Брайен, Кейтлин Э.; Румбедакис, Катина; Винкельманн, Ингер Э. (6 июня 2018 г.). «Текущее состояние науки о головоногих и взгляды на наиболее важные предстоящие проблемы от трех молодых исследователей» . Границы в физиологии . 9 : 700. doi : 10.3389/fphys.2018.00700 . ISSN 1664-042X . ПМК 6014164 . ПМИД 29962956 .
- ^ Jump up to: Перейти обратно: а б с Альбертин, Кэролайн Б.; Симаков Олег; Митрос, Тереза; Ван, З. Ян; Пунгор, Юдит Р.; Эдсингер-Гонсалес, Эрик; Бреннер, Сидней; Рэгсдейл, Клифтон В.; Рохсар, Дэниел С. (август 2015 г.). «Геном осьминога и эволюция нейронных и морфологических новинок головоногих» . Природа . 524 (7564): 220–224. Бибкод : 2015Natur.524..220A . дои : 10.1038/nature14668 . ISSN 0028-0836 . ПМЦ 4795812 . ПМИД 26268193 .
- ^ Jump up to: Перейти обратно: а б Геринг, Мэри А. (4 февраля 2013 г.). «Экспрессия импринтированных генов и вклад мобильных элементов». Растительные транспозоны и динамика генома в эволюции . Уайли-Блэквелл. стр. 117–142. дои : 10.1002/9781118500156.ch7 . ISBN 978-1-118-50015-6 .
- ^ Эрвин, Дженнифер А.; Маркетто, Мария К.; Гейдж, Фред Х. (август 2014 г.). «Мобильные элементы ДНК в формировании разнообразия и сложности в мозге» . Обзоры природы Неврология . 15 (8): 497–506. дои : 10.1038/nrn3730 . ISSN 1471-003X . ПМЦ 4443810 . ПМИД 25005482 .
- ^ Альбертин, Кэролайн Б.; Медина-Руис, София; Митрос, Тереза; Шмидбаур, Ханна; Санчес, Густаво; Ван, З. Ян; Гримвуд, Джейн; Розенталь, Джошуа Дж. К.; Рэгсдейл, Клифтон В.; Симаков Олег; Рохсар, Дэниел С. (04 мая 2022 г.). «Механизмы генома и транскриптома, управляющие эволюцией головоногих моллюсков» . Природные коммуникации . 13 (1): 2427. Бибкод : 2022NatCo..13.2427A . дои : 10.1038/s41467-022-29748-w . hdl : 1912/29234 . ПМЦ 9068888 . ПМИД 35508532 .
- ^ Шмидбаур, Ханна; Кавагути, Аканэ; Кларенс, Тереза; Фу, Сяо; Хоанг, Ой Пуи; Циммерманн, Боб; Ритчард, Елена А.; Вайсенбахер, Антон; Фостер, Джейми С.; Нихолм, Спенсер В.; Бейтс, Пол А.; Альбертин, Кэролайн Б.; Танака, Элли; Симаков Олег (21 апреля 2022 г.). «Появление новой регуляции и экспрессии генов головоногих моллюсков посредством крупномасштабной реорганизации генома» . Природные коммуникации . 13 (1): 2172. Бибкод : 2022NatCo..13.2172S . дои : 10.1038/s41467-022-29694-7 . hdl : 1912/29187 . ПМЦ 9023564 . ПМИД 35449136 .
- ^ Уэлен, Кристофер Д.; Ландман, Нил Х. (08 марта 2022 г.). «Ископаемые колеоидные головоногие моллюски из Миссисипского Медвежьего ущелья Лагерштетте проливают свет на раннюю эволюцию вампиропод» . Природные коммуникации . 13 (1): 1107. Бибкод : 2022NatCo..13.1107W . дои : 10.1038/s41467-022-28333-5 . ISSN 2041-1723 . ПМЦ 8904582 . ПМИД 35260548 .
- ^ Страгнелл, Дж.; Норман, М.; Джексон, Дж.; Драммонд, А.; Купер, А. (2005). «Молекулярная филогения колеоидных головоногих моллюсков (Mollusca: Cephalopoda) с использованием мультигенного подхода; влияние разделения данных на разрешение филогений в байесовской системе». Молекулярная филогенетика и эволюция . 37 (2): 426–441. Бибкод : 2005МОЛПЭ..37..426С . дои : 10.1016/j.ympev.2005.03.020 . ПМИД 15935706 .
- ^ Страгнелл, Дж.; Джексон, Дж.; Драммонд, Эй Джей; Купер, А. (2006). «Оценки времени расхождения для основных групп головоногих моллюсков: данные из нескольких генов». Кладистика . 22 (1): 89–96. дои : 10.1111/j.1096-0031.2006.00086.x . ПМИД 34892892 . S2CID 84743000 .
- ^ Карлини, Д.Б.; Рис, Канзас; Грейвс, Дж. Э. (2000). «Эволюция семейства генов актина и филогения колеоидных головоногих моллюсков (Mollusca: Cephalopoda)» . Молекулярная биология и эволюция . 17 (9): 1353–1370. doi : 10.1093/oxfordjournals.molbev.a026419 . ПМИД 10958852 .
- ^ Бергманн, С.; Либ, Б.; Рут, П.; Маркл, Дж. (2006). «Гемоцианин из живого ископаемого, головоногих моллюсков Nautilus pompilius : структура белка, организация генов и эволюция». Журнал молекулярной эволюции . 62 (3): 362–374. Бибкод : 2006JMolE..62..362B . дои : 10.1007/s00239-005-0160-x . ПМИД 16501879 . S2CID 4389953 .
- ^ Батер, Ф.А. (1888b). «Профессор Блейк и рост панциря головоногих» . Анналы и журнал естественной истории . 6. 1 (6): 421–426. дои : 10.1080/00222938809460761 .
- ^ Шевырев, А.А. (2005). «Макросистема головоногих моллюсков: исторический обзор, современное состояние знаний и нерешенные проблемы: 1. Основные особенности и общая классификация головоногих моллюсков». Палеонтологический журнал . 39 (6): 606–614. Перевод из Палеонтологического журнала № 6, 2005, 33–42.
- ^ Шевырев, А.А. (2006). «Макросистема головоногих моллюсков; исторический обзор, современное состояние знаний и нерешенные проблемы; 2. Классификация наутилоидных головоногих моллюсков». Палеонтологический журнал . 40 (1): 46–54. Бибкод : 2006PalJ...40...46S . дои : 10.1134/S0031030106010059 . S2CID 84616115 .
- ^ Крогер Б. «Рецензия в российском «Палеонтологическом журнале» » . Архивировано из оригинала 31 августа 2009 г.
- ^ Батер, Ф.А. (1888a). «Рост панциря головоногих (Siphonopoda)» . Анналы и журнал естественной истории . 6. 1 (4): 298–310. дои : 10.1080/00222938809460727 .
- ^ Бертольд, Томас; Энгезер, Тео (1987). «Филогенетический анализ и систематизация головоногих моллюсков». Переговоры Ассоциации естественных наук в Гамбурге . 29 : 187-220.
- ^ Энгезер, Тео (1997). «Страница ископаемых наутилоидей» . Архивировано из оригинала 25 сентября 2006 г.
- ^ Линдгрен, Арканзас; Гирибет, Г.; Нисигути, МК (2004). «Комбинированный подход к филогении головоногих (Mollusca)». Кладистика . 20 (5): 454–486. CiteSeerX 10.1.1.693.2026 . дои : 10.1111/j.1096-0031.2004.00032.x . ПМИД 34892953 . S2CID 85975284 .
- ^ Хоган, К. Майкл (22 декабря 2007 г.). «Кносские полевые заметки» . Современный антиквар .
- ^ Уилк, Стивен Р. (2000). Медуза: Разгадка тайны Горгоны . Издательство Оксфордского университета. ISBN 978-0-19-988773-6 .
- ^ «Система природы Карла Линнея, содержащая три царства природы » . google.com
- ^ Смедли, Эдвард; Роуз, Хью Джеймс; Роуз, Генри Джон (1845). Энциклопедия Metropolitana, или Универсальный словарь знаний: сочетающий в себе двойное преимущество философского и алфавитного расположения, с соответствующими гравюрами . Б. Феллоуз. стр. 255–.
- ^ Диксон, Роланд Беррейдж (1916). Океанический . Мифология всех рас. Том. 9. Компания Маршалл Джонс. стр. 2–.
- ^ Бэтчелор, Джон (1901). Айны и их фольклор . Лондон: Общество религиозных трактатов.
- ^ Чисхолм, Хью , изд. (1911). . Британская энциклопедия (11-е изд.). Издательство Кембриджского университета.
- ^ Коэн-Врино, Жерар (2012). «Об осьминогах, или Анатомия женской власти». Различия . 23 (2): 32–61. дои : 10.1215/10407391-1533520 .
- ^ Фрице, Корд; Суойоки, Саара (2000). Запрещенные изображения: эротическое искусство японского периода Эдо (на финском языке). Городской художественный музей Хельсинки. п.п. 23–28. ISBN 978-951-8965-54-4 .
- ^ Уленбек, Крис; Маргарита Винкель; Эллис Тиниос; Эми Рейгл Ньюленд (2005). Японские эротические фантазии: сексуальные образы периода Эдо . Хотей. п. 161. ИСБН 978-90-74822-66-4 .
- ^ Бриль, Хольгер (2010). Бернингер, Марк; Экке, Йохен; Хаберкорн, Гидеон (ред.). Блуждающий взгляд встречается с фотографиями путешествий: поле зрения и глобальный рост популярности манги для взрослых . МакФарланд. п. 203. ИСБН 978-0-7864-3987-4 .
{{cite book}}:|work=игнорируется ( помогите ) - ^ Смит, С. (26 февраля 2010 г.). «Почему карикатура Марка Цукерберга на осьминога напоминает «нацистскую пропаганду», - извиняется немецкая газета» . iMediaЭтика . Проверено 31 мая 2017 г.
Дальнейшее чтение [ править ]
- Барсков И.С.; Бойко, М.С.; Коновалова В.А.; Леонова, Т.Б.; Николаева, СВ (2008). «Головоногие моллюски в морских экосистемах палеозоя». Палеонтологический журнал . 42 (11): 1167–1284. Бибкод : 2008PalJ...42.1167B . дои : 10.1134/S0031030108110014 . S2CID 83608661 . Комплексный обзор палеозойских головоногих моллюсков.
- Кэмпбелл, Нил А.; Рис, Джейн Б.; Митчелл, Лоуренс Г. (1999). Биология, пятое издание . Addison Wesley Longman, Inc. Менло-Парк, Калифорния: ISBN 978-0-8053-6566-5 .
- Фелли Дж., Веккьоне М., Ропер CFE, Суини М. и Кристенсен Т., 2001–2003: Текущая классификация современных головоногих моллюсков . Национальный музей естественной истории: Отдел систематической биологии: зоология беспозвоночных: головоногие моллюски.
- Хэнлон, Роджер; Веккьоне, Майк; Олкок, Луиза (2018). Осьминоги, кальмары и каракатицы: наглядное научное руководство по наиболее развитым беспозвоночным океана . Издательство Чикагского университета. ISBN 978-0226459561 .
- Н. Джоан Эбботт, Родди Уильямсон, Линда Мэддок. Нейробиология головоногих . Издательство Оксфордского университета, 1995. ISBN 0-19-854790-0
- Мэрион Никсон и Джон З. Янг . Мозг и жизнь головоногих . Издательство Оксфордского университета, 2003. ISBN 0-19-852761-6
- Хэнлон, Роджер Т. и Джон Б. Мессенджер. Поведение головоногих . Издательство Кембриджского университета, 1996. ISBN 0-521-42083-0
- Мартин Стивенс и Сами Мерайлайта. Камуфляж животных: механизмы и функции . Издательство Кембриджского университета, 2011. ISBN 0-521-19911-5
- Родхаус, П.Г.; Нигматуллин, Ч. М. (1996). «Роль потребителей». Философские труды Королевского общества B: Биологические науки . 351 (1343): 1003–1022. дои : 10.1098/rstb.1996.0090 .
- Классификационный ключ к современным головоногим моллюскам: ftp://ftp.fao.org/docrep/fao/009/a0150e/a0150e03.pdf. [ постоянная мертвая ссылка ]
Внешние ссылки [ править ]
Головоногие .
- Рыбы против головоногих
- TONMO.COM - Интернет-журнал Octopus News Magazine - статьи и обсуждения о головоногих моллюсках.
- Scientific American: Может ли кальмар вылететь из воды?
- Семинар Роджера Хэнлона: «Быстрая адаптивная маскировка и передача сигналов у головоногих моллюсков»
- Глубоководные щетинистые черви, обитающие в море