Фосфоглицерат киназа
| Фосфоглицерат киназа | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 2.7.2.3 | ||
| Номер CAS. | 9001-83-6 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| Фосфоглицерат киназа | |||
|---|---|---|---|
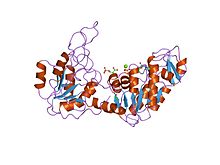 Структура дрожжевой фосфоглицераткиназы. [1] | |||
| Идентификаторы | |||
| Символ | ПГК | ||
| Пфам | PF00162 | ||
| ИнтерПро | ИПР001576 | ||
| PROSITE | PDOC00102 | ||
| СКОП2 | 3пгк / СКОПе / СУПФАМ | ||
| |||
Фосфоглицераткиназа ( EC 2.7.2.3 ) (PGK 1) представляет собой фермент , который катализирует обратимый перенос фосфатной группы от 1,3-бисфосфоглицерата (1,3-BPG) к ADP с образованием 3-фосфоглицерата (3-PG) и АТФ. :
- 1,3-бисфосфоглицерат + АДФ ⇌ глицерат-3-фосфат + АТФ.
Как и все киназы, это трансфераза . PGK является основным ферментом, используемым в гликолизе , на первом этапе гликолитического пути, генерирующего АТФ. При глюконеогенезе реакция, катализируемая ПГК, протекает в противоположном направлении, образуя АДФ и 1,3-БФГ.
У человека два изозима к настоящему времени идентифицированы PGK: PGK1 и PGK2. Изозимы имеют идентичность аминокислотных последовательностей на 87-88% и, хотя они структурно и функционально схожи, имеют разную локализацию: PGK2, кодируемый аутосомным геном, уникален для мейотических и постмейотических сперматогенных клеток, тогда как PGK1, кодируемый X -хромосома , повсеместно экспрессируется во всех клетках. [2]
Биологическая функция
[ редактировать ]PGK присутствует во всех живых организмах как один из двух ферментов, генерирующих АТФ при гликолизе. В глюконеогенном пути PGK катализирует обратную реакцию. В стандартных биохимических условиях предпочтение отдается гликолитическому направлению. [1]
В цикле Кальвина у фотосинтезирующих организмов PGK катализирует фосфорилирование 3-PG с образованием 1,3-BPG и ADP в рамках реакций, регенерирующих рибулозо-1,5-бисфосфат .
Сообщалось, что PGK проявляет тиолредуктазную активность в отношении плазмина , что приводит к образованию ангиостатина , который ингибирует ангиогенез и рост опухоли . Было также показано, что фермент участвует в репликации и репарации ДНК в ядрах клеток млекопитающих . [3]
Было показано, что человеческий изофермент PGK2, который экспрессируется только во время сперматогенеза, необходим для функции сперматозоидов у мышей. [4]
Интерактивная карта маршрутов
[ редактировать ]Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [§ 1]

- ^ Интерактивную карту маршрутов можно редактировать на WikiPathways: «Гликолиз-Глюконеогенез_WP534» .
Структура
[ редактировать ]Обзор
[ редактировать ]PGK обнаружен во всех живых организмах, и его последовательность высоко консервативна на протяжении всей эволюции. Фермент существует в виде из 415 остатков мономера , содержащего два домена почти одинакового размера, которые соответствуют N- и C-концам белка. [5] 3-фосфоглицерат (3-PG) связывается с N-концевым доменом, тогда как нуклеотидные субстраты, MgATP или MgADP, связываются с C-концевым доменом фермента. Эта расширенная двухдоменная структура связана с крупномасштабными конформационными изменениями «шарнирного изгиба», подобными тем, которые обнаружены у гексокиназы . [6] Два домена белка разделены щелью и соединены двумя альфа-спиралями . [2] В ядре каждого домена находится 6-нитевой параллельный бета-лист, окруженный альфа-спиралями. Две доли способны сворачиваться независимо, что согласуется с наличием промежуточных продуктов на пути сворачивания с одним свернутым доменом. [7] [8] Хотя связывание любого субстрата вызывает конформационные изменения , только за счет связывания обоих субстратов происходит закрытие домена, что приводит к переносу фосфатной группы. [2]
Фермент имеет тенденцию существовать в открытой конформации с короткими периодами закрытия и катализа, что обеспечивает быструю диффузию субстрата и продуктов через места связывания; открытая конформация PGK более конформационно стабильна из-за обнажения гидрофобной области белка при закрытии домена. [7]
Роль магния
[ редактировать ]Ионы магния обычно образуют комплексы с фосфатными группами – нуклеотидными субстратами PGK. Известно, что в отсутствие магния ферментативная активность не возникает. [9] Двухвалентный металл ферментов помогает лигандам экранировать отрицательные заряды связанной фосфатной группы, позволяя нуклеофильной атаке произойти ; эта стабилизация заряда является типичной характеристикой реакции фосфопереноса. [10] Предполагается, что ион может также способствовать закрытию домена, когда PGK связала оба субстрата. [9]
Механизм
[ редактировать ]
Без связи с каким-либо субстратом PGK существует в «открытой» конформации . После того, как триозный и нуклеотидный субстраты связываются с N- и C-концевыми доменами соответственно, происходит обширное шарнирно-изгибающее движение, приводящее домены и связанные с ними субстраты в непосредственной близости и приводящее к «закрытой» конформации. [11] Затем, в случае прямой гликолитической реакции, бета-фосфат АДФ инициирует нуклеофильную атаку на 1-фосфат 1,3-БФГ. Lys219 фермента направляет фосфатную группу к субстрату.
со стабилизированным зарядом PGK протекает через переходное состояние , которое предпочтительнее расположения связанного субстрата в закрытом ферменте, поскольку в переходном состоянии все три атома кислорода фосфата стабилизируются лигандами , в отличие от только двух стабилизированных атомов кислорода в исходном связанном состоянии. . [12]
В гликолитическом пути 1,3-БФГ является донором фосфата и обладает высоким потенциалом переноса фосфорила. Катализируемый PGK перенос фосфатной группы от 1,3-BPG к ADP с образованием АТФ может привести к увеличению энергии. [ нужны разъяснения ] реакция окисления углерода на предыдущем гликолитическом этапе (превращение глицеральдегид-3-фосфата в 3-фосфоглицерат ). [ нужна ссылка ]
Регулирование
[ редактировать ]Фермент активируется низкими концентрациями различных многовалентных анионов, таких как пирофосфат, сульфат, фосфат и цитрат. Высокие концентрации MgATP и 3-PG активируют PGK, тогда как Mg2+ в высоких концентрациях неконкурентно ингибирует фермент. [13]
PGK проявляет широкую специфичность по отношению к нуклеотидным субстратам. [14] Его активность ингибируется салицилатами, которые, по-видимому, имитируют нуклеотидный субстрат фермента. [15]
Было показано, что макромолекулярная скученность увеличивает активность PGK как при компьютерном моделировании, так и в средах in vitro, имитирующих внутреннюю часть клетки; в результате скученности фермент становится более ферментативно активным и более компактным. [5]
Актуальность заболевания
[ редактировать ]Дефицит фосфоглицераткиназы (PGK) является Х-сцепленным рецессивным признаком, связанным с гемолитической анемией , психическими расстройствами и миопатией у человека. [16] [17] в зависимости от формы – существует гемолитическая форма и миопатическая форма. [18] Поскольку признак является Х-сцепленным, он обычно полностью выражен у мужчин, имеющих одну Х-хромосому; пораженные женщины обычно протекают бессимптомно. [2] [17] Это состояние возникает в результате мутаций в Pgk1, гене, кодирующем PGK1, и было идентифицировано двадцать мутаций. [17] [2] На молекулярном уровне мутация Pgk1 ухудшает термическую стабильность и ингибирует каталитическую активность фермента. [2] PGK — единственный фермент непосредственного гликолитического пути, кодируемый Х-связанным геном. При гемолитической анемии в эритроцитах возникает дефицит ПГК . В настоящее время не существует окончательного лечения дефицита PGK. [19]
Сверхэкспрессия PGK1 связана с раком желудка и, как было обнаружено, увеличивает инвазивность клеток рака желудка in vitro . [20] Фермент секретируется опухолевыми клетками и участвует в ангиогенном процессе, приводя к высвобождению ангиостатина и ингибированию роста кровеносных сосудов опухоли. [3]
Известно , что благодаря своей широкой специфичности по отношению к нуклеотидным субстратам PGK участвует в фосфорилировании и активации ВИЧ антиретровирусных препаратов против , основанных на нуклеотидах. [14] [21]
Человеческие изоферменты
[ редактировать ]
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
Ссылки
[ редактировать ]- ^ Jump up to: а б Уотсон Х.К., Уокер Н.П., Шоу П.Дж., Брайант Т.Н., Венделл П.Л., Фотергилл Л.А., Перкинс Р.Э., Конрой С.К., Добсон М.Дж., Туит М.Ф. (1982). «Последовательность и структура дрожжевой фосфоглицераткиназы» . Журнал ЭМБО . 1 (12): 1635–40. дои : 10.1002/j.1460-2075.1982.tb01366.x . ПМК 553262 . ПМИД 6765200 .
- ^ Jump up to: а б с д и ж Кьярелли Л.Р., Морера С.М., Бьянки П., Фермо Э., Занелла А., Галицци А., Валентини Г. (2012). «Молекулярные данные о патогенных эффектах мутаций, вызывающих дефицит фосфоглицераткиназы» . ПЛОС ОДИН . 7 (2): e32065. Бибкод : 2012PLoSO...732065C . дои : 10.1371/journal.pone.0032065 . ПМК 3279470 . ПМИД 22348148 .
- ^ Jump up to: а б Лэй Эй Джей, Цзян XM, Кискер О, Флинн Э, Андервуд А, Кондрон Р, Хогг П.Дж. (декабрь 2000 г.). «Фосфоглицераткиназа действует при опухолевом ангиогенезе как дисульфидредуктаза». Природа . 408 (6814): 869–73. Бибкод : 2000Natur.408..869L . дои : 10.1038/35048596 . ПМИД 11130727 . S2CID 4340557 .
- ^ Даньшина П.В., Гейер CB, Дай Кью, Гулдинг Э.Х., Уиллис В.Д., Китто ГБ, Маккерри Дж.Р., Эдди Э.М., О'Брайен Д.А. (январь 2010 г.). «Фосфоглицерат киназа 2 (PGK2) необходима для функции сперматозоидов и мужской фертильности у мышей» . Биология размножения . 82 (1): 136–45. дои : 10.1095/biolreprod.109.079699 . ПМК 2802118 . ПМИД 19759366 .
- ^ Jump up to: а б Дхар А., Самиотакис А., Эббингауз С., Ниенхаус Л., Хомуз Д., Грюбеле М., Чунг М.С. (октябрь 2010 г.). «Структура, функция и укладка фосфоглицераткиназы сильно нарушаются скученностью макромолекул» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (41): 17586–91. Бибкод : 2010PNAS..10717586D . дои : 10.1073/pnas.1006760107 . ПМК 2955104 . ПМИД 20921368 .
- ^ Кумар С., Ма Б., Цай С.Дж., Вольфсон Х., Нусинов Р. (1999). «Складчатые воронки и конформационные переходы посредством шарнирно-изгибательных движений» . Клеточная биохимия и биофизика . 31 (2): 141–64. дои : 10.1007/BF02738169 . ПМИД 10593256 . S2CID 41924983 .
- ^ Jump up to: а б Йон Дж.М., Демадрил М., Беттон Дж.М., Минард П., Баллери Н., Миссиакас Д., Гайяр-Миран С., Перахия Д., Муавад Л. (1990). «Гибкость и сворачивание фосфоглицераткиназы». Биохимия . 72 (6–7): 417–29. дои : 10.1016/0300-9084(90)90066-п . ПМИД 2124145 .
- ^ Зеррад Л., Мерли А., Шредер Г.Ф., Варга А., Грацер Э, Перно П., Раунд А, Вас М., Боулер М.В. (апрель 2011 г.). «Пружинный механизм высвобождения регулирует движение доменов и катализ фосфоглицераткиназы» . Журнал биологической химии . 286 (16): 14040–8. дои : 10.1074/jbc.M110.206813 . ПМК 3077604 . ПМИД 21349853 .
- ^ Jump up to: а б Варга А, Пальмаи З, Гуголя З, Грацер Э, Вондервизт Ф, Заводски П, Балог Е, Вас М (декабрь 2012 г.). «Важность остатков аспартата в балансировании гибкости и точной настройке катализа 3-фосфоглицераткиназы человека». Биохимия . 51 (51): 10197–207. дои : 10.1021/bi301194t . ПМИД 23231058 .
- ^ Клифф М.Дж., Боулер М.В., Варга А., Марстон Дж.П., Сабо Дж., Хаунслоу А.М., Бакстер Н.Дж., Блэкберн ГМ, Вас М., Валто Дж.П. (май 2010 г.). «Аналоговые структуры переходного состояния человеческой фосфоглицераткиназы устанавливают важность баланса зарядов в катализе». Журнал Американского химического общества . 132 (18): 6507–16. дои : 10.1021/ja100974t . ПМИД 20397725 .
- ^ Бэнкс, РД; Блейк, CCF; Эванс, PR; Хазер, Р.; Райс, Д.В.; Харди, GW; Мерретт, М.; Филлипс, AW (28 июня 1979 г.). «Последовательность, структура и активность фосфоглицераткиназы: возможный фермент, изгибающий шарнир». Природа . 279 (5716): 773–777. Бибкод : 1979Natur.279..773B . дои : 10.1038/279773a0 . ПМИД 450128 . S2CID 4321999 .
- ^ Бернштейн Б.Е., Хол В.Г. (март 1998 г.). «Кристаллические структуры субстратов и продуктов, связанных с активным центром фосфоглицераткиназы, раскрывают каталитический механизм». Биохимия . 37 (13): 4429–36. дои : 10.1021/bi9724117 . ПМИД 9521762 .
- ^ Ларссон-Разникевич М. (январь 1967 г.). «Кинетические исследования реакции, катализируемой фосфоглицераткиназой. II. Кинетические взаимоотношения между 3-фосфоглицератом, MgATP2- и активирующим ионом металла». Biochimica et Biophysical Acta (BBA) - Энзимология . 132 (1): 33–40. дои : 10.1016/0005-2744(67)90189-1 . ПМИД 6030358 .
- ^ Jump up to: а б Варга А., Чалоин Л., Саги Г., Сендула Р., Грацер Э., Лилиом К., Заводски П., Лионн К., Вас М (июнь 2011 г.). «Нуклеотидная распущенность 3-фосфоглицераткиназы находится в центре внимания: значение для разработки лучших аналогов против ВИЧ». Молекулярные биосистемы . 7 (6): 1863–73. дои : 10.1039/c1mb05051f . ПМИД 21505655 .
- ^ Ларссон-Разникевич, Марта; Викселл, Ева (1 марта 1978 г.). «Ингибирование фосфоглицераткиназы салицилатами». Biochimica et Biophysical Acta (BBA) - Энзимология . 523 (1): 94–100. дои : 10.1016/0005-2744(78)90012-8 . ПМИД 343818 .
- ^ Ёсида А, Тани К (1983). «Нарушения фосфоглицераткиназы: функциональные, структурные и геномные аспекты». Биомедика Биохимика Акта . 42 (11–12): С263-7. ПМИД 6689547 .
- ^ Jump up to: а б с Бейтлер Э. (январь 2007 г.). «Дефицит ПГК» . Британский журнал гематологии . 136 (1): 3–11. дои : 10.1111/j.1365-2141.2006.06351.x . ПМИД 17222195 . S2CID 21111736 .
- ^ Домашний справочник NIH по генетике
- ^ Роудс М., Эшфорд Л., Манес Б., Колдер С., Домм Дж., Франгул Х. (февраль 2011 г.). «Трансплантация костного мозга при дефиците фосфоглицераткиназы (PGK)» . Британский журнал гематологии . 152 (4): 500–2. дои : 10.1111/j.1365-2141.2010.08474.x . ПМИД 21223252 . S2CID 37605904 .
- ^ Цикер Д., Кенигсрайнер И., Тричлер И., Леффлер М., Беккерт С., Трауб Ф., Низельт К., Бюлер С., Веллер М., Гаедке Дж., Тайхман Р.С., Нортофф Х., Брюхер Б.Л., Кенигсрайнер А. (март 2010 г.). «Фосфоглицераткиназа 1 — фермент, способствующий перитонеальной диссеминации при раке желудка» . Международный журнал рака . 126 (6): 1513–20. дои : 10.1002/ijc.24835 . ПМК 2811232 . ПМИД 19688824 .
- ^ Галлуа-Монбрен С., Фарах А., Секламан Э., Соммадосси Ж.П., Девиль-Бонн Д., Верон М. (ноябрь 2004 г.). «Широкая специфичность человеческой фосфоглицераткиназы в отношении противовирусных аналогов нуклеозидов». Биохимическая фармакология . 68 (9): 1749–56. дои : 10.1016/j.bcp.2004.06.012 . ПМИД 15450940 .
Внешние ссылки
[ редактировать ]- Фосфоглицерат + киназа Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Иллюстрация на arizona.edu









