Обыкновенный осьминог
| Обыкновенный осьминог Временной диапазон: | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Моллюска |
| Сорт: | Головоногие моллюски |
| Заказ: | Осьминог |
| Семья: | Октоподиды |
| Род: | Осьминог |
| Разновидность: | О. обыкновенный |
| Биномиальное имя | |
| Осьминог обыкновенный Кювье , 1797 г. | |
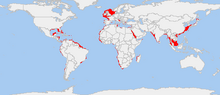 | |
| Распространение обыкновенного осьминога по всему миру | |
| Синонимы | |
| |

Осьминог обыкновенный ( Octopus vulgaris ) — моллюск, принадлежащий к классу головоногих . Octopus vulgaris — один из наиболее изученных видов осьминогов , а также один из самых умных. Он простирается от восточной Атлантики, простирается от Средиземного моря и южного побережья Англии до южного побережья Южной Африки. Встречается также у Азорских , Канарских и Кабо-Верде островов . Вид также распространен в Западной Атлантике.
Характеристики
[ редактировать ]





Octopus vulgaris вырастает до 25 см (10 дюймов) в длину мантии и имеет руки до 1 м (3,3 фута) в длину. [3] Он живет 1–2 года и может весить до 9 кг (20 фунтов). [4] [5] Спаривание может стать каннибалистическим . [6] O. vulgaris в огромных масштабах вылавливается донными тралами у северо-западного побережья Африки. Ежегодно собирают более 20 000 тонн (22 000 коротких тонн). [3]
Обыкновенный осьминог охотится в сумерках. Предпочтительны крабы, раки и двустворчатые моллюски (например, моллюски), хотя осьминог ест почти все, что может поймать. Он способен менять цвет, чтобы сливаться с окружающей средой, и способен прыгнуть на любую неосторожную добычу, оказавшуюся на его пути. Используя свой клюв, он способен разламывать раковины очищенных моллюсков. Он также обладает ядом, позволяющим подчинить себе добычу. [7]
В ходе эволюции у них появилась крупная нервная система и мозг. В организме человека насчитывается около 500 миллионов нейронов, что почти сравнимо с собаками. Они достаточно умны, чтобы различать яркость, перемещаться по лабиринтам, узнавать отдельных людей, учиться отвинчивать банки или совершать набеги на ловушки для омаров. [8] [9] [10] Также было замечено, что они содержат «сады», в которых собирают различные морские растения и водоросли, а также коллекции ракушек и камней; такое поведение, возможно, послужило вдохновением для названия группы «Битлз» 1969 года « Сад осьминога ». O. vulgaris был первым беспозвоночным животным, защищенным Законом о животных (научные процедуры) 1986 года в Великобритании. [11] Дрессирующие эксперименты показали, что осьминог обыкновенный может различать яркость, размер, форму, а также горизонтальную или вертикальную ориентацию объектов.
Физиология
[ редактировать ]Среда обитания и требования
[ редактировать ]Обыкновенный осьминог широко распространен в тропических, субтропических и умеренных водах по всему миру. [12] [13] [14] Они предпочитают дно относительно мелких, каменистых прибрежных вод, часто не глубже 200 м (660 футов). [14] Хотя они предпочитают около 36 граммов на литр (0,0013 фунта на кубический дюйм), соленость во всей их глобальной среде обитания составляет примерно от 30 до 45 граммов на литр (0,0011 и 0,0016 фунта на кубический дюйм). [15] Они подвергаются воздействию самых разных температур окружающей среды, но их предпочтительная температура колеблется от 15 до 16 ° C (от 59 до 61 ° F). [15] В особенно теплые сезоны осьминога часто можно встретить глубже обычного, чтобы избежать более теплых слоев воды. [16] Двигаясь вертикально по воде, осьминог подвергается воздействию различных давлений и температур, которые влияют на концентрацию кислорода, доступного в воде. [15] Это можно понять с помощью закона Генри , который гласит, что концентрация газа в веществе пропорциональна давлению и растворимости, на которую влияет температура. Эти различные различия в доступности кислорода предъявляют требования к методам регулирования. [17]
В первую очередь осьминог располагается в укрытии, где минимальная часть его тела представлена внешней воде. [18] Когда он действительно движется, то большую часть времени он движется по дну океана или моря, и в этом случае нижняя часть осьминога все еще не видна. [18] Такое ползание значительно увеличивает метаболические потребности, требуя от них увеличения потребления кислорода примерно в 2,4 раза по сравнению с количеством, необходимым для отдыхающего осьминога. [19] Эта повышенная потребность удовлетворяется увеличением ударного объема сердца осьминога. [20]
Осьминог иногда плавает по воде, полностью обнажая себя. [15] При этом он использует реактивный механизм, который предполагает создание гораздо более высокого давления в его мантийной полости , что позволяет ему двигаться сквозь воду. [20] Поскольку сердце и жабры обыкновенного осьминога расположены внутри его мантии, это высокое давление также сужает и накладывает ограничения на различные сосуды, возвращающие кровь к сердцу. [20] В конечном итоге это создает проблемы с кровообращением и не является устойчивым видом транспорта, поскольку осьминог не может обеспечить потребление кислорода, которое могло бы сбалансировать метаболические потребности при максимальных нагрузках. [20]
Дыхание
[ редактировать ]осьминог использует жабры В качестве дыхательной поверхности . Жабры состоят из жаберных ганглиев и ряда складчатых пластинок . Первичные ламели вытягиваются, образуя полуветви, а затем сгибаются, образуя вторичные свободно сложенные ламели, которые прикреплены только вверху и внизу. [21] Третичные ламели образуются путем сгибания вторичных ламелей веерообразной формы. [21] Вода медленно движется в одном направлении по жабрам и пластинкам, в мантийную полость и из воронки осьминога. [22]
Структура жабр осьминога обеспечивает поглощение большого количества кислорода; до 65% в воде при 20 °C (68 °F). [22] на тонкую кожу осьминога приходится большая часть поглощения кислорода В исследовании in vitro ; По оценкам, около 41% всего поглощения кислорода в состоянии покоя происходит через кожу. [18] На это число влияет активность животного – поглощение кислорода увеличивается, когда осьминог тренируется, поскольку все его тело постоянно подвергается воздействию воды, но общее количество поглощения кислорода через кожу фактически снижается до 33% в результате метаболические издержки плавания. [18] Когда животное сворачивается калачиком после еды, поглощение кислорода через кожу может упасть до 3% от общего потребления кислорода. [18] Дыхательный пигмент осьминога, гемоцианин , также способствует увеличению поглощения кислорода. [17] Осьминоги могут поддерживать постоянное потребление кислорода, даже когда концентрация кислорода в воде снижается примерно до 3,5 кПа (0,51 фунта на квадратный дюйм). [22] или насыщение 31,6% (стандартное отклонение 8,3%). [17] Если насыщение кислородом морской воды упадет примерно до 1–10%, это может быть смертельным для Octopus vulgaris в зависимости от веса животного и температуры воды. [17] Вентиляция может усилиться, чтобы перекачивать больше воды, переносящей кислород через жабры, но благодаря рецепторам, расположенным на жабрах, потребление энергии и поглощение кислорода остаются на стабильном уровне. [22] Высокий процент извлечения кислорода позволяет экономить энергию и дает преимущества при проживании в зоне с низкой концентрацией кислорода. [21]
Вода закачивается в мантийную полость осьминога, где соприкасается с внутренними жабрами. Вода имеет более высокую концентрацию кислорода по сравнению с кровью, возвращающейся из вен, поэтому кислород диффундирует в кровь. Ткани и мышцы осьминога используют кислород и выделяют углекислый газ при расщеплении глюкозы в цикле Кребса . Затем углекислый газ растворяется в крови или соединяется с водой с образованием углекислоты , что снижает pH крови. Эффект Бора объясняет, почему концентрация кислорода в венозной крови ниже, чем в артериальной, и почему кислород диффундирует в кровоток. На скорость диффузии влияет расстояние, которое кислород должен пройти из воды в кровоток, как указано в законах диффузии Фика . Законы Фика объясняют, почему жабры осьминогов содержат множество мелких складок с высокой васкуляризацией. Они увеличивают площадь поверхности, тем самым также увеличивая скорость диффузии . Капилляры, выстилающие складки жаберного эпителия. имеют очень тонкий тканевый барьер (10 мкм), что обеспечивает быструю и легкую диффузию кислорода в кровь. [23] В ситуациях, когда парциальное давление кислорода в воде низкое, диффузия кислорода в кровь снижается. [24] Закон Генри может объяснить это явление. Закон гласит, что в состоянии равновесия парциальное давление кислорода в воде будет равно парциальному давлению кислорода в воздухе; но концентрации будут различаться из-за разной растворимости. Этот закон объясняет, почему O. vulgaris должен изменять количество воды, проходящей через мантийную полость, при изменении концентрации кислорода в воде. [22]
Жабры находятся в прямом контакте с водой, переносящей больше кислорода, чем кровь, которая попала в мантийную полость осьминога. Жаберные капилляры довольно малы и многочисленны, что увеличивает площадь поверхности, с которой может контактировать вода, что приводит к усилению диффузии кислорода в кровь. Некоторые данные указывают на то, что пластинки и сосуды внутри пластинок на жабрах сжимаются, помогая продвижению крови через капилляры. [25]
Тираж
[ редактировать ]У осьминога три сердца: одно главное двухкамерное сердце, отвечающее за доставку насыщенной кислородом крови в организм, и два жаберных сердца меньшего размера, по одному рядом с каждой парой жабр. Кровеносный контур отправляет насыщенную кислородом кровь из жабр в предсердие большого сердца, а затем в его желудочек, который перекачивает эту кровь в остальную часть тела. Дезоксигенированная кровь из организма поступает в жаберные сердца, которые перекачивают кровь через жабры, чтобы насытить ее кислородом, а затем кровь возвращается в системное предсердие, чтобы процесс начался снова. [26] От системного сердца отходят три аорты: две второстепенные (брюшная аорта и гонадная аорта) и одна большая, дорсальная аорта, которая обслуживает большую часть тела. [27] У осьминога также есть большие кровеносные пазухи вокруг кишечника и за глазами, которые служат резервами во время физиологического стресса. [28]
Частота сердечных сокращений осьминога существенно не меняется при физической нагрузке, хотя временная остановка сердца в системном сердце может быть вызвана недостатком кислорода, практически любым внезапным раздражителем или давлением в мантии во время реактивного движения. [29] Единственной компенсацией нагрузки является увеличение ударного объема сердца до трех раз. [29] это означает, что он испытывает кислородный дефицит практически при любом быстром движении. [29] [30] Однако осьминог способен контролировать, сколько кислорода он вытягивает из воды при каждом вдохе, используя рецепторы на своих жабрах. [22] позволяя ему поддерживать постоянное поглощение кислорода в диапазоне давлений кислорода в окружающей воде. [29] Три сердца также зависят от температуры и кислорода, и ритм сокращений трех сердец обычно находится в фазе: два жаберных сердца бьются вместе, за которыми следует системное сердце. [26] Закон Франка-Старлинга также способствует общей функции сердца через сократимость и ударный объем, поскольку общий объем кровеносных сосудов должен поддерживаться и оставаться относительно постоянным внутри системы, чтобы сердце функционировало правильно. [31]
Кровь осьминогов состоит из богатого медью гемоцианина, который менее эффективен, чем богатый железом гемоглобин позвоночных, поэтому не увеличивает сродство к кислороду в такой же степени. [32] Оксигенированный гемоцианин в артериях связывается с CO 2 , который затем высвобождается, когда кровь в венах дезоксигенируется. Выброс CO 2 в кровь вызывает ее закисление с образованием угольной кислоты. [33] Эффект Бора объясняет, что концентрация углекислого газа влияет на pH крови и выделение или потребление кислорода. Цикл Кребса использует кислород из крови для расщепления глюкозы в активных тканях или мышцах и высвобождает углекислый газ в качестве ненужного продукта, что приводит к выделению большего количества кислорода. Кислород, попадающий в ткани или мышцы, создает дезоксигенированную кровь, которая возвращается к жабрам вен. Два плечевых сердца осьминога перекачивают кровь из вен через жаберные капилляры. Вновь насыщенная кислородом кровь оттекает из жаберных капилляров в сердце, откуда затем разносится обратно по всему телу. [26]
Объем крови в организме осьминога составляет около 3,5% от массы его тела. [28] но способность крови переносить кислород составляет всего около 4 объемных процентов. [29] Это способствует их восприимчивости к кислородному долгу, упомянутому ранее. Шедвик и Нильссон [30] пришел к выводу, что кровеносная система осьминога «принципиально непригодна для высокой физиологической работоспособности». Поскольку связывающий агент находится в плазме, а не в клетках крови, существует ограничение на поглощение кислорода, которое может испытывать осьминог. Если бы в кровотоке увеличилось содержание гемоцианина, жидкость стала бы слишком вязкой для миогенных клеток. [34] сердца качать. [31] Закон Пуазейля объясняет скорость потока основной жидкости по всей системе кровообращения за счет разницы кровяного давления и сосудистого сопротивления. [31]
Как и у позвоночных, кровеносные сосуды осьминогов очень эластичны, их устойчивость к физиологическому давлению составляет 70%. В основном они состоят из эластичного волокна, называемого артериальным эластомером осьминога, с более жесткими коллагеновыми волокнами, которые рекрутируются под высоким давлением, чтобы помочь сосуду сохранять свою форму без чрезмерного растяжения. [35] Шедвик и Нильссон [30] предположил, что все кровеносные сосуды осьминогов могут использовать сокращения гладких мышц, чтобы способствовать передвижению крови по телу, что имело бы смысл в контексте их жизни под водой с сопутствующим давлением.
Эластичность и сократительная природа аорты осьминога служат для сглаживания пульсирующего характера кровотока от сердца, когда импульсы проходят по длине сосуда, в то время как полая вена служит хранилищем энергии. [30] Ударный объём системного сердца изменяется обратно пропорционально разнице между входным давлением крови через полую вену и выходным противодавлением через аорту .
Осморегуляция
[ редактировать ]
Гемолимфа головоногих моллюсков, в том , перикардиальная жидкость и моча числе обыкновенного осьминога, изоосмотичны друг другу, а также окружающей морской воде. [36] Было высказано предположение, что головоногие моллюски не осуществляют осморегуляции , что указывает на то, что они являются конформерами. [36] Это означает, что они адаптируются, чтобы соответствовать осмотическому давлению окружающей среды, а поскольку осмотического градиента нет, нет чистого движения воды из организма в морскую воду или из морской воды в организм. [36] Среднее минимальное требование к солености осьминогов составляет 27 г/л (0,00098 фунта/куб. дюйм), и любое нарушение, приводящее к попаданию значительного количества пресной воды в их среду, может оказаться фатальным. [37]
Однако, что касается ионов , то, по-видимому, действительно существует несоответствие между концентрациями ионов, обнаруженными в морской воде, и концентрациями, обнаруженными в головоногих моллюсках. [36] В целом, в отличие от соленой воды, в них сохраняется гипоионная концентрация натрия, кальция и хлоридов. [36] Сульфат и калий также существуют в гипоионном состоянии, за исключением выделительной системы головоногих моллюсков, где моча гиперионна. [36] Эти ионы могут свободно диффундировать, и поскольку они существуют в организме в гипоионных концентрациях, они будут поступать в организм из морской воды. [36] Тот факт, что организм может поддерживать гипоионные концентрации, предполагает не только то, что у головоногих существует форма ионной регуляции, но также и то, что они также активно выделяют определенные ионы, такие как калий и сульфат, для поддержания гомеостаза . [36]
O. vulgaris систему моллюскового типа имеет почечную , которая сильно отличается от таковой у млекопитающих. Система построена вокруг придатка каждой жаберной части сердца, который по сути является продолжением перикарда . [36] Эти длинные ресничные протоки фильтруют кровь в пару почечных мешочков, одновременно активно реабсорбируя глюкозу и аминокислоты в кровоток. [36] Почечные мешочки активно регулируют концентрацию ионов в моче и активно добавляют в мочу азотистые соединения и другие продукты метаболизма. [36] После завершения фильтрации и реабсорбции моча опорожняется в мантийную полость O. vulgaris через пару почечных сосочков, по одному из каждого почечного мешка. [36]
Температура и размер тела напрямую влияют на потребление кислорода O. vulgaris , что изменяет скорость метаболизма. [19] Когда потребление кислорода уменьшается, количество выделения аммиака также уменьшается из-за замедления скорости обмена веществ. [19] В организме O. vulgaris находятся четыре различные жидкости: кровь, перикардиальная жидкость, моча и почечная жидкость. Моча и почечная жидкость имеют высокие концентрации калия и сульфатов, но низкие концентрации хлоридов. В моче низкая концентрация кальция, что позволяет предположить, что он активно выводится. Почечная жидкость имеет такую же концентрацию кальция, как и кровь. Концентрация хлоридов в крови высока, а концентрация натрия варьируется. Перикардиальная жидкость имеет концентрации натрия, калия, хлора и кальция, аналогичные концентрациям в соленой воде, что подтверждает идею о том, что O. vulgaris не осморегулирует, а подчиняется. Однако он имеет более низкую концентрацию сульфатов. [36] Перикардиальный проток содержит ультрафильтрат крови, известный как перикардиальная жидкость, и скорость фильтрации частично контролируется богатыми мышцами и нервами жаберными сердцами. [36] Почечные придатки перемещают азотистые и другие продукты жизнедеятельности из крови в почечные мешочки, но не увеличивают объем. Почечная жидкость имеет более высокую концентрацию аммиака, чем моча или кровь, поэтому почечные мешочки остаются кислыми, что помогает выводить аммиак из почечных придатков. Аммиак диффундирует по градиенту концентрации в мочу или кровь, где он прокачивается через жаберные сердца и диффундирует через жабры. [36] Выделение аммиака O. vulgaris делает их аммонотелическими организмами. выделяет несколько других азотистых отходов, Помимо аммиака, O. vulgaris таких как мочевина , мочевая кислота , пурины и некоторые свободные аминокислоты, но в меньших количествах. [36]
В почечных мешочках за регуляцию ионов отвечают две признанные специфические клетки. Два типа клеток — это клетки, образующие лакуны, и эпителиальные клетки , типичные для почечных канальцев. Клетки эпителия реснитчатые , цилиндрические и поляризованные , с тремя отдельными участками. Эти три области — апикальная, средняя цитоплазматическая и базальная пластинка. Средняя цитоплазматическая область является наиболее активной из трех из-за концентрации внутри нее множества органелл, таких как митохондрии, гладкая и шероховатая эндоплазматическая сеть и другие. Увеличение активности происходит за счет взаимосвязанного лабиринта базальной пластинки, создающего перекрестную активность, аналогичную активности богатых митохондриями клеток, обнаруженных у костистых морских рыб. Лакунообразующие клетки характеризуются контактом с базальной пластинкой, но не достигают апикального края ассоциированных эпителиальных клеток и располагаются в жаберном эпителии сердца. Форма широко варьируется и иногда бывает более электронно-плотной, чем эпителиальные клетки, что рассматривается как «диффузная почка», регулирующая концентрации ионов. [38]
Одна из адаптаций O. vulgaris — это некоторый прямой контроль над почками. [36] Он может по желанию переключаться между правой и левой почкой, выполняющей основную часть фильтрации, а также может регулировать скорость фильтрации, чтобы скорость не увеличивалась, когда кровяное давление животного повышается из-за стресса или физических упражнений. [36] Некоторые виды осьминогов, в том числе O. vulgaris , также имеют проток, идущий из гонадного пространства в жаберный перикард. [36] Уэллс [36] предположили, что этот проток, который сильно васкуляризирован и иннервирован, может способствовать реабсорбции важных метаболитов из яйцевой жидкости беременных женщин, направляя эту жидкость в почечные придатки.
Терморегуляция
[ редактировать ]Как океанический организм, O. vulgaris испытывает колебания температуры из-за многих факторов, таких как время года, географическое положение и глубина. [39] Например, осьминоги, живущие в окрестностях Неаполя , могут испытывать температуру 25 °C (77 °F) летом и 15 °C (59 °F) зимой. [39] Однако эти изменения будут происходить довольно постепенно и, следовательно, не потребуют какого-либо жесткого регулирования.
Обыкновенный осьминог — пойкилотермный , эвритермный эктотермный организм , что означает, что он соответствует температуре окружающей среды. [40] Это означает, что между организмом и окружающей средой не наблюдается реального температурного градиента, и они быстро выравниваются. Если осьминог плывет в более теплое место, он получает тепло от окружающей воды, а если он плывет в более холодное место, он теряет тепло аналогичным образом.
O. vulgaris может применять изменения в поведении, чтобы управлять широким диапазоном температур окружающей среды. Частота дыхания осьминогов чувствительна к температуре — дыхание увеличивается с температурой. [41] Его потребление кислорода увеличивается при температуре воды от 16 до 28 ° C (от 61 до 82 ° F), достигает максимума при 28 ° C (82 ° F), а затем начинает падать при 32 ° C (90 ° F). [41] Оптимальная температура для метаболизма и потребления кислорода составляет от 18 до 24 °C (от 64 до 75 °F). [41] Изменения температуры также могут вызывать изменение уровня белка гемолимфы в зависимости от потребления кислорода. [41] По мере повышения температуры концентрация белка увеличивается, чтобы приспособиться к температуре. Также кооперативность гемоцианина увеличивается, но сродство снижается. [42] И наоборот, снижение температуры приводит к снижению кооперативности дыхательных пигментов и увеличению сродства. [42] Небольшое повышение P50 , происходящее при изменении температуры, позволяет давлению кислорода оставаться высоким в капиллярах , что обеспечивает повышенную диффузию кислорода в митохондрии в периоды высокого потребления кислорода. [42] Повышение температуры приводит к повышению активности ферментов , однако снижение сродства к гемоцианину позволяет активности ферментов оставаться постоянной и поддерживать гомеостаз. Самые высокие концентрации белка гемолимфы наблюдаются при температуре 32 ° C (90 ° F), а затем падают при температурах выше этой. [41] Сродство к кислороду в крови снижается на 0,20 кПа/°C (0,016 фунтов на квадратный дюйм/°F) при pH 7,4. [42] Термическая устойчивость осьминога ограничена его способностью потреблять кислород, и когда он не может обеспечить достаточное количество кислорода для циркуляции при экстремальных температурах, последствия могут быть фатальными. [41] O. vulgaris обладает независимым от pH венозным резервом, который представляет собой количество кислорода, которое остается связанным с дыхательным пигментом при постоянном давлении кислорода. Этот резерв позволяет осьминогам переносить широкий диапазон pH в зависимости от температуры. [42]
Являясь конформером температуры, [43] У O. vulgaris нет какого-либо специального органа или структуры, предназначенной для производства тепла или теплообмена. Как и все животные, они выделяют тепло в результате обычных метаболических процессов, таких как переваривание пищи, [39] но не принимайте специальных средств для поддержания температуры тела в определенном диапазоне. Их предпочтительная температура напрямую отражает температуру, к которой они акклиматизировались. [43] Приемлемый диапазон температуры окружающей среды составляет 13–28 °C (55–82 °F). [43] оптимальная температура для максимальной метаболической эффективности составляет около 20 ° C (68 ° F). [40]
Обыкновенные осьминоги, как эктотермные животные, сильно подвержены влиянию изменений температуры. У всех видов есть температурные предпочтения, при которых они могут функционировать на своем основном уровне метаболизма . [43] Низкая скорость метаболизма обеспечивает быстрый рост, поэтому эти головоногие спариваются, когда вода становится ближе к предпочтительной зоне. Повышение температуры вызывает увеличение потребления кислорода O. vulgaris . [19] Повышенное потребление кислорода может быть напрямую связано со скоростью метаболизма, поскольку распад таких молекул, как глюкоза, требует поступления кислорода, что объясняется циклом Кребса. Количество выделяемого аммиака, наоборот, уменьшается с повышением температуры. [19] Уменьшение выделения аммиака также связано с обменом веществ осьминога из-за его необходимости тратить больше энергии при повышении температуры. Octopus vulgaris уменьшит количество выделяемого аммиака, чтобы использовать излишки растворенных веществ, которые в противном случае были бы выделены из-за увеличения скорости метаболизма. Осьминоги не регулируют свою внутреннюю температуру до тех пор, пока она не достигнет порога, когда они должны начать регулировать ее, чтобы предотвратить смерть. [19] Увеличение скорости метаболизма, наблюдаемое с повышением температуры, вероятно, связано с тем, что осьминог плавает на меньшую или большую глубину, чтобы оставаться в пределах своей предпочтительной температурной зоны.
Воспроизведение
[ редактировать ]Нерест O. vulgaris в этом районе продолжается с декабря по сентябрь с уникальным пиком в весенние месяцы. [44]
См. также
[ редактировать ]- Мой учитель-осьминог (документальный фильм, 2020 г.)
Ссылки
[ редактировать ]- ^ Кинтейро, Хавьер; Родригес-Кастро, Хорхе; Кинг-Мендес, Мануэль; Гонсалес-Энрикес, Сноу (19 марта 2020 г.). «Филогеография островных популяций обыкновенного осьминога Octopus vulgaris Cuvier, 1797 в Атлантической Макаронезии» . ПЛОС ОДИН 15 (3): e0230294. Бибкод : 2020PLoSO..1530294Q дои : 10.1371/journal.pone.0230194 . ПМК 7082011 . ПМИД 32191765 .
- ^ Олкок, Л.; Хедлам, Дж.; Аллен, Г. (2018). « Осьминог обыкновенный » . Красный список исчезающих видов МСОП . 2018 : e.T162571A918906. doi : 10.2305/IUCN.UK.2018-2.RLTS.T162571A918906.en . Проверено 18 марта 2022 г.
- ^ Перейти обратно: а б Норман, доктор медицинских наук, 2000. Головоногие моллюски: Всемирный справочник . ConchBooks.
- ^ «Обыкновенный осьминог» . Животные . Нэшнл Географик . 10 мая 2011 г. Архивировано из оригинала 17 октября 2016 г.
- ^ Пералес-Райя, Каталина; Альманса, Эдуардо; Бартоломе, Аврора; Фелипе, Беатрис К.; Иглесиас, Хосе; Санчес, Франсиско Хавьер; Карраско, Хосе Франсиско; Родригес, Кармен (2014). «Подтверждение возраста клювов Octopus vulgaris во всем онтогенетическом диапазоне: клювы как регистраторы жизненных событий у осьминогов» . Журнал исследований моллюсков . 33 (2): 481–493. дои : 10.2983/035.033.0217 . hdl : 10508/9595 . ISSN 0730-8000 . S2CID 85772903 .
- ^ Эрнандес-Урсера, Хорхе; Гарси, Мануэль Э.; Роура, Альваро; Гонсалес, Анхель Ф.; Кабанельяс-Реборедо, Мигель; Моралес-Нин, Беатрис; Герра, Анхель (ноябрь 2014 г.). «Каннибалистическое поведение осьминога (Octopus vulgaris) в дикой природе». Журнал сравнительной психологии . 128 (4): 427–430. дои : 10.1037/a0036883 . hdl : 10261/123375 . ПМИД 25198542 .
- ^ Канда, Ацухиро; Ивакоси-Укена, Эйко; Такува-Курода, Кёко; Минаката, Хироюки (январь 2003 г.). «Выделение и характеристика новых тахикининов из задней слюнной железы обыкновенного осьминога Octopus vulgaris». Пептиды . 24 (1): 35–43. дои : 10.1016/s0196-9781(02)00274-7 . ПМИД 12576083 . S2CID 21487225 .
- ^ «Осьминог открывает банку» . Ютуб. 16 сентября 2010 г. Архивировано из оригинала 15 декабря 2021 г. Проверено 5 февраля 2021 г.
- ^ «Осьминог в ловушке для креветок» . Ютуб. 12 февраля 2007 г. Архивировано из оригинала 15 декабря 2021 г. . Проверено 24 апреля 2019 г.
- ^ Годфри-Смит, Питер (2016). Другие умы: осьминог, море и глубокие истоки сознания . Нью-Йорк: Фаррар, Штраус и Жиру.
- ^ «Закон о животных (научные процедуры) (поправка) 1993 года» . 23 августа 1993 года . Проверено 22 февраля 2013 г.
- ^ «Рыболовство и аквакультура ФАО – Водные виды» .
- ^ «Осьминог обыкновенный» . Сеть разнообразия животных .
- ^ Перейти обратно: а б Белкари П., Кукку Д., Гонсалес М., Сраири А. и Видорис П. (2002) Распространение и численность Octopus vulgaris Cuvier 1797 (Cepphalopoda: Octopoda) в Средиземном море. Морские науки, 66 (S2): 157–166. два : 10.3989/scimar.2002.66s2157 .
- ^ Перейти обратно: а б с д Морено А., Лоуренсу С., Перейра Ж., Гаспар МБ, Кабрал Х.Н., Пирс Г.Дж. и др. (2013). Основные привычки для предварительного набора Octopus vulgaris на португальском побережье. Исследования рыболовства, 152 : 74–85. doi : 10.1016/j.fishres.2013.08.005 .
- ^ Кацаневакис, С. и Верриопулос, Г. (2004). Обилие Octopus vulgaris на мягких отложениях. Сциентия Марина, 68, 553–560. два : 10.3989/scimar.2004.68n4553 .
- ^ Перейти обратно: а б с д Вальверде, Хесус К. и Гарсия, Бенджамин. (2005). Подходящие уровни растворенного кислорода для осьминога обыкновенного ( Octopus vulgaris cuvier, 1797) при разном весе и температуре: анализ респираторного поведения. Аквакультура. 244: 303–314. doi : 10.1016/j.aquacultural.2004.09.036 .
- ^ Перейти обратно: а б с д и Мадан, Джей-Джей и Уэллс, MJ (1996). Кожное дыхание Octopus vulgaris . Журнал экспериментальной биологии, 199 : 2477–2483.
- ^ Перейти обратно: а б с д и ж Кацаневакис С., Стефанопулу С., Милиу Х., Мораиту-Апостолопулу М. и Верриопулос Г. (2005). Потребление кислорода и выделение аммиака Octopus vulgaris (Cephalopoda) в зависимости от массы тела и температуры. Морская биология, 146, 725–732. два : 10.1007/s00227-004-1473-9 .
- ^ Перейти обратно: а б с д Уэллс, М.Дж., Дати, Г.Г., Хулихан, Д.Ф., Смит, П.Дж.С. и Уэллс, Дж. (1987). Изменения кровотока и давления у тренирующихся осьминогов (Octopus vulgaris). Журнал экспериментальной биологии, 131, 175–187.
- ^ Перейти обратно: а б с Янг, Ричард Э. и Веккьоне, Майкл. (2002). Эволюция жабр у восьминогих. Вестник морской науки. 71(2): 1003–1017
- ^ Перейти обратно: а б с д и ж Уэллс, М.Дж., и Уэллс, Дж. (1995). Контроль респираторной и сердечной реакции на изменения напряжения кислорода в окружающей среде и потребности в кислороде у осьминога. Журнал экспериментальной биологии, 198, 1717–1727 гг.
- ^ Ино, Северная Каролина (август 1994 г.). Морфометрия жабр головоногих моллюсков. Журнал Морской биологической ассоциации Соединенного Королевства, 74 (3), 687–706.
- ^ Мельцнер Ф., Блок К. и Пертнер Х.О. (2006). Температурно-зависимая экстракция кислорода из вентиляционного потока и затраты на вентиляцию у головоногих Sepia officinalis. Журнал сравнительной физиологии B, 176, 607–621.
- ^ Уэллс, MJ, и Смит, PJS (1987). Производительность кровеносной системы осьминога: триумф инженерии над дизайном. Экспериментиа, 43, 487–499.
- ^ Перейти обратно: а б с Уэллс, MJ (1980). Нервный контроль сердцебиения у осьминога . Журнал экспериментальной биологии, 85, 111–128. ПМИД 7373208 .
- ^ Смит, PJS (1981). Роль венозного давления в регуляции выброса из сердца осьминога Eledone cirrosa (Lam.) . Журнал экспериментальной биологии, 93, 243–255.
- ^ Перейти обратно: а б О'Дор, Р.К., и Уэллс, М.Дж. (1984). Время кровообращения, запасы крови и внеклеточное пространство головоногих моллюсков. Журнал экспериментальной биологии, 113, 461–464. hdl : 10222/29342 .
- ^ Перейти обратно: а б с д и Уэллс, MJ (1979). Сердцебиение Octopus vulgaris . Журнал экспериментальной биологии, 78, 87–104.
- ^ Перейти обратно: а б с д Шедвик, Р.Э. и Нильссон ЕК. (1990). Значение сосудистой эластичности в системе кровообращения головоногих Octopus vulgaris . Журнал экспериментальной биологии, 152, 471–484.
- ^ Перейти обратно: а б с Хилл, Ричард В., Гордон А. Вайс и Маргарет Андерсон. Физиология животных . 3-е изд. Сандерленд, Массачусетс: (635–636, 654–657, 671–672) Sinauer Associates, 2012. Печать
- ^ «Информационный бюллетень об осьминоге» (PDF) . Всемирный фонд животных.
- ^ Пертнер, Х.О. (январь 1995 г.). «Координация обмена веществ, кислотно-основная регуляция и функция гемоцианина у головоногих моллюсков» (PDF) . Морское и пресноводное поведение и физиология . 25 (1–3): 131–148. Бибкод : 1995MFBP...25..131P . дои : 10.1080/10236249409378913 .
- ^ Связь возбуждения-сокращения
- ^ Шедвик, Роберт Э. и Гослайн, Джон М. (1985). Механические свойства аорты осьминога . Журнал экспериментальной биологии, 114, 259–284.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Уэллс, MJ (1978). Осьминог: физиология и поведение продвинутых беспозвоночных . Кембридж: Университетская типография.
- ^ Ваз-Пирес, Пауло; Сейшас, Педро; Барбоза, Александра (сентябрь 2004 г.). «Аквакультурный потенциал осьминога обыкновенного (Octopus vulgaris Cuvier, 1797): обзор». Аквакультура . 238 (1–4): 221–238. Бибкод : 2004Aquac.238..221V . doi : 10.1016/j.aquacultural.2004.05.018 .
- ^ Витмер, А. (1975). Тонкое строение реноперикардиальной полости головоногих моллюсков ocotopus dofleine martini. Журнал исследований ультраструктуры, (53), 29–36.
- ^ Перейти обратно: а б с Хилл, Р.В., Вайс, Джорджия, и Андерсон, М. (2012). Физиология животных. Сандерленд: SinauerAssociates, стр. 164–165.
- ^ Перейти обратно: а б Кацаневакис С., Протопапас Н., Милиу Х. и Верриопулос Г. (2005). Влияние температуры на специфическое динамическое действие осьминога обыкновенного Octopus vulgaris (Cephalopoda). Морская биология, 146, 733–738.
- ^ Перейти обратно: а б с д и ж Инь, Фэй; Сунь, Пэн; Пэн, Шимин; Тан, Баоцзюнь; Чжан, Донг; Ван, Чуньлинь; Му, Чанкао; Ши, Чжаохун (июль 2013 г.). «Дыхание, выделение и биохимическая реакция молоди обыкновенной китайской каракатицы Sepiella maindroni при разных температурах». Аквакультура . 402–403: 127–132. Бибкод : 2013Aquac.402..127Y . doi : 10.1016/j.aquacultural.2013.03.018 .
- ^ Перейти обратно: а б с д и Зейлински С. и др. 2001. Влияние температуры на связывание кислорода гемоцианином у антарктических головоногих моллюсков. Биологический вестник. 200 (1): 67–76. дои : 10.2307/1543086 . ПМИД 11249213 .
- ^ Перейти обратно: а б с д Нойола Дж., Каамаль-Монреаль К., Диас Ф., Ре Д., Санчес А. и Росас К. (2013). Термопредпочтение, толерантность и скорость метаболизма молодых особей Octopus maya на ранних стадиях акклиматизации к различным температурам. Журнал термической биологии, 38, 14–19.
- ^ Отеро, Гонсалес, А. Ф., Сиейро, член парламента, и Герра, А. (2007). Репродуктивный цикл и распределение энергии Octopus vulgaris в водах Галиции, северо-восток Атлантики. Исследования рыболовства, 85 (1), 122–129. https://doi.org/10.1016/j.fishres.2007.01.007
Внешние ссылки
[ редактировать ]- Кинтейро, Хавьер; Байбай, Тарик; Ухаттар, Лейла; Сукри, Абдельазиз; Сейшас, Пабло; Рей-Мендес, Мануэль (2011). «Множественное отцовство у обыкновенного осьминога Octopus vulgaris (Cuvier, 1797), выявленное с помощью микросателлитного анализа ДНК» (PDF) . Моллюсковые исследования . 31 (1): 15–20. CiteSeerX 10.1.1.363.8342 . дои : 10.11646/mr.31.1.2 .
- «CephBase: Обыкновенный осьминог» . Архивировано из оригинала 17 августа 2005 года.
- Фотографии обыкновенного осьминога из коллекции Sealife
- Виды, находящиеся в Красном списке МСОП, вызывающие наименьшее беспокойство
- Осьминоги
- Моллюски Атлантического океана
- Моллюски Средиземного моря
- Морские моллюски Африки
- Морские моллюски Европы
- Фауна Макаронезии
- Фауна острова Гоф
- Головоногие моллюски описаны в 1797 году.
- Таксоны, названные Жоржем Кювье
- Головоногие моллюски Европы