Глюкоза
 Скелетная формула d -глюкоза | |
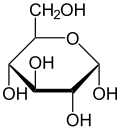 Проекция Хаворта α- d -глюкопираноза | |
 Фишера Проекция d -глюкоза | |
| Имена | |
|---|---|
| Произношение | / ˈ ɡ l uː k oʊ z / , / ɡ l uː k oʊ s / |
| Название ИЮПАК Разрешенные тривиальные имена: [1]
| |
| Предпочтительное название ИЮПАК ПИН-коды не идентифицируются для натуральных продуктов. | |
Систематическое название ИЮПАК
| |
| Другие имена Сахар в крови Декстроза Кукурузный сахар г -Глюкоза Виноградный сахар | |
| Идентификаторы | |
3D model ( JSmol ) |
|
| 3DMeet | |
| Сокращения | Глк |
| 1281604 | |
| ЧЭБИ | |
| ЧЕМБЛ | |
| ХимическийПаук | |
| Номер ЕС |
|
| 83256 | |
| КЕГГ | |
| МеШ | Глюкоза |
ПабХим CID | |
| номер РТЭКС |
|
| НЕКОТОРЫЙ |
|
| Характеристики | |
| С 6 Н 12 О 6 | |
| Молярная масса | 180.156 g/mol |
| Появление | Белый порошок |
| Плотность | 1,54 г/см 3 |
| Температура плавления | а- d -Глюкоза: 146 °C (295 °F; 419 К) β- d -Глюкоза: 150 ° C (302 ° F; 423 К) |
| 909 г/л (25 °C (77 °F)) | |
| −101.5×10 −6 см 3 /моль | |
| 8.6827 | |
| Термохимия | |
Теплоемкость ( С ) | 218,6 Дж/(К·моль) [2] |
Стандартный моляр энтропия ( S ⦵ 298 ) | 209,2 Дж/(К·моль) [2] |
Стандартная энтальпия образование (Δ f H ⦵ 298 ) | −1271 кДж/моль [3] |
| 2805 кДж/моль (670 ккал/моль) | |
| Фармакология | |
| B05CX01 ( ВОЗ ) V04CA02 ( ВОЗ ), V06DC01 ( ВОЗ ) | |
| Опасности | |
| NFPA 704 (огненный алмаз) | |
| Паспорт безопасности (SDS) | КМГС 08655 |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |

Глюкоза – это сахар с молекулярной формулой С 6 Н 12 О 6 . Глюкоза в целом является наиболее распространенным моносахаридом . [4] подкатегория углеводов . Глюкоза в основном вырабатывается растениями и большинством водорослей в ходе фотосинтеза из воды и углекислого газа с использованием энергии солнечного света, при этом она используется для производства целлюлозы в клеточных стенках , самого распространенного углевода в мире, или АТФ ( аденозинтрифосфата ), который используется клеткой как энергия. [5] [6] [7]
В энергетическом обмене глюкоза является важнейшим источником энергии во всех организмах . Глюкоза для метаболизма запасается в виде полимера , у растений главным образом в виде крахмала и амилопектина , а у животных - в виде гликогена . Глюкоза циркулирует в крови животных в виде сахара в крови . [5] [7] Природной формой глюкозы является d -глюкоза, а ее стереоизомер L -глюкоза производится синтетическим путем в сравнительно небольших количествах и менее биологически активна. [7] Глюкоза представляет собой моносахарид, содержащий шесть атомов углерода и альдегидную группу, и, следовательно, является альдогексозой . Молекула глюкозы может существовать как в открытой (ациклической), так и в кольцевой (циклической) форме. Глюкоза встречается в природе и в свободном состоянии содержится во фруктах и других частях растений. У животных глюкоза высвобождается в результате распада гликогена в процессе, известном как гликогенолиз .
Глюкоза в виде раствора сахара для внутривенного введения включена в Список основных лекарственных средств Всемирной организации здравоохранения . [8] Он также находится в списке в сочетании с хлоридом натрия (поваренной солью). [8]
Название глюкозы происходит от древнегреческого γλεῦκος ( gleûkos , «вино, сусло»), от γλυκύς ( glykýs , «сладкое»). [9] [10] Суффикс « -оза » представляет собой химический классификатор, обозначающий сахар.
История [ править ]
Глюкозу впервые выделил из изюма в 1747 году немецкий химик Андреас Маргграф . [11] [12] Глюкоза была обнаружена в винограде другим немецким химиком — Иоганном Тобиасом Ловицем — в 1792 году и отличалась от тростникового сахара ( сахарозы ). Глюкоза — термин, введенный Жаном Батистом Дюма в 1838 году и преобладающий в химической литературе. Фридрих Август Кекуле предложил термин декстроза (от латинского dexter , что означает «правый»), поскольку в водном растворе глюкозы плоскость линейно поляризованного света повернута вправо. Напротив, l-фруктоза (обычно называемая d -фруктоза) (кетогексоза) и l-глюкоза ( l -глюкоза) поворачивают линейно поляризованный свет влево. От более ранних обозначений вращения плоскости линейно поляризованного света ( d и l -номенклатура) позже отказались в пользу д - и l -обозначение , которое относится к абсолютной конфигурации асимметричного центра, наиболее удаленного от карбонильной группы, и в соответствии с конфигурацией д - или л -глицеральдегид. [13] [14]
Поскольку глюкоза является основной потребностью многих организмов, правильное понимание ее химического состава и структуры во многом способствовало общему прогрессу органической химии . Это понимание возникло во многом в результате исследований Эмиля Фишера , немецкого химика, получившего Нобелевскую премию по химии 1902 года за свои открытия. [15] Синтез глюкозы установил структуру органического материала и, следовательно, стал первым окончательным подтверждением теорий Якобуса Хенрикуса ван 'т Хоффа о химической кинетике и расположении химических связей в углеродсодержащих молекулах. [16] Между 1891 и 1894 годами Фишер установил стереохимическую конфигурацию всех известных сахаров и правильно предсказал возможные изомеры , применив теорию Вант-Гоффа об асимметричных атомах углерода. Названия изначально относились к природным веществам. Их энантиомерам были присвоены одинаковые названия с введением систематических номенклатур, учитывающих абсолютную стереохимию (например, номенклатура Фишера, д / номенклатура ).
За открытие метаболизма глюкозы Отто Мейергоф получил Нобелевскую премию по физиологии и медицине в 1922 году. [17] Ганс фон Эйлер-Хелпин был удостоен Нобелевской премии по химии вместе с Артуром Харденом в 1929 году за «исследование ферментации сахара и доли ферментов в этом процессе». [18] [19] В 1947 году Бернардо Уссей (за открытие роли гипофиза в метаболизме глюкозы и производных углеводов), а также Карл и Герти Кори (за открытие превращения гликогена из глюкозы) получили Нобелевскую премию. Физиология или медицина. [20] [21] [22] В 1970 году Луис Лелуар был удостоен Нобелевской премии по химии за открытие сахарных нуклеотидов, производных глюкозы, в биосинтезе углеводов. [23]
Химические и физические свойства [ править ]
Глюкоза образует белые или бесцветные твердые вещества, хорошо растворимые в воде и уксусной кислоте, но плохо растворимые в метаноле и этаноле . Они плавятся при 146 °C (295 °F) ( α ) и 150 °C (302 °F) ( бета ), разлагаются , начиная с 188 °C (370 °F) с выделением различных летучих продуктов, в конечном итоге оставляя остаток углерод . [24] Глюкоза имеет значение pKa 12,16 при температуре 25 ° C (77 ° F) в воде. [25]
Имея шесть атомов углерода, он классифицируется как гексоза , подкатегория моносахаридов . d -Глюкоза — один из шестнадцати альдогексозы стереоизомеров . д - изомер , d -глюкоза, также известная как декстроза, широко встречается в природе, но l -изомер, л -глюкоза , нет. Глюкозу можно получить гидролизом углеводов, таких как молочный сахар ( лактоза ), тростниковый сахар (сахароза), мальтоза , целлюлоза , гликоген и т. д. Декстроза обычно коммерчески производится из крахмалов , таких как кукурузный крахмал в США и Японии, из картофеля. и пшеничный крахмал в Европе, а также крахмал тапиоки в тропических регионах. [26] В производственном процессе используется гидролиз посредством обработки паром под давлением при контролируемом pH в струе с последующей ферментативной деполимеризацией. [27] Несвязанная глюкоза является одним из основных ингредиентов меда . [28] [29] [30] [31] [32]
Термин «декстроза» часто используется в клиническом (связанном с состоянием здоровья пациента) или пищевом контексте (связанном с приемом пищи, например, на этикетках пищевых продуктов или в диетических руководствах), тогда как «глюкоза» используется в биологическом или физиологическом контексте (химические процессы и диетические рекомендации). молекулярные взаимодействия), [33] [34] [35] [36] но оба термина относятся к одной и той же молекуле, а именно к D-глюкозе. [35] [37]
Моногидрат декстрозы представляет собой гидратированную форму D-глюкозы, что означает, что это молекула глюкозы с присоединенной дополнительной молекулой воды. [38] Его химическая формула С 6 Н 12 О 6 · Н 2 О. [38] [39] Моногидрат декстрозы также называют гидратированной D-глюкозой и обычно производят из растительных крахмалов. [38] [40] Моногидрат декстрозы используется в качестве преобладающего типа декстрозы в пищевых продуктах, таких как смеси для напитков, — это распространенная форма глюкозы, широко используемая в качестве пищевой добавки при производстве пищевых продуктов. Моногидрат декстрозы в основном потребляется в Северной Америке в виде кукурузного сиропа или кукурузного сиропа с высоким содержанием фруктозы . [35]
С другой стороны, безводная декстроза представляет собой глюкозу, к которой не присоединены молекулы воды. [40] [41] Безводные химические вещества обычно производятся путем удаления воды из гидратированного вещества с помощью таких методов, как нагревание или сушка (высушивание). [42] [43] [44] Моногидрат декстрозы можно дегидратировать до безводной декстрозы в промышленных условиях. [45] [46] Моногидрат декстрозы состоит примерно из 9,5% воды по массе; в процессе обезвоживания эта вода удаляется с образованием безводной (сухой) декстрозы. [40]
Безводная декстроза имеет химическую формулу C 6 H 12 O 6 , без присоединенной молекулы воды, что аналогично глюкозе. [38] Безводная декстроза на открытом воздухе имеет тенденцию поглощать влагу и превращаться в моногидрат, и ее производство обходится дороже. [40] Безводная декстроза (безводная D-глюкоза) обладает повышенной стабильностью и увеличенным сроком хранения. [43] имеет медицинское применение, например, в пероральном тесте на толерантность к глюкозе (OGTT). [47]
Тогда как молекулярная масса (молярная масса) моногидрата D-глюкозы составляет 198,17 г/моль, [48] [49] что для безводной D-глюкозы составляет 180,16 г/моль. [50] [51] [52] Плотность этих двух форм глюкозы также различна. [ указать ]
По химической структуре глюкоза представляет собой моносахарид , то есть простой сахар. Глюкоза содержит шесть атомов углерода и альдегидную группу и, следовательно, является альдогексозой . Молекула глюкозы может существовать в форме с открытой цепью (ациклической), а также в кольцевой (циклической) форме — из-за присутствия спиртовых и альдегидных или кетоновых функциональных групп форма с прямой цепью может легко превращаться в полуацетальную форму, подобную стулу. кольцевая структура, обычно встречающаяся в углеводах. [53]
Структура и номенклатура [ править ]

Глюкоза присутствует в твердой форме в виде моногидрата с замкнутым пирановым кольцом (моногидрат α-глюкопиранозы, иногда менее точно известный как гидрат декстрозы). С другой стороны, в водном растворе он в небольшой степени имеет открытую цепь и присутствует преимущественно в виде α- или β- пиранозы , которые взаимно превращаются. Из водных растворов можно кристаллизовать три известные формы: α-глюкопиранозу, β-глюкопиранозу и моногидрат α-глюкопиранозы. [54] Глюкоза является строительным блоком дисахаридов лактозы и сахарозы (тростниковый или свекловичный сахар), олигосахаридов, таких как рафиноза , и полисахаридов, таких как крахмал , амилопектин , гликоген и целлюлоза . [7] [55] Температура стеклования глюкозы составляет 31 ° C (88 ° F), а константа Гордона – Тейлора (экспериментально определенная константа для прогнозирования температуры стеклования для разных массовых долей смеси двух веществ) [55] составляет 4,5. [56]
| Формы и проекции d -глюкоза в сравнении | ||
|---|---|---|
| Проекция Натта | Проекция Хауорта | |
 |  а- d -глюкофураноза а- d -глюкофураноза | 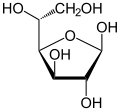 б- d -глюкофураноза б- d -глюкофураноза |
 а- d -глюкопираноза а- d -глюкопираноза |  б- d -глюкопираноза б- d -глюкопираноза | |
| а- d -глюкопираноза в (1) Толленсе/Фишере (2) проекции Хауорта (3) конформации стула (4) проекции Миллса | ||
 | ||
Открытая форма цепочки [ править ]

Форма глюкозы с открытой цепью составляет менее 0,02% молекул глюкозы в водном растворе в равновесии. [57] Остальное представляет собой одну из двух циклических полуацетальных форм. В своей форме с открытой цепью молекула глюкозы имеет открытую (в отличие от циклической ) неразветвленную основную цепь из шести атомов углерода, где C-1 является частью альдегидной группы. Н(С=О)- . Поэтому глюкозу также классифицируют как альдозу или альдогексозу . Альдегидная группа делает глюкозу редуцирующим сахаром, что дает положительную реакцию в тесте Фелинга .
Циклические формы [ править ]
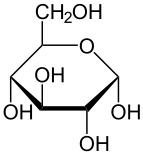
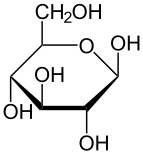
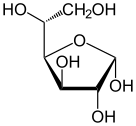
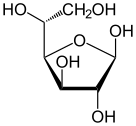
В растворах форма глюкозы с открытой цепью (либо « D- », либо « L- ») существует в равновесии с несколькими циклическими изомерами , каждый из которых содержит кольцо атомов углерода, замкнутое одним атомом кислорода. Однако в водном растворе более 99% молекул глюкозы существуют в форме пиранозы . Форма с открытой цепью ограничена примерно 0,25%, а формы фуранозы существуют в незначительных количествах. Термины «глюкоза» и « D -глюкоза» обычно также используются для этих циклических форм. Кольцо возникает из формы с открытой цепью в результате внутримолекулярной реакции нуклеофильного присоединения между альдегидной группой (при C-1) и гидроксильной группой C-4 или C-5, образуя полуацетальную связь. -C(OH)H-O- .
Реакция между C-1 и C-5 дает шестичленную гетероциклическую систему, называемую пиранозой, которая представляет собой моносахаридный сахар (отсюда и «-оза»), содержащий производное пиранового скелета. (Намного более редкая) реакция между C-1 и C-4 дает пятичленное фуранозное кольцо, названное в честь циклического эфира фурана . В любом случае к каждому углероду в кольце присоединен один водород и один гидроксил, за исключением последнего углерода (C-4 или C-5), где гидроксил заменен остатком открытой молекулы (который является −(C(CH 2 OH)HOH)−H или −(CHOH)−H соответственно).
Реакция замыкания цикла может дать два продукта, обозначенных «α-» и «β-». Когда молекулу глюкопиранозы рисуют в проекции Хаворта , обозначение «α-» означает, что гидроксильная группа присоединена к С-1, а Группа −CH 2 OH при C-5 лежит на противоположных сторонах плоскости кольца ( транс -расположение), а «β-» означает, что они находятся на одной стороне плоскости ( цис -расположение). Следовательно, изомер D- глюкозы с открытой цепью дает четыре различных циклических изомера: α- D -глюкопиранозу, β- D -глюкопиранозу, α- D -глюкофуранозу и β- D -глюкофуранозу. Эти пять структур существуют в равновесии и взаимопревращаются, причем взаимное превращение происходит гораздо быстрее при кислотном катализе .
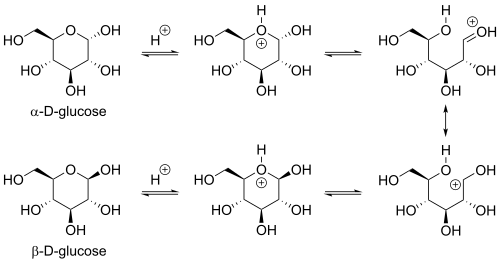

Другой изомер L -глюкозы с открытой цепью аналогичным образом дает начало четырем различным циклическим формам L -глюкозы, каждая из которых является зеркальным отражением соответствующей D -глюкозы.
Глюкопиранозное кольцо (α или β) может принимать несколько неплоских форм, аналогичных конформациям «кресло» и «лодочка» циклогексана . Точно так же глюкофуранозное кольцо может принимать несколько форм, аналогичных конформациям «конверта» циклопентана .
В твердом состоянии наблюдаются только формы глюкопиранозы.
Некоторые производные глюкофуранозы, такие как 1,2 -O -изопропилиден- D -глюкофураноза, стабильны и могут быть получены в чистом виде в виде кристаллических твердых веществ. [58] [59] Например, реакция α-D-глюкозы с пара -толилбороновой кислотой. H 3 C-(C 6 H 4 )-B(OH) 2 преобразует нормальное пиранозное кольцо с образованием 4-кратного сложного эфира α-D-глюкофуранозо-1,2:3,5-бис( п -толилборонат). [60]
Мутаротация [ править ]
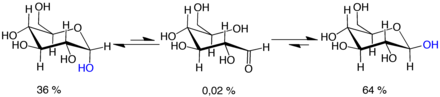
Мутаротация состоит из временного изменения направления реакции образования кольца, что приводит к образованию открытой цепи с последующим реформированием кольца. Этап замыкания кольца может использовать другой -OH- группа, чем та, которая воссоздана на стадии открытия (таким образом переключаясь между формами пиранозы и фуранозы), или новая полуацетальная группа, созданная на C-1, может иметь ту же или противоположную направленность, что и исходная (таким образом, переключаясь между α и β формы). Таким образом, хотя форма с открытой цепью едва обнаруживается в растворе, она является важным компонентом равновесия.
Форма с открытой цепью термодинамически нестабильна и спонтанно изомеризуется в циклические формы. (Хотя реакция замыкания кольца теоретически может создавать четырех- или трехатомные кольца, они будут сильно напряжены и не наблюдаются на практике.) В растворах при комнатной температуре четыре циклических изомера взаимопревращаются в течение часов, в процессе, называемом мутаротацией . [61] Начиная с любых пропорций, смесь сходится к стабильному соотношению α:β 36:64. Соотношение α:β было бы 11:89, если бы не влияние аномерного эффекта . [62] Мутаротация происходит значительно медленнее при температуре, близкой к 0 ° C (32 ° F).
Оптическая активность [ править ]
Будь то вода или твердая форма, d -(+)-глюкоза правовращающая , то есть она будет вращать направление поляризованного света по часовой стрелке, если смотреть в сторону источника света. Эффект обусловлен хиральностью молекул, да и вообще зеркальным изомером, l -(-)-глюкоза обладает левовращающим действием (вращает поляризованный свет против часовой стрелки) на такую же величину. Сила эффекта различна для каждого из пяти таутомеров .
Обратите внимание, что г – приставка не относится непосредственно к оптическим свойствам соединения. Это указывает на то, что хиральный центр С-5 имеет ту же направленность, что и d -глицеральдегид (который был назван так, потому что он правовращающий). Тот факт, что d -глюкоза имеет правовращающее действие – это комбинированный эффект ее четырех хиральных центров, а не только C-5; и вообще некоторые другие d- альдогексозы обладают левовращающими свойствами.
Превращение между двумя аномерами можно наблюдать в поляриметре, поскольку чистый α- d -глюкоза имеет удельный угол вращения +112,2° мл/(дм·г), чистая β- г - глюкоза +17,5° мл/(дм·г). [63] Когда равновесие достигается через определенное время за счет мутаротации, угол поворота составляет +52,7° мл/(дм·г). [63] При добавлении кислоты или основания это превращение значительно ускоряется. Уравновешивание происходит через альдегидную форму с открытой цепью.
Изомеризация [ править ]
В разбавленном гидроксиде натрия или других разбавленных основаниях моносахариды манноза , глюкоза и фруктоза взаимно превращаются (посредством превращения Лобри де Брюйна-Альберды-Ван Экенштейна ), так что формируется баланс между этими изомерами. Эта реакция протекает через эндиол :
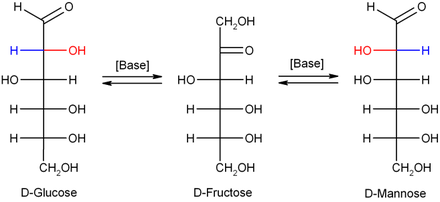
Биохимические свойства [ править ]
| Метаболизм обычных моносахаридов и некоторые биохимические реакции глюкозы. |
|---|

Глюкоза – самый распространенный моносахарид. Глюкоза также является наиболее широко используемой альдогексозой в большинстве живых организмов. Одним из возможных объяснений этого является то, что глюкоза имеет меньшую склонность, чем другие альдогексозы, к неспецифической реакции аминогруппами белков с . [64] Эта реакция — гликирование — нарушает или разрушает функцию многих белков. [64] например, в гликированном гемоглобине . Низкую скорость гликирования глюкозы можно объяснить тем, что она имеет более стабильную циклическую форму по сравнению с другими альдогексозами, что означает, что она тратит меньше времени, чем в ее реакционноспособной форме с открытой цепью . [64] Причина того, что глюкоза имеет наиболее стабильную циклическую форму из всех альдогексоз, заключается в том, что ее гидроксильные группы (за исключением гидроксигруппы на аномерном углероде d -глюкоза) находятся в экваториальном положении . Предположительно, глюкоза является наиболее распространенным природным моносахаридом, поскольку она менее гликируется с белками, чем другие моносахариды. [64] [65] Другая гипотеза состоит в том, что глюкоза, будучи единственной d -альдогексоза, имеющая все пять гидроксизаместителей в экваториальном положении в форме β- d -глюкоза более доступна для химических реакций, [66] : 194, 199 например, для этерификации [67] : 363 или образование ацеталя . [68] По этой причине, d -глюкоза также является весьма предпочтительным строительным блоком природных полисахаридов (гликанов). Полисахариды, состоящие исключительно из глюкозы, называются глюканами .
Глюкоза вырабатывается растениями путем фотосинтеза с использованием солнечного света. [69] [70] вода и углекислый газ и могут использоваться всеми живыми организмами в качестве источника энергии и углерода. Однако большая часть глюкозы встречается не в свободной форме, а в виде ее полимеров, т.е. лактозы, сахарозы, крахмала и других, которые являются запасными энергетическими веществами, а также целлюлозы и хитина , которые являются компонентами клеточной стенки растений или грибов. и членистоногих соответственно. Эти полимеры, потребляемые животными, грибами и бактериями, разлагаются до глюкозы с помощью ферментов. Все животные также способны сами производить глюкозу из определенных предшественников по мере возникновения необходимости. Нейроны , клетки мозгового слоя почек и эритроциты зависят от глюкозы для производства энергии. [70] У взрослого человека содержится около 18 г (0,63 унции) глюкозы, [71] из них около 4 г (0,14 унции) присутствует в крови. [72] Примерно 180–220 г (6,3–7,8 унций) глюкозы вырабатывается в печени взрослого человека за 24 часа. [71]
Многие из долгосрочных осложнений диабета (например, слепота , почечная недостаточность и периферическая нейропатия ), вероятно, обусловлены гликированием белков или липидов . [73] Напротив, регулируемое ферментами добавление сахаров к белку называется гликозилированием и имеет важное значение для функционирования многих белков. [74]
Поглощение [ править ]
Поступившая в организм глюкоза первоначально связывается с рецептором сладкого вкуса на языке человека. Этот комплекс белков T1R2 и T1R3 позволяет идентифицировать глюкозосодержащие источники пищи. [75] [76] Глюкоза в основном поступает с пищей — около 300 г (11 унций) в день производится путем переработки пищи. [76] но он также синтезируется из других метаболитов в клетках организма. У человека расщепление глюкозосодержащих полисахаридов происходит частично уже во время жевания посредством амилазы , содержащейся в слюне , а также мальтазы , лактазы и сахаразы на щеточной кайме тонкой кишки . Глюкоза является строительным блоком многих углеводов и может быть отделена от них с помощью определенных ферментов. Глюкозидазы , подгруппа гликозидаз, сначала катализируют гидролиз длинноцепочечных глюкозосодержащих полисахаридов, удаляя концевую глюкозу. В свою очередь, дисахариды в основном расщепляются специфическими гликозидазами до глюкозы. Названия разлагающих ферментов часто происходят от конкретных поли- и дисахаридов; в частности, для деградации полисахаридных цепей используются амилазы (названные в честь амилозы, компонента крахмала), целлюлазы (названные в честь целлюлозы), хитиназы (названные в честь хитина) и другие. Кроме того, для расщепления дисахаридов используются мальтаза, лактаза, сахараза, трегалаза и другие. У человека известно около 70 генов, кодирующих гликозидазы. Они участвуют в переваривании и расщеплении гликогена, сфинголипидов , мукополисахаридов и поли( АДФ-рибозы ). Люди не производят целлюлазы, хитиназы или трегалазы, но это делают бактерии в микробиоте кишечника .
Чтобы проникнуть в клеточные мембраны клеток и мембраны клеточных компартментов или выйти из них, глюкозе необходимы специальные транспортные белки из суперсемейства основных фасилитаторов . В тонком кишечнике (точнее, в тощей кишке ), [77] Глюкоза попадает в эпителий кишечника с помощью транспортеров глюкозы. [78] посредством вторичного активного механизма транспорта ионов натрия и глюкозы , называемого симпортом через котранспортер натрия/глюкозы 1 (SGLT1). [79] Дальнейший перенос происходит на базолатеральную сторону эпителиальных клеток кишечника через транспортер глюкозы GLUT2 . [79] а также поглощение клетками печени , клетками почек, клетками островков Лангерганса , нейронами , астроцитами и таницитами . [80] Глюкоза поступает в печень через воротную вену и сохраняется там в виде клеточного гликогена. [81] В клетке печени он фосфорилируется глюкокиназой в положении 6 с образованием глюкозо-6-фосфата , который не может покинуть клетку. Глюкозо-6-фосфатаза может превращать глюкозо-6-фосфат обратно в глюкозу исключительно в печени, поэтому организм может поддерживать достаточную концентрацию глюкозы в крови. В других клетках поглощение происходит путем пассивного транспорта через один из 14 белков GLUT. [79] В других типах клеток фосфорилирование происходит посредством гексокиназы , после чего глюкоза больше не может диффундировать из клетки.
Переносчик глюкозы GLUT1 вырабатывается большинством типов клеток и имеет особое значение для нервных клеток и β-клеток поджелудочной железы . [79] GLUT3 высоко экспрессируется в нервных клетках. [79] Глюкоза из кровотока поглощается GLUT4 из мышечных клеток ( скелетных мышц). [82] и сердечная мышца ) и жировые клетки . [83] GLUT14 экспрессируется исключительно в яичках . [84] Избыточная глюкоза расщепляется и превращается в жирные кислоты, которые сохраняются в виде триглицеридов . В почках глюкоза в моче всасывается через SGLT1 и SGLT2 в апикальных клеточных мембранах и передается через GLUT2 в базолатеральных клеточных мембранах. [85] Около 90% реабсорбции глюкозы в почках происходит через SGLT2 и около 3% через SGLT1. [86]
Биосинтез [ править ]
У растений и некоторых прокариот глюкоза — продукт фотосинтеза . [69] Глюкоза также образуется в результате распада полимерных форм глюкозы, таких как гликоген (у животных и грибов ) или крахмал (у растений). Расщепление гликогена называется гликогенолизом, расщепление крахмала — деградацией крахмала. [87]
Метаболический путь, который начинается с молекул, содержащих от двух до четырех атомов углерода (С), и заканчивается молекулой глюкозы, содержащей шесть атомов углерода, называется глюконеогенезом и встречается во всех живых организмах. Меньшие исходные материалы являются результатом других метаболических путей. В конечном счете почти все биомолекулы образуются в результате ассимиляции углекислого газа растениями и микробами в ходе фотосинтеза. [67] : 359 Свободная энергия образования α- d -глюкоза составляет 917,2 килоджоуля на моль. [67] : 59 У человека глюконеогенез происходит в печени и почках. [88] но и в других типах клеток. В печени хранится около 150 г (5,3 унции) гликогена, в скелетных мышцах — около 250 г (8,8 унции). [89] Однако глюкоза, высвобождаемая в мышечных клетках при расщеплении гликогена, не может быть доставлена в кровоток, поскольку глюкоза фосфорилируется гексокиназой, а глюкозо-6-фосфатаза не экспрессируется для удаления фосфатной группы. нет транспортного белка В отличие от глюкозы, для глюкозо-6-фосфата . Глюконеогенез позволяет организму накапливать глюкозу из других метаболитов, включая лактат или определенные аминокислоты , потребляя при этом энергию. Клетки почечных канальцев также могут производить глюкозу.
Глюкозу также можно найти вне живых организмов в окружающей среде. Концентрация глюкозы в атмосфере определяется путем сбора проб с самолета и, как известно, варьируется от места к месту. Например, концентрации глюкозы в атмосферном воздухе внутреннего Китая колеблются от 0,8 до 20,1 пг/л, тогда как концентрации глюкозы в восточном прибрежном Китае колеблются от 10,3 до 142 пг/л. [90]
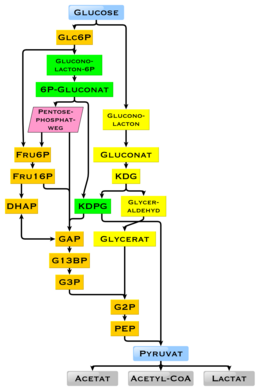
глюкозы Деградация
У человека глюкоза метаболизируется путем гликолиза. [91] и пентозофосфатный путь. [92] Гликолиз используется всеми живыми организмами, [66] : 551 [93] с небольшими вариациями, и все организмы генерируют энергию за счет распада моносахаридов. [93] В дальнейшем ходе метаболизма он может полностью разлагаться посредством окислительного декарбоксилирования , цикла лимонной кислоты (синоним цикл Кребса ) и дыхательной цепи до воды и углекислого газа. Если для этого недостаточно кислорода, разложение глюкозы у животных происходит анаэробно до лактата посредством ферментации молочной кислоты и высвобождается гораздо меньше энергии. Мышечный лактат попадает в печень через кровоток у млекопитающих, где происходит глюконеогенез ( цикл Кори ). При высоком запасе глюкозы метаболит ацетил-КоА из цикла Кребса также может использоваться для синтеза жирных кислот . [94] Глюкоза также используется для пополнения запасов гликогена в организме, который в основном содержится в печени и скелетных мышцах. Эти процессы регулируются гормонально .
У других живых организмов могут возникать и другие формы ферментации. Бактерия Escherichia coli может расти на питательных средах, содержащих глюкозу в качестве единственного источника углерода. [67] : 59 У некоторых бактерий, а в модифицированной форме также у архей, глюкоза расщепляется по пути Энтнера-Дудорова . [95] был обнаружен механизм регуляции генов Что касается глюкозы, то в E. coli — катаболитная репрессия (ранее известная как эффект глюкозы ). [96]
Использование глюкозы в качестве источника энергии в клетках осуществляется посредством аэробного дыхания, анаэробного дыхания или ферментации. [97] Первым этапом гликолиза является фосфорилирование глюкозы гексокиназой с образованием глюкозо-6-фосфата . Основная причина немедленного фосфорилирования глюкозы заключается в предотвращении ее диффузии из клетки, поскольку заряженная фосфатная группа предотвращает легкое прохождение глюкозо-6-фосфата через клеточную мембрану . [97] Кроме того, добавление высокоэнергетической фосфатной группы активирует глюкозу для последующего расщепления на более поздних стадиях гликолиза. [98] В физиологических условиях эта первоначальная реакция необратима. [ нужна медицинская ссылка ]
При анаэробном дыхании одна молекула глюкозы дает чистый прирост в размере двух молекул АТФ (четыре молекулы АТФ образуются во время гликолиза посредством фосфорилирования на уровне субстрата, но две требуются ферментам, используемым в этом процессе). [99] При аэробном дыхании молекула глюкозы гораздо более выгодна, поскольку генерируется максимальная чистая продукция 30 или 32 молекул АТФ (в зависимости от организма). [100]
Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [§ 1]

- ^ Интерактивную карту маршрутов можно редактировать на WikiPathways: «Гликолиз-Глюконеогенез_WP534» .
Опухолевые клетки часто растут сравнительно быстро и потребляют количество глюкозы, превышающее среднее, путем гликолиза. [101] что приводит к образованию лактата, конечного продукта брожения у млекопитающих, даже в присутствии кислорода. Это называется эффектом Варбурга . Для повышенного поглощения глюкозы опухолями чрезмерно вырабатываются различные SGLT и GLUT. [102] [103]
В дрожжах этанол ферментируется при высоких концентрациях глюкозы, даже в присутствии кислорода (что обычно приводит к дыханию, а не к брожению). Это называется эффектом Крэбтри .
Глюкоза также может разлагаться с образованием углекислого газа абиотическим путем. Экспериментально было показано, что это происходит посредством окисления и гидролиза при 22 ° C и pH 2,5. [104]
Источник энергии [ править ]
Глюкоза – повсеместное топливо в биологии . Он используется в качестве источника энергии в организмах, от бактерий до человека, посредством аэробного дыхания , анаэробного дыхания (у бактерий) или ферментации . Глюкоза является ключевым источником энергии человеческого организма посредством аэробного дыхания, обеспечивая около 3,75 килокалорий (16 килоджоулей ) пищевой энергии на грамм. [105] Расщепление углеводов (например, крахмала) приводит к образованию моно- и дисахаридов , большую часть которых составляет глюкоза. В результате гликолиза , а затем в реакциях цикла лимонной кислоты и окислительного фосфорилирования глюкоза окисляется с образованием углекислого газа и воды, выделяя энергию в основном в форме АТФ . Реакция инсулина и другие механизмы регулируют концентрацию глюкозы в крови. Физиологическая калорийность глюкозы в зависимости от источника составляет 16,2 килоджоуля на грамм. [106] или 15,7 кДж/г (3,74 ккал/г). [107] Высокая доступность углеводов из растительной биомассы привела к появлению в ходе эволюции, особенно у микроорганизмов, различных методов использования глюкозы для получения энергии и хранения углерода. Существуют различия, при которых конечный продукт больше не может использоваться для производства энергии. Наличие отдельных генов и их генных продуктов — ферментов — определяет возможные реакции. Метаболический путь гликолиза используется практически всеми живыми существами. Существенным отличием использования гликолиза является восстановление НАДФН в качестве восстановителя анаболизма , который в противном случае пришлось бы генерировать косвенно. [108]
почти всей энергией Глюкоза и кислород снабжают мозг . [109] поэтому его наличие влияет на психологические процессы. При низком уровне глюкозы психологические процессы, требующие умственных усилий (например, самоконтроль , принятие решений, требующих усилий). нарушаются [110] [111] [112] [113] В мозге, который зависит от глюкозы и кислорода как основных источников энергии, концентрация глюкозы обычно составляет от 4 до 6 мМ (5 мМ соответствует 90 мг/дл). [71] но снижается до 2–3 мМ при голодании. [114] Спутанность сознания возникает при концентрациях ниже 1 мМ и кома при более низких уровнях. [114]
Глюкоза в крови называется сахаром крови . Уровень сахара в крови регулируется глюкозосвязывающими нервными клетками гипоталамуса . [115] Кроме того, глюкоза в мозге связывается с рецепторами глюкозы системы вознаграждения в прилежащем ядре . [115] Связывание глюкозы с рецептором сладкого на языке вызывает высвобождение различных гормонов энергетического обмена либо через глюкозу, либо через другие сахара, что приводит к увеличению клеточного поглощения и снижению уровня сахара в крови. [116] Искусственные подсластители не снижают уровень сахара в крови. [116]
Содержание сахара в крови здорового человека в состоянии кратковременного голодания, например, после ночного голодания, составляет около 70–100 мг/дл крови (4–5,5 мМ). В плазме крови измеренные значения примерно на 10–15% выше. Кроме того, значения в артериальной крови выше, чем концентрации в венозной крови, поскольку глюкоза всасывается в ткани при прохождении по капиллярному руслу . Также в капиллярной крови, которую часто используют для определения сахара в крови, значения иногда выше, чем в венозной крови. Содержание глюкозы в крови регулируется гормонами инсулином , инкретином и глюкагоном . [115] [117] Инсулин снижает уровень глюкозы, глюкагон его повышает. [71] Кроме того, гормоны адреналин , тироксин , глюкокортикоиды , соматотропин и адренокортикотропин приводят к повышению уровня глюкозы. [71] Существует также гормононезависимая регуляция, которая называется ауторегуляцией глюкозы . [118] После приема пищи концентрация сахара в крови увеличивается. Значения выше 180 мг/дл в венозной цельной крови являются патологическими и называются гипергликемией , значения ниже 40 мг/дл называются гипогликемией . [119] При необходимости глюкоза высвобождается в кровоток под действием глюкозо-6-фосфатазы из глюкозо-6-фосфата, происходящего из гликогена печени и почек, тем самым регулируя гомеостаз концентрации глюкозы в крови. [88] [70] У жвачных животных концентрация глюкозы в крови ниже (60 мг/дл у крупного рогатого скота и 40 мг/дл у овец ), поскольку углеводы в большей степени преобразуются микробиотой кишечника в короткоцепочечные жирные кислоты . [120]
Некоторое количество глюкозы преобразуется астроцитами в , молочную кислоту которая затем используется клетками мозга в качестве источника энергии ; некоторая часть глюкозы используется клетками кишечника и эритроцитами , а остальная часть достигает печени , жировой ткани и мышечных клеток, где всасывается и сохраняется в виде гликогена (под влиянием инсулина ). Гликоген клеток печени может превращаться в глюкозу и возвращаться в кровь при низком уровне инсулина или его отсутствии; Гликоген мышечных клеток не возвращается в кровь из-за недостатка ферментов. В жировых клетках глюкоза используется для запуска реакций, которые синтезируют некоторые типы жиров и имеют другие цели. Гликоген — это механизм «накопления энергии глюкозы» в организме, поскольку он гораздо более «экономичен по пространству» и менее реактивен, чем сама глюкоза.
Из-за своей важности для здоровья человека глюкоза является аналитом в тестах на глюкозу , которые являются обычными медицинскими анализами крови . [121] Прием пищи или голодание перед взятием образца крови влияет на анализ глюкозы в крови; Высокий уровень глюкозы в крови натощак может быть признаком предиабета или сахарного диабета . [122]
Гликемический индекс — это показатель скорости резорбции и преобразования в уровень глюкозы в крови из принятых углеводов, измеряемый как площадь под кривой уровня глюкозы в крови после потребления по сравнению с глюкозой (глюкоза определяется как 100). [123] Клиническое значение гликемического индекса является спорным. [123] [124] так как продукты с высоким содержанием жиров замедляют всасывание углеводов и снижают гликемический индекс, например мороженое. [124] Альтернативным показателем является инсулиновый индекс , [125] измеряется как влияние потребления углеводов на уровень инсулина в крови. Гликемическая нагрузка — это показатель количества глюкозы, добавленной к уровню глюкозы в крови после потребления, основанный на гликемическом индексе и количестве потребляемой пищи.
Предшественник [ править ]
Организмы используют глюкозу в качестве предшественника для синтеза ряда важных веществ. Крахмал, целлюлоза и гликоген («животный крахмал») представляют собой распространенные полимеры глюкозы (полисахариды). Некоторые из этих полимеров (крахмал или гликоген) служат запасами энергии, тогда как другие (целлюлоза и хитин , который производится из производного глюкозы) играют структурную роль. Олигосахариды глюкозы в сочетании с другими сахарами служат важными запасами энергии. К ним относятся лактоза, преобладающий сахар в молоке, который представляет собой дисахарид глюкозы-галактозы, и сахароза, еще один дисахарид, состоящий из глюкозы и фруктозы. Глюкоза также добавляется к определенным белкам и липидам в процессе, называемом гликозилированием . Зачастую это имеет решающее значение для их функционирования. Ферменты, которые присоединяют глюкозу к другим молекулам, обычно используют фосфорилированную глюкозу для формирования новой связи путем ее соединения с разрывом глюкозо-фосфатной связи.
Помимо прямого использования в качестве мономера, глюкоза может расщепляться с целью синтеза множества других биомолекул. Это важно, поскольку глюкоза служит одновременно основным хранилищем энергии и источником органического углерода. Глюкоза может расщепляться и превращаться в липиды . Он также является предшественником синтеза других важных молекул, таких как витамин С (аскорбиновая кислота). В живых организмах глюкоза превращается в несколько других химических соединений, которые являются исходным материалом для различных метаболических путей . Среди них все остальные моносахариды [126] такие как фруктоза (по полиольному пути ), [79] манноза (эпимер глюкозы в положении 2), галактоза (эпимер в положении 4), фукоза, различные уроновые кислоты и аминосахара . Из глюкозы образуются [81] Помимо фосфорилирования до глюкозо-6-фосфата, являющегося частью гликолиза, глюкоза может окисляться при ее распаде до глюконо-1,5-лактона . Глюкоза используется у некоторых бактерий в качестве строительного блока при биосинтезе трегалозы или декстрана , а у животных — как строительный блок гликогена. Глюкоза также может превращаться из бактериальной ксилозоизомеразы во фруктозу. Кроме того, метаболиты глюкозы производят все заменимые аминокислоты, сахарные спирты, такие как маннит и сорбит , жирные кислоты , холестерин и нуклеиновые кислоты . [126] Наконец, глюкоза используется в качестве строительного материала при гликозилировании белков до гликопротеинов , гликолипидов , пептидогликанов , гликозидов и других веществ (катализируемого гликозилтрансферазами ) и может быть отщеплена от них гликозидазами .
Патология [ править ]
Диабет [ править ]
Диабет — это нарушение обмена веществ, при котором организм не может регулировать уровень глюкозы в крови либо из-за недостатка инсулина в организме, либо из-за неспособности клеток организма должным образом реагировать на инсулин. Каждая из этих ситуаций может быть вызвана устойчиво высоким уровнем глюкозы в крови вследствие истощения поджелудочной железы и резистентности к инсулину . – Поджелудочная железа орган, ответственный за секрецию гормонов инсулина и глюкагона. [127] Инсулин — это гормон, который регулирует уровень глюкозы, позволяя клеткам организма поглощать и использовать глюкозу. Без него глюкоза не может попасть в клетку и, следовательно, не может использоваться в качестве топлива для функций организма. [128] Если поджелудочная железа подвергается постоянному повышению уровня глюкозы в крови, инсулин-продуцирующие клетки поджелудочной железы могут быть повреждены, что приведет к нехватке инсулина в организме. Инсулинорезистентность возникает, когда поджелудочная железа пытается вырабатывать все больше и больше инсулина в ответ на постоянно повышенный уровень глюкозы в крови. В конце концов, остальная часть тела становится резистентной к инсулину, который вырабатывает поджелудочная железа, что требует большего количества инсулина для достижения того же эффекта снижения уровня глюкозы в крови и вынуждает поджелудочную железу вырабатывать еще больше инсулина, чтобы конкурировать с резистентностью. Эта негативная спираль способствует выгоранию поджелудочной железы и прогрессированию диабета.
Чтобы контролировать реакцию организма на сахароснижающую терапию, можно измерить уровень глюкозы. Мониторинг уровня глюкозы в крови может осуществляться несколькими методами, например, с помощью теста на глюкозу натощак, который измеряет уровень глюкозы в крови после 8 часов голодания. Другой тест — это 2-часовой тест на толерантность к глюкозе (GTT). Для этого теста человек проходит тест на глюкозу натощак, затем выпивает 75-граммовый напиток с глюкозой и проходит повторное тестирование. Этот тест измеряет способность организма человека перерабатывать глюкозу. Со временем уровень глюкозы в крови должен снизиться, поскольку инсулин позволяет ему поглощаться клетками и выходить из кровотока.
Лечение гипогликемии

Люди с диабетом или другими заболеваниями, которые приводят к низкому уровню сахара в крови, часто имеют при себе небольшое количество сахара в различных формах. Одним из наиболее часто используемых видов сахара является глюкоза, часто в форме таблеток глюкозы (глюкоза, спрессованная в форму таблетки, иногда с одним или несколькими другими ингредиентами в качестве связующего вещества), леденца или пакета сахара .
Источники [ править ]

Большинство пищевых углеводов содержат глюкозу либо в качестве единственного строительного блока (как в полисахаридах, крахмале и гликогене), либо вместе с другим моносахаридом (как в гетерополисахаридах, сахарозе и лактозе). [129] Несвязанная глюкоза является одним из основных ингредиентов меда. Глюкоза чрезвычайно распространена и была выделена из различных природных источников по всему миру, включая мужские шишки хвойного дерева Wollemia nobilis в Риме, [130] корни растений Ilex asprella в Китае, [131] и соломка из риса в Калифорнии. [132]
| Еда элемент | Углеводы, общий, [а] включая пищевые волокна | Общий сахар | Бесплатно фруктоза | Бесплатно глюкоза | Сахароза | Соотношение фруктоза/ глюкоза | Сахароза как доля общее количество сахаров (%) |
|---|---|---|---|---|---|---|---|
| Фрукты | |||||||
| Яблоко | 13.8 | 10.4 | 5.9 | 2.4 | 2.1 | 2.0 | 19.9 |
| Абрикос | 11.1 | 9.2 | 0.9 | 2.4 | 5.9 | 0.7 | 63.5 |
| Банан | 22.8 | 12.2 | 4.9 | 5.0 | 2.4 | 1.0 | 20.0 |
| Инжир , сушеный | 63.9 | 47.9 | 22.9 | 24.8 | 0.9 | 0.93 | 0.15 |
| Виноград | 18.1 | 15.5 | 8.1 | 7.2 | 0.2 | 1.1 | 1 |
| Пупок оранжевый | 12.5 | 8.5 | 2.25 | 2.0 | 4.3 | 1.1 | 50.4 |
| Персик | 9.5 | 8.4 | 1.5 | 2.0 | 4.8 | 0.9 | 56.7 |
| Груша | 15.5 | 9.8 | 6.2 | 2.8 | 0.8 | 2.1 | 8.0 |
| Ананас | 13.1 | 9.9 | 2.1 | 1.7 | 6.0 | 1.1 | 60.8 |
| слива | 11.4 | 9.9 | 3.1 | 5.1 | 1.6 | 0.66 | 16.2 |
| Овощи | |||||||
| Свекла , красная | 9.6 | 6.8 | 0.1 | 0.1 | 6.5 | 1.0 | 96.2 |
| Морковь | 9.6 | 4.7 | 0.6 | 0.6 | 3.6 | 1.0 | 77 |
| Красный перец , сладкий | 6.0 | 4.2 | 2.3 | 1.9 | 0.0 | 1.2 | 0.0 |
| Лук , сладкий | 7.6 | 5.0 | 2.0 | 2.3 | 0.7 | 0.9 | 14.3 |
| Сладкий картофель | 20.1 | 4.2 | 0.7 | 1.0 | 2.5 | 0.9 | 60.3 |
| Вещи | 27.9 | 0.5 | Следы | Следы | Следы | — | Следы |
| Сахарный тростник | 13–18 | 0.2–1.0 | 0.2–1.0 | 11–16 | 1.0 | высокий | |
| Сахарная свекла | 17–18 | 0.1–0.5 | 0.1–0.5 | 16–17 | 1.0 | высокий | |
| Зерна | |||||||
| Кукуруза , сладкая | 19.0 | 6.2 | 1.9 | 3.4 | 0.9 | 0.61 | 15.0 |
- ^ Количество углеводов рассчитывается в базе данных Министерства сельского хозяйства США и не всегда соответствует сумме сахаров, крахмала и «пищевых волокон».
Коммерческое производство [ править ]
Глюкозу производят в промышленности из крахмала путем ферментативного гидролиза с помощью глюкозамилазы или с использованием кислот . Ферментативный гидролиз в значительной степени вытеснил реакции гидролиза, катализируемые кислотами. [134] В результате получается сироп глюкозы (ферментативно с содержанием глюкозы более 90% в сухом веществе). [134] с годовым мировым объемом производства 20 миллионов тонн (по состоянию на 2011 год). [135] Это причина бывшего распространенного названия «крахмальный сахар». Амилазы чаще всего происходят из Bacillus licheniformis. [136] или Bacillus subtilis (штамм MN-385), [136] которые более термостабильны, чем первоначально используемые ферменты. [136] [137] Начиная с 1982 года пуллуланазы Aspergillus niger использовались в производстве глюкозного сиропа для превращения амилопектина в крахмал (амилозу), тем самым увеличивая выход глюкозы. [138] Реакцию проводят при pH = 4,6–5,2 и температуре 55–60 °С. [11] Кукурузный сироп содержит от 20% до 95% глюкозы в сухом веществе. [139] [140] Японская форма глюкозного сиропа « Мизуаме » изготавливается из сладкого картофеля или рисового крахмала. [141] Мальтодекстрин содержит около 20% глюкозы.
Многие сельскохозяйственные культуры могут использоваться в качестве источника крахмала. кукуруза , [134] рис, [134] пшеница , [134] маниока , [134] картофель , [134] ячмень , [134] сладкий картофель, [142] кукурузная шелуха и саго используются в разных частях мира. В Штатах Соединенных кукурузный крахмал почти исключительно используется (из кукурузы). Некоторая коммерческая глюкоза встречается в составе инвертного сахара — смеси глюкозы и фруктозы в соотношении примерно 1:1, получаемой из сахарозы. В принципе, целлюлозу можно гидролизовать до глюкозы, но этот процесс пока коммерчески непрактичен. [54]
Превращение во фруктозу [ править ]
, представляющей собой смесь глюкозы и фруктозы, используется почти исключительно кукуруза (точнее, кукурузный сироп) В США в качестве источника глюкозы для производства изоглюкозы , поскольку фруктоза обладает более высокой подслащивающей способностью – при той же физиологической калорийности в 374 килокалории. за 100 г. Ежегодное мировое производство изоглюкозы составляет 8 миллионов тонн (по состоянию на 2011 год). [135] При изготовлении кукурузного сиропа конечным продуктом является кукурузный сироп с высоким содержанием фруктозы (HFCS).
Коммерческое использование [ править ]

Глюкоза в основном используется для производства фруктозы и глюкозосодержащих продуктов. В пищевых продуктах он используется в качестве подсластителя и увлажнителя для увеличения объема и создания более мягкого вкуса . [134] Различные источники глюкозы, такие как виноградный сок (для вина) или солод (для пива), используются для ферментации до этанола при производстве алкогольных напитков . В большинстве безалкогольных напитков в США используется HFCS-55 (с содержанием фруктозы 55% в сухой массе), в то время как в большинстве других подслащенных HFCS продуктов питания в США используется HFCS-42 (с содержанием фруктозы 42% в сухой массе). ). [144] В Мексике, напротив, безалкогольные напитки подслащивают тростниковым сахаром, который обладает более высокой подслащивающей способностью. [145] Кроме того, сироп глюкозы используется, в частности, при производстве кондитерских изделий, таких как конфеты , ириски и помадка . [146] Типичными химическими реакциями глюкозы при нагревании в безводных условиях являются карамелизация и, в присутствии аминокислот, реакция Майяра .
Кроме того, из глюкозы можно биотехнологически получать различные органические кислоты, например, путем ферментации с Clostridium thermoaceticum для получения уксусной кислоты , с Penicillium notatum для производства арабоаскорбиновой кислоты , с Rhizopus delemar для производства фумаровой кислоты , с Aspergillus niger для производства фумаровой кислоты. производство глюконовой кислоты , с Candida brumptii для производства изолимонной кислоты , с Aspergillus terreus для производства итаконовой кислоты , с Pseudomonas fluorescens для производства 2-кетоглюконовой кислоты , с Gluconobacter suboxydans для производства 5-кетоглюконовой кислоты , с Aspergillus oryzae для производства койевой кислоты , с Lactobacillus delbrueckii для производства молочной кислоты , с Lactobacillus brevis для производства яблочной кислоты , с Propionibacter shermanii для производства пропионовой кислоты , с Pseudomonas aeruginosa для производства пировиноградной кислоты и с Gluconobacter suboxydans для производства винной кислоты . [147] [ необходимы дополнительные ссылки ] Недавно сообщалось о мощных биоактивных натуральных продуктах, таких как триптолид, которые ингибируют транскрипцию млекопитающих посредством ингибирования субъединицы XPB общего фактора транскрипции TFIIH в качестве конъюгата глюкозы для воздействия на гипоксические раковые клетки с повышенной экспрессией переносчика глюкозы. [148] В последнее время глюкоза получила коммерческое применение в качестве ключевого компонента «наборов», содержащих молочную кислоту и инсулин, предназначенных для индукции гипогликемии и гиперлактатемии для борьбы с различными видами рака и инфекциями. [149]
Анализ [ править ]
Когда молекулу глюкозы необходимо обнаружить в определенном положении в более крупной молекуле, спектроскопию ядерного магнитного резонанса , рентгеноструктурный анализ или лектина иммуноокрашивание проводят с использованием конъюгата репортерного фермента конканавалина А , который связывает только глюкозу или маннозу.
обнаружения качественные реакции Классические
Эти реакции имеют лишь историческое значение:
Тест Фелинга [ править ]
Проба Фелинга – классический метод выявления альдоз. [150] Из-за мутаротации глюкоза всегда присутствует в небольшой степени в виде альдегида с открытой цепью. При добавлении реагентов Фелинга (раствора Фелинга (I) и раствора Фелинга (II)) альдегидная группа окисляется до карбоновой кислоты , а Cu 2+ тартратный комплекс восстанавливается до Cu + и образует осадок кирпично-красного цвета (Cu 2 O).
Прохождение теста [ править ]
В тесте Толленса после добавления аммиачного AgNO 3 к раствору образца глюкоза снижает Ag + к элементарному серебру . [151]
Тест босиком [ править ]
В Барфоеда тесте [152] раствор растворенного ацетата меди , ацетата натрия К раствору испытуемого сахара добавляют и уксусной кислоты и затем нагревают на водяной бане в течение нескольких минут. Глюкоза и другие моносахариды быстро окрашиваются в красноватый цвет и образуют красновато-коричневый оксид меди(I) (Cu 2 O).
Тест Нюландера [ править ]
Глюкоза, являясь редуцирующим сахаром, реагирует в тесте Нюландера . [153]
Другие тесты [ править ]
При нагревании разбавленного раствора гидроксида калия с глюкозой до 100°С появляется сильное красновато-коричневое окрашивание и карамельный запах. [154] Концентрированная серная кислота растворяет сухую глюкозу без почернения при комнатной температуре с образованием сахарной серной кислоты. [154] [ нужна проверка ] В дрожжевом растворе при спиртовом брожении образуется углекислый газ в соотношении 2,0454 молекулы глюкозы на одну молекулу СО 2 . [154] Глюкоза образует черную массу с хлоридом олова . [154] В аммиачном растворе серебра глюкоза (а также лактоза и декстрин) приводит к отложению серебра. В аммиачном ацетата свинца растворе гликозиды свинца , которые при варке становятся менее растворимыми и становятся коричневыми. в присутствии глюкозы образуются [154] В аммиачном растворе меди оксида меди желтого цвета, а при кипячении - красный оксид меди (то же самое с декстрином, за исключением аммиачного раствора ацетата меди). с глюкозой при комнатной температуре образуется гидрат [154] При использовании реактива Хагера глюкоза образует оксид ртути . при кипячении [154] Щелочной раствор висмута используется для осаждения элементарного черно-коричневого висмута глюкозой. [154] Глюкоза, кипяченная в растворе молибдата аммония, окрашивает раствор в синий цвет. Раствор с индигокармином и карбонатом натрия окрашивается при кипячении с глюкозой. [154]
Инструментальная количественная оценка
Рефрактометрия и поляриметрия [ править ]
В концентрированных растворах глюкозы с низкой долей других углеводов ее концентрацию можно определить поляриметром. Для сахарных смесей концентрацию можно определить с помощью рефрактометра , например, при определении Охсле в процессе производства вина.
ферментативные методы растворе в Фотометрические
Фермент глюкозооксидаза (GOx) превращает глюкозу в глюконовую кислоту и перекись водорода, потребляя при этом кислород. Другой фермент, пероксидаза, катализирует хромогенную реакцию (реакцию Триндера). [155] фенола до с 4-аминоантипирином пурпурного красителя. [156]
тест- Фотометрический полосок метод
Метод тест-полосок использует вышеупомянутое ферментативное превращение глюкозы в глюконовую кислоту с образованием перекиси водорода. Реагенты иммобилизуются на полимерной матрице, так называемой тест-полоске, которая принимает более или менее интенсивный цвет. Это можно измерить рефлектометрически при длине волны 510 нм с помощью портативного фотометра на основе светодиодов. Это позволяет обычным специалистам определять уровень сахара в крови. Помимо реакции фенола с 4-аминоантипирином разработаны новые хромогенные реакции, позволяющие проводить фотометрию при более высоких длинах волн (550 нм, 750 нм). [156] [157]
Амперометрический датчик глюкозы [ править ]
Электроанализ глюкозы также основан на упомянутой выше ферментативной реакции. Полученную перекись водорода можно количественно определить амперометрически путем анодного окисления при потенциале 600 мВ. [158] GOx иммобилизуют на поверхности электрода или в мембране, расположенной вблизи электрода. В электродах используются драгоценные металлы, такие как платина или золото, а также электроды из углеродных нанотрубок, которые, например, легированы бором. [159] Нанопроволоки Cu–CuO также используются в качестве безэнзимных амперометрических электродов, предел обнаружения которых достигает 50 мкмоль/л. [160] Особенно перспективным методом является так называемая «ферментатная проводка», при которой электрон, текущий во время окисления, переносится по молекулярной проволоке непосредственно от фермента к электроду. [161]
методы сенсорные Другие
Существует множество других химических сенсоров для измерения глюкозы. [162] [163] Учитывая важность анализа глюкозы в науках о жизни, были также разработаны многочисленные оптические зонды для сахаридов на основе использования бороновых кислот. [164] которые особенно полезны для внутриклеточных сенсорных приложений, где другие (оптические) методы недоступны или применимы лишь условно. Помимо органических производных бороновой кислоты, которые часто высокоспецифично связываются с 1,2-диольными группами сахаров, существуют также другие концепции зондов, классифицированные по функциональным механизмам, которые используют селективные глюкозосвязывающие белки (например, конканавалин А) в качестве рецептора. . Кроме того, были разработаны методы, которые косвенно определяют концентрацию глюкозы через концентрацию продуктов метаболизма, например, путем потребления кислорода с использованием флуоресцентно-оптических датчиков. [165] Наконец, существуют концепции, основанные на ферментах, которые используют собственное поглощение или флуоресценцию (флуоресцентно-меченных) ферментов в качестве репортеров. [162]
Медная йодометрия [ править ]
Глюкозу можно определить количественно с помощью йодометрии меди. [166]
Хроматографические методы [ править ]
В частности, для анализа сложных смесей, содержащих глюкозу, например, в меде, используются хроматографические методы, такие как высокоэффективная жидкостная хроматография и газовая хроматография. [166] часто используются в сочетании с масс-спектрометрией . [167] [168] Принимая во внимание соотношение изотопов, этими методами также можно надежно обнаружить фальсификацию меда добавлением сахара. [169] Обычно используется дериватизация с использованием реагентов силилирования. [170] Также можно количественно определить пропорции ди- и трисахаридов.
Анализ in vivo [ править ]
Поглощение глюкозы клетками организмов измеряют с помощью 2-дезокси-D-глюкозы или фтордезоксиглюкозы . [114] ( 18 Е)фтордезоксиглюкоза используется в качестве индикатора при позитронно-эмиссионной томографии в онкологии и неврологии, [171] где это, безусловно, наиболее часто используемый диагностический агент. [172]
Ссылки [ править ]
- ^ Номенклатура углеводов (рекомендации 1996 г.) | 2-Carb-2. Архивировано 27 августа 2023 г. в Wayback Machine . iupac.qmul.ac.uk .
- ↑ Перейти обратно: Перейти обратно: а б Боэрио-Гоутс Дж. (1991), «Измерения теплоемкости и термодинамические функции кристаллической α-D-глюкозы при температурах от 10 К до 340 К», J. Chem. Термодин. , 23 (5): 403–09, doi : 10.1016/S0021-9614(05)80128-4
- ^ Пономарев В.В., Мигарская Л.Б. (1960), "Теплоты сгорания некоторых аминокислот", Рус. Дж. Физ. хим. (англ. пер.) , 34 : 1182–83.
- ^ Домб А.Дж., Кост Дж., Уайзман Д. (4 февраля 1998 г.). Справочник по биоразлагаемым полимерам . ЦРК Пресс. п. 275. ИСБН 978-1-4200-4936-7 .
- ↑ Перейти обратно: Перейти обратно: а б «NCATS Inxight Drugs — ДЕКСТРОЗА, НЕУКАЗАННАЯ ФОРМА» . Архивировано из оригинала 11 декабря 2023 г. Проверено 18 марта 2024 г.
- ^ Камид К. (2005). Целлюлозные продукты и производные целлюлозы: молекулярная характеристика и ее применение (1-е изд.). Амстердам: Эльзевир. п. 1. ISBN 978-0-08-045444-3 . Проверено 13 мая 2021 г.
- ↑ Перейти обратно: Перейти обратно: а б с д «L-глюкоза» . Статьи, учебные пособия и онлайн-словари по биологии . 07.10.2019. Архивировано из оригинала 25 мая 2022 г. Проверено 6 мая 2022 г.
- ↑ Перейти обратно: Перейти обратно: а б Всемирная организация здравоохранения (2019). Модельный список основных лекарственных средств Всемирной организации здравоохранения: 21-й список 2019 г. Женева: Всемирная организация здравоохранения. hdl : 10665/325771 . ВОЗ/MVP/EMP/IAU/2019.06. Лицензия: CC BY-NC-SA 3.0 IGO.
- ^ «Онлайн-этимологический словарь» . Этимонлайн.com . Архивировано из оригинала 26 ноября 2016 г. Проверено 25 ноября 2016 г.
- ^ Тенар, Гей-Люссак, Био и Дюма (1838) «Отчет о мемуарах М. Пелижио, озаглавленных: Исследования природы и химических свойств сахаров» . Архивировано 6 декабря 2015 г. в Wayback Machine (Отчет о мемуарах г-на Пелижио под названием: Исследования природы и химических свойств сахаров), Cadeaux , 7 : 106–113. Со страницы 109 . Архивировано 6 декабря 2015 г. в Wayback Machine : «Из сравнений, проведенных г-ном Пелиго, следует, что виноградный сахар, крахмальный сахар, диабетический сахар и медовый сахар имеют совершенно одинаковый состав и одинаковые свойства и составляют единое тело, которое мы предлагаем называть Глюкозу (1...(1) γλευχος, сусло, сладкое вино». Из сравнений, сделанных г-ном Пелиго, следует, что сахар из винограда, сахар из крахмала, сахар из диабета и сахар из меда имеют совершенно одинаковый состав и одинаковые свойства и составляют одно и то же вещество, которое мы предлагаем назвать глюкозой ( 1) ... (1) γλευχος, сусло, сладкое вино.
- ↑ Перейти обратно: Перейти обратно: а б Энциклопедия продуктов питания и здоровья . Академическая пресса. 2015. с. 239. ИСБН 978-0-12-384953-3 . Архивировано из оригинала 23 февраля 2018 г.
- ^ Маргграф (1747) «Химические эксперименты, проведенные с целью извлечения настоящего сахара из различных растений, растущих в наших странах». Архивировано 24 июня 2016 г. в Wayback Machine [Химические эксперименты, проведенные с целью извлечения настоящего сахара из различных растений. которые растут на наших землях], История Королевской академии наук и изящной словесности Берлина , стр. 79–90. Со страницы 90: Архивировано 27 октября 2014 г. в Wayback Machine. «Сушеный виноград, смоченный небольшим количеством воды, чтобы он стал мягким, затем можно раздавить, а выжатый сок очистить и сгустить. обеспечит тип сахара». (Виноград, смоченный небольшим количеством воды, чтобы он стал мягче, можно затем отжать, и выжатый сок, [после] очистки и сгущения, даст своего рода сахар.)
- ^ Джон Ф. Робит: Основы химии углеводов. Springer Science & Business Media, 2012, ISBN 978-1-461-21622-3 . п. 7.
- ^ Розанов М.А. (1906). «О классификации стереоизомеров Фишера.1» . Журнал Американского химического общества . 28 : 114–121. дои : 10.1021/ja01967a014 . Архивировано из оригинала 17 декабря 2019 г. Проверено 1 июля 2019 г.
- ^ Эмиль Фишер , Нобелевский фонд, архивировано из оригинала 3 сентября 2009 г. , получено 2 сентября 2009 г.
- ^ Fraser-Reid B, «Глюкоза Вант-Хоффа», Chem. англ. Новости , 77 (39): 8
- ^ «Отто Мейерхоф - Факты - NobelPrize.org» . Архивировано 15 июля 2018 г. в Wayback Machine . NobelPrize.org . Проверено 5 сентября 2018 г.
- ^ «Ганс фон Эйлер-Хельпин - Факты - NobelPrize.org». Архивировано 3 сентября 2018 г. в Wayback Machine . NobelPrize.org . Проверено 5 сентября 2018 г.
- ^ «Артур Харден - Факты - NobelPrize.org» . Архивировано 3 сентября 2018 г. в Wayback Machine . NobelPrize.org . Проверено 5 сентября 2018 г.
- ^ «Бернардо Уссей - Факты - NobelPrize.org» . Архивировано 15 июля 2018 г. в Wayback Machine . NobelPrize.org . Проверено 5 сентября 2018 г.
- ^ «Карл Кори - Факты - NobelPrize.org» . Архивировано 15 июля 2018 г. в Wayback Machine . NobelPrize.org . Проверено 5 сентября 2018 г.
- ^ «Герти Кори - Факты - NobelPrize.org» . Архивировано 15 июля 2018 г. в Wayback Machine . NobelPrize.org . Проверено 5 сентября 2018 г.
- ^ «Луис Лелуар - Факты - NobelPrize.org» . Архивировано 15 июля 2018 г. в Wayback Machine . NobelPrize.org . Проверено 5 сентября 2018 г.
- ^ Венюэ Кан и Чжицзюнь Чжан (2020): «Селективное производство уксусной кислоты посредством каталитического быстрого пиролиза гексоз над калийными солями», Катализаторы , том 10, страницы 502–515. дои : 10.3390/catal10050502
- ^ Бош Л., Файлс Т., Джеймс Т. (2004). «Бинарные и тройные комплексы фенилборной кислоты с сахаридами и основаниями Льюиса» . Тетраэдр . 60 (49): 11175–11190. дои : 10.1016/j.tet.2004.08.046 . ISSN 0040-4020 .
- ^ Йебра-Бюррун М (2005), «Подсластители», Энциклопедия аналитической науки , Elsevier, стр. 562–572, doi : 10.1016/b0-12-369397-7/00610-5 , ISBN 978-0-12-369397-6
- ^ «Глюкоза». Энциклопедия Колумбии, 6-е изд., 2015. Encyclepedia.com. 17 ноября 2015 г. http://www.encyclepedia.com. Архивировано 26 апреля 2009 г. в Wayback Machine .
- ^ Ага МБ, Шарма В., Дар А.Х., Даш К.К., Сингх А., Шамс Р., Хан С.А. (2023). «Комплексный обзор функциональных и нутрицевтических свойств меда» . Еда . 4 (2). дои : 10.1002/efd2.71 .
- ^ Бобиш О, Дезмирян Д.С., Моис А.Р. (2018). «Мед и диабет: важность натуральных простых сахаров в рационе для профилактики и лечения различных типов диабета» . Окислительная медицина и клеточное долголетие . 2018 : 1–12. дои : 10.1155/2018/4757893 . ПМК 5817209 . ПМИД 29507651 .
- ^ Вани Х.А., Маджид С., Хан М.С., Бхат А.А., Вани Р.А., Бхат С.А., Али С., Рехман М.У. (2020). «Применение меда при диабете и нарушениях обмена веществ» . Терапевтическое применение меда и его фитохимических веществ . Спрингер. стр. 195–217. дои : 10.1007/978-981-15-7305-7_9 . ISBN 978-981-15-7304-0 . Архивировано из оригинала 14 апреля 2021 г. Проверено 18 марта 2024 г.
- ^ Альварес-Суарес Х.М., Тулипани С., Романдини С., Бертоли Э., Баттино М. (2010). «Вклад меда в питание и здоровье человека: обзор» . Средиземноморский журнал питания и обмена веществ . 3 : 15–23. дои : 10.1007/s12349-009-0051-6 . Архивировано из оригинала 12 февраля 2024 г. Проверено 18 марта 2024 г.
- ^ Ищаек Дж.И., Керн М. (2006). «Американский мед с разным содержанием глюкозы и фруктозы имеет одинаковые гликемические индексы - Журнал Американской диетической ассоциации» . Журнал Американской диетической ассоциации . 106 (8): 1260–1262. дои : 10.1016/j.jada.2006.05.003 . ПМИД 16863724 .
- ^ «Потенциально важный вклад декстрозы, используемой в качестве разбавителя, в гипергликемию у госпитализированных пациентов | Лечение диабета | Американская диабетическая ассоциация» . Архивировано из оригинала 29 мая 2022 г. Проверено 18 марта 2024 г.
- ^ «Декстроза: почему она содержится в продуктах питания и лекарствах?» . 24 июня 2018 г. Архивировано из оригинала 13 февраля 2024 г. Проверено 18 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б с «Что такое декстроза, как она используется и полезна ли она для здоровья? - The Nutrition Insider» . 27 октября 2023 года. Архивировано из оригинала 14 февраля 2024 года . Проверено 18 марта 2024 г.
- ^ «Декстроза против глюкозы: равны ли эти сахара?» . Архивировано из оригинала 29 сентября 2023 г. Проверено 18 марта 2024 г.
- ^ Барон Д.Н., Макинтайр Н. (1976). «Письмо: Глюкоза — это декстроза — это глюкоза» . Британский медицинский журнал . 2 (6026): 41–42. дои : 10.1136/bmj.2.6026.41-c . ПМЦ 1687736 . ПМИД 938892 .
- ↑ Перейти обратно: Перейти обратно: а б с д «Пракаш Кемикалс Интернэшнл» . Архивировано из оригинала 6 июня 2023 г. Проверено 18 марта 2024 г.
- ^ «АФИ | моногидрат глюкозы» . Архивировано из оригинала 24 марта 2023 г. Проверено 18 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б с д «Разница между безводной декстрозой и моногидратом декстрозы» . 28 декабря 2022 года. Архивировано из оригинала 18 марта 2024 года . Проверено 18 марта 2024 г.
- ^ «Декстроза безводная» . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ Хворова Л.С., Андреев Н.Р., Лукин Н.Д. (2020). «Изучение условий применения поверхностно-активных веществ в производстве кристаллической глюкозы» . Российские сельскохозяйственные науки . 46 (1): 90–93. Бибкод : 2020РуАгС..46...90К . дои : 10.3103/S1068367420010048 . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б «Чем отличается безводная глюкоза от глюкозы» . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ «Безводный и моногидрат – в чем разница?» . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ Траси Н.С., Boerrigter SX, Бирн С.Р., Карвахал ТМ (15 марта 2011 г.). «Исследование влияния условий дегидратации на компактность глюкозы — ScienceDirect» . Международный фармацевтический журнал . 406 (1): 55–61. doi : 10.1016/j.ijpharm.2010.12.042 . ПМИД 21232587 . Архивировано из оригинала 18 марта 2024 года . Проверено 18 марта 2024 г.
- ^ Митра Б., Вулф С., Ву С. (2018). «Моногидрат декстрозы в качестве альтернативного разбавителя неживотного происхождения в рецептурах таблеток для влажной грануляции с высоким усилием сдвига» . Разработка лекарств и промышленная фармация . 44 (5): 817–828. дои : 10.1080/03639045.2017.1414231 . ПМИД 29300107 . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ «Тесты на диабет и предиабет - NIDDK» . Архивировано из оригинала 16 декабря 2023 г. Проверено 18 марта 2024 г.
- ^ «Декстроза моногидрат» . Архивировано из оригинала 02 декабря 2023 г. Проверено 18 марта 2024 г.
- ^ «D-(+)-глюкоза моногидрат» . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ «Д-Глюкоза» . Архивировано из оригинала 15 декабря 2023 г. Проверено 18 марта 2024 г.
- ^ «Д-(+)-Глюкоза» . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ «Д-(+)-Глюкоза» . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ «Глюкоза (Декстроза)» . 2 октября 2013 г. Архивировано из оригинала 21 декабря 2023 г. Проверено 18 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б Шенк ФВ (2006). «Глюкоза и глюкозосодержащие сиропы». Энциклопедия промышленной химии Ульмана . дои : 10.1002/14356007.a12_457.pub2 . ISBN 978-3-527-30673-2 .
- ↑ Перейти обратно: Перейти обратно: а б Патрик Ф. Фокс: Advanced Dairy Chemistry, том 3: Лактоза, вода, соли и витамины , Springer, 1992. Том 3, ISBN 9780412630200 . п. 316.
- ^ Бенджамин Кабальеро, Пол Финглас, Фидель Толдра: Энциклопедия продуктов питания и здоровья . Академическое издательство (2016). ISBN 9780123849533 , Том 1, с. 76.
- ^ «16.4: Циклические структуры моносахаридов» . Химия LibreTexts . 18 июля 2014 г. Архивировано из оригинала 17 апреля 2023 г. Проверено 17 апреля 2023 г.
- ^ Такаги С., Джеффри Г.А. (1979). «1,2-О-изопропилиден-D-глюкофураноза». Acta Crystallographica Раздел B. Б35 (6): 1522–1525. Бибкод : 1979AcCrB..35.1522T . дои : 10.1107/S0567740879006968 .
- ^ Белецкий М., Эггерт Х., Кристиан Норрильд Дж. (1999). «Флуоресцентный датчик глюкозы, ковалентно связывающийся со всеми пятью гидроксильными группами α-D-глюкофуранозы. Повторное исследование». Журнал Химического общества, Perkin Transactions . 2 (3): 449–456. дои : 10.1039/A808896I .
- ^ Чандран С.К., Нангия А (2006). «Модулированная кристаллическая структура (Z ′ = 2) α-d-глюкофуранозо-1,2:3,5-бис(п-толил)бороната». CrystEngComm . 8 (8): 581–585. дои : 10.1039/B608029D .
- ^ Макмерри Дж. Э. (1988), Органическая химия (2-е изд.), Brooks/Cole, p. 866, ISBN 0534079687 .
- ^ Хуаристи Э., Куэвас Дж. (1995), Аномерный эффект , CRC Press, стр. 9–10, ISBN 978-0-8493-8941-2
- ↑ Перейти обратно: Перейти обратно: а б Манфред Гессе, Герберт Мейер, Бернд Зе, Стефан Биенц, Лоран Биглер, Томас Фокс: Спектроскопические методы в органической химии . 8-е исправленное издание. Георг Тиме, 2011 г., ISBN 978-3-13-160038-7 , с. 34 (на немецком языке).
- ↑ Перейти обратно: Перейти обратно: а б с д Банн Х.Ф., Хиггинс П.Дж. (1981). «Реакция моносахаридов с белками: возможное эволюционное значение». Наука . 213 (4504): 222–24. Бибкод : 1981Sci...213..222B . дои : 10.1126/science.12192669 . ПМИД 12192669 .
- ^ Джереми М. Берг: Биохимия Страйера. Издательство Спрингер, 2017, ISBN 978-3-662-54620-8 , с. 531. (Немецкий)
- ↑ Перейти обратно: Перейти обратно: а б Гарретт Р.Х. (2013). Биохимия (5-е изд.). Бельмонт, Калифорния: Брукс/Коул, Cengage Learning. ISBN 978-1-133-10629-6 .
- ↑ Перейти обратно: Перейти обратно: а б с д Фут D, Фут JG (2011). Биохимия (4-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. ISBN 978-0-470-57095-1 .
- ^ Альберт Л. Ленинджер, Биохимия, 6-е издание , Worth Publishers Inc., 1972, ISBN 0-87901-009-6 стр. 228.
- ↑ Перейти обратно: Перейти обратно: а б «Химия для биологов: Фотосинтез» . www.rsc.org . Архивировано из оригинала 4 августа 2016 г. Проверено 5 февраля 2018 г.
- ↑ Перейти обратно: Перейти обратно: а б с Питер К. Генрих: Биохимия и патобиохимия Леффлера/Петрида. Спрингер Верлаг, 2014 г., ISBN 978-3-642-17972-3 , с. 195. (Немецкий)
- ↑ Перейти обратно: Перейти обратно: а б с д и У. Сатьянараяна: Биохимия. Elsevier Health Sciences, 2014 г., ISBN 978-8-131-23713-7 . п. 674.
- ^ Вассерман Д.Х. (2009). «Четыре грамма глюкозы» . Американский журнал физиологии. Эндокринология и обмен веществ . 296 (1): Е11–21. дои : 10.1152/ajpendo.90563.2008 . ПМК 2636990 . ПМИД 18840763 .
- ^ «Высокий уровень глюкозы в крови и осложнения диабета: накопление молекул, известных как AGE, может быть ключевым звеном» , Diabetes Forecast , Американская диабетическая ассоциация, 2010 г., ISSN 0095-8301 , заархивировано из оригинала 14 октября 2013 г. , получено в 2010 г. 05-20
- ^ Варки А., Каммингс Р.Д., Эско Дж.Д., Фриз Х.Х., Стэнли П., Бертоцци Ч.Р., Харт Г.В., Эцлер М.Э. (2009). Варки А (ред.). Основы гликобиологии (2-е изд.). Издательство Колд-Спринг-Харбор Лабораториз. ISBN 978-0-87969-770-9 . ПМИД 20301239 . Архивировано из оригинала 6 декабря 2016 г.
- ^ «Показ соединения D-глюкозы (FDB012530) - FooDB» . Архивировано из оригинала 06 декабря 2022 г. Проверено 18 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б Питер К. Генрих: Биохимия и патобиохимия Леффлера/Петрида. Спрингер Верлаг, 2014 г., ISBN 978-3-642-17972-3 , с. 404.
- ^ Гарольд А. Харпер: Медицинская биохимия. Спрингер Верлаг, 2013, ISBN 978-3-662-22150-1 , с. 641. (на немецком языке)
- ^ Навале А.М., Паранджапе А.Н. (2016). «Переносчики глюкозы: физиологические и патологические роли» . Биофизические обзоры . 8 (1): 5–9. дои : 10.1007/s12551-015-0186-2 . ПМЦ 5425736 . ПМИД 28510148 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж Питер К. Генрих: Биохимия и патобиохимия Леффлера/Петрида. Спрингер Верлаг, 2014 г., ISBN 978-3-642-17972-3 , с. 199, 200. (на немецком языке)
- ^ Торенс Б (2015). «GLUT2, чувствительность к глюкозе и гомеостаз глюкозы» (PDF) . Диабетология . 58 (2): 221–32. дои : 10.1007/s00125-014-3451-1 . ПМИД 25421524 . Архивировано (PDF) из оригинала 2 декабря 2023 г. Проверено 18 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б Питер К. Генрих: Биохимия и патобиохимия Леффлера/Петрида. Спрингер Верлаг, 2014 г., ISBN 978-3-642-17972-3 , с. 214. (на немецком языке)
- ^ Хуанг С., член парламента Чехии (2007 г.). «Переносчик глюкозы GLUT4» . Клеточный метаболизм . 5 (4): 237–52. дои : 10.1016/j.cmet.2007.03.006 . ПМИД 17403369 .
- ^ Говерс Р. (2014). Клеточная регуляция поглощения глюкозы транспортером глюкозы GLUT4 . Достижения клинической химии. Том. 66. стр. 173–240. дои : 10.1016/B978-0-12-801401-1.00006-2 . ISBN 978-0-12-801401-1 . ПМИД 25344989 .
- ^ Ву X, Freeze HH (декабрь 2002 г.). «GLUT14, дубликон GLUT3, специфически экспрессируется в семенниках как альтернативные формы сплайсинга». Геномика . 80 (6): 553–7. дои : 10.1006/geno.2002.7010 . ПМИД 12504846 .
- ^ Гецци С., Лу ДДФ, Райт Э.М. (2018). «Физиология обработки глюкозы почками посредством SGLT1, SGLT2 и GLUT2» . Диабетология . 61 (10): 2087–2097. дои : 10.1007/s00125-018-4656-5 . ПМК 6133168 . ПМИД 30132032 .
- ^ Поулсен С.Б., Фентон Р.А., Риг Т. (2015). «Котранспорт натрия-глюкозы» . Современное мнение по нефрологии и гипертонии . 24 (5): 463–9. дои : 10.1097/MNH.0000000000000152 . ПМК 5364028 . ПМИД 26125647 .
- ^ Смит, Элисон М., Зееман, Сэмюэл К., Смит, Стивен М. (2005). «Деградация крахмала». Анну. Преподобный Плант Биол . 56 : 73–98. doi : 10.1146/annurev.arplant.56.032604.144257 . ПМИД 15862090 .
- ↑ Перейти обратно: Перейти обратно: а б Лешек Шаблевский: Гомеостаз глюкозы и резистентность к инсулину. Издательство Бентам Сайенс, 2011 г., ISBN 978-1-608-05189-2 , с. 46.
- ^ Питер К. Генрих: Биохимия и патобиохимия Леффлера / Петрида. Спрингер Верлаг, 2014 г., ISBN 978-3-642-17972-3 , с. 389. (на немецком языке)
- ^ Ван Г, Кавамура К, Хатакеяма С, Таками А, Ли Х, Ван В (04 апреля 2007 г.). «Авиационное измерение органических аэрозолей над Китаем» . Экологические науки и технологии . 41 (9): 3115–3120. Бибкод : 2007EnST...41.3115W . дои : 10.1021/es062601h . ISSN 0013-936X . ПМИД 17539513 . Архивировано из оригинала 18 марта 2024 г. Проверено 9 ноября 2021 г.
- ^ Адева-Андани М.М., Перес-Фельпете Н., Фернандес-Фернандес С., Донапетри-Гарсия С., Пасос-Гарсия С. (2016). «Метаболизм глюкозы в печени человека» . Отчеты по биологическим наукам . 36 (6): e00416. дои : 10.1042/BSR20160385 . ПМЦ 5293555 . ПМИД 27707936 .
- ^ Х. Роберт Хортон, Лоуренс А. Моран, К. Грей Скримджер, Марк Д. Перри, Дж. Дэвид Роун: Биохимия . Исследования Пирсона; 4-е обновленное издание, 2008 г.; ISBN 978-3-8273-7312-0 ; п. 490–496. (Немецкий)
- ↑ Перейти обратно: Перейти обратно: а б Брайан К. Холл: Эволюция Стрикбергера. Издательство «Джонс и Бартлетт», 2013 г., ISBN 978-1-449-61484-3 , с. 164.
- ^ Джонс Дж.Г. (2016). «Печеночная глюкоза и липидный обмен» . Диабетология . 59 (6): 1098–103. дои : 10.1007/s00125-016-3940-5 . ПМИД 27048250 .
- ^ Энтнер Н., Дудоров М. (1952). «Окисление глюкозы и глюконовой кислоты Pseudomonas saccharophila» . J Биол Хим . 196 (2): 853–862. дои : 10.1016/S0021-9258(19)52415-2 . ПМИД 12981024 .
- ^ EM Ammar, X. Wang, CV Rao: Регуляция метаболизма Escherichia coli во время роста на смесях неглюкозных сахаров: арабинозы, лактозы и ксилозы. В: Научные отчеты . Группа 8, номер 1, январь 2018 г., с. 609, doi : 10.1038/s41598-017-18704-0 , PMID 29330542, PMC 5766520 .
- ↑ Перейти обратно: Перейти обратно: а б Бонадонна Р.К., Бонора Э., Дель Прато С., Саккомани М., Кобелли С., Натали А., Фрассерра С., Пекори Н., Ферраннини Е., Бир Д., ДеФронзо Р.А., Гулли Дж. (июль 1996 г.). «Роль транспорта глюкозы и фосфорилирования глюкозы в мышечной резистентности к инсулину при ИНСД» (PDF) . Диабет . 45 (7): 915–25. дои : 10.2337/диаб.45.7.915 . ПМИД 8666143 . S2CID 219249555 . Архивировано (PDF) из оригинала 06 марта 2017 г. Проверено 5 марта 2017 г.
- ^ «Глюкоза» . Архивировано из оригинала 5 декабря 2023 г. Проверено 18 марта 2024 г.
- ^ Краткий обзор медицинской биохимии @Google book , Blackwell Publishing, 2006, стр. 52, ISBN 978-1-4051-1322-9 , заархивировано из оригинала 23 февраля 2018 г.
- ^ Краткий обзор медицинской биохимии @Google book , Blackwell Publishing, 2006, стр. 50, ISBN 978-1-4051-1322-9 , заархивировано из оригинала 23 февраля 2018 г.
- ^ Аннибальди А, Видманн С (2010). «Метаболизм глюкозы в раковых клетках». Текущее мнение о клиническом питании и метаболической помощи . 13 (4): 466–70. дои : 10.1097/MCO.0b013e32833a5577 . ПМИД 20473153 . S2CID 205782021 .
- ^ Шаблевский Л (2013). «Экспрессия переносчиков глюкозы при раке». Biochimica et Biophysical Acta (BBA) - Обзоры о раке . 1835 (2): 164–9. дои : 10.1016/j.bbcan.2012.12.004 . ПМИД 23266512 .
- ^ Адекола К., Розен С.Т., Шанмугам М. (2012). «Переносчики глюкозы в метаболизме рака» . Современное мнение в онкологии . 24 (6): 650–4. дои : 10.1097/CCO.0b013e328356da72 . ПМК 6392426 . ПМИД 22913968 .
- ^ Шуман У, Грюндлер П (1998). «Электрохимическая деградация органических веществ на анодах PbO2: Мониторинг путем непрерывного измерения CO2» . Исследования воды . 32 (9): 2835–2842. дои : 10.1016/s0043-1354(98)00046-3 . ISSN 0043-1354 . Архивировано из оригинала 18 марта 2024 г. Проверено 9 ноября 2021 г.
- ^ «Глава 3: Расчет энергетического содержания пищевых продуктов – коэффициенты преобразования энергии» , Энергия пищевых продуктов – методы анализа и коэффициенты преобразования , Документ ФАО по продуктам питания и питанию 77, Рим: Продовольственная и сельскохозяйственная организация, 2003, ISBN 978-92-5-105014-9 , заархивировано из оригинала 24 мая 2010 г.
- ^ Георг Шведт: Сахарно-сладкая химия. Джон Уайли и сыновья, 2012 г., ISBN 978-3-527-66001-8 , с. 100 (на немецком языке) .
- ^ Шмидт, Ланг: Физиология человека , 30-е издание. Springer Verlag, 2007, с. 907 (на немецком языке) .
- ^ Дандекар Т., Шустер С., Снел Б., Хуйнен М., Борк П. (1999). «Выравнивание путей: применение к сравнительному анализу гликолитических ферментов» . Биохимический журнал . 343 (1): 115–124. дои : 10.1042/bj3430115 . ПМЦ 1220531 . ПМИД 10493919 .
- ^ Дэш П. «Гематоэнцефалический барьер и церебральный метаболизм (раздел 4, глава 11)» . Neuroscience Online: Электронный учебник по нейронаукам . Кафедра нейробиологии и анатомии – Медицинская школа Техасского университета в Хьюстоне. Архивировано из оригинала 17 ноября 2016 г.
- ^ Fairclough SH, Houston K (2004), «Метаболический показатель умственных усилий», Biol. Психол. , 66 (2): 177–190, doi : 10.1016/j.biopsycho.2003.10.001 , PMID 15041139 , S2CID 44500072
- ^ Gailliot MT, Baumeister RF, DeWall CN, Plant EA, Brewer LE, Schmeichel BJ, Tice DM, Maner JK (2007), «Самоконтроль зависит от глюкозы как ограниченного источника энергии: сила воли - это больше, чем метафора» (PDF) , Дж. Перс. Соц. Психол. , 92 (2): 325–336, CiteSeerX 10.1.1.337.3766 , doi : 10.1037/0022-3514.92.2.325 , PMID 17279852 , S2CID 7496171 , заархивировано (PDF) из оригинала 18 августа 2017 г.
- ^ Gailliot MT, Baumeister RF (2007), «Физиология силы воли: связь уровня глюкозы в крови с самоконтролем», Personal. Соц. Психол. Rev. , 11 (4): 303–327, CiteSeerX 10.1.1.475.9484 , doi : 10.1177/1088868307303030 , PMID 18453466 , S2CID 14380313
- ^ Масикампо Э.Дж., Баумайстер Р.Ф. (2008), «К физиологии двухпроцессного рассуждения и суждения: лимонад, сила воли и дорогой анализ, основанный на правилах», Psychol. наук. , 19 (3): 255–60, doi : 10.1111/j.1467-9280.2008.02077.x , PMID 18315798 , S2CID 38596025
- ↑ Перейти обратно: Перейти обратно: а б с Донард Дуайер: Метаболизм глюкозы в мозге. Академическое издательство, 2002, ISBN 978-0-123-66852-3 , с. XIII.
- ↑ Перейти обратно: Перейти обратно: а б с Куккук Л.Л., Мул Дж.Д., Ла Флер С.В. (2017). «Ощущение глюкозы в системе вознаграждения» . Границы в неврологии . 11 : 716. дои : 10.3389/fnins.2017.00716 . ПМЦ 5742113 . ПМИД 29311793 .
- ↑ Перейти обратно: Перейти обратно: а б Такер Р.М., Тан С.Ю. (2017). «Влияют ли некалорийные подсластители на острый гомеостаз глюкозы у людей? Систематический обзор» . Физиология и поведение . 182 : 17–26. дои : 10.1016/j.physbeh.2017.09.016 . ПМИД 28939430 . S2CID 38764657 . Архивировано из оригинала 29 июля 2020 г. Проверено 7 июня 2020 г.
- ^ Ла Флер С.Э., Флайерс Э., Калсбек А. (2014). «Нейронаука гомеостаза глюкозы». Диабет и нервная система . Справочник по клинической неврологии. Том. 126. стр. 341–351. дои : 10.1016/B978-0-444-53480-4.00026-6 . ISBN 978-0-444-53480-4 . ПМИД 25410233 . .
- ^ Бисшоп П.Х., Флайерс Э., Калсбек А. (2015). «Вегетативная регуляция продукции глюкозы печенью». Комплексная физиология . 5 (1): 147–165. дои : 10.1002/cphy.c140009 . ПМИД 25589267 .
- ^ WA Scherbaum, BM Lobnig, В: Ханс-Петер Вольф, Томас Р. Вейраух: Внутренняя терапия 2006, 2007 . 16-е издание. Эльзевир, 2006, ISBN 3-437-23182-0 , с. 927, 985 (на немецком языке) .
- ^ Гарольд А. Харпер: Медицинская биохимия . Спрингер Верлаг, 2013, ISBN 978-3-662-22150-1 , с. 294.
- ^ Кларк С.Ф., Фостер-младший (2012). «История глюкометров и их роль в самоконтроле сахарного диабета». Британский журнал биомедицинской науки . 69 (2): 83–93. CiteSeerX 10.1.1.468.2196 . дои : 10.1080/09674845.2012.12002443 . ПМИД 22872934 . S2CID 34263228 .
- ^ «Диагностика диабета и изучение предиабета» . Американская Диабетическая Ассоциация . Архивировано из оригинала 28 июля 2017 г. Проверено 20 февраля 2018 г.
- ↑ Перейти обратно: Перейти обратно: а б Ричард А. Харви, Дениз Р. Ферье: Биохимия . 5-е издание, Липпинкотт Уильямс и Уилкинс, 2011 г., ISBN 978-1-608-31412-6 , с. 366.
- ↑ Перейти обратно: Перейти обратно: а б У Сатьянараяна: Биохимия . Elsevier Health Sciences, 2014 г., ISBN 978-8-131-23713-7 , с. 508.
- ^ Холт С.Х., Миллер Дж.К., Петоч П. (1997). «Инсулиновый индекс пищевых продуктов: потребность в инсулине, создаваемая порциями обычных продуктов питания массой 1000 кДж» . Американский журнал клинического питания . 66 (5): 1264–1276. дои : 10.1093/ajcn/66.5.1264 . ПМИД 9356547 .
- ↑ Перейти обратно: Перейти обратно: а б Питер К. Генрих: Биохимия и патобиохимия Леффлера/Петрида. Спрингер Верлаг, 2014 г., ISBN 978-3-642-17972-3 , с. 27. (на немецком языке)
- ^ Рёдер П.В., Ву Б, Лю Ю, Хан В (2016). «Поджелудочная регуляция гомеостаза глюкозы» . Эксп. Мол. Мед . 48 (3 марта): e219–. дои : 10.1038/emm.2016.6 . ПМЦ 4892884 . ПМИД 26964835 .
- ^ Эстела, Карлос (2011) «Уровни глюкозы в крови», Студентский журнал математического моделирования: один + два: Том. 3: Вып. 2, статья 12.
- ^ «Углеводы и уровень сахара в крови» . Источник питания . 05.08.2013. Архивировано из оригинала 30 января 2017 г. Получено 30 января 2017 г. - через Гарвардскую школу общественного здравоохранения им. Т.Ч. Чана.
- ^ Вендитти А, Фрецца С, Винченти Ф, Броделла А, Скиубба Ф, Монтесано С, Франческин М, Сержи М, Фоддай С, Ди Кокко МЭ, Курини Р (2019). «Производное син-энт-лабдадиена с редкой функцией спиро-β-лактона из мужских шишек Wollemia nobilis» . Фитохимия . 158 : 91–95. Бибкод : 2019PChem.158...91В . doi : 10.1016/j.phytochem.2018.11.012 . ISSN 0031-9422 . ПМИД 30481664 . S2CID 53757166 . Архивировано из оригинала 18 марта 2024 г. Проверено 11 ноября 2021 г.
- ^ Лей Ю, Ши СП, Сон ЮЛ, Би Д, Ту ПФ (2014). «Тритерпеновые сапонины из корней Ilex asprella » . Химия и биоразнообразие . 11 (5): 767–775. дои : 10.1002/cbdv.201300155 . ISSN 1612-1872 . ПМИД 24827686 . S2CID 40353516 . Архивировано из оригинала 18 марта 2024 г. Проверено 11 ноября 2021 г.
- ^ Балан В., Балс Б., Чундават С.П., Маршалл Д., Дейл Б.Е. (2009), «Предварительная обработка лигноцеллюлозной биомассы с использованием AFEX», Биотопливо , Методы молекулярной биологии, том. 581, Тотова, Нью-Джерси: Humana Press, стр. 61–77, Bibcode : 2009biof.book...61B , doi : 10.1007/978-1-60761-214-8_5 , ISBN 978-1-60761-213-1 , PMID 19768616 , заархивировано из оригинала 18 марта 2024 г. , получено 11 ноября 2021 г.
- ^ «ФудДата Централ» . fdc.nal.usda.gov . Архивировано из оригинала 3 декабря 2019 г. Проверено 18 марта 2024 г.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я PJ Fellows: Технология пищевой промышленности. Издательство Вудхед , 2016, ISBN 978-0-081-00523-1 , с. 197.
- ↑ Перейти обратно: Перейти обратно: а б Томас Беккер, Дитмар Брайтаупт, Хорст Вернер Дёлле, Армин Фихтер, Гюнтер Шлегель, Сакаю Симидзу, Хидеаки Ямада: Биотехнология , в: Энциклопедия промышленной химии Ульмана , 7-е издание, Wiley-VCH, 2011. ISBN 978-3-527-32943-4 . Том 6, с. 48.
- ↑ Перейти обратно: Перейти обратно: а б с Японское общество исследования амилаз: Справочник по амилазам и родственным ферментам. Эльзевир, 2014, ISBN 978-1-483-29939-6 , с. 195.
- ^ Мэдсен ГБ, Норман Б.Е., Слотт С. (1973). «Новая термостабильная бактериальная амилаза и ее использование при высокотемпературном сжижении». Крахмал – Старке . 25 (9): 304–308. дои : 10.1002/star.19730250906 .
- ^ Норман Б.Е. (1982). «Новый разветвляющий фермент для применения в производстве глюкозных сиропов». Крахмал – Старке . 34 (10): 340–346. дои : 10.1002/star.19820341005 .
- ^ Джеймс Н. БеМиллер, Рой Л. Уистлер (2009). Крахмал: химия и технология . Пищевая наука и технология (3-е изд.). Нью-Йорк: Академическая пресса. ISBN 978-0-08-092655-1 .
- ^ БеМиллер, Джеймс Н., Уистлер, Рой Л., ред. (2009). Крахмал: химия и технология . Пищевая наука и технология (3-е изд.). Нью-Йорк: Академическая пресса. ISBN 978-0-08-092655-1 . Проверено 25 ноября 2016 г. .
- ^ Алан Дэвидсон: Оксфордский компаньон по еде (1999). «Мизуаме», с. 510 ISBN 0-19-211579-0 .
- ^ Алан Дэвидсон: Оксфордский справочник по еде . ОУП Оксфорд, 2014 г., ISBN 978-0-191-04072-6 , с. 527.
- ^ «Сахар» . Обучение, Продовольственные ресурсы. food.oregonstate.edu . Университет штата Орегон , Корваллис, Орегон. 23 мая 2012 г. Архивировано из оригинала 18 июля 2011 г. Проверено 28 июня 2018 г.
- ^ «Кукурузный сироп с высоким содержанием фруктозы: вопросы и ответы» . Управление по контролю за продуктами и лекарствами США. 05.11.2014. Архивировано из оригинала 25 января 2018 г. Проверено 18 декабря 2017 г.
- ↑ Кевин Пэнг: Мексиканская кока-кола стала хитом в США. В: Seattle Times , 29 октября 2004 г.
- ^ Стив Т. Беккет: Промышленное производство и использование шоколада Беккета . Джон Уайли и сыновья, 2017, ISBN 978-1-118-78014-5 , с. 82.
- ^ Джеймс А. Кент: Справочник Ригеля по промышленной химии . Springer Science & Business Media, 2013, ISBN 978-1-475-76431-4 , с. 938.
- ^ Датан Э, Минн И, Пэн X, Хэ QL, Ан Х, Ю Б, Помпер МГ, Лю Джо (2020). «Конъюгат глюкозы-триптолида избирательно воздействует на раковые клетки в условиях гипоксии» . iScience . 23 (9): 101536. Бибкод : 2020iSci...23j1536D . дои : 10.1016/j.isci.2020.101536 . ПМЦ 7509213 . ПМИД 33083765 .
- ^ Гудвин М.Л., Глэдден Л.Б., Нейстен М.В. (3 сентября 2020 г.). «Лактат-защищенная гипогликемия (ЛПГ)» . Границы в неврологии . 14 : 920. дои : 10.3389/fnins.2020.00920 . ISSN 1662-453X . ПМЦ 7497796 . ПМИД 33013305 .
- ^ Х. Фелинг: Количественное определение сахара в моче . В: Архив физиологической медицины (1848), том 7, с. 64–73 (на немецком языке).
- ^ Б. Толленс: О аммонийно-щелочном растворе серебра в качестве реагента для определения альдегида. Архивировано 19 февраля 2022 г. в Wayback Machine . В «Отчетах Немецкого химического общества» (1882 г.), том 15, с. 1635–1639 (на немецком языке).
- ^ Барфоед С. (1873 г.). «Об обнаружении глюкозы рядом с декстрином и родственными ему тельцами» . Журнал аналитической химии (на немецком языке). 12 :27-32. дои : 10.1007/BF01462957 . S2CID 95749674 . Архивировано из оригинала 29 июля 2020 г. Проверено 1 июля 2019 г.
- ^ Эмиль Нюландер: О щелочном растворе висмута как реагенте для определения глюкозы в моче , Журнал физиологической химии . Том 8, выпуск 3, 1884 г., с. 175-185 Аннотация . Архивировано 23 сентября 2015 г. в Wayback Machine (на немецком языке).
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я Георг Шведт: Сахарно-сладкая химия . Джон Уайли и сыновья, 2012 г., ISBN 978-3-527-66001-8 , с. 102 (на немецком языке).
- ^ Триндер П. (1969). «Определение глюкозы в крови с помощью глюкозооксидазы с альтернативным акцептором кислорода» . Анналы клинической биохимии . 6 : 24–27. дои : 10.1177/000456326900600108 . S2CID 58131350 .
- ↑ Перейти обратно: Перейти обратно: а б Чжан Ц, Чжао Г, Ян Н, Чжан Л (2019). «Уровень глюкозы в крови натощак у больных с различными видами заболеваний» . Часть А. Гликаны и гликозаминогликаны как клинические биомаркеры и терапевтические средства . Прогресс молекулярной биологии и трансляционной науки. Том. 162. стр. 277–292. дои : 10.1016/bs.pmbts.2019.01.004 . ISBN 978-0-12-817738-9 . ПМИД 30905457 . Архивировано из оригинала 18 марта 2024 г. Проверено 18 марта 2024 г.
- ^ Мидзогучи М., Исияма М., Сига М. (1998). «Водорастворимый хромогенный реагент для колориметрического обнаружения перекиси водорода - альтернатива 4-аминоантипирину, работающему на длинной волне». Аналитические коммуникации . 35 (2): 71–74. дои : 10.1039/A709038B .
- ^ Ван Дж (2008). «Электрохимические биосенсоры глюкозы». Химические обзоры . 108 (2): 814–825. дои : 10.1021/cr068123a . ПМИД 18154363 . .
- ^ Чен X, Чен J, Дэн C, Сяо C, Ян Y, Не Z, Яо С (2008). «Амперометрический биосенсор глюкозы на основе модифицированного электрода из углеродных нанотрубок, легированных бором». Таланта . 76 (4): 763–767. дои : 10.1016/j.talanta.2008.04.023 . ПМИД 18656655 .
- ^ Ван Г, Вэй Ю, Чжан В, Чжан Икс, Фан Б, Ван Л (2010). «Безферментное амперометрическое определение глюкозы с использованием композитов нанопроволок Cu-CuO». Микрохимика Акта . 168 (1–2): 87–92. дои : 10.1007/s00604-009-0260-1 . S2CID 98567636 .
- ^ Охара Т.Дж., Раджагопалан Р., Хеллер А. (1994). « Проводные» ферментные электроды для амперометрического определения глюкозы или лактата в присутствии мешающих веществ». Аналитическая химия . 66 (15): 2451–2457. дои : 10.1021/ac00087a008 . ПМИД 8092486 .
- ↑ Перейти обратно: Перейти обратно: а б Борисов С.М., Вольфбайс О.С. (2008). «Оптические биосенсоры». Химические обзоры . 108 (2): 423–461. дои : 10.1021/cr068105t . ПМИД 18229952 .
- ^ Ферри С., Кодзима К., Соде К. (2011). «Обзор глюкозооксидаз и глюкозодегидрогеназ: взгляд на ферменты, чувствительные к глюкозе», с высоты птичьего полета . Журнал науки и технологий о диабете . 5 (5): 1068–76. дои : 10.1177/193229681100500507 . ПМЦ 3208862 . ПМИД 22027299 .
- ^ Мадер Х.С., Wolfbeis OS (2008). «Зонды на основе бороновой кислоты для микроопределения сахаридов и гликозилированных биомолекул». Микрохимика Акта . 162 (1–2): 1–34. дои : 10.1007/s00604-008-0947-8 . S2CID 96768832 .
- ^ Вольфбайс О.С., Эме И., Папковская Н., Климант И. (2000). «Золь-гель биосенсоры глюкозы с использованием оптических преобразователей кислорода и метод компенсации переменного кислородного фона». Биосенсоры и биоэлектроника . 15 (1–2): 69–76. дои : 10.1016/S0956-5663(99)00073-1 . ПМИД 10826645 .
- ↑ Перейти обратно: Перейти обратно: а б Галант А.Л., Кауфман Р.К., Уилсон Дж.Д. (2015). «Глюкоза: обнаружение и анализ». Пищевая химия . 188 : 149–160. doi : 10.1016/j.foodchem.2015.04.071 . ПМИД 26041177 .
- ^ Санс М.Л., Санс Дж., Мартинес-Кастро I (2004 г.). «Газохроматографо-масс-спектрометрический метод качественного и количественного определения дисахаридов и трисахаридов в меде». Журнал хроматографии А. 1059 (1–2): 143–148. дои : 10.1016/j.chroma.2004.09.095 . ПМИД 15628134 .
- ^ Институт молекулярной физиологии растений Макса Планка в базе данных Гольма (19 июля 2007 г.). «Масс-спектр глюкозы» . База данных метаболомов Голма . Архивировано из оригинала 9 сентября 2018 г. Проверено 4 июня 2018 г.
- ^ Кабаньеро А.И., Ресио Х.Л., Руперес М. (2006). «Жидкостная хроматография в сочетании с масс-спектрометрией изотопного соотношения: новый взгляд на обнаружение фальсификации меда». J Agric Food Chem . 54 (26): 9719–9727. дои : 10.1021/jf062067x . ПМИД 17177492 .
- ^ Беккер М., Либнер Ф., Розенау Т., Поттаст А. (2013). «Подход этоксимации-силилирования для анализа моно- и дисахаридов и характеристики параметров их идентификации с помощью ГХ/МС». Таланта . 115 : 642–51. дои : 10.1016/j.talanta.2013.05.052 . ПМИД 24054643 .
- ↑ Общество немецких химиков : wayback=20100331071121 Приложения к позиционному документу Отдела ядерной химии. Архивировано 31 марта 2010 г. в Wayback Machine , февраль 2000 г.
- ^ Машауэр С., Пранте О (2014). «Подслащивание фармацевтической радиохимии путем (18) f-фторгликозилирования: краткий обзор» . БиоМед Исследования Интернэшнл . 2014 : 1–16. дои : 10.1155/2014/214748 . ПМЦ 4058687 . ПМИД 24991541 .










Виды углеводов |
|---|
Сахар как пищевой продукт |
|---|